
Abstract
Levan is a functional homopolymer of fructose with considerable applications in food, pharmaceutical and cosmetic industries. To improve the levan production in Bacillus amyloliquefaciens, the regulatory elements of sacB (encoding levansucrase) expression and levansucrase secretion were optimized. Four heterologous promoters were evaluated for sacB expression, and the Pgrac promoter led to the highest level for both sacB transcription and levansucrase enzyme activity. The levan production in the corresponding recombinant strain ΔLP-pHTPgrac reached 30.5 g/L, which was 114% higher than that of the control strain NK-ΔLP. In a further step, eight signal peptides were investigated (with Pgrac as the promoter for sacB expression) for their effects on the levansucrase secretion and levan production. The signal peptide yncM was identified as the optimal one, with a secretion efficiency of approximately 90%, and the levan production in the corresponding recombinant strain ΔLP-Y reached 37.4 g/L, which was 161% higher when compared with the control strains NK-ΔLP. Finally, fed-batch fermentation was carried out in 5-L bioreactors for levan production using the recombinant strain ΔLP-Y. A final levan concentration of 102 g/L was achieved, which is very close to the ever reported highest levan production level from the literature. To our best knowledge, this is the first report of the improvement of levan production through metabolic optimization for sacB expression and levansucrase secretion. The results from this study provided essential insights for systematically metabolic engineering of microbial cell factories for enhanced biochemical production.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Levan is a homopolymer of fructose polymerized by β-(2 → 6) glycosidic bonds in a main chain with occasional β-(2 → 1) linked branching points (Nakapong et al. 2013). Besides the common features of natural polysaccharide like safety, biocompatibility and biodegradability, levan also exhibits many favorable properties such as moisturizing, anti-obesity, anti-tumor, anti-oxidant, anti-inflammatory, hyperglycemic inhibitor and other biomedical functions (Dahech et al. 2011; Kucukasik et al. 2011; Dahech et al. 2012, 2013 ). Therefore, levan has been widely applied in food, prebiotics, cosmetics and medicines.
Levan biosynthesis usually involves the enzyme levansucrase (EC 2.4.1.10), which belongs to the glycoside hydrolases family 68 according to Carbohydrate-Active enzymes Database (Alamäe et al. 2012; Li et al. 2015). Levansucrase hydrolyzes sucrose by cleaving β-(1 → 2) linkage and transfers the fructosyl group to an acceptor molecule such as sucrose or fructooligosaccharide chain to generate levan. The molecular weight of the generated levan depends on specific features of the levansucrase (Li et al. 2015). Levansucrase is an bacterial extracellular enzyme, which can be produced by a wide range of bacterial strains, including Bacillus subtilis, Bacillus amyloliquefaciens, Zymomonas mobilis, Pseudomonas fluorescens and Lactobacillus reuteri (Abdel-Fattah et al. 2005; de Paula et al. 2008; Sims et al. 2011; Jathore et al. 2012; Feng et al. 2015a).
From the literature, there are several strategies that have been employed to increase the production of levan. First, intensive studies have been focused on selection of good natural producers, optimization of fermentation media and improvement of fermentation process design (Silbir et al. 2014; Wu et al. 2013). Second, sucrose metabolic pathways have been engineered to increase the synthesis of levan. The extracellular sucrase gene (sacC) disruption strain Z. mobilis showed 3-fold higher levansucrase activity than the wild-type strain, and the levan production was improved from 15.5 to 21.2 g/L (Senthilkumar et al. 2004). Similar genetic modification was carried out in L. reuteri, and results demonstrated that the deletion of the sucrose phosphorylase gene scrP also increased levan production (Teixeira et al. 2013). Recently, protease-deficient strains were constructed to improve the levan production. The B. amyloliquefaciens NK-Q-7 strain with six extracellular protease genes that were deleted showed the highest levan production of 31.1 g/L, representing a 102% increase compared to the control strain (Feng et al. 2015b).
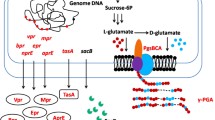
In this study, we attempted to achieve enhanced levan production from a different aspect, that is by optimizing the regulatory elements for levansucrase activity, including identifying the optimal strong promoter and signal peptide. The promoter of P43 is a well-characterized strong constitutive promoter in Bacillus and has been widely used for enzyme overexpression and metabolic engineering (Zhang et al. 2005, 2011). The inducible promoters Pgrac (IPTG inducible), PxylA (xylose inducible) and PsacB (sucrose inducible) have been successfully applied for the protein expression and claimed to bring about high-level expression of the target protein (Liu and Du 2012; Phan et al. 2012).
