
Abstract
In this paper, we study a three-patch two-species Lotka–Volterra competition patch model over a stream network. The individuals are subject to both random and directed movements, and the two species are assumed to be identical except for the movement rates. The environment is heterogeneous, and the carrying capacity is lager in upstream locations. We treat one species as a resident species and investigate whether the other species can invade or not. Our results show that the spatial heterogeneity of environment and the magnitude of the drift rates have a large impact on the competition outcomes of the stream species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The species living in stream environment is subject to both passive random movement and directed drift [47]. Intuitively, the drift will carry individuals to the downstream end, which may be crowded or hostile. However, random dispersal may drive the individuals to the upper stream locations, which are usually more favorable for the species [23]. Therefore, the joint impact of both undirectional and directed dispersal rates on the population dynamics of the species is usually complicated and has attracted increasing research interests recently [22, 25, 34, 40,41,42, 47].
Dispersal has profound effects on the distribution and abundance of organisms, and understanding the mechanisms for the evolution of dispersal is a fundamental question related to dispersal [26]. In the seminal works of Hastings [19] and Dockery et al. [15], it has been shown that in a spatially heterogeneous environment, when two competing species are identical except for the random dispersal rate, evolution of dispersal favors the species with a smaller dispersal rate. However, in an advective environment when individuals are subject to both undirectional random dispersal and directed movement, species with a faster dispersal rate can be selected [3, 4, 11].
Two-species reaction–diffusion–advection competition models of the following form have been proposed to study the evolution of dispersal for stream species [28, 34,35,36, 38, 43, 48, 49, 51,52,53]:
In [28, 34, 49], the authors have treated species u as a resident species and studied the conditions under which the species u only semitrivial equilibrium is stable/unstable. Various results on the global dynamics of (1.1) are presented in [36, 38, 43, 52, 53]. In particular, if r(x) is constant, the works [36, 38, 53] show that the species with a larger diffusion rate and/or a smaller advection rate wins the competition. If r(x) is a decreasing function, the authors in [37, 52] use \(q_1\) and \(q_2\) as bifurcation parameters to study the global dynamics of (1.1) and the related results will be discussed later (see Remark 3.14).
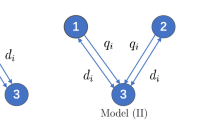
To study the evolution of dispersal in a river network, the authors in [23, 24] propose and investigate three-patch two-species Lotka–Volterra competition models. Let \(\varvec{u}=(u_1, u_2, u_3)\) and \(\varvec{v}=(v_1, v_2, v_3)\) be the population density of two competing species, respectively, where \(u_i\) and \(v_i\) are the densities in patch i. Suppose that the dispersal patterns of the individuals and the configuration of the patches are shown in Fig. 1.

A stream with three patches, where d is the random movement rate and q is the directed drift rate. Patch 1 is the upstream end, and patch 3 is the downstream end
The competition patch model over the stream network in Fig. 1 (with \(r_1=r_2=r_3\)) in [23, 24] is:
where \(d_1\) and \(d_2\) are random movement rates; \(q_1\) and \(q_2\) are directed movement rates; \(\varvec{r}=(r_1, r_2, r_3)\) is the growth rate; \(\varvec{k}=(k_1, k_2, k_3)\) is the carrying capacity; and two \(3\times 3\) matrices \(D=(D_{ij})\) and \(Q=(Q_{ij})\) represent the random movement pattern and directed drift pattern of individuals, respectively, where
We can write the model as
We assume \(d_1, d_2, q_1, q_2>0\) and \(r_i, k_i>0\) for \(i=1, 2, 3\). We adopt the same assumption in [23] on \(\varvec{k}=(k_1, k_2, k_3)\):
- \((\textbf{H})\):
-
\(k_1> k_2> k_3>0\).
Biologically, \((\mathbf {}{H})\) means that the upstream locations are more favorable for both species.
