
Abstract
Rationale
Aggressive behavior and impaired impulse control have been associated with dysregulations in the serotonergic system and with impaired functioning of the prefrontal cortex. 5-HT1B receptors have been shown to specifically modulate several types of offensive aggression.
Objective
This study aims to characterize the relative importance of two populations of 5-HT1B receptors in the dorsal raphé nucleus (DRN) and infralimbic cortex (ILC) in the modulation of aggressive behavior.
Methods
Male CFW mice were conditioned on a fixed-ratio 5 schedule of reinforcement to self-administer a 6% (w/v) alcohol solution. Mice repeatedly engaged in 5-min aggressive confrontations until aggressive behavior stabilized. Next, a cannula was implanted into either the DRN or the ILC. After recovery, mice were tested for aggression after self-administration of either 1.0 g/kg alcohol or water prior to a microinjection of the 5-HT1B agonist, CP-93,129 (0–1.0 μg/infusion).
Results
In both the DRN and ILC, CP-93,129 reduced aggressive behaviors after both water and alcohol self-administration. Intra-raphé CP-93,129 dose-dependently reduced both aggressive and locomotor behaviors. However, the anti-aggressive effects of intra-cortical CP-93,129 were behaviorally specific.
Conclusions
These findings highlight the importance of the serotonergic system in the modulation of aggression and suggest that the behaviorally specific effects of 5-HT1B receptor agonists are regionally selective. 5-HT1B receptors in a medial subregion of the prefrontal cortex, the ILC, appear to be critically involved in the attenuation of species-typical levels of aggression.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
High rates of violence and aggression occur under the influence of alcohol in a subset of humans (Virkkunen et al. 1994), primates (Miczek et al. 1993; Weerts et al. 1993), and rodents (Lister and Hilakivi 1988; Miczek et al. 1998; Miczek and O’Donnell 1980; Miczek et al. 1992) who show a consistent pro-aggressive behavioral response to moderate doses of alcohol. Those individuals who are sensitive to the aggression-heightening effects of alcohol have a distinct neurochemical, pharmacological, and behavioral profile characterized by low central levels of the 5-HT metabolite-5-hydroxy-indole acetic acid (5-HIAA) and 5-HT transporter (5-HTT). In both humans and non-human primates, reduced levels of these markers of 5-HT metabolism and activity are inversely correlated with heightened aggressive and violent behavior (Brown et al. 1979; Ferrari et al. 2005; Higley et al. 1996a; Reif et al. 2007), impulsive and risky behavior (Fairbanks et al. 2001; Frankle et al. 2005; Linnoila et al. 1983; Mehlman et al. 1994; Virkkunen et al. 1996; Virkkunen and Linnoila 1993), excessive alcohol consumption (Higley and Linnoila 1997; Higley et al. 1996b), and a family history of alcoholism (Cloninger 1987; Higley and Linnoila 1997; Higley et al. 1996b). Preclinical studies in rodents have similarly found blunted levels of 5-HT metabolites in aggressive mice (Giacalone et al. 1968; van der Vegt et al. 2003), observed that genetically modified mice that lack the 5-HTT gene are less aggressive than their wild-type counterparts (Holmes et al. 2002) and, using in vivo microdialysis, shown that cortical levels of 5-HT in the rat are significantly blunted during and after an aggressive encounter (van Erp and Miczek 2000). Together, these data point to an impulsive, pathological phenotype that is characterized by escalated levels of aggression and dampened serotonergic activity.
Mechanistically, the neural circuitry of species-typical aggression and aggression after alcohol self-administration may share common elements in the prefrontal cortex (PFC; Giancola 2000). One key role of the ventral medial prefrontal cortex is to exert inhibitory control over impulsive behaviors; impaired function of the prefrontal cortex is a hallmark feature of individuals diagnosed with high trait impulsivity (Best et al. 2002; Davidson et al. 2000; Fineberg et al. 2010; Kable and Glimcher 2007; Sripada et al. 2010). Likewise, highly impulsive and violent individuals have reduced metabolic activity in the frontal cortex and are less able to regulate their behavioral actions (Raine et al. 1994; Volkow et al. 1994; Yang and Raine 2009). In rodents, levels of c-Fos immunoreactivity are significantly increased in the prefrontal cortex immediately after an aggressive encounter (Halasz et al. 2006; Haller et al. 2006) and extracellular levels of 5-HT are elevated in the medial but not orbital prefrontal cortex of rats during performance of a delay discounting task (Winstanley et al. 2006). Similarly, mice that have been selectively bred for high and low levels of aggression have increased cortical levels of 5-HT after repeated resident–intruder confrontations, although this increase is blunted in the highly aggressive animals (Caramaschi et al. 2008). Specifically, the experience of repeatedly “winning” aggressive confrontations seems to functionally alter serotonergic tone in the prefrontal cortex. Together, these studies provide evidence to suggest that engaging in acts of aggression and impulsive behavior leads to an acute increase in cortical activity yet repeated bouts of high levels of aggression downregulates basal 5-HT activity relative to non-aggressive counterparts. These data indicate that the prefrontal cortex is actively recruited during performance of impulsive and aggressive behaviors and that prefrontal cortical 5-HT regulates both the immediate and long-lasting consequences of these behaviors.
5-HT1B receptors are located both pre- and post-synaptically throughout the central nervous system and modulate a wide variety of behaviors including sex, feeding, drug self-administration, sleep, anxiety, stress, depression, and aggression (Ahlenius and Larsson 1998; Barnes and Sharp 1999; Bouwknecht et al. 2001; Clark and Neumaier 2001; Clark et al. 2002, 2004; Fish et al. 1999; Lee and Simansky 1997; Moret and Briley 2000; Parsons et al. 1998; Stern et al. 1998; Zhuang et al. 1999). Genetic deletion of the 5-HT1B receptor can lead to heightened aggression and impulsivity, and allelic variation of the 5-HT1B receptor has been linked to impulsive aggression, antisocial personality disorder, and alcoholism (Fehr et al. 2000; Lappalainen et al. 1998, but see New et al. 2001; Sinha et al. 2003). The association of this polymorphism with impulsive aggression suggests that a higher frequency of the 5-HT1B receptor gene may contribute to a trait-like phenotype that predisposes individuals to engage in maladaptive patterns of aggression and alcohol drinking. Pharmacologically, 5-HT1B receptor agonists selectively attenuate alcohol-heightened aggression with greater potency than non-heightened aggression, in both mice and humans (Fish et al. 1999, 2008; Gowin et al. 2010). Site-specific microinjection of the 5-HT1B agonists, CP-94,253, and CP-93,129 into the ventral orbital prefrontal cortex (VO-PFC), potently and efficaciously reduce species-typical and escalated offensive aggression (Centenaro et al. 2008; De Almeida et al. 2006). In contrast, alcohol-heightened aggression is significantly increased after microinjection of CP-94,253 into the infralimbic (ILC) but not orbitofrontal region of the prefrontal cortex (Faccidomo et al. 2008). Collectively, these studies suggest that prefrontal 5-HT1B receptors are critically involved in the modulation of both species-typical and escalated aggression, an effect that depends on the targeted subregion.
