
Abstract
Rationale
Mesolimbic dopamine (DA) signaling is essential for the high maternal caregiving characteristic of the early postpartum period, but little is known about dopamine’s role in the expression of maternal caregiving thereafter.
Objectives
We tested the hypothesis that decreased mesolimbic dopaminergic signaling is particularly responsible for the natural decline in maternal caregiving that occurs as the postpartum period progresses.
Methods
Sprague-Dawley (SD) mother rats received intraperitoneal injections of either vehicle, the DA D1 receptor agonist SKF38393, the DA D2 receptor agonist quinpirole, or both agonists twice daily from postpartum days 9 to 15. In a separate experiment involving Long-Evans (LE) rats, we examined whether DA D1 and D2 receptor mRNAs in the nucleus accumbens (NA) shell and ventral tegmental area (VTA), along with DA turnover in the VTA, decline across the postpartum period in parallel with the decreasing maternal behavior.
Results
All drug treatments significantly maintained higher frequencies of active maternal behaviors (nesting, pup licking, retrieval) compared to vehicle. Furthermore, the majority of mothers treated with SKF38393 either alone or combined with quinpirole maintained full expression of maternal behavior during behavioral testing. D2 receptor mRNA levels were found to be lower in the late postpartum NA shell and VTA compared to early postpartum, but D1 receptor mRNA levels in the NA shell were higher in the late postpartum period. Furthermore, both late postpartum and recently parturient LE mothers had higher VTA DA turnover compared to nulliparae, suggesting changes in mesolimbic signal-to-noise ratio both at the end and beginning of motherhood.
Conclusions
Collectively, our results suggest that alterations in mesolimbic DA is part of the neural substrate responsible for dynamic maternal caregiving across the entire postpartum period.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Maternal caregiving that is sensitive to the needs of the offspring is essential for normal development in many animals (Curley and Champagne 2016; Mills-Koonce et al. 2015; Worobey et al. 2009). This sensitivity requires changes in maternal motivation that match the almost constantly evolving offspring demands, thus resulting in very high levels of caregiving during the early postpartum period that progressively wane as the young mature and gain biobehavioral independence (Grieb et al. 2018; Grieb et al. 2017; Grota and Ader 1969; Pereira and Morrell 2009; Reisbick et al. 1975). Similar to other naturally rewarding behaviors such as eating (Murray et al. 2014), drinking (Bromberg-Martin and Hikosaka 2009), exercise (Greenwood and Fleshner 2011), and sexual interactions (Brom et al. 2014), the mesolimbic dopamine (DA) system underlies the motivational aspects of maternal caregiving behaviors (Numan and Stolzenberg 2009). Numerous studies have shown elevated DA release in the nucleus accumbens (NA) of postpartum rats just before and while they retrieve and lick their pups (i.e., active maternal behaviors) (Afonso et al. 2009; Champagne et al. 2004; Hansen et al. 1993; Robinson et al. 2011). Interfering with this accumbens DA neurotransmission by inactivating the ventral tegmental area (VTA) (Gaffori and Le Moal 1979; Hansen et al. 1991; Numan and Smith 1984; Numan et al. 2009), or by administering DA D1 or D2 receptor antagonists systemically (Li et al. 2004; Pereira et al. 2011; Pereira and Ferreira 2006; Silva et al. 2001; Stern and Taylor 1991; Zhao and Li 2009) or centrally (Keer and Stern 1999; Miller and Lonstein 2005; Numan et al. 2005; Parada et al. 2008; Silva et al. 2003), severely disrupts active maternal behaviors in early postpartum rats. These manipulations also reduce pup-seeking behavior and/or the encoding of pups’ incentive salience, as indicated by less bar pressing for pups in an instrumental conditioning task (Lee et al. 2000) and reduced maternal preference for the pup-associated chamber in a conditioned place preference paradigm (Fleming et al. 1994; Seip and Morrell 2009).
Importantly, almost all studies emphasizing DA’s role in maternal motivation and caregiving behaviors in laboratory rats and mice have been conducted during the early postpartum period (from parturition to postpartum day (PPD) 8), when the expression of maternal behavior is very high. As noted just above, though, the frequency and duration of maternal caregiving behaviors notably decline across the postpartum period as the offspring develop (Champagne et al. 2007; Curley et al. 2009; Grieb et al. 2017; Grota and Ader 1969; Moltz and Robbins 1965; Pereira and Morrell 2009; Pereira et al. 2008; Reisbick et al. 1975). The neurobiological factors underlying this decline are almost completely unknown (as discussed in Grieb et al. 2017), but changes in the maternal mesolimbic DA system across postpartum time may be responsible. This is suggested by the fact that DA-dependent motivation to be in contact with pups drops during the late postpartum period, simultaneous with emerging offspring independence (Mattson et al. 2001; Wansaw et al. 2008).
In the present study we hypothesized that the reduced expression of active maternal caregiving behaviors in the late postpartum period is partly due to decreased DAergic signaling. To test this hypothesis, two sets of experiments were conducted in parallel. One set of experiments examined whether repeated stimulation of DA D1 and/or D2 receptors would prevent the natural decline in maternal caregiving behaviors of late postpartum Sprague-Dawley (SD) mother rats. A separate set of experiments determined whether early and late postpartum Long-Evans (LE) mother rats differ in their DA D1 and D2 receptor gene expression in the NA shell and VTA, as well as in their levels of tyrosine hydroxylase (TH; the rate-limiting enzyme in DA synthesis) and DA turnover in the VTA. Both the SD and LE rat strains have commonly been used in the neurobiological study of maternal behavior and demonstrate a similar repertoire of active caregiving activities during the early postpartum period that are dependent on mesolimbic dopamine (Afonso et al. 2009; Champagne et al. 2004; Hansen et al. 1993; Miller and Lonstein 2005; Numan et al. 2005; Pereira et al. 2011; Robinson et al. 2011). In addition, SD and LE mothers exhibit a similar decline in active caregiving activities across the late postpartum period to match the developmental needs of their offspring (Grieb et al. 2017; McIver and Jeffrey 1967; Pereira et al. 2008; Reisbick et al. 1975; Winokur et al. 2019).
Materials and methods
Experiment 1: Effects of repeated administration of DA D1 and D2 receptor agonists on late postpartum maternal behavior
Subjects
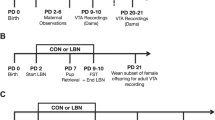
Subjects were female SD rats descended from rats purchased from Charles River Laboratories (Kingston, NY), born and raised at the Rutgers University Laboratory Animal Facility. Females’ estrous cycles were monitored daily by vaginal smearing, and females on a day of proestrus were placed overnight with a sexually experienced male from the colony. Pregnancy was confirmed the next day by semen in a vaginal smear or by the presence of a vaginal plug. Pregnant females were group housed with food (Lab Diet 2008, PMI Nutrition International, LLC, Brentwood, MO, USA) and water ad libitum, and the room was maintained on a 12-h light/dark cycle (light on at 0700 h). A few days before giving birth, females were housed in individual cages (48.5 cm × 38.5 cm × 20.5 cm) lined with fresh woodchip bedding and containing shredded paper towels as nest-building material. Soon after parturition (PPD0), litters were culled to contain 8 pups (4 males, 4 females). All procedures were performed in accordance with the principles of the National Institutes of Health Guide for Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee at Rutgers University.
