
Abstract
Species assemblages are shaped by local and continental-scale processes that are seldom investigated together, due to the lack of surveys along independent gradients of latitude and habitat types. Our study investigated changes in the effects of forest composition and structure on bat and bird diversity across Europe. We compared the taxonomic and functional diversity of bat and bird assemblages in 209 mature forest plots spread along gradients of forest composition and vertical structure, replicated in 6 regions spanning from the Mediterranean to the boreal biomes. Species richness and functional evenness of both bat and bird communities were affected by the interactions between latitude and forest composition and structure. Bat and bird species richness increased with broadleaved tree cover in temperate and especially in boreal regions but not in the Mediterranean where they increased with conifer abundance. Bat species richness was lower in forests with smaller trees and denser understorey only in northern regions. Bird species richness was not affected by forest structure. Bird functional evenness increased in younger and denser forests. Bat functional evenness was also influenced by interactions between latitude and understorey structure, increasing in temperate forests but decreasing in the Mediterranean. Covariation between bat and bird abundances also shifted across Europe, from negative in southern forests to positive in northern forests. Our results suggest that community assembly processes in bats and birds of European forests are predominantly driven by abundance and accessibility of feeding resources, i.e., insect prey, and their changes across both forest types and latitudes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The hierarchical framework of community assembly processes states that large-scale changes in species diversity are mainly driven by latitudinal gradients of climate stability and energy input (Ricklefs 2004; Clarke and Gaston 2006). At the local scale, fine-grained habitat composition and structure drive local species assemblages through resource availability and niche opportunities, but also interact with regional processes to shape patterns of diversity (Ferger et al. 2014; Kozlov et al. 2015a). At larger scales, global energy intake and temperature decrease towards higher latitudes (Pautasso and Gaston 2005). Accordingly, the general pattern of declining species richness with latitude was assumed to result from decreasing energetic input and increasing resource instability (Hurlbert 2004; Evans et al. 2005). Species assemblages are thus thought to be shaped by a combination of large-scale variations in environmental constraints and local changes in habitat or resource quality (Kissling et al. 2012; Ikin et al. 2014). Properly testing this hypothesis requires sampling species assemblages along a similar resource or habitat gradient, at multiple localities spread along wide latitudinal gradients (Willig and Bloch 2006).
In the present work, we sampled bat and bird diversity within a network of mature forest plots that are distributed along a tree species diversity gradient replicated on six countries spread along a continent-wide latitudinal gradient, from Mediterranean to boreal biomes (Baeten et al. 2013). European bats and birds are similar in diet (mostly insectivorous during the breeding season), have wide geographical distributions and use a variety of habitats that results in high habitat selectivity and low species turnover at the biogeographical scale (Dietz et al. 2009; Kissling et al. 2012; Stratford and Sekercioglu 2015). Bat and bird species living in mature forests depend on small-scale variation in canopy and understorey structure, e.g., vertical stratification, and composition, e.g., relative proportion of broadleaves and conifers (Lee and Rotenberry 2005; Jung et al. 2012; Ferger et al. 2014; Charbonnier et al. 2016), due to their high trophic position in the food web, well-defined foraging behaviours and relatively specialized habitat preferences (Müller et al. 2013; Zhang et al. 2013).
We specifically tested the hypothesis that forest structure and composition would have stronger effects on bat and bird species richness and functional diversity at higher latitudes. Studying the covariation between species assemblages from two functionally similar taxa is useful to detect local congruence in community structure (Eglington et al. 2015). Comparing changes in taxonomical and functional diversity of two taxa along the same environmental gradients at different latitudes may also help to understand the mechanisms underlying macro-scale patterns of variation (Mason et al. 2013). In addition, focusing on the functional structure of communities can provide insights into how environmental drivers shape species assemblage irrespective of changes in species richness (Mason et al. 2013; Mouillot et al. 2013; Cisneros et al. 2015; Lindenmayer et al. 2015). For example, differences in functional evenness, which measures the extent to which functional traits are equally distributed within species assemblages, would reflect the level of co-occurrence between functionally redundant species (Luck et al. 2013). Functional evenness is actually considered as a more adequate measure of functional diversity than other functional indices for reflecting ecosystem processes and efficiency in resource use (Petchey et al. 2007; Hillebrand et al. 2008; Barbaro et al. 2014). Last, studying the covariation between bat and bird functional diversity allows inference about intraguild competition between two potentially co-occurring insectivorous taxa (Speakman et al. 2000). From an applied point of view, this might help to understand the complementary role of bats and birds in providing pest control services to forests and surrounding landscapes (Maas et al. 2015; Whelan et al. 2015).