Therefore, these promoters were first evaluated for obtaining high levansucrase activity and enhanced levan production. On the other hand, the signal peptides located at the N-terminus of secretory protein play an important role in targeting the protein to the translocation machinery and transporting the protein from cytoplasm into the growth medium (Ng and Sarkar 2013). In the previous research, the secretion systems in Bacillus have been extensively investigated and many exceptional signal peptides have been identified and characterized (Zhang et al. 2013). Although the primary amino acid sequences of Bacillus signal peptides are not conserved, they share certain common features. Generally, a typical signal peptide contains three distinct regions, including a positively charged N-terminal domain, a hydrophobic core region (H region) and a C-terminal region with a signal peptidase cleavage site (Song et al. 2015). On the basis of the signal peptidase (SPase) recognition sequence, signal peptides are classified into four major families, and the Sec-dependent signal peptides have attracted the most attention due to its widespread and potential application for the secretion of enzyme production (Chen et al. 2007; Ying et al. 2012). However, the adaptation mechanism between the target protein and the signal peptide was still unclear, which might involve many aspects, such as molecular mass, folding rate and charge of the target protein (Zhang et al. 2013). Therefore, screening the optimal signal peptide was usually carried out by non-rational approaches, including single specific verification, random mutagenesis and high throughput screening (Brockmeier et al. 2006; Caspers et al. 2010). In this study, we attempt to improve levansucrase secretion through screening the optimal signal peptide.
Based on the levan producing B. amyloliquefaciens NK-ΔLP strain (Feng et al. 2015a), here, we further optimized the regulatory elements of levansucrase to achieve high-level levan production. Pgrac, PxylA, PA2up and PC2up were evaluated for their effects on the transcriptional level of sacB (encoding levansucrase). The strong promoter Pgrac was eventually identified for optimal levan production. Then, eight signal peptides were screened for the enhanced secretion of levansucrase. It was found that when fused with the yncM signal peptide, the levansucrase activity was 19.1-fold higher than the wild-type strain, and the levan production was improved from 14.2 (in control) to 37.4 g/L in flask fermentation. Our results from this study provided valuable reference for boosting bioproduct production in relevant hosts through rational metabolic engineering approaches.
Materials and methods
Strains and growth conditions
All the strains used in this work are listed in Table 1. Escherichia coli DH5α was used for plasmid propagation and transformation. Plasmid demethylation was carried out in E. coli GM2163. B. amyloliquefaciens NK-ΔLP that was derived from B. amyloliquefaciens LL3 (CCTCC M 208109) was used as the starting host strain (Feng et al. 2015a). E. coil and B. amyloliquefaciens cells were cultured at 37 °C in Luria-Bertani (LB) medium. Selection of E. coil was done with ampicillin (100 μg/mL). For B. amyloliquefaciens, chloramphenicol (6 μg/mL) was used.
The genome sequence accession number of B. amyloliquefaciens LL3 is CP002634. And the sacB gene ID is LL3_RS20055.
DNA manipulation and plasmid construction
All plasmids and primers used in this study are listed in Tables 1 and 2. The sacB gene was amplified from the genomic DNA of B. amyloliquefaciens NK-ΔLP, using primers sacB-F and sacB-R. The 1.4-kb fragment was digested with BamHI/XbaI and cloned into the E. coli-B. subtilis shuttle vector pHT01, which was digested with same enzymes, resulting in pHTPgrac (Fig. S1). This plasmid has an IPTG-inducible promoter Pgrac and served as a backbone into which four different promoters were inserted individually to drive the expression of levansucrase in B. amyloliquefaciens NK-ΔLP. Two constitutive promoters PA2cup and PC2up were amplified from the plasmids pUC-A2cup and pUC-C2up using primer pairs M13F/M13R, respectively (The sequence information of these two promoters is shown in Fig. S1). The xylose-inducible promoter PxylA was cloned from the plasmid pWH1520. The native promoter of sacB was amplified from the genomic DNA of B. amyloliquefaciens NK-ΔLP. After digested with KpnI and BamHI, these amplified fragments were cloned into the corresponding sites of pHTPgrac, resulting in pHTPA2cup, pHTPC2up, pHTPxylA and pHTPsacB, respectively (Fig. S2).
To investigate the influence of signal peptides on the expression of levansucrase in B. amyloliquefaciens NK-ΔLP, the native signal peptide SPsacB of levansucrase was replaced by seven different ones (SPamyE, SPbgls, SPbpr, SPnprB, SPnprE, SPwapA and SPyncM), respectively.
The seven signal peptides genes were amplified using B. subtilis 168 genome as the template, and the levansucrase gene lacking the signal peptide was cloned from B. amyloliquefaciens NK-ΔLP. The two DNA fragments (the signal peptides and the truncated levansucrase) were joined by overlap PCR. These seven fusion fragments were digested by BamHI and XbaI individually and ligated into the vector pHT01, which was digested with same enzymes, generating pHTPA (SPamyE), pHTPB (SPbgls), pHTPR (SPbpr), pHTPN (SPnprB), pHTPE (SPnprE), pHTPW (SPwapA) and pHTPY (SPyncM), respectively (Fig. S3).
Construction of the recombinant strains
All plasmid propagation was carried out in E. coli DH5α, and the constructed plasmids were verified by DNA sequencing. The plasmid DNA was transformed into the host strain B. amyloliquefaciens NK-ΔLP by electroporation (Feng et al. 2014), and the empty vector pHT01 was used as the control. The transformed strains were confirmed by PCR using primes pHT-F/R.