Pharmacological agents and treatments
The DA D1 receptor agonist, SKF38393, and the DA D2 receptor agonist, quinpirole (Sigma Chemical, St. Louis, MO, USA), were dissolved in 0.9% saline, which also served as the vehicle control treatment. The doses and injection times of SKF38393 and quinpirole (0.5 and 0.05 mg/kg, respectively, intraperitoneal (IP), 20 min before testing) were selected based on previously published reports showing no effect of these low doses on locomotion, oral stereotypies or grooming behavior in nonmaternal animals (e.g., Berridge and Aldridge 2000; Henry et al. 1998; Hu et al. 1992), and our preliminary studies of their effects on maternal behaviors in female rats.
Experimental design and procedure
Separate groups of postpartum females were randomly assigned to receive twice-daily (0900 and 1700 h) IP injections of the following: (1) 0.5 mg/kg SKF38393 (n = 11), (2) 0.05 mg/kg quinpirole (n = 8), (3) a combination of 0.5 mg/kg SKF38393 and 0.05 mg/kg quinpirole in a single injection (n = 10), or (4) saline vehicle (n = 7). All injection volumes were 1.0 mL/kg body weight. Mother rats were treated and tested daily from PPD9 to PPD15. Twenty minutes before behavior testing, both mothers and litters were removed from their home cages, the mothers were injected and then immediately returned to their home cages. The floors of these cages were divided into four equal compartments by 5-cm high Plexiglas dividers to impede the pups from crawling and grouping themselves into the nest. Litters were individually held in small cages at room temperature for the 20 min before behavior testing.
Maternal behavior testing
Each 30-min test started with the female’s litter scattered in the home cage opposite the nest site and the frequency of each of the following maternal behaviors was continuously recorded: retrieval of pups to the nest, mouthings (oral repositioning of the pups within the nest), full body and anogenital licking of pups, and nest building. In addition, the total duration of hovering over the pups in the nest while actively performing other behaviors (e.g., licking pups or self-grooming) and quiescent nursing were recorded. Total time in contact with pups was calculated as the summed durations of hovering over the pups and nursing them. The latencies for females to begin retrieving pups, to reunite the entire litter in the nest, and to begin hovering over and nursing were recorded. The latency to begin hovering over or nursing the pups was the first occurrence of a bout of each behavior that was ≥ 2 min in duration. A latency of 1800s was assigned for any behavior that was not initiated within a 30-min observation period. Because DA agonists can induce hyperactivity and oral stereotypies (Delfs and Kelley 1990), other behaviors recorded included general exploration, self-grooming, and eating or drinking. Oral stereotypic behavior was observed on the minute during the maternal behavior test and scored if sniffing, licking, biting and/or gnawing were present. Mild stereotyped sniffing was induced in quinpirole-treated SD mothers exclusively on PPD9, and is expressed as the cumulative score over 30 min.
Statistical analyses
Maternal behavior data are expressed as means ± SEMs and as the proportion of females showing full maternal behavior during the 30-min tests as defined by retrieving all pups to the nest, licking them, and adopting a nursing posture over them. McNemar’s chi-square tests were used to analyze within-group categorical data across postpartum testing days. Between-group proportion comparisons were analyzed with Fisher’s exact test. Behavioral data were also analyzed with linear mixed models, using the best-fitting covariance structure, with drug treatment included as the between-subjects factor and postpartum testing day (PPD9–PPD15) as the within-subjects factor. When there was a statistically significant effect, non-orthogonal planned comparisons using the error term from the overall ANOVA were used to assess differences between each drug treatment and the control condition. Statistical significance was indicated by p < 0.05.
Experiment 2: Reproductive state changes in the NA and VTA DA system
Subjects
Subjects were female LE rats descended from rats purchased from Harlan Laboratories (Indianapolis, IN), born and raised in the Lonstein breeding colony at Michigan State University. Females were housed after weaning with 2 or 3 same-sex littermates in clear polypropylene cages (48 cm × 28 cm × 16 cm) containing wood chip bedding, with food (Tekland rat chow, Indianapolis, IN) and water ad libitum; the room was maintained on a 12:12 light/dark cycle (lights on at 0700 h). Females’ estrous cycles were monitored daily by vaginal smearing. For the postpartum groups, females on a day of proestrus were placed overnight with a sexually experienced male from the colony. Pregnancy was confirmed the next day by semen in a vaginal smear or by the presence of a vaginal plug. Pregnant subjects were then housed with 1–2 other pregnant females until 5–7 days before expected parturition, after which they were singly housed (diestrous virgins remained with their cage-mates until sacrifice). Soon after parturition (PPD0), litters were culled to contain 8 pups (4 males, 4 females). All procedures were performed in accordance with the principles of the National Institutes of Health Guide for Care and Use of Laboratory Animals and approved by the Institutional Animal Care and Use Committee at Michigan State University.