We tested four predictions: (1) bat and bird species richness in European forests decrease with increasing latitude, from Mediterranean to boreal forests (Hurlbert 2004); (2) bat and bird richness and functional evenness increase with the amount of broadleaved tree species because they are more likely to provide more nesting and feeding resources than evergreen trees (Russ and Montgomery 2002; Lindenmayer et al. 2015; Charbonnier et al. 2016); (3) forest understorey structure (e.g., vertical stratification) have opposite effects on the two predatory vertebrate taxa, being negative for bats which use acoustic signals to locate their prey, and positive for birds which are mostly foliage gleaners, thus ultimately resulting in reduced intraguild competition (Müller et al. 2013; Maas et al. 2015; Stratford and Sekercioglu 2015) and (4) the strength of forest habitat effects on bat and bird diversity increases with latitude as local environmental filters become more dominant (Ricklefs 2004).
Methods
Study area
The FunDivEurope Exploratory Platform was set up to investigate the effects of tree diversity on multiple forest ecosystem functions in a network of 209 forest plots in Europe (Baeten et al. 2013). The network was established in six forested regions ranging from 40°N to 63°N, separated by ca. 3200 km, and located in Mediterranean (Alto Tajo in Spain and Colline Metallifere in Italy), temperate (Hainich in Germany and Râsca in Romania), and boreal areas (North Karelia in Finland and Bialowieza in Poland) (Fig. 1). In each region, 28–43 forest plots were spread along a gradient of tree diversity ranging from monocultures of the locally most common tree species (target species) to mixtures of two, three, four or five of these target species, depending on the regional tree species pool. The gradient of sampled forest habitats was therefore designed to be strictly independent from the latitudinal gradient in terms of tree composition. Special attention was paid to establish all plots in mature forests of similar age, under homogeneous abiotic and management conditions (Baeten et al. 2013). A plot was delimited by a 90 m2 surrounded by a 20 m-large buffer area to avoid edge effects. In all plots, 14 variables of forest composition and structure were recorded: total tree species richness, conifer and broadleaved tree species richness and cover, total basal area, mean tree diameter at breast height and mean tree height, and shrub and soil cover. In addition, all vascular plant species <1.3 m in height were recorded in three 5 m × 5 m subplots, embedded into the main plot, to estimate understory species richness and cover (Ampoorter et al. 2014). We averaged the percentage cover of trees, shrubs, understory and bare soil for the three subplots to calculate (Eq. 1) a plot-level Shannon index of vertical stratification diversity (SVS) based on the percentage cover (pc i ) of each of the four vegetation strata (Ferger et al. 2014).

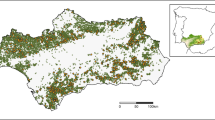
Map of the FunDivEUROPE exploratory network of 209 mature forest plots, with bar plots indicating mean (±SE) species richness for birds (green bars) and bats (blue bars). The number of sampled forest plots and tree species occurring per country are also indicated (colour figure online)
Bat and bird diversity
Bat and bird communities were sampled in all 209 plots. The plots in Italy, Germany and Finland were, respectively sampled in late April, mid-May and early June 2012, and those in Spain, Romania and Poland were sampled during the same periods in 2013, to account for latitudinal changes in bird and bat period of presence and peak in breeding activity. We visited all forest plots once to trade off sample size and spatial coverage against temporal replications, and to sample the entire gradient at every location within a short time period, i.e., within a single breeding season.