The primary goal of this study was to compare the relative importance of two distinct populations of 5-HT1B receptors, those located in the dorsal raphé nucleus (DRN) and those in the PFC, on the modulation of aggressive behavior. To achieve this goal, we used a 5-HT1B receptor agonist that was highly selective for the 5-HT1B receptor, relative to other 5-HT receptors (i.e., 5-HT1A and 5-HT1D receptors that are abundant in these regions). CP-94,253 and CP-93,129 have different dissociation constants for the 5-HT1B receptor (5-HT1B K d = 2 nM vs. 55 nM, respectively; Koe et al. 1992a, b) with CP-94,253 having a slightly greater affinity for the 5-HT1B receptor than CP-93,129. However, CP-93,129 has a much lower affinity for the 5-HT1A receptor than does CP-94,253 (5-HT1A K d = 1,500 nM vs. 89 nM, respectively) and is therefore more selective for the 5-HT1B receptor. Moreover, Bannai et al. (2007) previously found that intra-raphé injection of CP-93,129 decreased escalated aggression that is observed in animals when they perform an operant conditioning task that is reinforced by the opportunity to engage in an aggressive confrontation. However, the importance of this population of 5-HT1B receptors in the DRN on species-typical and aggression after alcohol self-administration has not been fully characterized. Thus, we chose to use the 5-HT1B agonist, CP-93,129, to preferentially activate 5-HT1B receptors in the DRN and ILC, in order to assess the importance of these two populations of receptors on the modulation of aggressive behavior.
Materials and methods
Subjects
Male “resident” Swiss Webster mice (n = 23; CFW, Charles River Labs, Wilmington, MA) were pair-housed upon arrival with a conspecific female in clear, polycarbonate cages (28 × 17 cm) lined with pine shavings. Purina rodent chow was freely available through the cage lid and water was available for 3 h/day. Mouse litters were weaned at 21 days postpartum. “Intruders” were male CFW mice (n = 46) that were housed in groups of 8–12 in a large polycarbonate cage (48 × 26 cm) lined with corn cob bedding, with unlimited access to food and water. The vivarium was maintained on a 12-h light/dark photocycle (lights off at 0700 hours), with temperature set at 21 ± 1°C and 23% humidity. All mice were cared for according to the Guide for the Care and Use of Laboratory Animals (National Research Council 1996) and the Tufts University IACUC approved all experimental procedures.
Alcohol self-administration apparatus
Resident male mice were trained to emit an operant response (nose-poke) on a custom-designed aluminum panel (16.5 × 3.8 × 15.9 cm) that was inserted into their home cage with thumb screws (see Miczek and de Almeida 2001 and Faccidomo et al. 2008 for details). Briefly, each side of the panel contained a cue light positioned above a drinking trough (3 × 5 cm; Med Associates; Georgia, VT). A photobeam spanned the entrance of the drinking trough and a nose-poke was recorded when the photobeam was disrupted, causing a relay click as auditory feedback. Each trough was connected to a syringe pump (Med Associates). The panel and pump were connected to an interface and computer that recorded the behavior of each mouse (MED-PC for Windows v.4.1; Med Associates). A white house light was illuminated throughout the session and a 28-V fan minimized external noise.
Acquisition of alcohol self-administration
A modified sucrose fading procedure was used to facilitate alcohol self-administration (Faccidomo et al. 2008; Samson 1986). Prior to the first drinking session, the resident was acclimated to the initial reinforcing solution by presentation of a plastic bottle containing a sucrose (10% w/v) solution through the cage lid for 16 h. The next day, fluid access was restricted for 21 h prior to the first self-administration session. Each session began by removing the female and pups, then placing the operant conditioning panel into the home cage. Initially, every nose-poke into either trough was reinforced (fixed-ratio 1 schedule; FR 1) by a 0.05-ml delivery of a 10% sucrose solution. Each reinforcement was accompanied by a brief noise and absence of the house light, both of which served as secondary cues. Responses that occurred during delivery of the reinforcement (1.26 s) were recorded but were not reinforced. During the second session, responding on only the “active” nose-poke (“active” side was counterbalanced across mice) was reinforced and the reinforcement schedule was increased to FR 5. Responding on the “inactive” nose-poke was recorded but held no contingencies. In subsequent sessions, alcohol was gradually added to the reinforcing solution in 1% increments up to 6%. Next, the sucrose concentration in the reinforcing solution was decreased in 1% decrements until residents were self-administering unsweetened alcohol (6% w/v). Initially, sessions lasted for 30 min, after which the mice were reunited with their cage mate and given access to water for 3 h. When the mice began self-administering a 5% sucrose/6% alcohol solution, the duration of the session was shortened to prevent severe intoxication. Sessions were terminated after each resident consumed 1.0 g/kg alcohol. Drinking sessions occurred 5 days per week between 0700 and 1400 hours.
Apparatus and measurements
Aggressive confrontations were videotaped using a low-lux video camera (Panasonic BL-200) connected to a VCR and monitor. All confrontations were analyzed by trained observers using a custom-designed keyboard connected to a computer running The Observer software (Noldus, The Observer v.5.0; Wageningen, The Netherlands). The frequencies and durations of salient aggressive (attack bites, sideways threat, tail rattles, and pursuit) and non-aggressive (grooming, rearing, and walking) behaviors were quantified according to the operational descriptions provided by Grant and Macintosh (1963) and Miczek and O’Donnell (1978).
Resident–intruder confrontations
After 3 weeks of pair-housing, aggression-naive residents were screened for aggressive behavior until stable baseline levels of aggression emerged (ca. 6–10 confrontations with less than 15% variability; Miczek and O’Donnell 1978). Aggressive confrontations began by removing the female and pups and introducing a male “intruder” mouse into the home cage of the resident. Confrontations lasted for 5 min after the first attack bite or for 5 min if no attack was initiated. After the confrontation, the female and pups were reunited with their resident cage mate. Importantly, each resident repeatedly confronted the same intruder and aggressive intruders (ca. one intruder/cage) were excluded as stimulus animals.