SDS-PAGE analysis
The transformant cell cultures were harvested through centrifugation at 8000 rpm for 20 min at 4 °C. The supernatant was precipitated with 10% trichloroacetic acid (TCA) at 4 °C for 20 min, centrifuged at 12,000 rpm (4 °C) for 10 min, and the pellets were washed twice with 80% acetone. The soluble enzymes were subjected to a 12% (w/v) SDS-PAGE. Protein bands were stained with Coomassie Brilliant Blue R-250 solution.
Real-time quantitative reverse transcription PCR (qRT-PCR) analysis
qRT-PCR was performed to further determine the transcriptional level of sacB with various promoters. Bacterial cultures were harvested after 24 h incubation. Total RNA were extracted using the commercial RNApure Bacteria kit (DNase I) (Cwbio, China), and the samples were treated with RNase-free DNase I (Qiagen), following the manufacturer’s protocol. Complementary DNA (cDNA) was obtained by reverse transcription using a HiFi-MMLV cDNA Kit (Cwbio, China). qRT-PCR was performed using the RealMasterMix (SYBR Green) Kit (Cwbio, China). The transcription levels of these genes were normalized against the levels of rspU (Reiter et al. 2011). The primers used in qRT-PCR are listed in Table 2. Samples from five independent experiments were analyzed for each gene. The results were standardized as fold changes compared with that from the wild-type strain.
Levansucrase activity assay
The supernatants were recovered by centrifugation at 8000 rpm for 20 min at 4 °C, and then dialysis through a membrane with a cutoff of 8000 Da were performed for 48 h at 4 °C. Afterwards, the extracellular levansucrase activity of the supernatant was determined. For the intracellular levansucrase activity measurement, the cell pellets were resuspended in a 20 mM potassium phosphate buffer (pH 6.0) containing 1% Triton X-100 and then disrupted by ultrasonication (600 W for 30 min with cycles of sonication of 3 s on and 3 s off). The unbroken cells and cellular debris were precipitated by centrifugation (12,000 rpm for 20 min) at 4 °C, and the intracellular levansucrase extract was recovered for activity determination.
The levansucrase activity was assayed by using sucrose as a substrate. The enzymatic reactions were initiated by adding 0.1 mL enzyme solution with 0.4 mL 20 mM potassium phosphate buffer (pH 6.0) and 0.5 mL of 20% sucrose (w/v) in 20 mM potassium phosphate buffer (pH 6.0) preheated at 37 °C, and immediately the mixture was incubated at 37 °C for 20 min. The reaction was stopped by boiling for 10 min. One unit of total levansucrase activity is defined as the amount of the biocatalyst to liberate 1 μM of glucose from sucrose per minute. The concentrations of glucose in the enzymatic reaction were measured using a biosensor (SBA-40D, Biology Institute of the Shandong Academy of Sciences, China). Secretion efficiency was defined as the ratio of extracellular amylase activity to total levansucrase activity (sum of extracellular and intracellular levansucrase activities).
Levan production in flask fermentation
Cell culturing of various B. amyloliquefaciens strains was first started in the 250 mL flask containing 50 mL LB medium and aerobically cultured at 37 °C for 18 h with shaking at 180 rpm. Then, this seed culture was used to be inoculated into 100 mL fermentation medium in shaking flasks at a 3% (v/v) inoculum ratio. The culture was further cultivated at 37 °C and 180 rpm of agitation for 48 h. After growing to an optical density (OD600) of 0.7–0.8, the culture of ΔLP-pHTPgrac and ΔLP-pHTPxylA was induced with 0.1 mM IPTG and 0.5% (w/v) xylose, respectively. All fermentations were repeated for three times.
Batch and fed-batch fermentations for levan production in a 5-L fermenter
For batch fermentation in bioreactor, the seed culture (at an inoculum ratio of 10%, v/v) was inoculated into a 5-L bioreactor (Bailun Biological Technology Co. Ltd., China), containing 2.7 L of fermentation medium (Feng et al. 2015a). The agitation speed and sterile air rate were maintained at 300 rpm and 6 vvm, respectively. The pH was maintained at 6.0 by adding 3 M NaOH. The fermentation proceeded at 37 °C for 48 h, and samples were withdrawn at a 6-h interval for further analysis.
Fed-batch fermentation was carried out in a similar manner except that sucrose substrate was brought back up to 200 g/L at 24 and 36 h, respectively.
Analytical procedures
Cell growth was monitored by measuring the optical density at 600 nm (OD600) using a Shimadzu UV-1800 spectrophotometer (Kyoto, Japan). After fermentation, the culture was centrifuged (8000 rpm for 20 min) and the pellets were washed twice with distilled water and dried to a constant weight at 80 °C to determine the biomass of the culture (dry cell weight, DCW). The levan was recovered and purified following the method as described previously (Feng et al. 2015a). The culture was centrifuged at 4 °C, 8000 rpm for 20 min. Fourfold volume of absolute ethanol was added to the supernatant followed by incubation at 4 °C for overnight. The precipitate was resuspended in deionized water; then the solution was dialyzed through a membrane with a cutoff of 3500 Da for 48 h and afterwards lyophilized to obtain levan. The data reported are the average values ± standard deviation (SD) of three replicate experiments. The concentration of sucrose in the culture was measured by a high-performance liquid chromatography (HPLC) equipped with a differential refraction detector (Schambeck SFD) and a Prevail Carbohydrate ES 5u column (4.6 × 250 mm) set at a temperature of 30 °C. Seventy-five percent acetonitrile was used as the mobile phase at a flow rate of 1.0 mL/min. The concentration of the sucrose was estimated by constructing standard curve for sucrose. All assays were run in triplicate.