Bats were sampled during a complete night with an automatic bat recorder (Sound Meter SM2BAT, Wildlife acoustics) located at the centre of the plot (Müller et al. 2012). Detectors were calibrated to pick up all bat calls and were programmed to record from 1 h before sunset to 1 h after sunrise. Recordings were performed only during nights without rain, with low wind speeds (<30 km h−1) and with an ambient temperature above 10 °C. Bats were identified to the finest possible taxonomic level on the basis of their calls, using Sonochiro v3.2.3 and Batsound 4.1 softwares. The same trained operator (YC) validated the automatic identifications with identification keys and updated species lists for each region. Identification to species level was achieved for about 80 % of the calls, but two pairs of Myotis species (Myotis mystacinus and M. brandtii; M. myotis and M. blythii) could not be separated due to call overlaps and were therefore merged into species pairs. Bat species richness was defined as the number of bat species (or bat pairs of Myotis spp) recorded in a given forest plot. Total bat activity was the number of bat passes per night and was further used as a proxy for total bat abundance. Due to bad meteorological conditions and technical problems, we obtained useful bat data for 195 out of the 209 plots. Missing plots were in Finland (5) and Italy (9), ranging from one to four species mixtures, with broadleaved and conifers species (no bias).
Bird communities were sampled using point-counts within a distance of 80 m around the observer, which is accurate for single visits to a large number of sites (Bonthoux and Balent 2012; Klingbeil and Willig 2015). Four trained observers (YC, LB, JYB and JN) recorded all birds heard or seen within the plot, except flyovers, during a 15 min period. Bird counts were performed during the first 4 h after sunrise, on days without adverse weather conditions such as strong wind or heavy rain. Bird species richness was defined as the number of bird species recorded in a given plot. The total number of individual birds of all species recorded per forest plot was further used as a proxy for total bird abundance. As we controlled for age and structure of sampled plots, we assumed that bird species detectability was comparable between the forest types in the different exploratory regions (Baeten et al. 2013).
Functional evenness
We computed bat and bird functional evenness on the basis of the same set of ten ecological traits or attributes that are indicative for species response to environmental changes (Duchamp and Swihart 2008; Hanspach et al. 2012; Barbaro et al. 2014; Cisneros et al. 2015; Lindenmayer et al. 2015). They were: the conservation status, biogeographical range, trophic and foraging guilds, fecundity, nest or roost site location, body mass, home range size, migration status and breeding phenology (Online Resources 3 and 4). We did not use bat species wingspan because it was highly correlated to body mass (R 2 = 0.86). We computed functional evenness using the ‘FD’ R package. This index measures the homogeneity of dispersion, for a given species assemblage, within the multivariate trait space defined by a principal coordinate analysis of the Gower dissimilarity matrix of species traits (Laliberté and Legendre 2010; Mason et al. 2013).
Data analysis
We summarized the variables describing forest plot composition and structure with the first two axes of a principal component analysis (Fig. 2). The first axis (PCA1, accounting for 24.2 % of total variance) was associated with forest composition and ranged from plots dominated by conifers (negative values) to broadleaved trees (positive values), with mixed stands at intermediate values. The second axis (PCA2, 21.3 % of total variance) matched a gradient of forest vertical structure, ranging from plots with large, tall trees and clear understorey (negative values) to plots with smaller trees and denser, more stratified understorey (positive values). We checked the absence of correlation between latitude and the first two PCA axes, (respectively R 2 = 0.02 and R 2 = 0.13 for PCA1 and PCA2), which made it possible to use the three variables in the same model and test their interaction terms.

PCA ordination biplot of 14 forest composition and structure variables recorded in the 209 sampled plots. PCA1 is related to forest composition variables and PCA2 to forest vertical structure variables. Up-right eigenvalues histogram
We tested for latitudinal changes in the effects of forest habitat composition and structure on bat and bird functional evenness with linear mixed models using a Poisson distribution for bat and bird species richness, and a Gaussian distribution for functional evenness. We accounted for the hierarchical structure of the dataset by adding country (n = 6) as a random effect. We also controlled for differences in target tree species identity across regions with an additional random effect corresponding to target tree species combinations (Baeten et al. 2013). The covariate structure included additive effects of forest composition (PCA1), forest structure (PCA2), latitude and interactions between latitude and PCA1 and PCA2.