Two-species Lotka–Volterra competition patch models have attracted many research interests recently. Model (1.2) with n patches in spatially homogeneous environment (i.e., \(r_1=\dots =r_n\) and \(k_1=\dots =k_n\)) has been considered in our earlier papers [7, 10], but many techniques and results there cannot be generalized to the situation when \(k_1=\dots =k_n\) is not assumed. The authors in [18, 45] have studied the global dynamics of model (1.2) with two patches and \(q:=q_1=q_2\). They have showed that there exists a critical drift rate such that below it the species with a smaller dispersal rate wins the competition, while above it the species with a larger dispersal rate wins. In a competition model with two patches, the authors in [12, 17, 32] have showed that the species with more evenly distributed resources has less competition advantage. In [8], the global dynamics of a Lotka–Volterra competition patch model is classified under some assumptions on patches, which requires \(d_1/q_1=d_2/q_2\) in terms of (1.2). For more studies on competition patch models, we refer to the works [2, 5, 6, 27, 30, 33, 44, 46, 50].
We will take an adaptive dynamics approach [14, 16] to analyze (1.4) by viewing species \(\varvec{u}\) as the resident species and species \(\varvec{v}\) as the mutant species. Model (1.4) has two semitrivial equilibria \((\varvec{u}^*,\varvec{0})\) and \((\varvec{0},\varvec{v}^*)\). We fix parameters \(d_1\) and \(q_1\) and vary \(d_2\) and \(q_2\). We show that there exists a curve \(q=q_{\varvec{u}}^*(d)\) dividing the \((d_2, q_2)\)-plane into two regions such that \((\varvec{u}^*, \varvec{0})\) is stable if and only if \((d_2, q_2)\) is above the curve. Our results complement those in [23] by defining and analyzing the curve \(q=q_{\varvec{u}}^*(d)\) and obtaining the global dynamics of model (1.4). In particular, we show that if \(q_1<{\underline{q}}\) the curve \(q=q_{\varvec{u}}^*(d)\) is bounded (see Fig. 3) and if \(q_1>{{\overline{q}}}\) it is unbounded (see Fig. 4). This result is in sharp contrast with the corresponding one for the model in spatially homogeneous environment (\(k_1=k_2=k_3\)) [7], where the curve \(q=q_{\varvec{u}}^*(d)\) is always unbounded. We give explicitly parameter ranges for competitive exclusion and conditions for coexistence/bistability in three cases (\(q_1<{\underline{q}}\), \({\underline{q}}\le q_1\le {{\overline{q}}}\) and \(q_1>{{\overline{q}}}\)). Our results show that the magnitude of the drift rates and the spatial heterogeneity of environment have a large impact on the competition outcomes of the stream species.
Our paper is organized as follows. In Sect. 2, we list some preliminary results. In Sect. 3, we state the main results on model (1.4). We give some conclusive remarks and numerical simulations in Sect. 4. The proofs of the main results are presented in Sect. 5. In the Appendix, we show the relations of \({\underline{q}}\), \({{\overline{q}}}\), and \(q_0\). These relations are implicitly included in the main results, and we prove them for reader’s convenience.
2 Preliminary
Let \(A=(a_{ij})_{n\times n}\) be a square matrix with real entries, \(\sigma (A)\) be the set of all eigenvalues of A, and s(A) be the spectral bound of A, i.e., \(s(A)=\max \{\textrm{Re} \lambda : \lambda \in \sigma (A)\}\). The matrix A is called irreducible if it cannot be placed into block upper triangular form by simultaneous row and column permutations and essentially nonnegative if \(a_{ij}\ge 0\) for all \(1\le i, j\le n\) and \(i\ne j\). By the Perron–Frobenius theorem, if A is irreducible and essentially nonnegative, then s(A) is an eigenvalue of A (called the principal eigenvalue of A), which is the unique eigenvalue associated with a nonnegative eigenvector. The following result on the monotonicity of spectral bound can be found in [1, 9]:
Lemma 2.1
Let \(A=(a_{ij})_{n\times n}\) be an irreducible and essentially nonnegative matrix and \(M=\text {diag}(m_i)\) be a real diagonal matrix. If \(s(A)=0\), then
for \(\mu \in (0, \infty )\) and the inequality is strict except for the case \(m_1=\cdots =m_n\). Moreover,
where \(\theta _i\in (0, 1)\), \(1\le i\le n\), is determined by A and \(\displaystyle \sum _{i=1}^n{\theta _i}=1\) (if A has each column sum equaling zero, then \(\varvec{\theta }=(\theta _1,\dots ,\theta _n)^T\) is a positive eigenvector of A corresponding to eigenvalue 0).
