
Abstract
Microbial product lipopolysaccharide has been shown to be involved in the pathogenesis of inflammatory skin diseases. Parthenolide present in extracts of the herb feverfew has demonstrated an anti-inflammatory effect. However, the effect of parthenolide on the Akt/mTOR and NF-κB pathway activation-induced productions of inflammatory mediators in keratinocytes has not been studied. Using human keratinocytes, we investigated the effect of parthenolide on the inflammatory mediator production in relation to the Toll-like receptor-4-mediated-Akt/mTOR and NF-κB pathways, which regulate the transcription genes involved in immune and inflammatory responses. Parthenolide, Akt inhibitor, Bay 11–7085, and N-acetylcysteine each attenuated the lipopolysaccharide-induced production of IL-1β and PGE2, increase in the levels of cyclooxygenase, formation of reactive oxygen species, increase in the levels of Toll-like receptor-4, and activation of the Akt/mTOR and NF-κB in keratinocytes. The results show that parthenolide appears to attenuate the lipopolysaccharide-stimulated production of inflammatory mediators in keratinocytes by suppressing the Toll-like receptor-4-mediated activation of the Akt, mTOR, and NF-κB pathways. The activation of signaling transduction pathways appear to be regulated by reactive oxygen species. Parthenolide appears to attenuate the microbial product-mediated inflammatory skin diseases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Keratinocytes are considered to be implicated in the inflammatory and immune responses in skin through the production of proinflammatory cytokines and chemokines (Pastore et al. 2006; Tsuruta 2009). Keratinocytes respond to microbial products, such as lipopolysaccharide, through the activation of Toll-like receptors, thereby producing various cytokines (Baker 2006; Begon et al. 2007; Kang et al. 2006). Lipopolysaccharide induces immune and inflammatory responses by expressing Toll-like receptors (Takeda and Akira 2005; Thomson et al. 2009). Toll-like receptor 4, one of the Toll-like receptors, plays a critical role in recognizing microbial lipopolysaccharide (Takeda and Akira 2005). Lipopolysaccharide binding to keratinocyte CD14 and subsequent activation of Toll-like receptor 4 causes an activation of nuclear factor (NF)-κB and the secretion of proinflammatory cytokines and chemokines (Song et al. 2002; Takeda and Akira 2005). Lipopolysaccharide induces the activation of the phosphatidylinositol (PI) 3-kinase/Akt (protein kinase B)/mammalian target of rapamycin (mTOR) signaling pathways, which is followed by activation of transcription factors, including activator protein-1 and NF-κB (Thomson et al. 2009; Zhong et al. 2012). NF-κB regulates genes responsible for the innate and adaptive immune responses as well as inflammation (Ghosh and Hayden 2008). NF-κB activation is triggered by a variety of agents, including cytokine TNF-α, oxidative stress, and DNA damage (Ghosh and Hayden 2008; Gloire et al. 2006; Siomek 2012). Reactive oxygen species play a critical role in the physiological regulation of cellular functions and are involved in pathologic conditions, such as inflammation and cell death (Marchi et al. 2012). They have been shown to induce the activation of NF-κB (Gloire et al. 2006; Siomek 2012).
Parthenolide is a major sesquiterpene lactone present in extracts of the herb feverfew Tanacetum parthenium and is found in highest concentration in the flowers and fruit (Kwok et al. 2001; Nam 2006; Yuan et al. 2006). Parthenolide has been shown to exhibit anti-inflammatory effects by suppressing NF-κB activation (Juliana et al. 2010; Kwok et al. 20001; Magni et al. 2012; Nam 2006). Parthenolide blocks the hyperalgesic response and attenuates the edema response in the rat carrageenan inflammation model (Feltenstein et al. 2004). Parthenolide inhibits the activity of multiple inflammasomes, which process the inactive pro-IL-1β and pro-IL-18 into the active proinflammatory cytokines IL-1β and IL-18, in macrophages by directly inhibiting the protease activity of caspase-1 (Juliana et al. 2010). Parthenolide suppresses the IL-4 protein levels in anti-CD3/CD28-stimulated peripheral blood T cells from allergic and normal donors (Li-Weber et al. 2002). Unlike this report, in the spleens of mice treated with lipopolysaccharide, the induction of IL-6 messenger RNA (mRNA) only decreased, while mRNAs of TNF-alpha and COX-2 are not changed, and IL-1beta mRNA increased (Smolinski and Pestka 2005).
Parthenolide is demonstrated to have an anti-inflammatory effect. Keratinocytes are known to be involved in the skin inflammatory and immune responses. However, the effect of parthenolide on the Akt/mTOR and NF-κB pathway activation-induced productions of inflammatory mediators in keratinocytes has not been studied. Using human keratinocytes, we investigated the effect of parthenolide on lipopolysaccharide-induced inflammatory mediator production in relation to the Toll-like receptor 4 expression-mediated activations of the Akt, mTOR, and NF-κB pathways. Then, we assessed the effect and action of parthenolide as a preventative compound to treat microbial product-mediated inflammatory skin diseases.