Results
Cloning of sacB gene from B. amyloliquefaciens NK-ΔLP
The sacB gene encoding levansucrase was amplified from NK-ΔLP genome and cloned into pUC19-simple for DNA sequencing. Sequence analysis showed that the sacB contains one 1419-bp open reading frame (ORF) and encodes 472 amino acids. Based on the prediction using the SignalP software (Brockmeier et al. 2006), it was revealed that the levansucrase consists of a Sec-type signal peptide of 29 aa and a 443 aa mature protein. BLAST search showed that the deduced amino acid sequence of the levansucrase from NK-ΔLP shares the highest similarity of 97% with the levansucrase of B. amyloliquefaciens (accession nos. AGJ52124.1 and KMO06910.1), and shares about 75–90% similarity with the levansucrase from other Bacillus strains (accession nos. CAA26513.1, CBI68350.1, AGZ16261.1 and ACI15886.1). However, it showed very low similarity with the levansucrase from Z. mobilis, Pseudomonas and Leuconostoc mesenteroides. Figure 1 presents the alignment of the NK-ΔLP levansucrase amino acid sequence with those of other Bacillus strains using the Clustal X software.

The alignment of the amino acid sequences of levansucrase from NK-ΔLP with six previously reported levansucrase from other Bacillus strains
Effect of different promoters on sacB transcription and levansucrase expression
The recombinant plasmids pHTPA2cup, pHTPC2up, pHTPxylA, pHTPgrac and pHTPsacB (containing the selected four promoters PA2cup, PC2up, PxylA and Pgrac and the native promoter of levansucrase PsacB, respectively) were transformed into NK-ΔLP. The desirable transformants were screened and identified with PCR. After growing to an optical density (OD600) of 0.7–0.8, the culture of ΔLP-pHTPgrac and ΔLP-pHTPxylA was induced with 0.1 mM IPTG and 0.5% (w/v) xylose, respectively. The transcription level of sacB in all these strains was determined with qRT-PCR after 24 h of incubation in a levan fermentation medium.
As shown in Fig. 2a, the transcription level of sacB in ΔLP-pHTPgrac was significantly higher than that in other strains, which was about 22.1- and 7.21-fold of the control strain NK-ΔLP and the strain ΔLP-pHTPsacB, respectively. The transcription level of sacB in ΔLP-pHTPC2up strain was about 6.34-fold of that in the control strain. The sacB transcription levels in other three recombinants were comparable. The results demonstrated that the promoter Pgrac is much more efficient than promoters PA2cup, PC2up, PxylA and PsacB in driving transcription of sacB in B. amyloliquefaciens NK-ΔLP.

The effects of different promoters on sacB transcription and expression. a qRT-PCR analysis of sacB gene transcription. b Extracellular levansucrase activity measurement. c SDS-PAGE analysis of levansucrase production in culture supernatants. Samples 1–7 correspond to NK-ΔLP, ΔLP-pHT01, ΔLP-pHTPA2cup, ΔLP-pHTPC2up, ΔLP-pHTPxylA, ΔLP-pHTPsacB and ΔLP-pHTPgrac, respectively. Lane M—molecular weight marker. Data are means ± SD from three replicates
To verify the transcription efficiency of sacB using the selected promoters, the extracellular levansucrase activity of the recombination strains was measured after 48 h of incubation in the levan fermentation medium (Fig. 2b). Obviously, the recombinant strain ΔLP-pHTPgrac generated the highest extracellular levansucrase activity, which was about 5–8-fold higher than that of the other strains. In addition, SDS-PAGE analysis was carried out to confirm the levansucrase expression. As shown in Fig. 2c, distinct bands with a molecular mass of about 52 kDa were observed, which is in good agreement with the predicted value. The recombinant strain ΔLP-pHTPgrac generated a much thicker band than others. This was consistent with the result of enzyme activity measurement. Therefore, Pgrac was identified as the optimal promoter to drive sacB expression in this study.
Effect of different promoters on levan production
The five recombinant strains as well as the control strain were cultured in flask to investigate the effect of different promoters on levan production and DCW. As shown in Fig. 3, the levan production of ΔLP-pHTPgrac reached 30.5 g/L, and it was 114% higher than that of the control strain ΔLP-pHT01 (14.2 g/L). The recombinant strains ΔLP-pHTPsacB, ΔLP-pHTPxylA and ΔLP-pHTPC2up produced 18.6, 18.6 and 18.8 g/L levan, respectively, which was about 30.3% higher than that of the control strain. The levan production of the ΔLP-pHTPA2cup was comparable with the control strain. On the other hand, interestingly, the DCW of the five recombinant strains were approximately the same as that of the control strain ΔLP-pHT01 and that of the wild-type NK-ΔLP. It was thus concluded here that the different enzyme activities and levan production were resulted from promoter strength not from cell growth.