Let \(\varvec{m}=(m_1, m_2, m_3)\) be a real vector. We write \(\varvec{m}\gg \varvec{0}\) if \(m_i>0\) for all \(i=1, 2, 3\), and \(\varvec{m}>\varvec{0}\) if \(\varvec{m}\ge \varvec{0}\) and \(\varvec{m}\ne \varvec{0}\). Matrix \(dD+qQ+\text {diag}(m_i)\) is irreducible and essentially nonnegative for any \(d, q>0\), where D and Q are defined by (1.3). By the Perron–Frobenius theorem, \(s\left( dD+qQ+\text {diag}(m_i)\right) \) is the principal eigenvalue of the following eigenvalue problem:
We need to consider the following single-species patch model:
The global dynamics of (2.2) is as follows:
Lemma 2.2
Let D and Q be defined in (1.3), \(\varvec{r}, \varvec{k}\gg \varvec{0}\), \(d>0\), and \(q\ge 0\). Then, model (2.2) admits a unique positive equilibrium \(\varvec{u}^*\gg \varvec{0}\), which is globally asymptotically stable.
Proof
By [13, 31, 39], it suffices to show that \(\varvec{0}\) is unstable, i.e.,
Let \(\phi ^T=(\phi _1, \phi _2, \phi _3)^T\gg \varvec{0}\) with \(\sum _{i=1}^3\phi _i=1\) be the positive eigenvector of \(dD+qQ+\text {diag}(r_i)\) corresponding to s. Multiplying (1, 1, 1) to the left of \(dD\phi +qQ\phi +\text {diag}({r_i})\phi =s\phi \), we get \(s=\sum _{i=1}^3{{r_i\phi _i}}>0\). This proves the result. \(\square \)
By Lemma 2.2, model (1.4) has two semitrivial equilibria \((\varvec{u}^*, \varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\), where \(\varvec{u}^* (\text {resp., }\varvec{v}^*)\gg \varvec{0}\) is the positive equilibrium of (2.2) with (d, q) replaced by \((d_1, q_1)\) (resp., \((d_2, q_2)\)). Linearizing model (1.4) at \((\varvec{u}^*,\varvec{0})\), we can easily see that its stability is determined by the sign of \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) \), which is the principal eigenvalue of the following eigenvalue problem:
In particular, \((\varvec{u}^*,\varvec{0})\) is locally asymptotically stable if \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) <0\) and unstable if \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) >0\). Here, we abuse the notation by denoting \(\varvec{1}-{\varvec{u}^*}/{\varvec{k}}:=(1-u_1^*/k_1, 1-u_2^*/k_2, 1-u_3^*/k_3)\).
3 Main results
We fix \(d_1, q_1>0, \varvec{r}, \varvec{k}\gg \varvec{0}\) and view species \(\varvec{u}\) as the resident species and \(\varvec{v}\) as the mutant species. We investigate the dynamics of model (1.4) varying \((d_2, q_2)\). For this purpose, we divide the first quadrant of the (d, q)-plane into six regions:

Illustration of the six regions of (d, q)-plane
For readers’ convenience, we graph the six regions in Fig. 2.