Materials and methods
Materials
Parthenolide, rapamycin, Bay 11–7085 ((2E)-3-[[4-(1,1-dimethylethyl)phenyl]sulfonyl]-2-propenenitrile), Akt inhibitor (type II, SH-5) and horseradish peroxidase-conjugated anti-mouse IgG were purchased from EMD-Calbiochem. Co. (La Jolla, CA, USA). Enzyme-linked immunosorbent assay (ELISA) kits for human CXCL1/IL1β, human cyclooxygenase-2, human/mouse/rat phospho-Akt (Pan), and a parameter assay kit for prostaglandin E2 were purchased from R&D systems, Inc. (Minneapolis, MN, USA). Antibodies (Toll-like receptor (TLR)-4, NF-κB p65, NF-κB p50, phospho-IκB-α, phospho-Akt1, Akt1, mTOR, and β-actin) were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). The TransAMTM NF-κB assay kit was purchased from Active Motif® (Carlsbad, CA, USA). Lipopolysaccharide (from Escherichia coli), cell counting kit-8, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), N-acetylcysteine and other chemicals were purchased from Sigma-Aldrich Inc. (St. Louis, MO, USA).
Keratinocyte culture
Human keratinocytes (HEK001, tissue: skin; morphology: epithelial; cell type: human papillomavirus 16 E6/E7 transformed) were purchased from American Type Culture Collection (Manassas, VA, USA) and cultured in keratinocyte serum-free media supplemented with bovine pituitary extract, recombinant epidermal growth factor, 100 units/ml penicillin and 100 μg/ml streptomycin (GIBCO®, Invitrogen Co., Grand Island, NY, USA).
Immunoassays for IL-1β, PGE2, and cyclooxygenase-2
Keratinocytes (1 × 105 cells/300 μl for the cytokine assay or 4 x 104 cells/400 μl for the cyclooxygenase-2 assay) were grown in a 24-well plate and treated with 1 μg/ml lipopolysaccharide for 24 h. After centrifugation at 412g for 10 min, the amounts of IL-1β and PGE2 in the culture supernatants were analyzed using the ELISA and parameter kits, according to the manufacturer’s instructions. The cytosolic levels of cyclooxygenase-2 in cell lysates were analyzed using the ELISA kit, according to the manufacturer’s instruction. Absorbance was measured at 450 nm using a microplate reader (Magellan, TECAN, Salzburg, Austria).
Preparation of cytosolic and nuclear extracts
Keratinocytes (5 × 106 cells/ml) were treated with lipopolysaccharide for 30 min at 37 °C. Exceptionally, to assay the levels of TLR-4, keratinocytes were treated with lipoteichoic acid 24 h. Keratinocyte cytosolic and nuclear extracts were prepared, as a previously described (Schreiber et al. 1989). Keratinocytes were harvested by centrifugation at 412g for 10 min and washed twice with phosphate buffered saline (PBS). The cells were suspended in 400-μl lysis buffer (10 mM KCl, 1.5 mM MgCl2, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM dithiothreitol, 0.5 mM PMSF, 1 mM sodium orthovanadate, 2 μg/ml aprotinin, 2 μg/ml leupeptin, and 10 mM HEPES-KOH, pH 7.8) and were allowed to swell on ice for 15 min. Next, 25 μl of a 10 % Nonidet NP-40 solution (final concentration approximately 0.6 %) was added, and the tubes were vigorously vortexed for 10 s. The homogenates were centrifuged at 12,000g for 10 min at 4 °C. The supernatants were stored as cytoplasmic extracts and kept at −70 °C. The nuclear pellets were resuspended in 50 μl of an ice-cold hypertonic solution containing 5 % glycerol and 0.4 M NaCl lysis buffer. The tubes were incubated on ice for 30 min and then centrifuged at 12,000g for 15 min at 4 °C. The supernatants were collected as the nuclear extracts and stored at −70 °C. Protein concentrations were determined using the Bradford method according to the manufacturer’s instructions (Bio-Rad Laboratories, Hercules, CA, USA).
Western blot for phospho-IκB and NF-κB levels
The cytosolic fraction for the TLR-4 assay and the cytosolic and nuclear extracts for the NF-κB assay were mixed with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer and boiled for 5 min. Samples (30 μg protein/well) were loaded onto each lane of a 10–12 % SDS-polyacrylamide gel and transferred onto polyvinylidene difluoride membranes (GE Healthcare Chalfont St. Giles, Buckinghamshire, UK). Membranes were blocked for 2 h in Tris-buffered saline (TBS) (50 mM Tris–HCl, pH 7.5 and 150 mM NaCl) containing 0.1 % Tween 20 and 5 % non-fat dried milk. Each membrane was labeled with the appropriate antibody (TLR-4, NF-κB p65, NF-κB p50, phospho-IκB-α, Akt1, mTOR, or β-actin) overnight at 4 °C with gentle agitation. After four washes in TBS containing 0.1 % Tween 20, the membranes were incubated with horseradish peroxidase-conjugated anti-mouse IgG for 2 h at room temperature. Membranes were treated with SuperSignal® West Pico chemiluminescence substrate and protein bands were visualized by detecting the enhanced chemiluminescence in an appropriate image analyzer (Lite for Las-1000 plus version 1.1; Fuji Photo Film Co., Tokyo, Japan).
Assay for DNA binding activity of NF-κB
The biding of NF-κB p65 to DNA was determined according to the user’s manual for the TransAMTM NF-κB kit. Keratinocytes (2 × 106 cells/ml) were treated with lipopolysaccharide for 30 min. Nuclear extracts were prepared according to the procedure described in the Active Motif® protocol and added to a 96-well plate to which oligonucleotides containing an NF-κB consensus binding site (5ʹ-GGGACTTTCC-3ʹ) were immobilized. The active NF-κB p65 bound to DNA was exposed to a primary antibody specific for NF-κB p65, which was reacted with anti-rabbit horseradish peroxidase-conjugated IgG. Next, the color developer and stop solutions were added to the plate. The absorbance of samples was measured at 450 nm with a reference wavelength of 655 nm in a microplate reader.