In addition, we estimated the direction and magnitude of covariation between total bat activity (as a proxy for abundance) and total bird abundance per country along the latitudinal gradient using generalized linear mixed models (with Poisson distribution). All models were checked for overdispersion and spatial autocorrelation, but no residual structure justified further model complexity. We performed all analyses in R software using the lme4 package (Bates et al. 2015).
Results
Because we used a single-visit sampling method to cover the entire study area (209 plots) within two summer seasons, we verified that we sampled a significant proportion of the entire European forest species pool for both bats and birds. We recorded a total of 26 bat species in 192 plots and 76 bird species in 209 plots, which represent 79–72 % of the European forest species, respectively (Huntley et al. 2007; Dietz et al. 2009, Online Resources 1 and 2).
Models for bat and bird species richness and functional evenness showed that all but one of the response variables were affected by interacting effects of latitude and forest composition and/or structure (Table 1). Bat species richness increased with the proportion of broadleaved trees (PCA1) in temperate forests and especially in boreal regions, while it tended to decrease in the Mediterranean (Table 1; Fig. 3a). Bat species richness decreased in forests with smaller trees and denser, more stratified understorey in northern regions but it was not significantly affected by forest structure in Mediterranean regions (Table 1; Fig. 3b). Bird species richness showed exactly the same response pattern to forest composition as for bats, with a significant interaction between latitude and proportion of broadleaved trees (Fig. 3c). By contrast, bird species richness was not affected by the vertical structure of forests (Table 1).

Forest composition (PCA1) and/or structure (PCA2) effects on bat species richness (a, b), bird species richness (c) and bat functional evenness (d) along the latitudinal gradient from boreal (blue, Finland and Poland); to temperate (green, Germany and Romania) and Mediterranean (red, Italy and Spain) regions. PCA1 accounts for forest composition, ranging from plots dominated by conifers (negative values) to broadleaved trees (positive values). PCA2 accounts for forest vertical structure ranging from plots with large, tall trees and clear understorey (negative values) to plots with smaller trees and denser, more stratified understorey (positive values) (colour figure online)
Bat functional evenness was not influenced by the composition of forest stands but by the interaction between forest structure and latitude (Table 1). It decreased in younger, denser Mediterranean forests while it increased in temperate forests (Table 1; Fig. 3d) and was not affected by forest structure in boreal regions. Bird functional evenness was not either affected by forest composition but significantly increased in younger, denser forests, irrespective of latitude (Table 1).
Finally, covariations between bird abundance and bat activity gradually changed along the latitudinal gradient (Fig. 4), with regression coefficients shifting from significantly negative in Mediterranean countries (Italy and Spain) to significantly positive in boreal countries (Poland and Finland).

Estimates (±confidence intervals) for generalized linear mixed models correlating total bat activity to total bird abundance per region, ranked by latitude
Discussion
How the influence of habitat on the composition of species assemblages changes along latitudinal gradients has seldom been explicitly addressed in continent-scale surveys. Here, based on an extensive sampling of bird and bat species assemblages, we demonstrated that the effects of forest habitat composition and structure significantly changed with latitude. The species-energy theory predicts a positive correlation between species richness and available energy (Pautasso and Gaston 2005), particularly for insectivores (Kissling et al. 2012). However, we did not find any consistent pattern of reduced bat and bird species richness towards higher latitudes, suggesting a stronger effect of local habitat features than large-scale climatic and energy gradients. Most forest bats and birds have widespread European distributions and their regional diversity is relatively low compared to other continents (e.g., Cisneros et al. 2015), making local changes in resource availability and habitat suitability more important drivers of local communities than the composition of regional species pools (Kissling et al. 2012; Ramos Pereira and Palmeirim 2013).