3.1 Invasion curve
We consider the local stability of \((\varvec{u}^*,\varvec{0})\) in this subsection. Biologically, if \((\varvec{u}^*,\varvec{0})\) is stable, then a small amount of species \(\varvec{v}\) cannot invade species \(\varvec{u}\); if \((\varvec{u}^*,\varvec{0})\) is unstable, then a small amount of species \(\varvec{v}\) may be able to invade species \(\varvec{u}\). We prove that there exists a curve \(q=q_{\varvec{u}}^*(d)\) in the (d, q)-plane such that \((\varvec{u}^*,\varvec{0})\) is locally asymptotically stable if \((d_2, q_2)\) is above the curve and \((\varvec{u}^*,\varvec{0})\) is unstable if it is below the curve. To this end, we define
where \(d=d_0>0\) is the unique root of \(\lambda _1(d, 0, \varvec{1}-\varvec{u^*/k})=0\) if \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) < 0\) (see the existence of \(d_0\) in Lemma 5.2). We have the following result about the local stability/instability of the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\):
Theorem 3.1
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, there exists a continuous function \(q=q_{\varvec{u}}^*(d): (0, d^*)\rightarrow {\mathbb {R}}_+\) passing through \((d_1, q_1)\) such that the following statements hold for model (1.4):
-
(i)
If \((d_2, q_2)\in S_1\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is locally asymptotically stable;
-
(ii)
If \((d_2, q_2)\in S_2\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is unstable.
Here, \(S_1\cup S_2\) is a partition of the first quadrant of the (d, q)-plane defined as follows:
where
Remark 3.2
We call the curve in the first quadrant of (d, q)-plane defined by the function \(q=q_{\varvec{u}}^*(d)\) in Theorem 3.1 the invasion curve. This curve consists with all the points \((d, q_{\varvec{u}}^*(d))\) such that \(\lambda _1(d, q_{\varvec{u}}^*(d), \varvec{1}-{\varvec{u}^*}/{\varvec{k}})=0\), i.e., \((\varvec{u}^*,\varvec{0})\) is linearly neutrally stable. The invasion curve divides the first quadrant into \(S_1\cup S_2\), where \(S_1\) is the region above the curve and \(S_2\) is the region below it. By Theorem 3.1, \((\varvec{u}^*,\varvec{0})\) is locally asymptotically stable if \((d_2, q_2)\in S_1\) and unstable if \((d_2,q_2)\in S_2\).
In the following of this paper, we denote
We take \({\underline{q}}\) and \({{\overline{q}}}\) as the threshold values for the drift rates. Specifically, if a drift rate is below \({\underline{q}}\) (above \({{\overline{q}}}\)), we call it a slow (large) drift; if a drift rate is between \({\underline{q}}\) and \({{\overline{q}}}\), we call it an intermediate drift. These definitions coincide with those in [23] if \(r_1=r_2=r_3\). It turns out that the magnitude of drift rate \(q_1\) will have a large impact on the shape of the invasion curve and the dynamics of the model.
We have the following result about the invasion curve:
Proposition 3.3
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Let \(S_1\) and \(S_2\) be defined in Theorem 3.1. Then, the following statements hold:
-
(i)
\( G_{11}\subset S_1\) and \(G_{21}\subset S_2\);
-
(ii)
If \(q_1>{{\overline{q}}}\), then \(G_{12}\subset S_1\) and \(G_{22}\subset S_2\);
-
(iii)
If \(q_1<{\underline{q}}\), then \(G_{13}\subset S_1\) and \(G_{23}\subset S_2\)
We explore further properties of the invasion curve:
Proposition 3.4
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Let \(q=q_{\varvec{u}}^*(d): (0, d^*)\rightarrow {\mathbb {R}}_+\) be defined in Theorem 3.1. Then, the following statements hold:
-
(i)
\(\lim _{d\rightarrow 0}q_{\varvec{u}}^*(d)=q_0\), where
$$\begin{aligned} q_0=\max \left\{ r_1\left( 1-\frac{u_1^*}{k_1}\right) ,\ r_2\left( 1-\frac{u_2^*}{k_2}\right) \right\} ; \end{aligned}$$(3.6) -
(ii)
If \(q_1<{\underline{q}}\), then
$$\begin{aligned} d^*=d_0\;\;\text {and}\;\; \lim _{d\rightarrow d^*} q_{\varvec{u}}^*(d)=0; \end{aligned}$$(3.7) -
(iii)
If \(q_1>{{\overline{q}}}\), then
$$\begin{aligned} d^*=\infty \;\;\text {and}\;\;\lim _{d\rightarrow \infty } \displaystyle \frac{q_{\varvec{u}}^*(d)}{d}=\theta \end{aligned}$$(3.8)for some \(\theta \in \left( 0,{q_1}/{d_1}\right) \);
-
(iv)
If \({\underline{q}}\le q\le {{\overline{q}}}\), then (3.7) holds when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) < 0\), (3.8) holds with \(\theta \in \left( 0,{q_1}/{d_1}\right) \) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) > 0\), and (3.8) holds with \(\theta =0\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) =0\).