Enzyme-linked immunosorbent assays for phosphorylated Akt
Keratinocytes (1 × 106 cells/ml) were treated with lipopolysaccharide for 4 h. Cells were harvested by centrifugation at 412g for 10 min, washed twice with PBS, and suspended in the lysis buffer provided by R&D systems for whole cell lysates. The homogenates were centrifuged at 2000g for 5 min and the supernatant was used for the ELISA. The amounts of phosphorylated Akt were determined according to the manufacturer’s instructions for the immunoassays. The supernatants were exposed first to the antibodies for the phosphorylated forms of the kinases, next to the biotinylated detection antibodies, and finally to streptavidin-horseradish-peroxidase. The absorbance was measured at 405 nm.
Measurement of intracellular reactive oxygen species formation
The dye DCFH2-DA, which is oxidized to fluorescent dichlorofluorescein (DCF) by hydroperoxides, was used to measure relative levels of cellular peroxides (Fu et al. 1998). Keratinocytes (1 × 105 cells/400 μl in 24-well plate) were treated with lipopolysaccharide 24 h at 37 °C. The cells were washed, suspended in fetal bovine serum-free RPMI-1640, incubated with 50 μM dye for 30 min at 37 °C and washed with phosphate buffered saline. The cell suspensions were centrifuged at 412g for 10 min and medium was removed. The cells were dissolved with 1 % Triton X-100 and fluorescence was measured at an excitation wavelength of 485 nm and an emission wavelength of 530 nm using a fluorescence microplate reader (SpectraFluor, TECAN, Salzburg, Austria).
Measurement of cell viability
Cell viability was measured using an MTT reduction assay, which is based on the conversion of MTT to formazan crystals by mitochondrial dehydrogenases (Mosmann 1983). Cells (3 × 104) were treated with parthenolide for 24 h at 37 °C. The medium (200 μl) was incubated with 10 μl of a 10 mg/ml MTT solution for 2 h at 37 °C. After centrifugation at 412g for 10 min, the culture medium was removed and 100 μl of dimethyl sulfoxide was added to each well to dissolve the formazan. The absorbance was measured at 570 nm using a microplate reader (Magellan, TECAN, Salzburg, Austria). Cell viability was expressed as a percentage of the value measured from the control cultures in which the cells did not receive any treatments.
Cell viability was also measured by monitoring the reduction of 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8) to a water-soluble formazan dye (Ishiyama et al. 1997). Cells (3 × 104) were treated with parthenolide for 24 h at 37 °C. The medium (200 μl) was incubated with 10 μl of cell counting kit-8 (Sigma-Aldrich Inc.) for 2 h at 37 °C. After incubation, the reaction was stopped by the addition of 1 % (wt/vol) sodium dodecylsulfate to each well. The absorbance was measured at 450 nm using a microplate reader with a reference wavelength at 650 nm.
Statistical analysis
The data are expressed as the mean ± SEM. Statistical analyses were performed by one-way analysis of variance (ANOVA). When significance was detected, the post hoc comparisons between the different groups were made using Duncan’s test for multiple comparisons. A probability value of p < 0.05 was considered statistically significant.
Results
The effect of parthenolide on the production and levels of inflammatory mediators
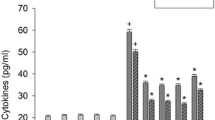
The effect of parthenolide on the production and changes in the levels of inflammatory mediators in keratinocytes exposed to lipopolysaccharide was examined. Keratinocytes treated with parthenolide exhibited a concentration-dependent reduction in lipopolysaccharide-induced production of IL-1β and PGE2 (Fig. 1). Parthenolide also inhibited lipopolysaccharide-induced increase in the levels of an inducible enzyme cyclooxygens-2, which plays a critical role in inflammation. Treatment with parthenolide alone, up to 10 μM, did not significantly induce cytokine production in cells not treated with lipopolysaccharide. We examined whether the lipopolysaccharide-induced production of inflammatory mediators was mediated by the NF-kB, Akt, and mTOR signaling pathways. Treatment with 2.5 μM Bay 11–7085 (an irreversible inhibitor of IkB-α phosphorylation), 0.5 μM Akt inhibitor or 0.5 μM mTOR inhibitor rapamycin reduced the lipopolysaccharide-induced production and changes in the levels of IL-1β, PGE2, and cyclooxygens-2 (Fig. 2). Alone, the inhibitors did not induce production and changes in the levels of IL-1β, PGE2, and cyclooxygens-2. We examined whether lipopolysaccharide-induced production of inflammatory mediators was mediated by reactive oxygen species. One millimeter Thiol compound N-acetylcysteine reduced the lipopolysaccharide-induced production and changes in the levels of IL-1β, PGE2, and cyclooxygens-2 (Fig. 2).