Drugs
Ninety-five percent ethyl alcohol (Pharmco Products Inc.; Brookfield, CT) was diluted with tap water to 6% (w/v). The 5-HT1B agonist CP-93,129 (3-(1,2,5,6-tetrahydropyrid-4-yl)pyrrollo[3,2-b]pyrid-5-one, generously donated by Pfizer, Groton, CT) was freshly dissolved in aCSF (in millimoles, 147 NaCl, 1.3 anhydrous CaCl2, 0.9 anhydrous MgCl2, 4.0 KCl, pH = 6.7–7).
Experiment 1: intra-raphé microinjection and aggression
Residents (n = 14) were anesthetized with a ketamine (100 mg/kg)/xylazine (10 mg/kg) cocktail, i.p., placed into a stereotaxic frame (Kopf Instruments; Tujunga, CA) and implanted with a 26-gauge guide cannula (Plastics One; Roanoke, VA) aimed at the DRN (AP −4.4 mm; ML ±0; DV −1.7 mm from bregma; 26° angle; Franklin and Paxinos 2001). A 33-gauge obdurator (Plastics One), extending 0.5 mm beyond the cannula tip, was inserted after surgery and moved daily to prevent blockage and scarring. An aversive tasting quinine polish (Bite It©) coated the headmount and obdurator to prevent gnawing damage by the female cagemate. After 1–2 weeks recovery, residents resumed alcohol self-administration and aggression testing.
On test days, mice consumed water or 1.0 g/kg alcohol immediately before microinjection of either artificial cerebrospinal fluid (aCSF) or CP-93,129 (0.1–1.0 μg) via a 33 gauge injector that extended 2 mm beneath the guide cannulae. Eight microinjections were conducted using a Latin Square design with a minimum of 2 tests/week. The injector was connected to a glass syringe (CMA Microdialysis, North Chelmsford, MA) and pump that infused 0.5 μl over 4 min (0.125 μl/min). The injector remained in place for 1 min after the infusion to allow for diffusion and to minimize vertical capillary action along the injection tract. Mice were unrestrained during the infusion and a 5 min aggressive confrontation commenced 10 min after the injection.
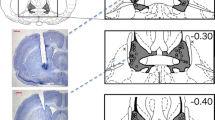
Immediately after the final aggression test, mice were deeply anesthesized (Avertin®) and intracardially perfused with 0.9% saline and 4% paraformaldehyde. To verify implant position, the brains were sliced on a sliding microtome in 60-μm coronal sections, and stained with cresyl violet. Histological verification of cannula placement revealed that four residents had cannulae placements outside of the DRN; these residents were excluded from the final analysis (Fig. 1a, b). The cannula of one mouse became obstructed after completion of four out of eight treatment conditions; this resident was also excluded from the final analysis.

A schematic representation of mouse dorsal raphé (a) and infralimbic cortex (c) coronal sections adapted from Franklin and Paxinos (2001). Filled circles indicate the approximate site of an accurately placed injection (n = 9 for DRN; n = 7 for ILC). Filled triangles indicate the approximate site of missed placements (n = 4 for DRN; n = 2 for ILC). b and d are representative photomicrographs of mouse brain coronal sections (×2) that were stained with cresyl violet to visualize the injection site
Experiment 2: intra-cortical microinjection and aggression
A second group of residents (n = 9) was anesthetized with Avertin® (2,2,2 tribromoethanol; 400 mg/kg, i.p.) and implanted with a cannula aimed at either the right or the left infralimbic cortex (ILC; AP +1.7 mm; ML ±0.4 mm; DV −1.2 mm, from dura). They were tested for aggression 10 min after consuming water or 1.0 g/kg alcohol and a microinjection of either aCSF or CP-93,129 (0.1–1.0 μg). Eight microinjections were conducted using a Latin Square design with a minimum of two tests per week. Histological verification of cannula placement revealed that two residents had cannulae placements within the lateral septum; these residents were excluded from the final analysis (Fig. 1c, d).
Statistical analysis
The frequencies of the aggressive behaviors and the durations of the non-aggressive behaviors were separately analyzed using a two-way repeated measures ANOVA. The Holm–Sidak post-hoc test was run when appropriate, using the aCSF and water tests as the control conditions. α was set at 0.05 for all comparisons.
Results
Experiment 1: intra-raphé microinjection and aggression
Intra-raphé administration of the selective 5-HT1B agonist, CP-93,129 produced a dose-dependent decrease in aggressive behavior after consumption of 1.0 g/kg alcohol or water. Specifically, a significant main effect of drug was found on the frequency of attack bites (F(3,24) = 22.0, p < 0.001; Fig. 2, Table 1), sideways threats (F(3,24) = 15.8, p < 0.001; Table 1), and tail rattles (F(3,24) = 9.9, p < 0.001; Table 1). Post-hoc tests revealed that this effect was due to a reduction of these aggressive behaviors after infusion of 0.5 and 1.0 μg CP-93,129, irrespective of whether the resident consumed water or alcohol prior to the infusion. There was also a significant main effect of CP-93,129 on the duration of attack bites (F(3,24) = 14.4, p < 0.001; Table 1) and sideways threats (F(3,24) = 6.8, p = 0.002; Table 1). Post-hoc tests revealed that this effect was due to a reduction of both behaviors after infusion of 1.0 μg CP-93,129 and a reduction in the duration of attack bites after infusion of 0.5 μg CP-93,129. Aggressive pursuit of the intruder rarely occurred and was unaffected by any treatment condition.