Levan fermentation results with recombinant strains containing various promoters. Values represent the means ± SD
Effect of different signal peptides on the secretion of levansucrase
An appropriate signal peptide is a critical factor for the extracellular protein expression. So far, although numerous signal peptides have been investigated and characterized, there has been no specific rational approach or algorithm to identify the optimal signal peptide for a target protein. To find the best signal peptide for levansucrase, 14 candidate signal peptides from B. subtilis 168 were compared based on their compositions, especially the positively charged N-domain, the hydrophobic H-domain and the C-domain (for recognition and cleavage) (Table 3). Based on this preliminary analysis, seven different signal peptides amyE, bglS, bpr, nprB, nprE, wapA and yncM were chosen for further evaluation to improve the levansucrase secretion. We used Pgrac to drive the expression of levansucrase fused to different signal peptides. The seven plasmids pHTPA (SPamyE), pHTPB (SPbglS), pHTPR (SPbpr), pHTPN (SPnprB), pHTPE (SPnprE), pHTPW (SPwapA) and pHTPY (SPyncM) were constructed and transformed into NK-ΔLP, generating recombinant strains ΔLP-A, ΔLP-B, ΔLP-R, ΔLP-N, ΔLP-E, ΔLP-W and ΔLP-Y, respectively.
The extracellular levansucrase activity was analyzed for each strain after 48 h incubation in the levan fermentation medium. As shown in Fig. 4a, the strain ΔLP-Y produced the highest extracellular levansucrase titer, which was about 19.1-fold of the control strain NK-ΔLP. Moreover, the extracellular levansucrase activity of the recombinant strain ΔLP-B was secondly higher among all, which was 11.2-fold higher than the control strain. In addition, SDS-PAGE assay was carried out to further confirm the levansucrase secretion. As shown in Fig. 4b, the strains ΔLP-Y and ΔLP-B showed apparently deeper bands than others, which was consistent with the results of enzyme activity assay. Then, it was concluded that the signal peptide SPyncM was an excellent candidate for levansucrase secretory expression.

Comparison of levansucrase production in recombinant strains containing different signal peptides. a Extracellular enzyme activity. Values represent the means ± SD. b SDS-PAGE analysis of levansucrase secretion. Samples 1–9 corresponds to ΔLP-pHT01, ΔLP-A, ΔLP-B, ΔLP-R, ΔLP-N, ΔLP-E, ΔLP-W, ΔLP-Y and ΔLP-pHTPgrac, respectively. Lane M—molecular weight marker
Effect of different signal peptides on levan production
As shown above, different signal peptides led to various levels of levansucrase secretion. We thus further investigated their effects on levan production. The recombinant strains as well as the control strain were cultured in flask. As shown in Fig. 5, all the recombinant strains exhibited higher levan production than the control strain ΔLP-pHT01, and ΔLP-Y generated the highest levan titer of 37.4 g/L, which was about 161% improvement compared with ΔLP-pHT01. The levan production of ΔLP-B strain reached 32.7 g/L, which was about 130% higher than that of ΔLP-pHT01. This result was well consistent with the result of enzymatic assay.

Levan fermentation results with recombinant strains containing different signal peptides. Values represent the means ± SD
Scale-up of levan production in a 5-L fermenter
To obtain more practical parameters for industrial levan production, the scale-up fermentation was conducted for the recombinant ΔLP-Y strain in a 5-L fermenter. As shown in Fig. S4, it took about 18 h for the strain to reach the logarithmic growth phase, and the highest levan production of 51.2 g/L was achieved at 30 h, which was 3.58-fold higher than that was obtained from NK-ΔLP strain in flask. Based on previous reports, levansucrase has dual roles of hydrolysis and transfructosylation activities. The ratio of hydrolysis and transfructosylation activities is highly dependent on the sucrose concentration. High concentration of sucrose can promote transfructosylation activity, but when sucrose is close to depletion, hydrolysis tends to become predominant (Öner et al. 2016). Therefore, the depletion of sucrose at 30 h in our fermentation may hinder the further levan synthesis. We thus further carried out the fermentation in a fed-batch mode by supplementing sucrose to the concentration of 200 g/L at 24 and 36 h, respectively. As shown in Fig. 6, the highest levan production of 102 g/L was obtained at 48 h, which was 7.12-fold higher than that obtained from NK-ΔLP strain in flask. So far, the highest levan production reported is 112 g/L by a B. subtilis (natto) CCT7712 strain. Our result is very close to this value, indicating that the strain that we generated in this study has a great potential for industrial scale levan production (Öner et al. 2016).

Fermentation profiles from a fed-batch cultivation of the recombinant strain ΔLP-Y in a 5-L fermenter. Cell growth (filled diamond); levan production (filled triangles); dissolved oxygen (DO, filled cycle); residual sucrose (filled square)
Discussion
In our previous study, we generated the NK-ΔLP strain, which can produce levan at the molecular weight around 5 kDa that is significantly lower than other reported levan products (Feng et al. 2015a). The levan of low molecular weight from NK-ΔLP can potentially be used as a prebiotic compound (Feng et al. 2015a). However, the levan production was only about 13.9 g/L (flask fermentation), which is far from the desirable level for an economic industrial scale production. Therefore, in this study, we attempted to enhance the levan production in this strain through rational metabolic engineering approaches.