Effect of parthenolide on the production and changes in the levels of inflammatory mediators. HEK001 keratinocytes were pre-treated with 0.5–10 μM parthenolide for 20 min and exposed to 1 μg/ml lipopolysaccharide (LPS) in combination with parthenolide for 24 h. The levels of IL-1β, PGE2, and cyclooxygenase-2 (COX-2) were measured using ELISA or parametric kits. Data represent the mean ± SEM (n = 6). + p < 0.05 compared to the control; *p < 0.05 compared to lipopolysaccharide alone

Effect of signaling inhibitors and N-acetylcysteine on the production and changes in the levels of inflammatory mediators. Keratinocytes were treated with lipopolysaccharide in the presence of 2.5 μM Bay 11–7085, 0.5 μM Akt inhibitor, 0.5 μM rapamycin, or 1 mM N-acetylcysteine for 24 h. The levels of IL-1β, PGE2, and cyclooxygenase-2 (COX-2) were measured using ELISA or parametric kits. Data represent the mean ± SEM (n = 6). + p < 0.05 compared to the control; *p < 0.05 compared to lipopolysaccharide alone
The effect of parthenolide on NF-κB activation
The effect of parthenolide on lipopolysaccharide-induced activation of Toll-like receptor 4, which results in the stimulated response of keratinocytes, was examined. Lipopolysaccharide increased the Toll-like receptor 4 levels, which was inhibited by the addition of 5 μM parthenolide or 1 mM N-acetylcysteine (Fig. 3a). Next, we examined whether the effect of parthenolide on the lipopolysaccharide-induced production of inflammatory mediators in keratinocytes resulted from altered NF-κB activation. Treatment with lipopolysaccharide produced an increase in NF-κB p65, NF-κB p50, and phospho-IκB levels in keratinocytes (Fig. 3b). Treatment with 5 μM parthenolide, 2.5 μM Bay 11–7085, 0.5 μM Akt inhibitor, or 0.5 μM rapamycin prevented the lipopolysaccharide-induced IkB-α phosphorylation and NF-κB activation.

Effect of parthenolide on Toll-like receptor 4 levels and NF-κB activation. Keratinocytes were pre-treated with compounds (5 μM parthenolide, 2.5 μM Bay 11–7085, 0.5 μM Akt inhibitor or 0.5 μM rapamycin, or 1 mM N-acetylcysteine (NAC)) for 20 min and exposed to 1 μg/ml lipopolysaccharide in combination with compounds for 30 min (for NF-κB) or 24 h (for Toll-like receptor 4). In A and B, the levels of Toll-like receptor 4, NF-κB p65, NF-κB p50, phospho-IκB-α, and β-actin were analyzed by Western blotting with specific antibodies. Data are representative of three different experiments. The densities of protein bands were determined by densitometry and the data represent a fold increase from the control density. In C, the NF-κB to DNA binding activity was measured using an assay kit. Data represent the mean ± SEM (n = 5). + p < 0.05 compared to the control; *p < 0.05 compared lipopolysaccharide alone
The inhibitory effect of parthenolide on lipopolysaccharide-induced activation of NF-κB was confirmed by monitoring the binding of NF-κB to DNA. Non-stimulated cells exhibited a small increase in NF-κB-DNA binding activity. Treatment with lipopolysaccharide produced a marked increase in the NF-κB-DNA binding activity, which was reduced by the addition of parthenolide, Bay 11–7085, Akt inhibitor rapamycin, or N-acetylcysteine (Fig. 3c).
The effect of parthenolide on Akt activation
The involvement of the Akt/mTOR pathways in the lipopolysaccharide-induced production of inflammatory mediators was examined. Treatment with lipopolysaccharide increased the levels of phospho-Akt and mTOR in keratinocytes. The levels of phospho-Akt increased over time and reached a peak value after 4 h of treatment, after which the levels slightly declined (data not shown). To explore, we assessed the effect on phospho-Akt levels with a 4-h exposure to lipopolysaccharide. The lipopolysaccharide-induced activation of Akt was confirmed by the preventive effect of the specific Akt inhibitor. The addition of parthenolide or N-acetylcysteine inhibited the lipopolysaccharide-induced increase in the phospho-Akt levels (Fig. 4a). Akt inhibitor, parthenolide, or N-acetylcysteine alone did not induce phosphorylation of Akt. We further examined the inhibitory effect of parthenolide on changes in the levels of mTOR by Western blot analysis. Parthenolide, Akt inhibitor, or rapamycin inhibited the lipopolysaccharide-induced increase in the levels of mTOR (Fig. 4b). Akt inhibitor, parthenolide, or rapamycin alone did not induce changes in the levels of mTOR (data not shown).

Effect of parthenolide on activation of Akt. a keratinocytes were treated with 1 μg/ml lipopolysaccharide in the presence of compounds (5 μM parthenolide, 0.5 μM Akt inhibitor, or 1 mM N-acetylcysteine (NAC)) for 4 h, and then the level of phospho-Akt was measured by ELISA. Data represent the mean ± SEM (n = 6). + p < 0.05 compared to the control; *p < 0.05 compared to lipopolysaccharide treatment alone. b HEK001 keratinocytes were treated with 1 μg/ml lipopolysaccharide in the presence of compounds (5 μM parthenolide, 0.5 μM Akt inhibitor, 0.5 μM rapamycin or 1 mM N-acetylcysteine (NAC)) for 4 h, and the levels of mTOR were analyzed by Western blotting and data are representative of three to four different experiments. The densities of protein bands were determined by densitometry and the data represent a fold increase from the control density
The effect of parthenolide on formation of reactive oxygen species
The effect of parthenolide on the formation of reactive oxygen species as the responses of stimulated keratinocytes was examined. The formation of reactive oxygen species within cells was determined by monitoring a conversion of DCFH2-DA to fluorescent DCF. Keratinocytes treated with lipopolysaccharide for 24 h showed a significant increase in DCF fluorescence, which was inhibited by the addition of 1 mM N-acetylcysteine or 40 μM trolox (a scavenger of hydroxyl radicals and peroxynitrite) (Fig. 5). Treatment with 2.5–5 μM parthenolide attenuated the lipopolysaccharide-induced increase in DCF fluorescence.