Microinjection of 5-HT1B receptor agonist CP-93,129 into the dorsal raphé nucleus. a shows the effect of CP-93,129 on the mean (±SEM, vertical lines), frequency of attack bites after the consumption of water (+H2O, open circles and bar), or 1.0 g/kg alcohol (+1.0 Alc, filled circles and bar). b shows the effects of CP-93,129 on the duration of walking after the consumption of water (+H2O, open circles and bar) or 1.0 g/kg alcohol (+1.0 Alc, filled circles and bar). N = 7; asterisks denote significant decrease from aCSF and p < 0.05 for all comparisons
Multiple indices of motor activity were dose-dependently reduced by intra-raphé administration of the 5-HT1B agonist, CP-93,129 (Table 1). Specifically, there was a significant main effect of drug on the duration of walking (F(3,24) = 10.6, p < 0.001; Fig. 2; Table 1) and rearing (F(3,24) = 4.8, p = 0.009; Table 1). The frequencies of walking (F(3,24) = 29.4, p < 0.001), rearing (F(3,24) = 5.0, p = 0.008), and self-grooming (F(3,24) = 4.0, p = 0.019) were also dose-dependently reduced by drug treatment (Fig. 2, Table 1). Post-hoc tests revealed that infusion of the highest dose of CP-93,129 (1.0 μg) significantly decreased frequencies of all of these motor behaviors and walking frequency was also significantly affected by a moderate dose of CP-93,129 (0.5 μg). Social contact with the intruder is rarely observed and was not changed by any of the drug treatments. Consumption of 1.0 g/kg alcohol did not affect any of these aggressive and non-aggressive behaviors.
Experiment 2: intra-cortical microinjection and aggression
Intra-cortical administration of CP-93,129 produced a significant decrease in aggressive behavior after consumption of 1.0 g/kg alcohol or water. Specifically, a significant main effect of drug was found on the frequency of attack bites (F(3,18) = 3.2, p = 0.049; Fig. 3, Table 2) and on the frequency (F(3,18) = 3.9, p = 0.026; Table 2) and duration of sideways threats (F(3,18) = 3.7, p = 0.026; Table 2;). Post-hoc tests revealed that microinjection of 0.1 and 1.0 μg CP-93,129 significantly decreased the frequency of attack bites and that microinjection of 0.1 and 0.5 μg CP-93,129 significantly reduced the frequency and duration of sideways threats. Tail rattle frequencies (F(1,18) = 8.5, p = 0.027; Table 2) and durations (F(1,18) = 7.7, p = 0.033; Table 2) were significantly suppressed after self-administration of 1.0 g/kg alcohol, but not water, irrespective of CP-93,129 dose. Aggressive pursuit of the intruder rarely occurred and was unaffected by any treatment condition.

Microinjection of the 5-HT1B receptor agonist CP-93,129 into the infralimbic region of the prefrontal cortex. a shows the effect of CP-93,129 on the mean (±SEM, vertical lines) frequency of attack bites after the consumption of water (+H2O, open circles and bar) or 1.0 g/kg alcohol (+1.0 Alc, filled circles and bar). b shows the effects of CP-93,129 on the duration of walking after the consumption of water (+H2O, open circles and bar) or 1.0 g/kg alcohol (+1.0 Alc, filled circles and bar). N = 9; asterisks denote significant decrease from aCSF and p < 0.05 for all comparisons
The predominant motor activity behaviors, walking, rearing, and contact with the intruder, were unaffected by alcohol or CP-93,129 (Fig. 3, Table 2 ). However, the duration of self-grooming was significantly decreased after microinjection of 1.0 μg of CP-93,129 (F(3,18) = 3.3, p = 0.046; Table 2) and the frequency of grooming was significantly reduced after microinjection of 0.1 and 1.0 μg of CP-93,129 (F(3,18) = 7.0, p = 0.002; Table 2).
Discussion
The relationship between serotonin and aggressive behavior has been studied for several decades and our understanding of how serotonin acts continues to be refined. Of the many 5-HT receptor subtypes, the 5-HT1B receptor is especially interesting because systemic administration of 5-HT1B receptor agonists has been found to decrease aggressive behavior without non-selective effects on motor behaviors (for review, see Olivier and van Oorschot 2005). The current studies extend these previous studies to brain regions that contain 5-HT1B receptors and that are thought to be necessary for aggressive behavior, the infralimbic cortex and the dorsal raphé nucleus. Infusion of the 5-HT1B receptor agonist CP-93,129 into the infralimbic cortex and the dorsal raphé nucleus similarly decreased aggression that occurred after consumption of both water and 1.0 g/kg alcohol. Nonetheless, the behavioral specificity of the anti-aggressive effect of CP-93,129 differed between brain regions. When injected into the dorsal raphé, CP-93,129 decreased both aggressive and motor behavior; on the other hand, infralimbic injection decreased aggression without affecting motor behavior. Furthermore, CP-93,129 was more efficacious at reducing aggressive behavior when infused into the ILC—the lowest tested dose (0.1 μg CP-93,129) significantly reduced aggression after intra-ILC but not intra-DRN microinjection. This dissociation is consistent with the literature indicating blunted serotonergic tone in the PFC of highly aggressive mice and suggests a behaviorally specific role for the infralimbic region of the prefrontal cortex in the modulation of aggression, whereas the dorsal raphé may control both aggressive and motor behaviors (Caramaschi et al. 2008). These studies further elucidate the importance of the serotonergic system in the modulation of aggression and highlight the infralimbic cortex as a critical region of interest.
The neural circuitry of aggressive behavior is becoming more widely understood and developed. Early studies correlated levels of activation or immediate early gene expression with a history of violent and/or aggressive behavior and found that the amygdala, prefrontal cortex, and hypothalamus are key regions that modulate and are activated in association with aggressive behavior (Caramaschi et al. 2008; Halasz et al. 2006; Haller et al. 2006; Kollack-Walker and Newman 1995; Lin et al. 2011; Veening et al. 2005). Recently, a distinction has emerged between cortical subregions that are activated in response to social interaction versus aggressive confrontations. Specifically, the infralimbic and medial orbital cortices show greater c-fos immunoreactivity 1 h after an aggressive, but not social, encounter. The ventral and lateral orbital cortices were only activated by social encounters demonstrating a specific role of the ILC in the modulation of aggressive behavior (Halasz et al. 2006). The ILC contains the 5-HT1B receptors that are hypothesized to be important for 5-HT’s anti-aggressive effects. In our studies, these receptors were targeted by microinjecting the selective 5-HT1B receptor agonist, CP-93,129. Administration of this agonist reduced aggressive behaviors without affecting motor behaviors such as walking, rearing, and grooming. The behaviorally specific reduction in aggression confirms results from other microinjection studies showing that 5-HT1B activation in the ventral orbitofrontal cortex selectively reduces species-typical, maternal, and aggression escalated by social instigation (Centenaro et al. 2008; da Veiga et al. 2011; De Almeida et al. 2006). However, they oppose the results of microinjection of a different 5-HT1B receptor agonist, CP-94,253, that was shown to increase alcohol-heightened aggression and decrease extracellular 5-HT (Faccidomo et al. 2008). Nonetheless, these studies provide evidence that cortical 5-HT1B receptors are essential for aggressive behavior.