Dopamine receptor RT-qPCR
LE females were rendered unconscious with CO2 and rapidly decapitated on a day of diestrous for the nulliparous virgins (DV), within 3 h after delivery of the last pup (PPD0), during the early postpartum period on PPD7, or late postpartum on PPD18 (ns = 7–9/group). Brains were removed from the skull, flash frozen with isopentane, and stored at − 80 °C until sectioning. Brains were cut coronally into 300-μm-thick sections using a cryostat (Leica CM1950, Nussloch, Germany) to obtain three sections that included the NA shell and three sections that included the VTA (NA: plates 17–23; VTA: plates 76–82) (Swanson 2004). The NA shell and VTA were each extracted from the sections using a 1-mm-diameter micropuncher (Harris Micropunch, Hatfield, PA). The NA shell, rather than the NA core, was analyzed because of the association between NA shell DA and active maternal behaviors in rats (Afonso et al. 2013; Keer and Stern 1999; Li and Fleming 2003). The tissue was homogenized in RLT buffer (79216, Qiagen, Valencia, CA) containing β-mercaptoethanol by pulsed sonication for 20 s at 20% amplitude (Fisher Scientific, Pittsburgh, PA). mRNAs were then extracted using the RNeasy Plus Mini Kit (74134, Qiagen, Valencia, CA) per the manufacturer’s instructions. The extracted mRNAs were quantified using a gene quant 100 spectrophotometer (General Electric, Marlborough, MA) by measuring the 260-nm absorbance values. One hundred nanograms of mRNAs was then converted to cDNA using a high-capacity reverse transcription kit (Applied Biosystems, Foster City, CA) per the manufacturer’s instructions. After conversion to cDNA, samples were stored at − 20 °C until being analyzed with real-time RT-PCR. DA D1 and D2 receptor mRNAs were analyzed using previously used methods (Grieb et al. 2017). Briefly, each sample was run in triplicate and included cDNA, primers, and SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA) in a 25-μL reaction. A QuantStudio 3 Real-Time PCR Instrument (Product No. A28131, Applied Biosystems, Foster City, CA) was used for quantification, with the following settings: 50 °C for 2 min, 95 °C for 10 min, and 40 cycles of 95 °C for 15 s and 60 °C for 1 min. A dissociation curve was run for each sample to ensure that only a single product was transcribed. To analyze changes in DA D1 and D2 receptor mRNA, three transcripts were run: D1 (200 nM primers: Forward: 5′- GCA TGG CTT GGA TTG CTA CG -3′; Reverse: 5′- CCA GTT GCT GCC TGG ACT AA -3′), D2 (200 nM primers: Forward: 5′- GAG CCA ACC TGA AGA CAC CA –3′; Reverse: 5′- GCA TCC ATT CTC CGC CTG TT -3′), and HPRT-1 as a control gene (200 nM primers: Forward: 5′- GAA ATG TCT GTT GCT GCG TCC -3′; Reverse: 5′- GCC TAC AGG CTC ATA GTG CAA -3′). All primers were from Integrated DNA Technologies (Coralville, IA). During quantification, a no-template control was run alongside the samples to ensure that no primer-dimer amplification had occurred. In addition, mRNA samples not run through the reverse transcription kit were run simultaneously to ensure no gDNA contamination. Amplification efficiencies were calculated for each primer set, and each was within the accepted range (1.90–2.10; 2.0 is indicative of a doubling of replicants per cycle) to use the ΔΔCT method to calculate fold change between groups (Livak and Schmittgen 2001; Schmittgen and Livak 2008).
Tyrosine hydroxylase immunohistochemistry
Groups of female rats were overdosed with sodium pentobarbital either as virgins on a day of diestrous (DV), within 3 h after delivery of the last pup (PPD0), on PPD7, or on PPD18 (ns = 5/group). Parturition was monitored by spot checks every 30 min during the light photophase on the expected day of delivery, and only females whose parturitions were observed were included in the PPD0 group. Subjects were perfused transcardially with saline followed by 4% paraformaldehyde, the brains extracted, postfixed overnight, and submerged in 30% sucrose. Brains were cut into 40-μm sections in four series and sections stored in a sucrose-based cryoprotectant until processing. Three matched sections per subject containing the VTA (− 5.65 to − 6.06 mm from bregma; sections ~ 160 μm apart) were selected for analysis. Immunohistochemistry was conducted using methods previously reported in detail elsewhere (Miller and Lonstein 2009). All rinses were in 0.1 M Tris-buffered saline (TBS). Briefly, sections were incubated in 0.1% sodium borohydride for 15 min, followed by a 10-min incubation in 1% hydrogen peroxide diluted in 0.3% Triton-X TBS. Tissue was then blocked in a solution containing 20% Normal Goat Serum (NGS) in 0.3% Triton-X TBS for 1 h at room temperature. Sections were then incubated in a Triton-TBS solution containing 2% NGS and a mouse anti-tyrosine hydroxylase polyclonal antiserum (AB5986; Millipore, Burlington, MA; 1:2000), for ~ 16 h at room temperature, then in a biotinylated goat anti-mouse secondary antiserum (BA-9200; Vector Labs, Burlingame, CA; 1:500) for 1 h at room temperature. Sections were incubated in ABC solution (PK 6100, Vectors Labs, Burlingame, CA) for 1 h at room temperature. TH immunoreactivity was visualized using Vector-SG (SK-4700; Vector Labs, Burlingame, CA), and the slides were mounted and coverslipped. The number of TH-immunoreactive (TH-ir) cells in the VTA was counted bilaterally on each section under ×40 magnification (ROI superimposed on each hemisection = 245 μm × 190 μm) by experimenters naive to the subjects’ experimental condition using a Nikon Eclipse E600 light microscope. Given the very low background staining (see Fig. 4), somata with any visible TH immunoreactivity were included in the quantification. The summed number of TH-ir cells counted in all sections per subject was used for data analyses. In addition to the number of TH-ir cells in the VTA, the percentage of total area in the ROI that was covered by TH-ir pixels was also analyzed. TH-ir pixels were counted if they were darker than a standardized threshold for optical density that was set across sections and subjects, and data were represented as the percentage of TH-ir pixels/Total number of pixels in the ROI. All images were taken at a standardized light level.
High performance liquid chromatography
Additional LE females either virgins on a day of diestrus (DV), within 3 h of parturition (PPD0), on PPD7, or on PPD18 (ns = 7–8/group) were rendered unconscious with CO2 and rapidly decapitated. Brains were removed, quickly frozen with dry ice, and stored at − 80 °C until sectioning. Brains were cut coronally into 300-μm-thick sections containing the VTA using a cryostat (Leica CM 1950, Nussloch, Germany), and the VTA was removed using a 1-mm-diameter micropuncher (Harris Micropunch, Hatfield, PA) to analyze tissue levels of DA and its major metabolites. Serotonin and its major metabolite were also measured. Tissue samples were sonicated in 200 μL of an antioxidant solution (0.4 N perchloric acid, 1.34 mM EDTA, and 0.53 mM sodium metabisulfite), and 10 μL was removed and placed into 2% sodium dodecyl sulfate (SDS) for protein quantification using a BCA protein assay (Thermo-Fisher Scientific, Waltham, MA). Following protein quantification, samples were centrifuged at 10,000 rpm for 10 min. The supernatant was removed and separated by a 150 × 4.6 mm Microsorb MV C8 100-5 column (Agilent Technologies, Santa Clara, CA), and simultaneously examined for dopamine (DA), homovanillic acid (HVA), 3,4-dihydroxyphenylacetic acid (DOPAC), 5-hydroxytryptophan (5-HT), and 5-hydroxyindoleacetic acid (5-HIAA). Compounds were detected using a 12-channel coulometric array detector (CoulArray 5200, ESA, Paris, FR) attached to a Waters 2695 Solvent Delivery System (Waters, Milford, MA) under the following conditions: flow rate of 1 mL/min; detection potentials of 25, 85, 180, 420, 480 mV; and scrubbing potential of 750 mV. The mobile phase consisted of 100 mm citric acid (#A104, Thermo-Fisher Scientific, Waltham, MA), 75 mM disodium phosphate (#BP332; Thermo-Fisher Scientific, Waltham, MA), and 80 mm heptane sulfonate monohydrate (#51832, Thermo-Fisher Scientific, Waltham, MA) pH 4.25 in 5% methanol. Sample values were calculated based on a six-point standard curve of the analytes. Data are expressed as nanogram/milligram protein.