Effect of parthenolide on formation of reactive oxygen species. Keratinocytes were treated with treated with 1 μg/ml lipopolysaccharide in the presence of compounds (2.5–5 μM parthenolide, 1 mM N-acetylcysteine (NAC) or 40 μM trolox) for 24 h. Then, the changes in DCF fluorescence, which indicates formation of reactive oxygen species formation, were measured and data are expressed as arbitrary units (a.u.) of fluorescence. Data represent mean ± SEM (n = 6). + p < 0.05 compared to control; *p < 0.05 compared to lipopolysaccharide alone
The effect of parthenolide on cell viability
To examine whether the inhibitory effect of parthenolide on the stimulated keratinocyte response can be attributed to an effect on cell viability, we assessed the cytotoxic effect of parthenolide by using an MTT assay, which provides rapid and precise results for cellular growth and survival. When the keratinocytes were treated with parthenolide for 24 h, cell viability decreased with concentration. At 5 μM, parthenolide showed an approximately 4 % loss in cell viability (Fig. 6). At 7.5 and 10 μM, treatment with parthenolide resulted in approximately 14 and 24 % cell viability loss, respectively.

Effect of parthenolide on cell viability. Keratinocytes were treated with 0.5–10 μM of parthenolide for 24 h and then cell viability was determined using an MTT reduction assay or a WST-8 reduction assay. Data represent mean ± SEM (n = 6). + cpr < 0.05 compared to the control (percentage of control)
We further examined the effect of parthenolide on cell viability by monitoring a WST-8 reduction using cell counting kit-8. When the keratinocytes were treated with parthenolide for 24 h, cell viability decreased with concentration. At 5 μM, parthenolide showed an approximately 4 % loss in cell viability (Fig. 6). At 7.5 and 10 μM, treatment with parthenolide resulted in approximately 16 and 27 % cell viability loss, respectively
Discussion
Keratinocytes treated with lipopolysaccharide produced a significant amount of cytokine IL-1β and proinflammatory mediator PGE2, and increased the levels of an inducible inflammatory enzyme cyclooxygenase-2. Parthenolide has been shown to have an anti-inflammatory effect (Feltenstein et al. 2004; Juliana et al. 2010; Kwok et al. 2001). Nevertheless, the effect of parthenolide on the production and changes in the levels of inflammatory mediators in kerstinocytes exposed to lipopolysaccharide has not been studied. In the present study, parthenolide significantly inhibited the lipopolysaccharide-induced production of proinflammatory mediators in keratinocytes. Proinflammatory cytokines elicit the amplification and persistence of inflammatory and immune responses in skin (Pastore et al. 2006). Cyclooxygenase-2 is upregulated during inflammatory states and produces prostanoids responsible for the generation of pain and inflammation (Vane and Botting 1998). Thus, parthenolide appears to reduce inflammatory reaction and immune cell function by suppressing the productions of cytokine and proinflammatory mediator and the expression of an inducible inflammatory enzyme.
Lipopolysaccharide induces the production of cytokines by activating the Toll-like receptor 4-mediated NF-κB pathway (Takeda and Akira 2005; Thomson et al. 2009). NF-κB is required for cell survival and immunity, but the abnormal expression or activation of NF-κB leads to the chronic and acute inflammation (Kwak et al. 2011; Sun and Zhang 2007). The aberrant activation of NF-κB in both keratinocytes and lymphocytes may be involved in the development of inflammatory skin diseases (Rebholz et al. 2007). The basal NF-κB-DNA binding activity in peripheral blood mononuclear cells was found to be significantly higher in atopic eczema patients than healthy age-matched control patients (Angelini et al. 2007). Lipopolysaccharide induces the production of cytokines in keratinocytes through the activation of NF-κB (Ge et al. 2012; Lee et al. 2010). The phosphorylation and proteolytic degradation of IκB induce the release of NF-κB dimers (Napetschnig and Wu 2013). The translocation of active NF-κB dimers to the nucleus elicits the activation of target genes responsible for the synthesis of cytokines and chemokines (Ghosh and Hayden 2008; Vestergaard et al. 2005). In the present study, lipopolysaccharide increased the levels of the Toll-like receptor 4, the levels of phospho-IκB and NF-κB p65/50, and the binding of NF-κB to DNA in keratinocytes. The lipopolysaccharide-induced activation of Toll-like receptor 4 elicits a nuclear translocation of NF-κB and the secretion of proinflammatory cytokines and chemokines (Song et al. 2002). Changes in the levels of Toll-like receptor 4 and activation of NF-κB suggest that lipopolysaccharides stimulate production of inflammatory mediators by activating the Toll-like receptor 4-mediated NF-κB pathway, i.e., the nuclear translocation and specific DNA binding of activated NF-κB dimers. Then, we measured whether the preventive effect of parthenolide on the lipopolysaccharide-induced production of inflammatory mediators in keratinocytes was mediated by its inhibitory effect on the Toll-like receptor 4-mediated activation of NF-κB. Parthenolide has been shown to inhibit NF-κB activation (Juliana et al. 2010; López-Franco et al. 2006; Nam 2006). In the present study, parthenolide inhibited the lipopolysaccharide-induced changes in the levels of Toll-like receptor 4 and activation of NF-κB in keratinocytes. Accordingly, these results suggest that parthenolide may inhibit the lipopolysaccharide-induced production of inflammatory mediators by suppressing the Toll-like receptor 4-mediated activation of NF-κB. The NF-κB regulates the transcription genes involved in the inflammatory response (Ghosh and Hayden 2008). Based on this report, parthenolide appears to reduce the lipopolysaccharide-induced production of inflammatory mediators by suppressing the production of mRNAs responsible for producing cytokines.