Since levan is the catalytic product of the secreted levansucrase, we hypothesized that we can improve levan production by increasing the expression level of levansucrase in the host strain. Therefore, the transcriptional and translational regulations of levansucrase were primarily targeted. Up to now, the most economic and effective approach for regulation modification still remains the optimization of promoters at transcriptional level (Song et al. 2015). The activity of the promoter directly influences the production efficiency of the target protein. In our previous work, when a strong promoter P43 from B. subtilis 168 replaced the sacB native promoter, the transcription level of sacB was not increased but was reduced by ten times, and the levan production has remarkably declined (Fig. S5). Through the promoter engineering, we identified a few promoters better than P43, including PA2cup, PC2up and PxylA (unpublished data), and thus we further evaluated them for levansucrase expression in this study. The promoters PA2cup and PC2up are both strong and constitutively expressed promoters from B. subtilis phage Ф29 that is recognized by sigma A factor (Meijer and Salas 2004). Pgrac promoter, a widely used IPTG-inducible promoter, is a combination of the B. subtilis groESL promoter, the E. coli lac operator and the gsiB Shine-Dalgarno sequence (Phan et al. 2012). The promoter PxylA, originating from Bacillus megaterium, is a strong xylose-inducible promoter (Radha and Gunasekaran 2008). As a control, the native promoter PsacB of levansucrase from B. amyloliquefaciens NK-ΔLP was also tested. The result suggested that Pgrac promoter is the most effective promoter in driving the transcription of sacB in B. amyloliquefaciens NK-ΔLP. To our surprise, the strong xylose-inducible promoter PxylA produced only modest amount of levansucrase. Although the transcriptional level of the promoter PC2up was doubled than that of PsacB promoter, the levansucrase activity and levan production of strain ΔLP-pHTPC2up were very similar to that of the strain ΔLP-pHTPsacB. The results indicate that not all strong promoters are suitable for levansucrase expression in NK-ΔLP. The expression of the target protein may be influenced by other factors, such as characteristics of the protein, promoter type, promoter-gene interactions and type of inducers (Ying et al. 2012).
For many years, B. subtilis and related Bacilli are known to naturally produce various proteins and secreted into the medium with high concentrations. In addition, Bacillus strains are generally classified as generally recognized as safe (GRAS) organisms (Pohl and Harwood 2010). Thus, people paid tremendous attention to them aiming to exploit them as major “cell factories” for secreted heterologous protein of interest (van Dijl and Hecker 2013). Although many studies have focused on the effect of different signal peptides on heterologous proteins expression (Wei et al. 2015; Ying et al. 2012; Zhang et al. 2013), few efforts have been made to study their effects on endogenous proteins production. On the other hand, it was reported that the signal peptide structure itself affects the secretion efficiency. Zanen et al. (2005) reported that the hydrophobicity of the signal peptide was critical for the early stage recognition of the signal peptide by signal recognition particle (SRP) in B. subtilis. By modifying the signal peptide with higher positive charge in the N-terminal region and higher hydrophobicity in the core H region, the yield of tuna growth hormone (tGH) was increased by 10-fold (Sagiya et al. 1994). This means that the appropriate signal peptide structure can improve the efficiency of protein secretion. To screen a compatible signal peptide for the levansucrase secretion, seven different signal peptides were selected and evaluated from B. subtilis 168. As described previously, the signal peptide of SPyncM can greatly improve the levansucrase secretion and levan production, which has never been reported in levan producing strain before.
Moreover, the intracellular levansucrase activity of the mentioned recombinant strains was characterized in the levan fermentation medium (Fig. S6a), and it was showed that the strain ΔLP-Y possessed the highest secretion efficiency of 89.9% (Fig. S6b). In addition, the different signal peptides have resulted in different levansucrase production rates (both extracellularly and intracellularly), indicating that the secretory expression of the protein has a close relationship with transportation rate directed by signal peptide. In fact, many previous studies have proposed that the balance between the amount of translocated proteins emerging from the trans-side of the membrane and their extracytosolic folding capacity played an important role on the secretory expression of the target protein (Beena et al. 2004; Brockmeier et al. 2006; Zhang et al. 2013). This could also be the reason for us to achieve the highest levansucrase secretion with appropriate combination of promoter and signal peptide.
In summary, in order to improve the levan production in strain NK-ΔLP, the regulatory elements for levansucrase expression were investigated and optimized. Five promoters were tested, and the strong promoter Pgrac was eventually identified as the optimal. Then eight signal peptides were screened and evaluated for their effects on the levansucrase secretory expression and the levan production. The signal peptide yncM showed the highest secretion efficiency and resulted in the high production of levansucrase and levan in flask fermentation. Moreover, fed-batch fermentation was carried out with the optimized recombinant strain ΔLP-Y, and a very high production of levan 102 g/L was achieved after 48 h of fermentation; this level is comparable to the previous reported highest levan production. This is the first report about improving levan production by optimizing of the regulatory elements for levansucrase. The levan production has been significantly increased. We are trying additional efforts to further improve the levan production in order to meet the industrial scale levan production purpose.