Statistics
PCR and neurochemical data are expressed as group means ± SEMs and were analyzed using one-way ANOVAs, with LSD post-hoc tests used in cases of statistically significant ANOVAs. In cases of unequal variances between groups, Welch’s ANOVAs were used. Statistical significance was indicated by p < 0.05.
Results
Experiment 1: Effects of repeated administration of DA D1 and D2 receptor agonists on late postpartum maternal behavior
As shown in Fig. 1a, there were significant effects of drug treatment on the percentage of SD dams that exhibited full maternal behavior (i.e., retrieving and grouping all pups into the nest, licking them, and adopting a nursing posture over them) during the 30-min tests. Consistent with numerous prior studies showing a decline in maternal behavior during the late postpartum period, the majority of vehicle-treated dams did not group all pups into the nest and exhibited very low levels of active caregiving from PPD12 onward relative to PPD9 (McNemar’s chi-square test, PPD12–15 versus PPD9 all ps < 0.05; Fig. 1a). In contrast, repeated treatment with SKF38393 and SKF38393 + quinpirole maintained a significantly higher proportion of fully maternal females throughout PPD12–15 (Fisher’s exact test, all ps < 0.05, Fig. 1a). A similar effect was observed in quinpirole-treated mothers, but the effect was less pronounced and only statistically significant on PPD14 when compared to the vehicle-treated group (Fisher’s exact test, p < 0.05; Fig. 1a).

a Percentage of SD dams displaying full maternal behavior (retrieving and grouping all pups to the nest, licking, and adopting a nursing posture during the 30-min tests) on PPD9–15 after being treated with either vehicle, SKF38393 (SKF), quinpirole (Quin), or SKF38393 plus quinpirole (SKF + Quin). Mean ± SEM number of b pups retrievals, c mouthing, d anogenital and e corporal licking the pups, and f nest building by PPD9–15 dams treated with vehicle, SKF, Quin, or SKF + Quin. Letter a, b, or c above lines indicates significant difference when comparing between: (a) SKF vs. vehicle, (b) Quin vs. vehicle, and (c) SKF + Quin vs. vehicle on that particular postpartum day, p < 0.05. Note: The a percentage of dams displaying full maternal behavior, the b number of pups retrieved, and c the frequency of mouthing and d, e licking the pups significantly decreased across the late postpartum days
Treatment effects were also found for the expression of discrete components of maternal caregiving, with mothers treated with SKF38393, or combined SKF38393 + quinpirole, exhibiting higher levels of nearly all active components of maternal behavior compared to controls from PPD12 onward (Retrieval: treatment, F(3,32) = 4.8, p = 0.007, ηp2 = 0.31; day, F(6,192) = 29.6, p < 0.001, ηp2 = 0.48; treatment × day, F(18,192) = 1.69, p = 0.04, ηp2 = 0.137; Mouthing: treatment, F(3,32) = 8.12, p < 0.001, ηp2 = 0.43; day, F(6,192) = 5.56, p < 0.001, ηp2 = 0.15; treatment × day, F(18,192) = 2.34, p = 0.02, ηp2 = 0.18; Corporal licking: treatment, F(3,32) = 21.59, p < 0.001, ηp2 = 0.67; day, F(6,192) = 14.42, p < 0.001, ηp2 = 0.31; treatment × day, F(18,192) = 5.88, p < 0.001, ηp2 = 0.36; Anogenital licking: treatment, F(3,32) = 17.36, p < 0.001, ηp2 = 0.62; day, F(6,192) = 9.22, p < 0.001, ηp2 = 0.22; treatment × day, F(18, 192) = 4.87, p < 0.001, ηp2 = 0.31; Nest building: treatment, F(3,32) = 7.79, p < 0.001, ηp2 = 0.42; day, F(6,192) = 5.16, p < 0.001, ηp2 = 0.14; treatment × day, F(18, 192) = 2.45, p = 0.001, ηp2 = 0.19; Fig. 1b–f). Planned comparisons revealed that co-administration of SKF38393 + quinpirole significantly increased all active caregiving behaviors compared with the vehicle-treated group throughout PPD12 to PPD15 (all ps < 0.05; Fig. 1b–f). Similarly, SKF38393 significantly increased corporal and anogenital lickings compared to vehicle on PPD12 through PPD15 (all ps < 0.05). Quinpirole transiently reduced pup licking on PPD9 (p < 0.05), but thereafter generally facilitated maternal behavior, with treated dams exhibiting significantly higher levels of retrievals as well as corporal and anogenital lickings compared to vehicle-treated females from PPD12 onward (all ps < 0.05; Fig. 1b–f).
As expected, there were significant main effects of postpartum day, but no main effects of treatment or day × treatment interactions, on the latencies for postpartum females to retrieve their first pup or group all pups into the nest, with both latencies increasing across postpartum days (Latency to first retrieval: F(6,192) = 11.73, p < 0.001, ηp2 = 0.27; Latency to reunite with litter: F(6,192) = 21.68, p < 0.001, ηp2 = 0.4; Fig. 2a, b). Notably, a significantly higher proportion of SKF38393- and SKF38393 + quinpirole-treated females retrieved and grouped pups into the nest on PPD13, PPD14, and PPD15 compared to the vehicle-treated females that mostly did not (all ps < 0.05).

Mean ± SEM latency to a retrieve their first pup, and b group all their pups into the nest, and duration of c hovering over pups in the nest and d nursing by PPD9–15 SD dams treated with either vehicle, SKF38393 (SKF), quinpirole (Quin), or SKF38393 plus quinpirole (SKF + Quin). Note: The latency to a retrieve the first pup and b group the pups into the nest significantly increased across late postpartum days, while the duration of c hovering over the pups in the nest significantly decreased across late postpartum period
Lastly, there were significant main effects of treatment and day, as well as treatment × day interactions, on the durations and types of contact that mothers had with pups. Specifically, DA agonist-treated females generally spent more time with their pups (Hover over: treatment, F(3,32) = 6.62, p = 0.001, ηp2 = 0.30; day, F(6,192) = 10.27, p < 0.001, ηp2 = 0.2; treatment × day, F(18,192) = 1.897, p = 0.02; ηp2 = 0.12; Nursing: treatment, F(3,32) = 4.5, p = 0.01, ηp2 = 0.3; day, F(6,192) = 3.1, p = 0.006, ηp2 = 0.1; treatment × day, F(18, 192) = 3.62, p < 0.01, ηp2 = 0.25; Total time in contact: treatment, F(3,32) = 3.39, p = 0.03, ηp2 = 0.24; day, F(6,192) = 2.55, p = 0.021, ηp2 = 0.074; treatment × day, F(18,192) = 3.087, p < 0.01, ηp2 = 0.22; Fig. 2c, d). However, planned comparisons revealed no significant effects of any of the three drug treatments on the duration of hovering over or the duration of nursing across the late postpartum period (Fig. 2c, d).