Both bat and bird richness increased with the proportion of broadleaved trees in temperate and boreal forests, and with the proportion of conifers in Mediterranean forests (Fig. 3a–c). Habitat effects on species richness usually reflect differences in life histories and resource use strategies (Zhang et al. 2013; Ikin et al. 2014). The congruence of bat and bird richness along gradients of forest composition therefore implies that common ecological traits are involved, suggesting that insectivorous diet and foraging methods are key to explain these patterns. The positive effect of broadleaved trees under temperate and boreal climates is likely due to higher insect prey availability (Brändle and Brandl 2001). In these regions, broadleaved trees generally increase the abundance and diversity of available prey for both bats (Razgour et al. 2011; Fuentes-Montemayor et al. 2013; Charbonnier et al. 2016) and insectivorous birds that mostly locate their prey visually by gleaning on broadleaved tree foliage (Mäntylä et al. 2004; Barbaro et al. 2014; Maas et al. 2015). The positive effect of conifers on bat and bird species richness in Mediterranean regions is consistent with past observations of bats and birds being as much or even more abundant in native pine than in evergreen Mediterranean forests (Napal et al. 2010; Proença et al. 2010; Rey-Benayas et al. 2010). Moreover, many Mediterranean broadleaved tree species are evergreen and tend to host less insect herbivores than temperate and boreal broadleaved trees (Giffard et al. 2012) that are less defended against herbivory (Björkman and Larsson 1991).
Bat species richness was higher in older stands with less dense understorey in temperate and boreal regions, likely because bats locate their prey using acoustic signals that are more difficult to perceive or interpret in forests with complex vertical structure (Müller et al. 2012; Jung et al. 2012). Bird species richness was not affected by forest structure, irrespective of latitudes, probably because foraging strategies are more diverse in this taxon, making bird communities more flexible in resource use (Moorman et al. 2007).
Bird functional evenness was higher in forests with more complex understorey structure (e.g., in younger and denser forests), indicating lower functional redundancy when species richness is low (Petchey et al. 2007; Luck et al. 2013; Mouillot et al. 2013). The same pattern was observed with bat functional evenness, but only in temperate countries (Fig. 3d). The fact that functional evenness increased with forest structure complexity may reflect the efficiency of resource partitioning by these predatory taxa (Hillebrand et al. 2008). More complex forest structure would result in more abundant and diverse prey resources, allowing higher levels of predator functional diversity (Barbaro et al. 2014; Ferger et al. 2014). The opposite (negative) response pattern of Mediterranean bat evenness to forest structure may be due to difference in bat species composition (Cruz et al. 2016). Bat species assemblages in these forests were dominated by Pipistrellus spp., which have been found to cope with structurally complex habitat by adjusting echolocation processes (Kalko and Schnitzler 1993; Berger-Tal et al. 2008). This confers a benefit when dense understorey vegetation provides complementary prey resources (Müller et al. 2012) in forest habitats relatively poor in insects (Cruz et al. 2016).
Forest bats and birds are two groups of insectivorous vertebrates that share broad habitat and dietary requirements. As a result, the hypothesis of competitive exclusion among bats and birds along the independent gradients of habitat and latitude is worth being tested (Speakman et al. 2000). Here, we found that bats and birds tend to aggregate in the same forest habitats in northern countries while they tend to be segregated by habitat structures in southern countries. The positive covariation in northern countries may be due to a summer ‘resource pulse’ of invertebrate biomass concentrating predation over short time intervals and restricted spatial extents (Speakman et al. 2000). The positive covariation in southern countries could be explained by higher and more even insect abundance (Kozlov et al. 2015b), and higher habitat-diversity in forested landscapes, allowing habitat partitioning among competing predators with important dispersal capacities (Tilman 1994).
In accordance with both theoretical and empirical studies (Hurlbert 2004; Ramos Pereira and Palmeirim 2013; Zhang et al. 2013), our results showed that local habitat features and large-scale processes jointly shape bat and bird communities across a large diversity of mature European forests. This is consistent with the prediction that community assembly processes are driven by an interaction between resource and energy gradients at the continental-scale and habitat filters at the local scale (Ricklefs 2004). We thus advocate for taking into account biogeographical factors when studying large-scale variations in habitat-diversity relationships, since they appear important drivers of interactions between regional species pools and both taxonomical and functional composition of local communities.