References
Abdel-Fattah AF, Mahmoud DA, Esawy MA (2005) Production of levansucrase from Bacillus subtilis NRC 33a and enzymic synthesis of levan and fructo-oligosaccharides. Curr Microbiol 51(6):402–407
Alamäe T, Visnapuu T, Mardo K, Mäe A, Zamfir AD (2012) Levansucrases of Pseudomonas bacteria: novel approaches for protein expression, assay of enzymes, fructooligosaccharides and heterooligofructans. Carbohydr Polym 38:176–191
Beena K, Udgaonkar JB, Varadarajan R (2004) Effect of signal peptide on the stability and folding kinetics of maltose binding protein. Biochemistry 43(12):3608–3619
Brockmeier U, Caspers M, Freudl R, Jockwer A, Noll T, Eggert T (2006) Systematic screening of all signal peptides from Bacillus subtilis: a powerful strategy in optimizing heterologous protein secretion in Gram-positive bacteria. J Mol Biol 362(3):393–402
Caspers M, Brockmeier U, Degering C, Eggert T, Freudl R (2010) Improvement of Sec-dependent secretion of a heterologous model protein in Bacillus subtilis by saturation mutagenesis of the N-domain of the AmyE signal peptide. Appl Microbiol Biotechnol 86(6):1877–1885
Chen Z, Heng C, Li Z, Liang X, Xinchen S (2007) Expression and secretion of a single-chain sweet protein monellin in Bacillus subtilis by sacB promoter and signal peptide. Appl Microbiol Biotechnol 73(6):1377–1381
Dahech I, Belghith KS, Hamden K, Feki A, Belghith H, Mejdoub H (2011) Oral administration of levan polysaccharide reduces the alloxan-induced oxidative stress in rats. Int J Biol Macromol 49(5):942–947
Dahech I, Belghith KS, Belghith H, Mejdoub H (2012) Partial purification of a Bacillus licheniformis levansucrase producing levan with antitumor activity. Int J Biol Macromol 51(3):329–335
Dahech I, Harrabi B, Hamden K, Feki A, Mejdoub H, Belghith H, Belghith KS (2013) Antioxidant effect of nondigestible levan and its impact on cardiovascular disease and atherosclerosis. Int J Biol Macromol 58:281–286
de Paula VC, Pinheiro IO, Lopes CE, Calazans GC (2008) Microwave-assisted hydrolysis of Zymomonas mobilis levan envisaging oligofructan production. Bioresour Technol 99(7):2466–2470
Feng J, Gu Y, Wang J, Song C, Yang C, Xie H, Zhang W, Wang S (2013) Curing the plasmid pMC1 from the poly (gamma-glutamic acid) producing Bacillus amyloliquefaciens LL3 strain using plasmid incompatibility. Appl Biochem Biotechnol 171(2):532–542
Feng J, Gao W, Gu Y, Zhang W, Cao M, Song C, Zhang P, Sun M, Yang C, Wang S (2014) Functions of poly-gamma-glutamic acid (γ-PGA) degradation genes in γ-PGA synthesis and cell morphology maintenance. Appl Microbiol Biotechnol 98(14):6397–6407
Feng J, Gu Y, Han L, Bi K, Quan Y, Yang C, Zhang W, Cao M, Wang S, Gao W, Sun Y, Song C (2015a) Construction of a Bacillus amyloliquefaciens strain for high purity levan production. FEMS Microbiol Lett 362(11). doi:10.1093/femsle/fnv079
Feng J, Gu Y, Quan Y, Zhang W, Cao M, Gao W, Song C, Yang C, Wang S (2015b) Recruiting a new strategy to improve levan production in Bacillus amyloliquefaciens. Sci Rep 5:13814. doi:10.1038/srep13814
Jathore NR, Bule MV, Tilay AV, Annapure US (2012) Microbial levan from Pseudomonas fluorescens: characterization and medium optimization for enhanced production. Food Sci Biotechnol 21(4):1045–1053
Kucukasik F, Kazak H, Guney D, Finore I, Poli A, Yenigun O, Nicolaus B, Oner ET (2011) Molasses as fermentation substrate for levan production by Halomonas sp. Appl Microbiol Biotechnol 89(6):1729–1740
Li W, Yu S, Zhang T, Jiang B, Mu W (2015) Recent novel applications of levansucrases. Appl Microbiol Biotechnol 99(17):6959–6969
Liu SL, Du K (2012) Enhanced expression of an endoglucanase in Bacillus subtilis by using the sucrose-inducible sacB promoter and improved properties of the recombinant enzyme. Protein Expr Purif 83(2):164–168
Meijer W, Salas W (2004) Relevance of UP elements for three strong Bacillus subtilis phage phi29 promoter. Nucleic Acids Res 32(3):1166–1176
Nakapong S, Pichyangkura R, Ito K, Iizuka M, Pongsawasdi P (2013) High expression level of levansucrase from Bacillus licheniformis RN-01 and synthesis of levan nanoparticles. Int J Biol Macromol 54:30–36
Ng DT, Sarkar CA (2013) Engineering signal peptides for enhanced protein secretion from Lactococcus lactis. Appl Environ Microbiol 79(1):347–356
Öner ET, Hernández L, Combie J (2016) Review of Levan polysaccharide: from a century of past experiences to future prospects. Biotechnol Adv. doi:10.1016/j.biotechadv.2016.05.002