Importantly, home cage activity—including general exploration (as measured by line crosses across the different quadrants of the cage and rearing), self-grooming, eating, and drinking—did not differ among groups across PPD9-PPD15 (Crossing: treatment, F(3,32) = 0.86, p = ns, ηp2 = 0.075; day, F(6,192) = 0.15, p = ns, ηp2 = 0.038; treatment × day, F(18,192) = 0.29, p = ns, ηp2 = 0.057; Rearing: treatment, F(3,32) = 0.07, p = ns, ηp2 = 0.006; day, F(6,192) = 0.22, p = ns, ηp2 = 0.038; treatment × day, F(18,192) = 0.22, p = ns, ηp2 = 0.072; Self-grooming: F(3,32) = 0.86, p = ns, ηp2 = 0.075; day, F(6,192) = 0.15, p = ns, ηp2 = 0.038; treatment × day, F(18,192) = 0.29, p = ns, ηp2 = 0.057; Table 1). However, quinpirole induced mild stereotyped sniffing, exclusively in PPD9 females (treatment, F(3,32) = 3.7, p = 0.021, ηp2 = 0.26; day, F(6,192) = 20.4, p < 0.01, ηp2 = 0.39; treatment × day, F(18,192) = 3.7, p = 0.021, ηp2 = 0.26; PPD9Quinpirole 2.0 ± 0.7 vs PPD9Vehicle 0.0 ± 0.0, p = 0.014). Oral stereotypies were not observed on subsequent quinpirole treatment days or following any other drug treatment (all ps = ns; data not shown).
Experiment 2: Reproductive state changes in the NA and VTA dopamine system
In LE mothers, D2 mRNA expression in the NA shell was lower in both the PPD18 and recently parturient females compared to DVs (F(3, 28) = 4.22, p = 0.01, ηp2 = 0.31; Fig. 3a), and lower in the VTA of PPD18 dams compared to all other groups (F(3, 29) = 7.65, p < 0.01, ηp2 = 0.44; Fig. 3c). On the other hand, D1 mRNA expression in the NA shell was higher in the PPD7 and PPD18 mothers compared to DVs (F(3, 28) = 3.23, p = 0.04, ηp2 = 0.26; Fig. 3b), and there was no significant difference across reproductive state in D1 mRNA expression in the VTA (F(3, 29) = 1.44, p = 0.25, ηp2 = 0.13; Fig. 3d).

Dopamine D2 receptor mRNA expression (means ± SEMs) in the a nucleus accumbens (NA) shell and c ventral tegmental area (VTA) of LE female rats sacrificed as diestrus virgins (DV), 3 h after parturition (PPD0), on early PPD7, or on late PPD18. Dopamine D1 receptor mRNA expression (means ± SEMs) in the b NA shell and d VTA of females sacrificed as DV, PPD0, on PPD7, or on PPD18. Different letters above bars indicate significant differences between groups, p < 0.05
While the number of TH-ir cells did not differ across group (Fig 4a-c), levels of TH immunoreactivity in the VTA was significantly lower in PPD18 dams (F(3, 16) = 3.31, p < 0.05, ηp2 = 0.38; Fig. 4d) compared to all other groups. The groups also differed in their VTA DA turnover (i.e., HVA + DOPAC/DA) (F(3, 27) = 10.73, p < 0.01, ηp2 = 0.61; Fig. 4f), with PPD18 dams having higher turnover compared to DVs (p = 0.02). Recently parturient dams also had higher DA turnover in the VTA compared to DVs (p < 0.001), as well as compared to the PPD7 (p < 0.001) and PPD18 dams (p < 0.01). There were no significant effects of female reproductive state on VTA levels of DA (Fig. 4e), DOPAC, HVA, 5-HT, 5-HIAA, or 5-HT turnover (i.e., 5-HIAA/5-HT) (Table 2).

Photomicrographs of tyrosine hydroxylase (TH) immunoreactivity in the VTA of representative a diestrous virgin (DV) and b late postpartum day (PPD) 18 LE rats. c Mean ± SEM number of TH-ir cells in the VTA of females sacrificed as DV, 3 h after parturition (PPD0), on PPD7, or on PPD18. d Percentage of total VTA area covered by TH immunoreactivity of those same groups (normalized to DV group; means ± SEMs). e Mean ± SEM dopamine levels (ng/mg tissue) and f dopamine turnover (DOPAC + HVA)/DA) in the VTA. Different letters above bars indicate significant differences between groups, p < 0.05. IPN, interpeduncular nucleus; SNc, substantia nigra, pars compacta
Discussion
The present studies are important first steps in testing the hypothesis that decreased mesolimbic DA signaling at least partly underlies the decline in maternal behaviors across the early to late postpartum periods. In support of our hypothesis, in Experiment 1 we found that repeated treatment with DA agonist(s) (especially the combination of D1 and D2 receptor agonists) maintained higher expression of all active components of maternal caregiving behavior in postpartum SD rats compared to vehicle-treated controls, whose caregiving more rapidly declined during the late postpartum period. There was no consistent effect of DA agonist treatment on locomotor activity, oral stereotypies, or grooming behavior indicating that our maternal behavior results were not confounded by these known potential consequences of DA receptor agonism. In Experiment 2, we found that late postpartum LE rats had significantly lower D2 receptor expression in the NA shell and the VTA, and higher D1 receptor expression in the NA shell, compared to diestrous virgins. TH-ir optical density was also lower in the VTA of the late postpartum dams compared to either early postpartum mothers or diestrus virgins. This change in VTA TH levels was further accompanied by higher DA turnover in the late postpartum females compared to diestrus virgins. Given that SD and LE strains of rats have a very similar decline in dopamine-dependent active caregiving behaviors across the late postpartum period (Afonso et al. 2009; Champagne et al. 2004; Grieb et al. 2017; Hansen et al. 1993; Pereira et al. 2008; Reisbick et al. 1975; Winokur et al. 2019), these results collectively suggest that the mesolimbic DA system changes across female reproduction to not only stimulate maternal caregiving during the early postpartum period (Gaffori and Le Moal 1979; Hansen et al. 1991; Keer and Stern 1999; Li et al. 2004; Miller and Lonstein 2005; Numan et al. 2005; Numan and Smith 1984; Numan et al. 2009; Parada et al. 2008; Pereira et al. 2011; Pereira and Ferreira 2006; Silva et al. 2001; Stern and Taylor 1991; Zhao and Li 2009), but further contribute to the characteristic waning of caregiving as postpartum time progresses.