References
Ampoorter E, Baeten L, Koricheva J, Vanhellemont M, Verheyen K (2014) Do diverse overstoreys induce diverse understoreys? Lessons learnt from an experimental-observational platform in Finland. For Ecol Manag 318:206–215
Baeten L, Verheyen K, Wirth C, Bruelheide H, Bussotti F, Finér L, Jaroszewicz B, Selvi F, Valladares F, Allan E, Ampoorter E, Auge H, Avăcăriei D, Barbaro L, Bărnoaiea I, Bastias C, Bauhus J, Beinhoff C, Benavides R, Benneter A, Berger S, Berthold F, Boberg J, Bonal D et al (2013) A novel comparative research platform designed to determine the functional significance of tree species diversity in European forests. Perspect Plant Ecol Evol Syst 15:281–291
Barbaro L, Giffard B, Charbonnier Y, van Halder I, Brockerhoff EG (2014) Bird functional diversity enhances insectivory at forest edges: a transcontinental experiment. Divers Distrib 20:149–159
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. doi:10.18637/jss.v067.i01
Berger-Tal O, Berger-Tal R, Korine C, Holderied MW, Fenton MB (2008) Echolocation calls produced by Kuhl’s pipistrelles in different flight situations. J Zool 274:59–64
Björkman C, Larsson S (1991) Pine sawfly defence and variation in host plant resin acids: a trade-off with growth. Ecol Entomol 16:283–289
Bonthoux S, Balent G (2012) Point count duration: 5 minutes are sufficient to model the distribution of bird species and study the structure of communities in a French landscape. J Ornithol 153:491–504
Brändle M, Brandl R (2001) Species richness of insects and mites on trees: expanding Southwood. J Anim Ecol 70:491–504
Charbonnier Y, Gaüzère P, van Halder I, Nezan J, Barnagaud JY, Jactel H, Barbaro L (2016) Deciduous trees increase bat diversity at stand and landscape scales in mosaic pine plantations. Land Ecol 31:291–300
Cisneros LM, Fagan ME, Willig MR (2015) Effects of human-modified landscapes on taxonomic, functional, and phylogenetic dimensions of bat biodiversity. Divers Distrib 21:523–533
Clarke A, Gaston KJ (2006) Climate, energy and diversity. Proc R Soc B Biol Sci 273:2257–2266
Cruz J, Sarmento P, Rydevik G, Rebelo H, White PCL (2016) Bats like vintage: managing exotic eucalypt plantations for bat conservation in a Mediterranean landscape. Anim Conserv 19:53–64
Dietz C, von Helversen O, Nill D (2009) Bats of Britain. A & C Black, Europe and Northwest Africa
Duchamp JE, Swihart RK (2008) Shifts in bat community structure related to evolved traits and features of human-altered landscapes. Landsc Ecol 23:849–860
Eglington SM, Brereton TM, Tayleur CM, Noble D, Risely K, Roy DB, Pearce-Higgins JW (2015) Patterns and causes of covariation in bird and butterfly community structure. Landsc Ecol 30:1461–1472
Evans KL, Warren PH, Gaston KJ (2005) Species-energy relationships at the macroecological scale: a review of the mechanisms. Biol Rev 80:1–25
Ferger SW, Schleuning M, Hemp A, Howell KM, Böhning-Gaese K (2014) Food resources and vegetation structure mediate climatic effects on species richness of birds. Glob Ecol Biogeogr 23:541–549
Fuentes-Montemayor E, Goulson D, Cavin L, Wallace JM, Park KJ (2013) Fragmented woodlands in agricultural landscapes: the influence of woodland character and landscape context on bats and their insect prey. Agric Ecosyst Environ 172:6–15
Giffard B, Corcket E, Barbaro L, Jactel H (2012) Bird predation enhances tree seedling resistance to insect herbivores in contrasting forest habitats. Oecologia 168:415–424
Hanspach J, Fischer J, Ikin K, Stott J, Law BS (2012) Using trait-based filtering as a predictive framework for conservation: a case study of bats on farms in southeastern Australia. J Appl Ecol 49:842–850