Remark 3.5
By Propositions 3.3 and 3.4, the invasion curve lies in \(G_{12}\cup G_{22}\) when the drift rate \(q_1\) is small, and it lies in \(G_{13}\cup G_{23}\) when \(q_1\) is large. Moreover, if \(q_1\) is small, the invasion curve is defined on a bounded interval \((0, d_0)\); if \(q_1\) is large, it is defined on \((0, \infty )\) and has a slant asymptote \(q=\theta d\) for some \(\theta \in (0, q_1/d_1)\).
3.2 Competitive exclusion
In this subsection, we study the global dynamics of model (1.4) and find some parameter ranges of \((d_2, q_2)\) such that competitive exclusion happens. The relations of \({\underline{q}}\), \({{\overline{q}}}\) and \(q_0\) are implicitly included in the results below. However, for reader’s convenience, we include the proof in the Appendix.
Firstly, we consider the small drift case, i.e., \(q_1<{\underline{q}}\).
Theorem 3.6
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\) with \(q_1<{\underline{q}}\). Then, the following statements hold:
-
(i)
If \((d_2,q_2)\in G_{21}\cup G_{23}\), then the semitrivial equilibrium \((\varvec{0},\varvec{v}^*)\) of (1.4) is globally asymptotically stable;
-
(ii)
If \((d_2,q_2)\in G_{11}\cup G_{12}^*\cup G_{13}\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) of (1.4) is globally asymptotically stable.
Here, \(G_{12}^*\) is defined by

Illustration of the results for the case \(q_1<{\underline{q}}\). If \((d_2, q_2)\) is above the curve \(q=q_{\varvec{u}}^*(d)\), then \((\varvec{u}^*,\varvec{0})\) is stable; and if \((d_2, q_2)\) is under the curve, then \((\varvec{u}^*,\varvec{0})\) is unstable. If \((d_2,q_2)\in G_{21}\cup G_{23}\), \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable; if \((d_2,q_2)\in G_{11}\cup G_{12}^*\cup G_{13}\), \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable
Remark 3.7
Our results on model (1.4) for the small drift rate case are summarized in Fig. 3. We have proved that competitive exclusion appears if \((d_2, q_2)\) falls into the blue and yellow regions of Fig. 3.
Next, we consider the large drift case, i.e., \(q_1>{{\overline{q}}}\).
Theorem 3.8
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\) with \(q_1>{{\overline{q}}}\). Then, the following statements hold:
-
(i)
If \((d_2,q_2)\in G_{21}\cup G_{22}\cup G_{23}^*\), then the semitrivial equilibrium \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable;
-
(ii)
If \((d_2,q_2)\in G_{11}\cup G_{12}\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable.
Here, \(G_{23}^*\) is defined by
Remark 3.9
Our results on model (1.4) for the large drift rate case are summarized in Fig. 4. Different from the small drift rate case, the invasion curve is unbounded. Again, we are able to prove that competitive exclusion happens if \((d_2, q_2)\) falls into the blue and yellow regions of Fig. 4.
Then, we consider the intermediate drift case, i.e., \({\underline{q}}\le q_1\le {{\overline{q}}}\), and we have the following results on the global dynamics of model (1.4).
Theorem 3.10
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\) with \({\underline{q}}\le q_1\le {{\overline{q}}}\). Let \(G_{12}^*\) be defined by (3.9) and \(G_{23}^*\) be defined by (3.10). Then, the following statements hold:
-
(i)
If \((d_2,q_2)\in G_{21}\cup G_{23}^*\), then the semitrivial equilibrium \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable;
-
(ii)
If \((d_2,q_2)\in G_{11}\cup G^*_{12}\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable.