Toll-like receptor-4 has been established as the receptor for lipopolysaccharide (Pålsson-McDermott and O'Neill 2004; Takeda and Akira 2005). Circulating lipopolysaccharide-binding protein recognizes lipopolysaccharide in the plasma and brings it to CD14. This aids the loading of lipopolysaccharide onto the lipopolysaccharide receptor complex, which is composed of dimerized Toll-like receptor 4 receptors and two molecules of the extracellular adapter MD-2. The binding of lipopolysaccharide onto the lipopolysaccharide receptor complex is followed by binding of the adapter’s myeloid differentiation primary response protein 88 (MyD88) and MyD88-adapter-like (Mal) or not. The lipopolysaccharide signaling leads to the early activation of NF-κB, interferon regulatory factor-3, and MAPK kinase pathways, which is mediated by the adapters MyD88 and Mal (Pålsson-McDermott and O'Neill 2004). Parthenolide has been shown to inhibit the MyD88-dependent pathway by inhibiting the activity of inhibitor-κB kinase in RAW264.7 macrophages (Park et al. 2011). Parthenolide inhibits NF-κB and interferon regulatory factor 3 activation induced by lipopolysaccharide in macrophage cell lines. Thus, parthenolide appears to attenuate the MyD88-dependent-NF-κB and/or interferon regulatory factor 3 pathways. Parthenolide has been demonstrated to inhibit the lipopolysaccharide-induced expression of toll-like receptor 4 at both protein and mRNA levels in human leukemia monocytic THP-1 cells (Li et al. 2015). Parthenolide appears to reduce the levels of Toll-like receptor 4 by suppressing the synthesis of Toll-like receptor 4 in keratinocytes treated with lipopolysaccharide.
Lipopolysaccharide induces NF-κB activation by activating the phosphatidylinositol (PI) 3-kinase/Akt/mTOR pathway (Lee et al. 2008; Thomson et al. 2009; Zhong et al. 2012). We examined whether lipopolysaccharide-induced NF-κB activation and subsequent production of inflammatory mediators in keratinocytes was mediated by Akt/mTOR signaling. In the present study, Akt inhibitor and rapamycin reduced the lipopolysaccharide-induced activation of NF-κB and production of inflammatory mediators. Keratinocytes exposed to lipopolysaccharide had activated (phosphorylated) Akt and increased the levels of mTOR, which was inhibited by the addition of Akt inhibitor, and rapamycin. The inhibitory effect on the phosphorylation of Akt and on the changes in the levels of mTOR suggests that parthenolide appears to inhibit the lipopolysaccharide-induced production of inflammatory mediators by suppressing the Akt/mTOR-regulated NF-κB activation.
Parthenolide at 10 μM induced cell death by approximately 24 % in keratinocytes, whereas at the concentration of 5 μM used in the assays of signaling pathway activation, it induced cell death by approximately 4 %. Therefore, the inhibitory effect of parthenolide at 5 μM on the Akt/mTOR and NF-κB pathways involved in the production of inflammatory mediators not appear to be significantly affected by changes in cell viability.
Reactive oxygen species may act as key effectors in the regulation of cell functions and induce the activation of NF-κB (Gloire et al. 2006; Siomek 2012). We investigated the formation of reactive oxygen species in the lipopolysaccharide-stimulated keratinocytes, which may be involved in the NF-κB activation. N-acetylcysteine inhibits the TNF-α- or lipopolysaccharide-induced cytokine production by suppressing reactive oxygen species formation (Hsu and Wen 2002; Young et al. 2008). In this study, oxidant scavenger N-acetylcysteine attenuated the lipopolysaccharide-induced changes in the levels and activity of Toll-like receptor-4, NF-κB, Akt, and mTOR, and the formations of reactive oxygen species. Thus, the Akt, mTOR, and NF-κB pathways appear to be regulated by reactive oxygen species.
Parthenolide inhibits nitric oxide production and iNOS mRNA expression by a mechanism involving stabilization of the I-κBα/NF-κB complex in cultured rat aortic smooth muscle cells treated with lipopolysaccharide and interferon-γ (Wong and Menendez 1999). However, it is unclear that the inhibitory effect of parthenolide on the NF-κB-mediated inflammatory responses in experimental atherosclerosis is mediated by its effect on reactive oxygen species, because parthenolide affects neither apoptotic cell death nor oxidative stress (López-Franco et al. 2006). In the present study, parthenolide at the concentration, which did not affect significant cell viability, inhibited the lipopolysaccharide-induced changes in the levels and activity of signal transduction proteins, and the formation of reactive oxygen species. Thus, the present results suggest that parthenolide appears to reduce the lipopolysaccharide-induced signal transduction-mediated production of proinflammatory mediators by suppressing formation of reactive oxygen species.