Hillebrand H, Bennett DM, Cadotte MW (2008) Consequences of dominance: a review of evenness effects on local and regional ecosystem processes. Ecology 89:1510–1520
Huntley B, Green RE, Collingham YC, Willis SG (2007) A climatic atlas of European breeding birds. Lynx, Barcelona
Hurlbert AH (2004) Species-energy relationships and habitat complexity in bird communities. Ecol Lett 7:714–720
Ikin K, Barton PS, Stirnemann I, Stein JR, Michael D, Crane M, Okada S, Lindenmayer DB (2014) Multi-scale associations between vegetation cover and woodland bird communities across a large agricultural region. Plos One 9:e97029
Jung K, Kaiser S, Böhm S, Nieschulze J, Kalko EKV (2012) Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J Appl Ecol 49:523–531
Kalko EK, Schnitzler HU (1993) Plasticity in echolocation signals of European pipistrelle bats in search flight: implications for habitat use and prey detection. Behav Ecol Sociobiol 33:415–428
Kissling WD, Sekercioglu CH, Jetz W (2012) Bird dietary guild richness across latitudes, environments and biogeographic regions. Glob Ecol Biogeogr 21:328–340
Klingbeil BT, Willig MR (2015) Bird biodiversity assessments in temperate forest: the value of point count versus acoustic monitoring protocols. Peer J 3:e973
Kozlov MV, Stańska M, Hajdamowicz I, Zverev V, Zvereva EL (2015a) Factors shaping latitudinal patterns in communities of arboreal spiders in northern Europe. Ecography 38:1026–1035
Kozlov MV, Lanta V, Zverev V, Zvereva EL (2015b) Global patterns in background losses of woody plant foliage to insects. Global Ecol Biogeogr 24:1126–1135
Laliberté E, Legendre P (2010) A distance-based framework for measuring functional diversity from multiple traits. Ecology 91:299–305
Lee PY, Rotenberry JT (2005) Relationships between bird species and tree species assemblages in forested habitats of eastern North America. J Biogeogr 32:1139–1150
Lindenmayer D, Blanchard W, Tennant P, Barton P, Ikin K, Mortelliti A, Okada S, Crane M, Michael D (2015) Richness is not all: how changes in avian functional diversity reflect major landscape modification caused by pine plantations. Divers Distrib 21:836–847
Luck GW, Carter A, Smallbone L (2013) Changes in bird functional diversity across multiple land uses: interpretations of functional redundancy depend on functional group identity. Plos One 8:e63671
Maas B, Karp DS, Bumrungsri S, Darras K, Gonthier D, Huang JCC, Lindell CA, Maine JJ, Mestre L, Michel NL, Morrison EB, Perfecto I, Philpott SM, Sekercio CH, Silva RM, Taylor PJ, Tscharntke T, Van Bael SA, Whelan CJ, Williams-Guill K (2015) Bird and bat predation services in tropical forests and agroforestry landscapes. Biol Rev. doi:10.1111/brv.12211
Mäntylä E, Klemola T, Haukioja E (2004) Attraction of willow warblers to sawfly-damaged mountain birches: novel function of inducible plant defences? Ecol Lett 7:915–918
Mason NWH, de Bello F, Mouillot D, Pavoine S, Dray S (2013) A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J Veg Sci 24:794–806
Moorman CE, Bowen LT, Kilgo JC, Sorenson CE, Hanula JL, Horn S, Ulyshen MD (2007) Seasonal diets of insectivorous birds using canopy gaps in a bottomland forest. J Field Ornithol 78:11–20
Mouillot D, Graham NAJ, Villéger S, Mason NWH, Bellwood DR (2013) A functional approach reveals community responses to disturbances. Trends Ecol Evol 28:167–177
Müller J, Mehr M, Bässler C, Fenton M, Hothorn T, Pretzsch H, Klemmt HJ, Brandl R (2012) Aggregative response in bats: prey abundance versus habitat. Oecologia 169:673–684
Müller J, Brandl R, Buchner J, Pretzsch H, Seifert S, Stratz C, Veith M, Fenton B (2013) From ground to above canopy-bat activity in mature forests is driven by vegetation density and height. For Ecol Manag 306:179–184