Illustration of the results for the case \(q_1>{\overline{q}}\). If \((d_2, q_2)\) is above the curve \(q=q_{\varvec{u}}^*(d)\), then \((\varvec{u}^*,\varvec{0})\) is stable; and if \((d_2, q_2)\) is under the curve, then \((\varvec{u}^*,\varvec{0})\) is unstable. If \((d_2,q_2)\in G_{21}\cup G_{22}\cup G_{23}^*\), \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable; and if \((d_2,q_2)\in G_{11}\cup G_{12}\), \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable

Illustration of the results for the case \({\underline{q}}\le q_1\le {\overline{q}}\). If \((d_2,q_2)\in G_{21}\cup G_{23}^*\), \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable; and if \((d_2,q_2)\in G_{11}\cup G^*_{12}\), \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable
Remark 3.11
Our results on model (1.4) for the intermediate drift rate case are summarized in Fig. 5. In this case, the invasion curve may be defined on either a bounded or an unbounded interval. However, we know that it must locate between the yellow and blue regions in Fig. 5, where competitive exclusion happens.
In view of Theorems 3.6, 3.8, and 3.10, the global dynamics of model (1.4) in \(G_{11}\cup G_{21}\) is independent of \(q_1\):
Corollary 3.12
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, the following statements hold:
-
(i)
If \((d_2,q_2)\in G_{11}\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable;
-
(ii)
If \((d_2,q_2)\in G_{21}\), then the semitrivial equilibrium \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable.
More importantly, we have the following result about the evolution of random dispersal and directed drift rates.
Corollary 3.13
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, the following statements hold:
-
(i)
Fix \(d_1=d_2\). If \(q_1<q_2\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable; If \(q_1>q_2\), then the semitrivial equilibrium \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable;
-
(ii)
Fix \(q_1=q_2<{\underline{q}}\). If \(d_1<d_2\), then the semitrivial equilibrium \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable; If \(d_1>d_2\), then the semitrivial equilibrium \((\varvec{0}, \varvec{v}^*)\) is globally asymptotically stable;
-
(iii)
Fix \(q_1=q_2>{{\overline{q}}}\). If \(d_1<d_2\), then the semitrivial equilibrium \((\varvec{0}, \varvec{v}^*)\) is globally asymptotically stable; If \(d_1>d_2\), then the semitrivial equilibrium \((\varvec{u}^*, \varvec{0})\) is globally asymptotically stable.
Remark 3.14
By Corollary 3.13, the species with a smaller drift rate tends to have competitive advantage. If the drift rate is small, the species with smaller random dispersal rate has competitive advantage; if the drift rate is large, larger random dispersal rate is favored. We remark that Corollary 3.13 (i) was proved in [37] for the PDE case, and the corresponding results of 3.13 (ii)–(iii) for the PDE case in [37] are as follows: if \(d_1>d_2\), then there exists \({{\overline{q}}}(d_1,d_2)\) (resp. \({\underline{q}}(d_1,d_2)\)) such that \((\varvec{u}^*, \varvec{0})\) (resp. \((\varvec{0}, \varvec{v}^*)\)) is globally asymptotically stable for \(q_1=q_2>{{\overline{q}}}(d_1,d_2)\) (resp. \(q_1=q_2<{\underline{q}}(d_1,d_2)\)).
3.3 Coexistence and bistability
If \((d_2, q_2)\) is in the blank regions of Figs. 3, 4 and 5, we show that bistability and coexistence may occur. To this end, we explore the stability/instability of the semitrivial equilibrium \((\varvec{0},\varvec{v}^*(d_2,q_2))\) along the invasion curve \(q_2=q_{\varvec{u}}^*(d_2)\). Let
Then, \({\hat{\lambda }}_1(d_1)=0\), the semitrivial equilibrium \((\varvec{0},\varvec{v}^*(d_2,q^*_{\varvec{u}}(d_2)))\) is stable if \({\hat{\lambda }}_1(d_2)<0\), and \((\varvec{0},\varvec{v}^*(d_2,q^*_{\varvec{u}}(d_2)))\) is unstable if \({\hat{\lambda }}_1(d_2)>0\). The following result for the large drift case can be proved similarly as [7, Theorem 5.4], so we omit the proof here.