The medial prefrontal cortex is richly innervated with 5-HT projections originating from DRN neurons (O’Hearn and Molliver 1984). 5-HT1B receptors are inhibitory G-protein coupled receptors and function both as pre-synaptic autoreceptors in the PFC located at the terminals of serotonergic neurons, and as heteroreceptors on non-serotonergic interneurons (Bruinvels et al. 1993, 1994; Hoyer et al. 1985; Sari et al. 1999). Recent evidence suggests that this population of receptors might be vulnerable to alcohol-induced functional adaptations. Specifically, Chiavegatto et al. 2010 found that prefrontal 5-HT1B mRNA levels were significantly reduced in mice who repeatedly engaged in alcohol-heightened aggression vs. those mice whose aggressive behavior did not escalate after alcohol consumption. This result suggests that there is a functional difference between alcohol-heightened and non-heightened aggressors in their serotonergic system, which is inversely correlated with a heightened aggressive response to acute alcohol. Our prior study on CP-94,253 found that extracellular 5-HT levels following reverse perfusion in the PFC were blunted in mice with a history of alcohol consumption (Faccidomo et al. 2008). Local infusion of CP-93,129 into the PFC has been shown to significantly reduce levels of extracellular glutamate suggesting that 5-HT1B receptors are likely located on glutamatergic neurons in the PFC (Golembiowska and Dziubina 2002). These glutamate neurons may in turn project to the subcortical structures involved in the expression of aggression and similar impulsive-like behaviors. Together, these data suggest a neurochemical mechanism by which 5-HT in the PFC may regulate aggressive behaviors.
The dorsal raphé is also a site of action for the 5-HT1B receptor agonists. Microinjection of CP-93,129 decreased both species-typical and aggression after alcohol self-administration, but also disrupted motor behavior. This non-specific decrease in aggression is consistent with previous findings of dorsal raphé microinjection of 5-HT1A or 5-HT1B receptor agonists (Bannai et al. 2007; De Almeida and Lucion 1997; Faccidomo et al. 2008; Mos et al. 1993) and may be the result of slowing of 5-HT neuronal activity. Functionally, local administration of 5-HT1B receptor agonists generally decreases DRN and MRN cell firing, though regulation of 5-HT cell firing and release is chiefly mediated by somatodendritic 5-HT1A receptors (Adell et al. 2001; Evrard et al. 1999; Sprouse 1991; Sprouse and Aghajanian 1987; 1988; Verge et al. 1985). Several key findings argue against an essential and specific role for dorsal raphé 5-HT1B receptors in the anti-aggressive effects of 5-HT1B receptor agonists. First, systemic administration of anti-aggressive 5-HT1B/D agonists does not decrease motor activity; the behavioral effects are more similar to cortical microinjection (de Almeida et al. 2001; Fish et al. 1999). Second, the anti-aggressive effects do not appear to depend on somatodendritic 5-HT1B receptors because neurotoxic lesions of dorsal raphé 5-HT cell bodies using 5,7-dihydroxytryptamine do not prevent the anti-aggressive effects of systemically administered 5-HT1B/D agonists (de Almeida et al. 2001; Sijbesma et al. 1991). Although the dorsal raphé does not appear to be essential for the anti-aggressive effects of 5-HT1B receptor agonists, the dorsal raphé does appear to be essential for the actions of other neuromodulators. Intra raphé GABAA, CRF1, and GABAB receptor manipulations regulate alcohol-heightened and species-typical aggression (Takahashi et al. 2010a, b, 2011) and GABAergic interneurons regulate extracellular release of 5-HT in the prefrontal cortex (Takahashi et al. 2010b). Thus, GABA serves to functionally regulate serotonergic tone throughout this critical pathway which indirectly modulates of aggressive behavior.
In summary, these findings confirm and further demonstrate the importance of prefrontal cortex 5-HT1B receptors in the modulation of species-typical levels of aggression. Future studies aim to expand these findings to address whether these 5-HT1B-mediated effects on aggression can be mimicked by intra-cortical glutamatergic antagonism. Furthermore, given the ability of alcohol to down regulate 5-HT1B receptors (Chiavegatto et al. 2010) and the demonstration that 5-HT1B overexpression in the nucleus accumbens is linked to excessive alcohol self-administration (Furay et al. 2011; Hoplight et al. 2006), a key direction for future studies would be to investigate the neural circuitry that regulates 5-HT1B receptor activity, escalated aggressive behavior and excessive alcohol intake.