Repeated treatment with DA receptor agonist(s), especially the combined D1/D2 agonists treatment, maintained the full expression of maternal behavior into the late postpartum period. This was evidenced by a majority of treated SD females performing the complete repertoire of caregiving activities (i.e., completing retrieval, licking, and nursing their pups) and doing so at levels characteristics of the early postpartum period, when compared to controls, whose caregiving more rapidly declined across the late postpartum period. Although quinpirole-treated dams initially licked their pups less on PPD9, they subsequently exhibited higher levels of lickings from PPD12 to 15, compared to vehicle-treated dams. This initial disruptive effect of quinpirole on pup licking is likely related to its activation of DA D2 autoreceptors that decreases both excitability of DA neurons and DA release (Ford 2014). However, repeated administration of quinpirole reduces D2 autoreceptor sensitivity (downregulation), favoring activation of D2 postsynaptic receptors (Bartlett et al. 2005), which probably explains the subsequent higher expression of active caregiving in these females from PPD12 to 15. Another possible, non-exclusive explanation is that quinpirole’s initial effect on stereotyped sniffing might have briefly affected pup licking. Taken together, our results suggest that reduced postsynaptic DA receptor activation during mid-to-late postpartum is involved in the normal waning of active maternal behaviors. This finding is consistent with previous studies showing that interfering with DA receptor activity, via systemic or intra-accumbens administration of either DA D1 or D2 receptor antagonists, selectively disrupts active caregiving behaviors in early postpartum female rats, including both SD and LE mothers, without affecting general activity (Keer and Stern 1999; Li et al. 2004; Numan et al. 2005; Pereira and Ferreira 2006; Silva et al. 2001; Stern and Taylor 1991; Zhao and Li 2009). It is also consistent with prior results demonstrating less DA release in the NA of late postpartum female rats compared to early postpartum rats during interactions with pups (Pereira et al. 2013).
While DA receptor agonism in our study maintained significantly higher levels of active caregiving behaviors in mid-to-late lactation, it did not completely prevent the natural decline in caregiving. It may be the case that, due to decreased DA activity across the postpartum period, gradually increasing the doses of the DA agonists across testing would have been needed to fully maintain caregiving behaviors at early postpartum levels. Consistent with this possibility of decreased sensitivity of the DA system to the/its agonist(s), in Experiment 2 we did find lower DA D2 receptor mRNA in the NA shell of late postpartum compared to early postpartum LE dams. That experiment found no difference in DA D1 receptor mRNA between early and late postpartum rats, though, despite the fact that DA D2 and D1 receptor blockade produces very similar disruptions of active caregiving (Byrnes et al. 2002; Keer and Stern 1999; Silva et al. 2001). Thus, downregulation of DA D2 receptor mRNA might underlie the decline in mothering across the postpartum period, either alone or by altering the ratio of D1/D2 receptor content in the NA (Hull et al. 1989).
DA receptor agonism in Experiment 1 also may not have been able to fully maintain maternal behavior at early postpartum levels because continuous exposure to pups in the late postpartum female might have led to tolerance within the mesolimbic DA system. In this sense, long-term exposure to drugs and alcohol is associated with adaptations in the mesolimbic DA system, including decreased TH immunoreactivity (Mazei-Robison et al. 2011; Russo et al. 2007; Sklair-Tavron et al. 1996). Consistent with this, Experiment 2 found that late postpartum mothers had a significantly lower density of TH immunoreactivity in the VTA compared to early postpartum or diestrus virgin females. Low levels of TH immunoreactivity in the VTA is associated with reduced capacity for DA synthesis (Bacopoulos and Bhatnagar 1977) and less responsiveness to rewarding stimuli (Russo et al. 2007). We also found that the late postpartum dams had higher DA turnover in the VTA, which may have been driven by their decreased VTA D2 expression in response to chronically high DA (Bartlett et al. 2005; Ford 2014). Higher DA turnover within the VTA activates D2 autoreceptors on VTA neurons, subsequently decreasing DA cell firing (Beaulieu and Gainetdinov 2011; Ford 2014; Gantz et al. 2013; Pucak and Grace 1994) and distal DA release in regions such as the NA and mPFC (Andersson et al. 2006; Mebel et al. 2012; Rice and Patel 2015). Decreased DA release at target sites, including the NA shell, could ultimately lead to decreased caregiving during the late postpartum period (Andersson et al. 2006; Kliem et al. 2007; Pruett and Salvatore 2013).
We also found that recently parturient LE rats (PPD0) had higher DA turnover in the VTA compared to virgins. These diffferences in DA turnover in the VTA can be contrasted with the stable DA turnover previously reported in the NA across the postpartum period (Lonstein et al. 2003; Winokur et al. 2019). While both PPD0 and PPD18 dams had high DA turnover in the VTA, the behavioral consequences probably differ between the groups. For instance, if high DA turnover in recently parturient dams decreases basal DA release in the NA shell compared to virgins, it may enhance new mothers’ signal-to-noise ratio for DAergic responses to pup-related stimuli (Afonso et al. 2009; Afonso et al. 2013). This increased signal-to-noise ratio would only occur in the early postpartum period when there is high DA release in response to the pups’ cues (Afonso et al. 2009; Champagne et al. 2004; Hansen et al. 1993; Pereira et al. 2013). Therefore, increased VTA DA turnover soon after parturition could be associated with enhanced signal-to-noise ratio to help new mothers distinguish between basal and pup-stimulated DA release, while increased VTA DA turnover in the late postpartum period would likely lead to decreased DA output as discussed just above.
In conclusion, our results support the hypothesis that reduced expression of active maternal caregiving behaviors in the late postpartum period is partly due to decreasing DAergic function across postpartum time . Given that both offspring age and maternal postpartum stage drive this decline in maternal caregiving behaviors (Grieb et al. 2018; Pereira et al. 2008; Reisbick et al. 1975; Rosenblatt 1969), it would be valuable for future research to disentangle their respective roles in the dynamic maternal mesolimbic system and its contribution to behavioral change across motherhood.