Theorem 3.15
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\) with \(q_1>{{\overline{q}}}\). Let \(q=q_{\varvec{u}}^*(d): (0, \infty )\rightarrow {\mathbb {R}}_+\) be defined in Theorem 3.1 and Proposition 3.4 (iii). Then, for any \(d_2>0\), the following statements hold:
-
(i)
If \( {\hat{\lambda }}_1(d_2)<0\), then
$$\begin{aligned} {{\hat{q}}}(d_2):=\inf \left\{ q>0: \ q>q_{\varvec{u}}^*(d_2)\ \text {and} \ \lambda _1\left( d_1,q_1,\varvec{1}-\frac{{\varvec{v}^*}\left( d_2,q\right) }{\varvec{k}}\right) \ge 0\right\} \end{aligned}$$exists and satisfies
$$\begin{aligned} {\left\{ \begin{array}{ll} {{\hat{q}}}(d_2)\in \left( q_{\varvec{u}}^*(d_2),q_1 \right) \;\;&{}\text {for}\;\; d_2<d_1,\\ {{\hat{q}}}(d_2)\in \left( q_{\varvec{u}}^*(d_2),\displaystyle \frac{q_1}{d_1}d_2\right) \;\;&{}\text {for}\;\; d_2>d_1. \end{array}\right. } \end{aligned}$$(3.12)Moreover, for any \(q_2\in (q_{\varvec{u}}^*(d_2), {{\hat{q}}}(d_2))\), both semitrivial equilibria \((\varvec{u}^*,\varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\) are locally asymptotically stable and model (1.4) admits an unstable positive equilibrium.
-
(ii)
If \( {\hat{\lambda }}_1(d_2)>0\), then
$$\begin{aligned} {{\hat{q}}}(d_2):=\sup \left\{ q>0:\ q<q_{\varvec{u}}^*(d_2)\ \text {and}\ \lambda _1\left( d_1,q_1,\varvec{1}-\frac{{\varvec{v}^*}\left( d_2,q\right) }{\varvec{k}}\right) \le 0\right\} \end{aligned}$$exists and satisfies
$$\begin{aligned} {\left\{ \begin{array}{ll} {{\hat{q}}}(d_2)\in \left( \displaystyle \frac{q_1}{d_1}d_2, q_{\varvec{u}}^*(d_2)\right) \;\; &{}\text {for}\;\; d_2<d_1,\\ {{\hat{q}}}(d_2)\in \left( q_1, q_{\varvec{u}}^*(d_2)\right) \;\;&{}\text {for}\;\; d_2>d_1. \end{array}\right. } \end{aligned}$$Moreover, for any \(q_2\in ({{\hat{q}}}(d_2),q_{\varvec{u}}^*(d_2))\), both semitrivial equilibria \((\varvec{u}^*,\varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\) are unstable and model (1.4) admits a stable positive equilibrium.
Remark 3.16
In (ii) when both semitrivial equilibria are unstable, we may conclude that the solutions are uniform persistent. If \({\underline{q}}\le q_1\le {{\overline{q}}}\) (the intermediate drift case), Theorem 3.15 (i)–(ii) holds for any \(d_2<d_1\), and we omit the statement to save space here.
The small drift rate case will be handled slightly different from the large drift rate case. For any \(\theta >0\), by Lemma 2.1 and Proposition 3.3 (ii), the line \(q=d\theta \) and the invasion curve \(q=q_{\varvec{u}}^*(d)\) have exactly one intersection point \((d^*(\theta ), d^*(\theta )\theta )\). So we can reparameterize the invasion curve as follows:
Let
Then, the semitrivial equilibrium \((\varvec{0},\varvec{v}^*\left( d^*(\theta ),q^*(\theta )\right) )\) is stable if \({\tilde{\lambda }}_1(\theta )<0\) and unstable if \({\tilde{\lambda }}_1(\theta )>0\). Noticing that \(q_{\varvec{u}}^*(d_1)=q_1\), we have \(d^*(q_1/d_1)=d_1\) and
Theorem 3.17
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\) with \(0<q_1<{\underline{q}}\). Then, for any \(\theta >0\), the following statements hold:
-
(i)
If \( {\tilde{\lambda }}_1(\theta )<0\), then
$$\begin{aligned} {{\tilde{d}}}^*(\theta ):=\inf \left\{ d>0: \ d>d^*(\theta )\ \text {and} \ \lambda _1\left( d_1,q_1,\varvec{1}-\frac{{\varvec{v}^*}\left( d,d\theta \right) }{\varvec{k}}\right) \ge 0\right\} \end{aligned}$$exists with \(d^*(\theta )<{{\tilde{d}}}^*(\theta )\) such that for any \((d_2,q_2)\) with \(q_2=d_2\theta \) and \(d^*(\theta )<d_2<{{\tilde{d}}}^*(\theta )\) both semitrivial equilibria \((\varvec{u}^*,\varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\) are locally asymptotically stable and model (1.4) admits an unstable positive equilibrium.