Overall, the results show that parthenolide appears to attenuate lipopolysaccharide-stimulated production of inflammatory mediators in keratinocytes by suppressing the activation of the Akt, mTOR, and NF-κB pathways, which are mediated by activation of Toll-like receptor 4. The activation of signaling transduction pathways appears to be regulated by reactive oxygen species. Thereby, parthenolide appears to attenuate the microbial product-mediated inflammatory skin diseases.
References
Angelini F, Di Matteo G, Balestrero S, Brunetti E, Mancino G, Rossi P, Galli E (2007) Nuclear factor κB activity is increased in peripheral blood mononuclear cells of children affected by atopic and non-atopic eczema. Int J Immunopathol Pharmacol 20:59–67
Baker BS (2006) The role of microorganisms in atopic dermatitis. Clin Exp Immunol 144:1–9
Begon E, Michel L, Flageul B, Beaudoil I, Jean-Louis F, Bachelez H, Dubertret L, Musette P (2007) Expression, subcellular localization and cytokinic modulation of Toll-like receptors (TLRs) in normal human keratinocytes: TLR2 up-regulation in psoriatic skin. Eur J Dermatol 17:497–506
Feltenstein MW, Schühly W, Warnick JE, Fischer NH, Sufka KJ (2004) Anti-inflammatory and anti-hyperalgesic effects of sesquiterpene lactones from Magnolia and Bear's foot. Pharmacol Biochem Behav 79:299–302
Fu W, Luo H, Parthasarathy S, Mattson MP (1998) Catecholamines potentiate amyloid β-peptide neurotoxicity: involvement of oxidative stress, mitochondrial dysfunction, and perturbed calcium homeostasis. Neurobiol Dis 5:229–243
Ge Y, Xu Y, Sun W, Man Z, Zhu L, Xia X, Zhao L, Zhao Y, Wang X (2012) The molecular mechanisms of the effect of dexamethasone and cyclosporin A on TLR4 /NF-κB signaling pathway activation in oral lichen planus. 508:157–164.
Ghosh S, Hayden MS (2008) New regulators of NF-κB in inflammation. Nat Rev Immunol 8:837–848
Gloire G, Legrand-Poels S, Piette J (2006) NF-κB activation by reactive oxygen species: fifteen years later. Biochem Pharmacol 72:1493–1505
Hsu HY, Wen MH (2002) Lipopolysaccharide-mediated reactive oxygen species and signal transduction in the regulation of interleukin-1 gene expression. J Biol Chem 277:22131–22139
Ishiyama M, Miyazono Y, Sasamoto K, Ohkura Y, Ueno K (1997) A highly water-soluble disulfonated tetrazolium salt as a chromogenic indicator for NADH as well as cell viability. Talanta 44:1299–1305
Juliana C, Fernandes-Alnemri T, Wu J, Datta P, Solorzano L, Yu JW, Meng R, Quong AA, Latz E, Scott CP, Alnemri ES (2010) Anti-inflammatory compounds parthenolide and Bay 11–7082 are direct inhibitors of the inflammasome. J Biol Chem 285:9792–9802
Kang SS, Kauls LS, Gaspari AA (2006) Toll-like receptors: applications to dermatologic disease. J Am Acad Dermatol 54:951–983
Kwak JH, Jung JK, Lee H (2011) Nuclear factor-κB inhibitors; a patent review (2006–2010). Expert Opin Ther Pat 21:1897–1910
Kwok BH, Koh B, Ndubuisi MI, Elofsson M, Crews CM (2001) The anti-inflammatory natural product parthenolide from the medicinal herb Feverfew directly binds to and inhibits IκB kinase. Chem Biol 8:759–766
Lee SA, Park SH, Kim BC (2008) Raloxifene, a selective estrogen receptor modulator, inhibits lipopolysaccharide-induced nitric oxide production by inhibiting the phosphatidylinositol 3-kinase/Akt/nuclear factor-κB pathway in RAW264.7 macrophage cells. Mol Cells 26:48–52
Lee CS, Jang ER, Kim YJ, Lee MS, Seo SJ, Lee MW (2010) Hirsutenone inhibits lipopolysaccharide-activated NF-kappaB-induced inflammatory mediator production by suppressing Toll-like receptor 4 and ERK activation. Int Immunopharmacol 10:520–525
Li S, Gao X, Wu X, Wu Z, Cheng L, Zhu L, Shen D, Tong X (2015) Parthenolide inhibits LPS-induced inflammatory cytokines through the toll-like receptor 4 signal pathway in THP-1 cells. Acta Biochim Biophys Sin (Shanghai). Apr 4. pii: gmv019. [Epub ahead of print]
Li-Weber M, Giaisi M, Treiber MK, Krammer PH (2002) The anti-inflammatory sesquiterpene lactone parthenolide suppresses IL-4 gene expression in peripheral blood T. Eur J Immunol 32:3587–3597
López-Franco O, Hernández-Vargas P, Ortiz-Muñoz G, Sanjuán G, Suzuki Y, Ortega L, Blanco J, Egido J, Gómez-Guerrero C (2006) Parthenolide modulates the NF-kappaB-mediated inflammatory responses in experimental atherosclerosis. Arterioscler Thromb Vasc Biol 26:1864–1870