References
Afonso VM, King S, Chatterjee D, Fleming AS (2009) Hormones that increase maternal responsiveness affect accumbal dopaminergic responses to pup-and food-stimuli in the female rat. Horm Behav 56(1):11–23
Afonso VM, Shams WM, Jin D, Fleming AS (2013) Distal pup cues evoke dopamine responses in hormonally primed rats in the absence of pup experience or ongoing maternal behavior. J Neurosci 33(6):2305–2312
Andersson DR, Nissbrandt H, Bergquist F (2006) Partial depletion of dopamine in substantia nigra impairs motor performance without altering striatal dopamine neurotransmission. Eur J Neurosci 24(2):617–624
Bacopoulos NG, Bhatnagar RK (1977) Correlation between tyrosine hydroxylase activity and catecholamine concentration or turnover in brain regions. J Neurochem 29(4):639–643
Bartlett SE, Enquist J, Hopf FW, Lee JH, Gladher F, Kharazia V et al (2005) Dopamine responsiveness is regulated by targeted sorting of D2 receptors. Proc Natl Acad Sci 102(32):11521–11526
Beaulieu JM, Gainetdinov RR (2011) The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev 63(1):182–217
Berridge KC, Aldridge JW (2000) Super-stereotypy I: enhancement of a complex movement sequence by systemic dopamine D1 agonists. Synapse 37(3):194–204
Brom M, Both S, Laan E, Everaerd W, Spinhoven P (2014) The role of conditioning, learning and dopamine in sexual behavior: a narrative review of animal and human studies. Neurosci Biobehav Rev 38:38–59
Bromberg-Martin ES, Hikosaka O (2009) Midbrain dopamine neurons signal preference for advance information about upcoming rewards. Neuron 63(1):119–126
Byrnes EM, Rigero BA, Bridges RS (2002) Dopamine antagonists during parturition disrupt maternal care and the retention of maternal behavior in rats. Pharmacol Biochem Behav 73(4):869–875
Champagne FA, Chretien P, Stevenson CW, Zhang TY, Gratton A, Meaney MJ (2004) Variations in nucleus accumbens dopamine associated with individual differences in maternal behavior in the rat. J Neurosci 24(17):4113–4123
Champagne FA, Curley JP, Keverne EB, Bateson PP (2007) Natural variations in postpartum maternal care in inbred and outbred mice. Physiol Behav 91(2–3):325–334
Curley JP, Champagne FA (2016) Influence of maternal care on the developing brain: mechanisms, temporal dynamics and sensitive periods. Front Neuroendocrinol 40:52–66
Curley JP, Jordan ER, Swaney WT, Izraelit A, Kammel S, Champagne FA (2009) The meaning of weaning: influence of the weaning period on behavioral development in mice. Dev Neurosci 31(4):318–331
Delfs J, Kelley AJN (1990) The role of D1 and D2 dopamine receptors in oral stereotypy induced by dopaminergic stimulation of the ventrolateral striatum. Neuroscience 39(1):59–67
Fleming AS, Korsmit M, Deller M (1994) Rat pups are potent reinforcers to the maternal animal: effects of experience, parity, hormones, and dopamine function. Psychobiology 22(1):44–53
Ford CP (2014) The role of D2-autoreceptors in regulating dopamine neuron activity and transmission. Neuroscience 282:13–22
Gaffori O, Le Moal M (1979) Disruption of maternal behavior and appearance of cannibalism after ventral mesencephalic tegmentum lesions. Physiol Behav 23(2):317–323
Gantz SC, Bunzow JR, Williams JT (2013) Spontaneous inhibitory synaptic currents mediated by a G protein-coupled receptor. Neuron 78(5):807–812
Greenwood BN, Fleshner M (2011) Exercise, stress resistance, and central serotonergic systems. Exerc Sport Sci Rev 39(3):140–149
Grieb ZA, Tierney SM, Lonstein JS (2017) Postpartum inhibition of ovarian steroid action increases aspects of maternal caregiving and reduces medial preoptic area progesterone receptor expression in female rats. Horm Behav 96:31–41
Grieb ZA, Holschbach MA, Lonstein JS (2018) Interaction between postpartum stage and litter age on maternal caregiving and medial preoptic area orexin. Physiol Behav 194:430–436
Grota LJ, Ader R (1969) Continuous recording of maternal behaviour in Rattus norvegicus. Anim Behav 17(4):722–729
Hansen S, Harthon C, Wallin E, Löfberg L, Svensson K (1991) The effects of 6-OHDA-induced dopamine depletions in the ventral or dorsal striatum on maternal and sexual behavior in the female rat. Pharmacol Biochem Behav 39(1):71–77
Hansen S, Bergvall ÅH, Nyiredi S (1993) Interaction with pups enhances dopamine release in the ventral striatum of maternal rats: a microdialysis study. Pharmacol Biochem Behav 45(3):673–676
Henry DJ, Hu XT, White FJ (1998) Adaptations in the mesoaccumbens dopamine system resulting from repeated administration of dopamine D1 and D2 receptor-selective agonists: relevance to cocaine sensitization. Psychopharmacology 140(2):233–242
Hu XT, Brooderson RJ, White FJ (1992) Repeated stimulation of D1 dopamine receptors causes time-dependent alterations in the sensitivity of both D1 and D2 dopamine receptors within the rat striatum. Neuroscience 50(1):137–147
Hull EM, Warner RK, Bazzett TJ, Eaton RC, Thompson JT, Scaletta LL (1989) D2/D1 ratio in the medial preoptic area affects copulation of male rats. J Pharmacol Exp Ther 251(2):422–427
Keer SE, Stern JM (1999) Dopamine receptor blockade in the nucleus accumbens inhibits maternal retrieval and licking, but enhances nursing behavior in lactating rats. Physiol Behav 67(5):659–669
Kliem MA, Maidment NT, Ackerson LC, Chen S, Smith Y, Wichmann T (2007) Activation of nigral and pallidal dopamine D1-like receptors modulates basal ganglia outflow in monkeys. J Neurophysiol 98(3):1489–1500
Lee A, Clancy S, Fleming AS (2000) Mother rats bar-press for pups: effects of lesions of the mpoa and limbic sites on maternal behavior and operant responding for pup-reinforcement. Behav Brain Res 108(2):215–231
Li M, Fleming AS (2003) The nucleus accumbens shell is critical for normal expression of pup-retrieval in postpartum female rats. Behav Brain Res 145(1–2):99–111
Li M, Davidson P, Budin R, Kapur S, Fleming AS (2004) Effects of typical and atypical antipsychotic drugs on maternal behavior in postpartum female rats. Schizophr Res 70(1):69–80
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25(4):402–408
Lonstein JS, Dominguez JM, Putnam SK, De Vries GJ, Hull EM (2003) Intracellular preoptic and striatal monoamines in pregnant and lactating rats: possible role in maternal behavior. Brain Res 970(1–2):149–158
Mattson BJ, Williams S, Rosenblatt JS, Morrell JI (2001) Comparison of two positive reinforcing stimuli: pups and cocaine throughout the postpartum period. Behav Neurosci 115(3):683–694
Mazei-Robison MS, Koo JW, Friedman AK, Lansink CS, Robison AJ, Vinish M, Niswender KD (2011) Role for mTOR signaling and neuronal activity in morphine-induced adaptations in ventral tegmental area dopamine neurons. Neuron 72(6):977–990
McIver, A. H., & Jeffrey, W. E. J. B. (1967). Strain differences in maternal behavior in rats. 210-216
Mebel DM, Wong JC, Dong YJ, Borgland SL (2012) Insulin in the ventral tegmental area reduces hedonic feeding and suppresses dopamine concentration via increased reuptake. Eur J Neurosci 36(3):2336–2346