Phan TT, Nguyen HD, Schumann W (2012) Development of a strong intracellular expression system for Bacillus subtilis by optimizing promoter elements. J Biotechnol 157(1):167–172
Pohl S, Harwood CR (2010) Heterologous protein secretion by Bacillus species: from the cradle to the grave. Adv Appl Microbiol 73:1–25
Radha S, Gunasekaran P (2008) Sustained expression of keratinase gene under PxylA and PamyL promoters in the recombinant Bacillus megaterium MS941. Bioresour Technol 99(13):5528–5537
Reiter L, Kolsto AB, Piehler AP (2011) Reference genes for quantitative, reverse-transcription PCR in Bacillus cereus group strains throughout the bacterial life cycle. J Microbiol Methods 86(2):210–217
Sagiya Y, Yamagata H, Udaka S (1994) Direct high-level secretion into the culture medium of tuna growth hormone in biologically active form by Bacillus brevis. Appl Microbiol Biotechnol 42(2–3):358–363
Senthilkumar V, Rameshkumar N, Busby SJW, Gunasekaran P (2004) Disruption of the Zymomonas mobilis extracellular sucrase gene (sacC) improves levan production. J Appl Microbiol 96(4):671–676
Silbir S, Dagbagli S, Yegin S, Baysal T, Goksungur Y (2014) Levan production by Zymomonas mobilis in batch and continuous fermentation systems. Carbohydr Polym 99:454–461
Sims IM, Frese SA, Walter J, Loach D, Wilson M, Appleyard K, Eason J, Livingston M, Baird M, Cook G, Tannock GW (2011) Structure and functions of exopolysaccharide produced by gut commensal Lactobacillus reuteri 100-23. ISME J 5(7):1115–1124
Song Y, Nikoloff JM, Zhang D (2015) Improving protein production on the level of regulation of both expression and secretion pathways in Bacillus subtilis. J Microbiol Biotechnol 25(7):963–977
Teixeira JS, Abdi R, Su MS-W, Schwab C, Gänzle MG (2013) Functional characterization of sucrose phosphorylase and scrR, a regulator of sucrose metabolism in Lactobacillus reuteri. Food Microbiol 36(2):432–439
van Dijl JM, Hecker M (2013) Bacillus subtilis: from soil bacterium to supersecreting cell factory. Microb Cell Factories 12:3. doi:10.1186/1475-2859
Wei X, Zhou Y, Chen J, Cai D, Wang D, Qi G, Chen S (2015) Efficient expression of nattokinase in Bacillus licheniformis: host strain construction and signal peptide optimization. J Ind Microbiol Biotechnol 42(2):287–295
Wu FC, Chou SZ, Shih IL (2013) Factors affecting the production and molecular weight of levan of Bacillus subtilis natto in batch and fed-batch culture in fermenter. J Taiwan Inst Chem E 44(6):846–853
Ying Q, Zhang C, Guo F, Wang S, Bie X, Lu F, Lu Z (2012) Secreted expression of a hyperthermophilic alpha-amylase gene from Thermococcus sp. HJ21 in Bacillus subtilis. J Mol Microbiol Biotechnol 22(6):392–398
Zanen G, Houben EN, Meima R, Tjalsma H, Jongbloed JD, Westers H, Oudega B, Luirink J, van Dijl JM, Quax WJ (2005) Signal peptide hydrophobicity is critical for early stages in protein export by Bacillus subtilis. FEBS J 272(18):4617–4630
Zhang XZ, Cui ZL, Hong Q, Li SP (2005) High-level expression and secretion of methyl parathion hydrolase in Bacillus subtilis WB800. Appl Environ Microbiol 71(7):4101–4103
Zhang XZ, Sathitsuksanoh N, Zhu Z, Percival Zhang YH (2011) One-step production of lactate from cellulose as the sole carbon source without any other organic nutrient by recombinant cellulolytic Bacillus subtilis. Metab Eng 13(4):364–372
Zhang J, Kang Z, Ling Z, Cao W, Liu L, Wang M, Du G, Chen J (2013) High-level extracellular production of alkaline polygalacturonate lyase in Bacillus subtilis with optimized regulatory elements. Bioresour Technol 146:543–548
Acknowledgments
This work was supported by the National Key Technology Support Program 2015BAD16B04; the Natural Science Foundation of China Grant Nos. 81671842, 31470213 and 31670093; and the Project of Tianjin, China, i.e., 16JCZDJC37600, 15ZCZDNC00450 and 14ZCZDSF00009; and the program of State Key Laboratory of Medicinal Chemical Biology 201603007.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Electronic supplementary material
ESM 1
(PDF 424 kb)
Rights and permissions
About this article

Cite this article
Gu, Y., Zheng, J., Feng, J. et al. Improvement of levan production in Bacillus amyloliquefaciens through metabolic optimization of regulatory elements. Appl Microbiol Biotechnol 101, 4163–4174 (2017). https://doi.org/10.1007/s00253-017-8171-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-017-8171-2