References
Adell A, Celada P, Artigas F (2001) The role of 5-HT1B receptors in the regulation of serotonin cell firing and release in the rat brain. J Neurochem 79:172–182
Ahlenius S, Larsson K (1998) Evidence for an involvement of 5-HT1B receptors in the inhibition of male rat ejaculatory behavior produced by 5-HTP. Psychopharmacol (Berl) 137:374–382
Bannai M, Fish EW, Faccidomo S, Miczek KA (2007) Anti-aggressive effects of agonists at 5-HT1B receptors in the dorsal raphe nucleus of mice. Psychopharmacol (Berl) 193:295–304
Barnes NM, Sharp T (1999) A review of central 5-HT receptors and their function. Neuropharmacology 38:1083–1152
Best M, Williams JM, Coccaro EF (2002) Evidence for a dysfunctional prefrontal circuit in patients with an impulsive aggressive disorder. Proc Natl Acad Sci U S A 99:8448–8453
Bouwknecht JA, Hijzen TH, van der Gugten J, Maes RA, Hen R, Olivier B (2001) Absence of 5-HT(1B) receptors is associated with impaired impulse control in male 5-HT(1B) knockout mice. Biol Psychiatry 49:557–568
Brown GL, Goodwin FK, Ballenger JC, Goyer PF, Major LF (1979) Aggression in humans correlates with cerebrospinal fluid amine metabolites. Psychiatry Res 1:131–139
Bruinvels AT, Palacios JM, Hoyer D (1993) Autoradiographic characterisation and localisation of 5-HT1D compared to 5-HT1B binding sites in rat brain. Naunyn Schmiedebergs Arch Pharmacol 347:569–582
Bruinvels AT, Landwehrmeyer B, Gustafson EL, Durkin MM, Mengod G, Branchek TA, Hoyer D, Palacios JM (1994) Localization of 5-HT1B, 5-HT1D alpha, 5-HT1E and 5-HT1F receptor messenger RNA in rodent and primate brain. Neuropharmacology 33:367–386
Caramaschi D, de Boer SF, de Vries H, Koolhaas JM (2008) Development of violence in mice through repeated victory along with changes in prefrontal cortex neurochemistry. Behav Brain Res 189:263–272
Centenaro LA, Vieira K, Zimmermann N, Miczek KA, Lucion AB, de Almeida RM (2008) Social instigation and aggressive behavior in mice: role of 5-HT1A and 5-HT1B receptors in the prefrontal cortex. Psychopharmacol (Berl) 201:237–248
Chiavegatto S, Quadros IM, Ambar G, Miczek KA (2010) Individual vulnerability to escalated aggressive behavior by a low dose of alcohol: decreased serotonin receptor mRNA in the prefrontal cortex of male mice. Genes Brain Behav 9:110–119
Clark MS, Neumaier JF (2001) The 5-HT1B receptor: behavioral implications. Psychopharmacol Bull 35:170–185
Clark MS, Sexton TJ, McClain M, Root D, Kohen R, Neumaier JF (2002) Overexpression of 5-HT1B receptor in dorsal raphe nucleus using Herpes Simplex Virus gene transfer increases anxiety behavior after inescapable stress. J Neurosci 22:4550–4562
Clark MS, Vincow ES, Sexton TJ, Neumaier JF (2004) Increased expression of 5-HT1B receptor in dorsal raphe nucleus decreases fear-potentiated startle in a stress dependent manner. Brain Res 1007:86–97
Cloninger CR (1987) Neurogenetic adaptive mechanisms in alcoholism. Science 236:410–416
da Veiga CP, Miczek KA, Lucion AB, de Almeida RM (2011) Social instigation and aggression in postpartum female rats: role of 5-Ht1A and 5-Ht1B receptors in the dorsal raphe nucleus and prefrontal cortex. Psychopharmacol (Berl) 213:475–487
Davidson RJ, Jackson DC, Kalin NH (2000) Emotion, plasticity, context, and regulation: perspectives from affective neuroscience. Psychol Bull 126:890–909
De Almeida RM, Lucion AB (1997) 8-OH-DPAT in the median raphe, dorsal periaqueductal gray and corticomedial amygdala nucleus decreases, but in the medial septal area it can increase maternal aggressive behavior in rats. Psychopharmacol (Berl) 134:392–400
de Almeida RM, Nikulina EM, Faccidomo S, Fish EW, Miczek KA (2001) Zolmitriptan—a 5-HT1B/D agonist, alcohol, and aggression in mice. Psychopharmacol (Berl) 157:131–141
De Almeida RM, Rosa MM, Santos DM, Saft DM, Benini Q, Miczek KA (2006) 5-HT(1B) receptors, ventral orbitofrontal cortex, and aggressive behavior in mice. Psychopharmacol (Berl) 185:441–450
Evrard A, Laporte AM, Chastanet M, Hen R, Hamon M, Adrien J (1999) 5-HT1A and 5-HT1B receptors control the firing of serotoninergic neurons in the dorsal raphe nucleus of the mouse: studies in 5-HT1B knock-out mice. Eur J Neurosci 11:3823–3831
Faccidomo S, Bannai M, Miczek KA (2008) Escalated aggression after alcohol drinking in male mice: dorsal raphe and prefrontal cortex serotonin and 5-HT(1B) receptors. Neuropsychopharmacology 33:2888–2899
Fairbanks LA, Melega WP, Jorgensen MJ, Kaplan JR, McGuire MT (2001) Social impulsivity inversely associated with CSF 5-HIAA and fluoxetine exposure in vervet monkeys. Neuropsychopharmacology 24:370–378
Fehr C, Grintschuk N, Szegedi A, Anghelescu I, Klawe C, Singer P, Hiemke C, Dahmen N (2000) The HTR1B 861 G>C receptor polymorphism among patients suffering from alcoholism, major depression, anxiety disorders and narcolepsy. Psychiatry Res 97:1–10
Ferrari PF, Palanza P, Parmigiani S, de Almeida RM, Miczek KA (2005) Serotonin and aggressive behavior in rodents and nonhuman primates: predispositions and plasticity. Eur J Pharmacol 526:259–273
Fineberg NA, Potenza MN, Chamberlain SR, Berlin HA, Menzies L, Bechara A, Sahakian BJ, Robbins TW, Bullmore ET, Hollander E (2010) Probing compulsive and impulsive behaviors, from animal models to endophenotypes: a narrative review. Neuropsychopharmacology 35:591–604
Fish EW, Faccidomo S, Miczek KA (1999) Aggression heightened by alcohol or social instigation in mice: reduction by the 5-HT(1B) receptor agonist CP-94,253. Psychopharmacol (Berl) 146:391–399
Fish EW, McKenzie-Quirk SD, Bannai M, Miczek KA (2008) 5-HT(1B) receptor inhibition of alcohol-heightened aggression in mice: comparison to drinking and running. Psychopharmacol (Berl) 197:145–156
Frankle WG, Lombardo I, New AS, Goodman M, Talbot PS, Huang Y, Hwang DR, Slifstein M, Curry S, Abi-Dargham A, Laruelle M, Siever LJ (2005) Brain serotonin transporter distribution in subjects with impulsive aggressivity: a positron emission study with [11C]McN 5652. Am J Psychiatry 162:915–923