Napal M, Garin I, Goiti U, Salsamendi E, Aihartza J (2010) Habitat selection by Myotis bechsteinii in the southwestern Iberian Peninsula. Ann Zool Fennici 47:239–250
Pautasso M, Gaston KJ (2005) Resources and global avian assemblage structure in forests. Ecol Lett 8:282–289
Petchey OL, Evans KL, Fishburn IS, Gaston KJ (2007) Low functional diversity and no redundancy in British avian assemblages. J Anim Ecol 76:977–985
Proença VM, Pereira HM, Guilherme J, Vicente L (2010) Plant and bird diversity in natural forests and in native and exotic plantations in NW Portugal. Acta Oecol 36:219–226
Ramos Pereira MJ, Palmeirim JM (2013) Latitudinal diversity gradients in New World bats: are they a consequence of niche conservatism? Plos One 8:e69245
Razgour O, Hanmer J, Jones G (2011) Using multi-scale modelling to predict habitat suitability for species of conservation concern: the grey long-eared bat as a case study. Biol Conserv 144:2922–2930
Rey-Benayas JM, Galván I, Carrascal LM (2010) Differential effects of vegetation restoration in Mediterranean abandoned cropland by secondary succession and pine plantations on bird assemblages. For Ecol Manag 260:87–95
Ricklefs RE (2004) A comprehensive framework for global patterns in biodiversity. Ecol Lett 7:1–15
Russ J, Montgomery W (2002) Habitat associations of bats in Northern Ireland: implications for conservation. Biol Conserv 108:49–58
Speakman JR, Rydell J, Webb PI, Hayes JP, Hays GC, Hulbert IAR, McDevitt RM (2000) Activity patterns of insectivorous bats and birds in northern Scandinavia (69°N), during continuous midsummer daylight. Oikos 88:75–86
Stratford J, Sekercioglu CH (2015) Birds in forest ecosystems. In: Peh KSH, Corlett R, Bergeron Y (eds) Handbook of forest ecology. Routledge Press, London, pp 281–296
Tilman D (1994) Competition and biodiversity in spatially structured habitats. Ecology 75:2–16
Whelan CJ, Sekercioglu CH, Wenny DG (2015) Why birds matter: from economic ornithology to ecosystem services. J Ornithol 156:227–238
Willig MR, Bloch CP (2006) Latitudinal gradients of species richness: a test of the geographic area hypothesis at two ecological scales. Oikos 112:163–173
Zhang J, Kissling WD, He FL (2013) Local forest structure, climate and human disturbance determine regional distribution of boreal bird species richness in Alberta, Canada. J Biogeogr 40:1131–1142
Acknowledgments
We are grateful to FunDivEurope site managers and field teams for assistance in fieldwork. We also thank Inge van Halder, Fabrice Vetillard and Bastien Castagneyrol for help and advice at various stages of this study. JYB received financial support from the CIRCE project under the AU Ideas program. The research reported here was conducted as part of the European FunDivEUROPE project, which received funding from the European Commission’s Seventh Framework Program under the Grant Agreement No. 265171.
Author contribution statement
YC, LB, KV and HJ designed the study, YC, LB, JYB, EA and JN performed the field work, YC, LB and JYB analysed the data, YC, LB, JYB and HJ wrote the manuscript and all authors contributed to editing the final version.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David N. Koons.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Charbonnier, Y.M., Barbaro, L., Barnagaud, JY. et al. Bat and bird diversity along independent gradients of latitude and tree composition in European forests. Oecologia 182, 529–537 (2016). https://doi.org/10.1007/s00442-016-3671-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-016-3671-9