Magni P, Ruscica M, Dozio E, Rizzi E, Beretta G, Maffei Facino R (2012) Parthenolide inhibits the LPS-induced secretion of IL-6 and TNF-α and NF-κB nuclear translocation in BV-2 microglia. Phytother Res 26:1405–1409
Marchi S, Giorgi C, Suski JM, Agnoletto C, Bononi A, Bonora M, De Marchi E, Missiroli S, Patergnani S, Poletti F, Rimessi A, Duszynski J, Wieckowski MR, Pet P (2012) Mitochondria-ros crosstalk in the control of cell death and aging. J Signal Transduct 2012:329635
Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65:55–63
Nam NH (2006) Naturally occurring NF-κB inhibitors. Mini Rev Med Chem 6:945–951
Napetschnig J, Wu H (2013) Molecular basis of NF-κB signaling. Annu Rev Biophys 42:443–468
Pålsson-McDermott EM, O'Neill LA (2004) Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 113:153–162
Park SJ, Shin HJ, Youn HS (2011) Parthenolide inhibits TRIF-dependent signaling pathway of Toll-like receptors in RAW264.7 macrophages. Mol Cells 31:261–265
Pastore S, Mascia F, Girolomoni G (2006) The contribution of keratinocytes to the pathogenesis of atopic dermatitis. Eur J Dermatol 16:125–131
Rebholz B, Haase I, Eckelt B, Paxian S, Fliag MJ, Ghoreshi K, Nedopasov SA, Mailhammer R, Debey-Pascher S, Schultz JL, Weindl G, Fӧrster I, Huss R, Stratis A, Ruzicka T, Rӧcken PK, Schmid RM, Rupec RA (2007) Crosstalk between keratinocytes and adaptive immune cells in an IκBα protein-mediated inflammatory disease of the skin. Immunity 27:296–307
Schreiber E, Matthias P, Müller MM, Schaffner W (1989) Rapid detection of octamer binding proteins with 'mini-extracts', prepared from a small number of cells. Nucleic Acids Res 17:6419
Siomek A (2012) NF-κB signaling pathway and free radical impact. Acta Biochim Pol 59:323–331
Smolinski AT, Pestka JJ (2005) Comparative effects of the herbal constituent parthenolide (Feverfew) on lipopolysaccharide-induced inflammatory gene expression in murine spleen and liver. J Inflamm (Lond) 2:6
Song PI, Park YM, Abraham T, Harten B, Zivony A, Neparidze N, Armstrong CA, Ansel JC (2002) Human keratinocytes express functional CD14 and toll-like receptor 4. J Invest Dermatol 119:424–432
Sun XF, Zhang H (2007) NFκB and NFκBI polymorphisms in relation to susceptibility of tumour and other diseases. Histol Histopathol 22:1387–1398
Takeda K, Akira S (2005) Toll-like receptors in innate immunity. Int Immunol 17:1–14
Thomson AW, Turnquist HR, Raimondi G (2009) Immunoregulatory functions of mTOR inhibition. Nat Rev Immunol 9:324–337
Tsuruta D (2009) NF-κB links keratinocytes and lymphocytes in the pathogenesis of psoriasis. Recent Pat Inflamm Allergy Drug Discov 3:40–48
Vane JR, Botting RM (1998) Anti-inflammatory drugs and their mechanism of action. Inflamm Res 47(Suppl 2):S78–S87
Vestergaard C, Johansen C, Otkjaer K, Deleuran M, Iversen L (2005) Tumor necrosis factor-alpha-induced CTACK/CCL27 (cutaneous T-cell-attracting chemokine) production in keratinocytes is controlled by nuclear factor κB. Cytokine 29:49–55
Wong HR, Menendez IY (1999) Sesquiterpene lactones inhibit inducible nitric oxide synthase gene expression in cultured rat aortic smooth muscle cells. Biochem Biophys Res Commun 262:375–380
Young CN, Koepke JI, Terlecky LJ, Borkin MS, Boyd SL, Terlecky SR (2008) Reactive oxygen species in tumor necrosis factor-α-activated primary human keratinocytes: implications for psoriasis and inflammatory skin disease. J Invest Dermatol 128:2606–2614
Yuan G, Wahlqvist ML, He G, Yang M, Li D (2006) Natural products and anti-inflammatory activity. Asia Pac J Clin Nutr 15:143–152
Zhong LM, Zong Y, Sun L, Guo JZ, Zhang W, He Y, Song R, Wang WM, Xiao CJ, Lu D (2012) Resveratrol inhibits inflammatory responses via the mammalian target of rapamycin signaling pathway in cultured LPS-stimulated microglial cells. PLoS One 7, e32195
Acknowledgments
This research was supported by the Chung-Ang University Research Scholarship Grants in 2015, Chung-Ang University, Seoul, South Korea.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Nam, Y.J., Lee, D.H., Lee, M.S. et al. Sesquiterpene lactone parthenolide attenuates production of inflammatory mediators by suppressing the Toll-like receptor-4-mediated activation of the Akt, mTOR, and NF-κB pathways. Naunyn-Schmiedeberg's Arch Pharmacol 388, 921–930 (2015). https://doi.org/10.1007/s00210-015-1132-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00210-015-1132-3