Franklin KBJ, Paxinos G (2001) The mouse brain in sterotaxic coordinates, 2nd edn. Academic, New York
Furay AR, Neumaier JF, Mullenix AT, Kaiyala KK, Sandygren NK, Hoplight BJ (2011) Overexpression of 5-HT(1B) mRNA in nucleus accumbens shell projection neurons differentially affects microarchitecture of initiation and maintenance of ethanol consumption. Alcohol 45:19–32
Giacalone E, Tansella M, Valzelli L, Garattini S (1968) Brain serotonin metabolism in isolated aggressive mice. Biochem Pharmacol 17:1315–1327
Giancola PR (2000) Executive functioning: a conceptual framework for alcohol-related aggression. Exp Clin Psychopharmacol 8:576–597
Golembiowska K, Dziubina A (2002) Inhibition of amino acid release by 5-HT1B receptor agonist in the rat prefrontal cortex. Pol J Pharmacol 54:625–631
Gowin JL, Swann AC, Moeller FG, Lane SD (2010) Zolmitriptan and human aggression: interaction with alcohol. Psychopharmacol (Berl) 210:521–531
Grant E, Macintosh JH (1963) A comparison of the social postures of some common laboratory rodents. Behaviour 21:246–295
Halasz J, Toth M, Kallo I, Liposits Z, Haller J (2006) The activation of prefrontal cortical neurons in aggression—a double labeling study. Behav Brain Res 175:166–175
Haller J, Toth M, Halasz J, De Boer SF (2006) Patterns of violent aggression-induced brain c-fos expression in male mice selected for aggressiveness. Physiol Behav 88:173–182
Higley JD, Linnoila M (1997) A nonhuman primate model of excessive alcohol intake. Personality and neurobiological parallels of type I- and type II-like alcoholism. Recent Dev Alcohol 13:191–219
Higley JD, Suomi SJ, Linnoila M (1996a) A nonhuman primate model of type II alcoholism? Part 2. Diminished social competence and excessive aggression correlates with low cerebrospinal fluid 5-hydroxyindoleacetic acid concentrations. Alcohol Clin Exp Res 20:643–650
Higley JD, Suomi SJ, Linnoila M (1996b) A nonhuman primate model of type II excessive alcohol consumption? Part 1. Low cerebrospinal fluid 5-hydroxyindoleacetic acid concentrations and diminished social competence correlate with excessive alcohol consumption. Alcohol Clin Exp Res 20:629–642
Holmes A, Murphy DL, Crawley JN (2002) Reduced aggression in mice lacking the serotonin transporter. Psychopharmacol (Berl) 161:160–167
Hoplight BJ, Sandygren NA, Neumaier JF (2006) Increased expression of 5-HT1B receptors in rat nucleus accumbens via virally mediated gene transfer increases voluntary alcohol consumption. Alcohol 38:73–79
Hoyer D, Engel G, Kalkman HO (1985) Characterization of the 5-HT1B recognition site in rat brain: binding studies with (-)[125I]iodocyanopindolol. Eur J Pharmacol 118:1–12
Kable JW, Glimcher PW (2007) The neural correlates of subjective value during intertemporal choice. Nat Neurosci 10:1625–1633
Koe BK, Lebel LA, Fox CB, Macor JE (1992a) Binding and uptake studies with [H-3] Cp-93,129, a radiolabeled selective 5-Ht1B receptor ligand. Drug Dev Res 25:67–74
Koe BK, Nielsen JA, Macor JE, Heym J (1992b) Biochemical and behavioral-studies of the 5-Ht(1B) receptor agonist, Cp-94,253. Drug Dev Res 26:241–250
Kollack-Walker S, Newman SW (1995) Mating and agonistic behavior produce different patterns of Fos immunolabeling in the male Syrian hamster brain. Neuroscience 66:721–736
Lappalainen J, Long JC, Eggert M, Ozaki N, Robin RW, Brown GL, Naukkarinen H, Virkkunen M, Linnoila M, Goldman D (1998) Linkage of antisocial alcoholism to the serotonin 5-HT1B receptor gene in 2 populations. Arch Gen Psychiatry 55:989–994
Lee MD, Simansky KJ (1997) CP-94, 253: a selective serotonin1B (5-HT1B) agonist that promotes satiety. Psychopharmacol (Berl) 131:264–270
Lin D, Boyle MP, Dollar P, Lee H, Lein ES, Perona P, Anderson DJ (2011) Functional identification of an aggression locus in the mouse hypothalamus. Nature 470:221–226
Linnoila M, Virkkunen M, Scheinin M, Nuutila A, Rimon R, Goodwin FK (1983) Low cerebrospinal fluid 5-hydroxyindoleacetic acid concentration differentiates impulsive from nonimpulsive violent behavior. Life Sci 33:2609–2614
Lister RG, Hilakivi LA (1988) The effects of novelty, isolation, light and ethanol on the social behavior of mice. Psychopharmacol (Berl) 96:181–187
Mehlman PT, Higley JD, Faucher I, Lilly AA, Taub DM, Vickers J, Suomi SJ, Linnoila M (1994) Low CSF 5-HIAA concentrations and severe aggression and impaired impulse control in nonhuman primates. Am J Psychiatry 151:1485–1491
Miczek KA, de Almeida RM (2001) Oral drug self-administration in the home cage of mice: alcohol-heightened aggression and inhibition by the 5-HT1B agonist anpirtoline. Psychopharmacol (Berl) 157:421–429
Miczek KA, O’Donnell JM (1978) Intruder-evoked aggression in isolated and nonisolated mice: effects of psychomotor stimulants and L-dopa. Psychopharmacol (Berl) 57:47–55
Miczek KA, O’Donnell JM (1980) Alcohol and chlordiazepoxide increase suppressed aggression in mice. Psychopharmacol (Berl) 69:39–44
Miczek KA, Weerts EM, Tornatzky W, DeBold JF, Vatne TM (1992) Alcohol and “bursts” of aggressive behavior: ethological analysis of individual differences in rats. Psychopharmacol (Berl) 107:551–563
Miczek KA, Weerts EM, DeBold JF (1993) Alcohol, benzodiazepine-GABAA receptor complex and aggression: ethological analysis of individual differences in rodents and primates. J Stud Alcohol Suppl 11:170–179
Miczek KA, Barros HM, Sakoda L, Weerts EM (1998) Alcohol and heightened aggression in individual mice. Alcohol Clin Exp Res 22:1698–1705
Moret C, Briley M (2000) The possible role of 5-HT(1B/D) receptors in psychiatric disorders and their potential as a target for therapy. Eur J Pharmacol 404:1–12
Mos J, Olivier B, Poth M, Van Oorschot R, Van Aken H (1993) The effects of dorsal raphe administration of eltoprazine, TFMPP and 8-OH-DPAT on resident intruder aggression in the rat. Eur J Pharmacol 238:411–415
National Research Council (1996) Guide for the care and use of laboratory animals. National Academy Press, New York
New AS, Gelernter J, Goodman M, Mitropoulou V, Koenigsberg H, Silverman J, Siever LJ (2001) Suicide, impulsive aggression, and HTR1B genotype. Biol Psychiatry 50:62–65