Miller SM, Lonstein JS (2005) Dopamine d1 and d2 receptor antagonism in the preoptic area produces different effects on maternal behavior in lactating rats. Behav Neurosci 119(4):1072–1083
Miller SM, Lonstein JS (2009) Dopaminergic projections to the medial preoptic area of postpartum rats. Neuroscience 159(4):1384–1396
Mills-Koonce WR, Willoughby MT, Zvara B, Barnett M, Gustafsson H, Cox MJ, Family Life Project Key Investigators (2015) Mothers’ and fathers’ sensitivity and children’s cognitive development in low-income, rural families. J Appl Dev Psychol 38:1–10
Moltz H, Robbins D (1965) Maternal behavior of primiparous and multiparous rats. J Comp Physiol Psychol 60(3):417–421
Murray S, Tulloch A, Gold MS, Avena NM (2014) Hormonal and neural mechanisms of food reward, eating behaviour and obesity. Nat Rev Endocrinol 10(9):540
Numan M, Smith HG (1984) Maternal behavior in rats: evidence for the involvement of preoptic projections to the ventral tegmental area. Behav Neurosci 98(4):712–727
Numan M, Stolzenberg DS (2009) Medial preoptic area interactions with dopamine neural systems in the control of the onset and maintenance of maternal behavior in rats. Front Neuroendocrinol 30(1):46–64
Numan M, Numan MJ, Pliakou N, Stolzenberg DS, Mullins OJ, Murphy JM, Smith CD (2005) The effects of D1 or D2 dopamine receptor antagonism in the medial preoptic area, ventral pallidum, or nucleus accumbens on the maternal retrieval response and other aspects of maternal behavior in rats. Behav Neurosci 119(6):1588–1604
Numan M, Stolzenberg DS, Dellevigne AA, Correnti CM, Numan MJ (2009) Temporary inactivation of ventral tegmental area neurons with either muscimol or baclofen reversibly disrupts maternal behavior in rats through different underlying mechanisms. Behav Neurosci 123(4):740–751
Parada M, King S, Li M, Fleming AS (2008) The roles of accumbal dopamine D1 and D2 receptors in maternal memory in rats. Behav Neurosci 122(2):368–376
Pereira M, Ferreira A (2006) Demanding pups improve maternal behavioral impairments in sensitized and haloperidol-treated lactating female rats. Behav Brain Res 175(1):139–148
Pereira M, Morrell JI (2009) The changing role of the medial preoptic area in the regulation of maternal behavior across the postpartum period: facilitation followed by inhibition. Behav Brain Res 205(1):238–248
Pereira M, Seip K, Morrell JI, Bridges R (2008) Maternal motivation and its neural substrate across the postpartum period. Neurobiology of the parental brain:39–60
Pereira M, Farrar AM, Hockemeyer J, Müller CE, Salamone JD, Morrell JI (2011) Effect of the adenosine A 2A receptor antagonist MSX-3 on motivational disruptions of maternal behavior induced by dopamine antagonism in the early postpartum rat. Psychopharmacology 213(1):69–79
Pereira, M., Farrar, A. M., Morrell, J. I., & Abercrombie, E. D. (2013). Changes in nucleus accumbens dopamine release during mother-pup interactions at early and late postpartum stages. In V Parental brain conference, Germany
Pruett BS, Salvatore MF (2013) Nigral GFRα1 infusion in aged rats increases locomotor activity, nigral tyrosine hydroxylase, and dopamine content in synchronicity. Mol Neurobiol 47(3):988–999
Pucak ML, Grace AA (1994) Evidence that systemically administered dopamine antagonists activate dopamine neuron firing primarily by blockade of somatodendritic autoreceptors. J Pharmacol Exp Ther 271(3):1181–1192
Reisbick S, Rosenblatt JS, Mayer AD (1975) Decline of maternal behavior in the virgin and lactating rat. J Comp Physiol Psychol 89(7):722–732
Rice ME, Patel JC (2015) Somatodendritic dopamine release: recent mechanistic insights. Philosophical Transactions of the Royal Society B: Biological Sciences 370(1672):20140185
Robinson DL, Zitzman DL, Williams SK (2011) Mesolimbic dopamine transients in motivated behaviors: focus on maternal behavior. Frontiers in psychiatry 2:23
Rosenblatt JS (1969) The development of maternal responsiveness in the rat. Am J Orthop 39(1):36–56
Russo SJ, Bolanos CA, Theobald DE, DeCarolis NA, Renthal W, Kumar A et al (2007) IRS2-Akt pathway in midbrain dopamine neurons regulates behavioral and cellular responses to opiates. Nat Neurosci 10(1):93–99
Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by the comparative C T method. Nat Protoc 3(6):1101–1108
Seip KM, Morrell JI (2009) Transient inactivation of the ventral tegmental area selectively disrupts the expression of conditioned place preference for pup-but not cocaine-paired contexts. Behav Neurosci 123(6):1325–1338
Silva MRP, Bernardi MM, Felicio LF (2001) Effects of dopamine receptor antagonists on ongoing maternal behavior in rats. Pharmacol Biochem Behav 68(3):461–468
Silva MRP, Bernardi MM, Cruz-Casallas PE, Felicio LF (2003) Pimozide injections into the nucleus accumbens disrupt maternal behaviour in lactating rats. Pharmacol Toxicol 93(1):42–47
Sklair-Tavron L, Shi WX, Lane SB, Harris HW, Bunney BS, Nestler EJ (1996) Chronic morphine induces visible changes in the morphology of mesolimbic dopamine neurons. Proc Natl Acad Sci 93(20):11202–11207
Stern JM, Taylor LA (1991) Haloperidol inhibits maternal retrieval and licking, but enhances nursing behavior and litter weight gains in lactating rats. J Neuroendocrinol 3(6):591–596
Swanson, L. (2004). Brain maps: structure of the rat brain. Gulf Professional Publishing
Wansaw MP, Pereira M, Morrell JI (2008) Characterization of maternal motivation in the lactating rat: contrasts between early and late postpartum responses. Horm Behav 54(2):294–301
Winokur, S. B., Lopes, K. L., Moparthi, Y., & Pereira, M. (2019). Depression-related disturbances in rat maternal behaviour are associated with altered monoamine levels within mesocorticolimbic structures. J Neuroendocrinol, e12766
Worobey J, Lopez MI, Hoffman DJ (2009) Maternal behavior and infant weight gain in the first year. J Nutr Educ Behav 41(3):169–175
Zhao C, Li M (2009) The receptor mechanisms underlying the disruptive effects of haloperidol and clozapine on rat maternal behavior: a double dissociation between dopamine D2 and 5-HT2A/2C receptors. Pharmacol Biochem Behav 93(4):433–442
Acknowledgments
The authors would like to thank Dr. Jack W. Lipton and Allyson Cole-Strauss for their assistance with the HPLC analysis.
Funding
These studies were funded by a NARSAD Young Investigator Award and NIDA SOAR DA027945 awarded to MP, DA014025 to JIM, and HD097085 to JSL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to report and are in control of all primary data. On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Grieb, Z.A., Vitale, E.M., Morrell, J.I. et al. Decreased mesolimbic dopaminergic signaling underlies the waning of maternal caregiving across the postpartum period in rats. Psychopharmacology 237, 1107–1119 (2020). https://doi.org/10.1007/s00213-019-05441-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-019-05441-7