O’Hearn E, Molliver ME (1984) Organization of raphe-cortical projections in rat: a quantitative retrograde study. Brain Res Bull 13:709–726
Olivier B, van Oorschot R (2005) 5-HT1B receptors and aggression: a review. Eur J Pharmacol 526:207–217
Parsons LH, Weiss F, Koob GF (1998) Serotonin1B receptor stimulation enhances cocaine reinforcement. J Neurosci 18:10078–10089
Raine A, Buchsbaum MS, Stanley J, Lottenberg S, Abel L, Stoddard J (1994) Selective reductions in prefrontal glucose metabolism in murderers. Biol Psychiatry 36:365–373
Reif A, Rosler M, Freitag CM, Schneider M, Eujen A, Kissling C, Wenzler D, Jacob CP, Retz-Junginger P, Thome J, Lesch KP, Retz W (2007) Nature and nurture predispose to violent behavior: serotonergic genes and adverse childhood environment. Neuropsychopharmacology 32:2375–2383
Samson HH (1986) Initiation of ethanol reinforcement using a sucrose-substitution procedure in food- and water-sated rats. Alcohol Clin Exp Res 10:436–442
Sari Y, Miquel MC, Brisorgueil MJ, Ruiz G, Doucet E, Hamon M, Verge D (1999) Cellular and subcellular localization of 5-hydroxytryptamine1B receptors in the rat central nervous system: immunocytochemical, autoradiographic and lesion studies. Neuroscience 88:899–915
Sijbesma H, Schipper J, de Kloet ER, Mos J, van Aken H, Olivier B (1991) Postsynaptic 5-HT1 receptors and offensive aggression in rats: a combined behavioural and autoradiographic study with eltoprazine. Pharmacol Biochem Behav 38:447–458
Sinha R, Cloninger CR, Parsian A (2003) Linkage disequilibrium and haplotype analysis between serotonin receptor 1B gene variations and subtypes of alcoholism. Am J Med Genet B Neuropsychiatr Genet 121B:83–88
Sprouse JS (1991) Inhibition of dorsal raphe cell firing by MDL 73005EF, a novel 5-HT1A receptor ligand. Eur J Pharmacol 201:163–169
Sprouse JS, Aghajanian GK (1987) Electrophysiological responses of serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists. Synapse 1:3–9
Sprouse JS, Aghajanian GK (1988) Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: a comparative study with dorsal raphe neurons. Neuropharmacology 27:707–715
Sripada CS, Gonzalez R, Phan KL, Liberzon I (2010) The neural correlates of intertemporal decision-making: contributions of subjective value, stimulus type, and trait impulsivity. Hum Brain Mapp 32:1637–1648
Stern L, Zohar J, Cohen R, Sasson Y (1998) Treatment of severe, drug resistant obsessive compulsive disorder with the 5HT1D agonist sumatriptan. Eur Neuropsychopharmacol 8:325–328
Takahashi A, Kwa C, Debold JF, Miczek KA (2010a) GABA(A) receptors in the dorsal raphe nucleus of mice: escalation of aggression after alcohol consumption. Psychopharmacol (Berl) 211:467–477
Takahashi A, Shimamoto A, Boyson CO, DeBold JF, Miczek KA (2010b) GABA(B) receptor modulation of serotonin neurons in the dorsal raphe nucleus and escalation of aggression in mice. J Neurosci 30:11771–11780
Takahashi A, Quadros IM, de Almeida RM, Miczek KA (2011) Brain serotonin receptors and transporters: initiation vs. termination of escalated aggression. Psychopharmacol (Berl) 213:183–212
Van der Vegt BJ, Lieuwes N, Cremers TI, deBoer SF, Koolhaas JM (2003) Cerebrospinal fluid monoamine and metabolite concentrations and aggression in rats. Horm Behav 44:199–208
van Erp AM, Miczek KA (2000) Aggressive behavior, increased accumbal dopamine, and decreased cortical serotonin in rats. J Neurosci 20:9320–9325
Veening JG, Coolen LM, de Jong TR, Joosten HW, de Boer SF, Koolhaas JM, Olivier B (2005) Do similar neural systems subserve aggressive and sexual behaviour in male rats? Insights from c-Fos and pharmacological studies. Eur J Pharmacol 526:226–239
Verge D, Daval G, Patey A, Gozlan H, el Mestikawy S, Hamon M (1985) Presynaptic 5-HT autoreceptors on serotonergic cell bodies and/or dendrites but not terminals are of the 5-HT1A subtype. Eur J Pharmacol 113:463–464
Virkkunen M, Linnoila M (1993) Brain serotonin, type II alcoholism and impulsive violence. J Stud Alcohol Suppl 11:163–169
Virkkunen M, Rawlings R, Tokola R, Poland RE, Guidotti A, Nemeroff C, Bissette G, Kalogeras K, Karonen SL, Linnoila M (1994) CSF biochemistries, glucose metabolism, and diurnal activity rhythms in alcoholic, violent offenders, fire setters, and healthy volunteers. Arch Gen Psychiatry 51:20–27
Virkkunen M, Eggert M, Rawlings R, Linnoila M (1996) A prospective follow-up study of alcoholic violent offenders and fire setters. Arch Gen Psychiatry 53:523–529
Volkow ND, Wang GJ, Hitzemann R, Fowler JS, Overall JE, Burr G, Wolf AP (1994) Recovery of brain glucose metabolism in detoxified alcoholics. Am J Psychiatry 151:178–183
Weerts EM, Tornatzky W, Miczek KA (1993) Prevention of the pro-aggressive effects of alcohol in rats and squirrel monkeys by benzodiazepine receptor antagonists. Psychopharmacol (Berl) 111:144–152
Winstanley CA, Theobald DE, Dalley JW, Cardinal RN, Robbins TW (2006) Double dissociation between serotonergic and dopaminergic modulation of medial prefrontal and orbitofrontal cortex during a test of impulsive choice. Cereb Cortex 16:106–114
Yang Y, Raine A (2009) Prefrontal structural and functional brain imaging findings in antisocial, violent, and psychopathic individuals: a meta-analysis. Psychiatry Res 174:81–88
Zhuang X, Gross C, Santarelli L, Compan V, Trillat AC, Hen R (1999) Altered emotional states in knockout mice lacking 5-HT1A or 5-HT1B receptors. Neuropsychopharmacology 21:52S–60S
Acknowledgments
The authors would like to thank J Thomas Sopko for his outstanding technical assistance. All research was supported by AA13983 (KAM)
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Faccidomo, S., Quadros, I.M.H., Takahashi, A. et al. Infralimbic and dorsal raphé microinjection of the 5-HT1B receptor agonist CP-93,129: attenuation of aggressive behavior in CFW male mice. Psychopharmacology 222, 117–128 (2012). https://doi.org/10.1007/s00213-011-2629-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-011-2629-1