-
(ii)
If \( {\tilde{\lambda }}_1(\theta )>0\), then
$$\begin{aligned} {{\tilde{d}}}^*(\theta ):=\sup \left\{ d>0: \ d<d^*(\theta )\ \text {and} \ \lambda _1\left( d_1,q_1,\varvec{1}-\frac{{\varvec{v}^*}\left( d, d\theta \right) }{\varvec{k}}\right) \le 0\right\} \end{aligned}$$exists with \({{\tilde{d}}}^*(\theta )<d^*(\theta )\) such that for any \((d_2,q_2)\) with \(q_2=d_2\theta \) and \({{\tilde{d}}}^*(\theta )<d_2< d^*(\theta )\) both semitrivial equilibria \((\varvec{u}^*,\varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\) are unstable and model (1.4) admits a stable positive equilibrium.
Moreover, \({{\tilde{d}}}^*(\theta )\) satisfies
Proof
We prove (i), and (ii) can be proved similarly. Fix \(\theta >0\). Suppose \( {\tilde{\lambda }}_1(\theta )<0\). Let
By Theorem 3.6 (ii), \((\varvec{0},\varvec{v}^*)\) is unstable or neutrally stable if \((d_2, q_2)\in G_{11}\cup G_{13}\), which yields \(A\ne \emptyset \). Since \( {\tilde{\lambda }}_1(\theta )<0\), there exists \(\epsilon _0>0\) such that
Therefore, \({{\tilde{d}}}^*(\theta )\) exists with \(d^*(\theta )<{{\tilde{d}}}^*(\theta )\).
If \((d_2,q_2)\) satisfies \(q_2=d_2\theta \) and \({{\tilde{d}}}^*(\theta )<d_2< d^*(\theta )\), by the definition of \({{\tilde{d}}}^*(\theta )\), we have
which means that \((\varvec{0}, \varvec{v}^*)\) is locally asymptotically stable. By Theorem 3.1, \((\varvec{u}^*,\varvec{0})\) is also locally asymptotically stable. By the monotone dynamical system theory [20, 21, 46], model (1.4) admits an unstable positive equilibrium. Finally, it is easy to see that (3.15) holds by Theorem 3.6. \(\square \)
4 Discussions and numerical simulations
In this section, we discuss the results of the paper and present some numerical simulations.
4.1 Impact of spatial heterogeneity
If the environment is homogeneous, i.e., assumption (H) is replaced by \(k_1=k_2=k_3\), model (1.4) with n patches has been investigated in our recent paper [7]. The main results in [7] are summarized in Fig. 6. In particular, we prove that the invasion curve is between the lines \(q=q_1\) and \(q=q_1d/d_1\), \((\varvec{u}^*, \varvec{0})\) is globally asymptotically stable in \(G_1\), and \((\varvec{0}, \varvec{v}^*)\) is globally asymptotically stable in \(G_2\). These results are independent of the magnitude of drift rate \(q_1\) and are similar to the large drift rate case in this paper. Biologically, the downstream end is crowded due to the drift and thereby less friendly compared with the upstream end. If the environment perturbs from being uniformly distributed and the upstream locations become advantageous, e.g., assumption (H) holds, then a larger drift rate may compensate for it. This may explain why the homogeneous environment case is similar to the larger drift case in this paper.

Illustration of the main results for (1.4) with \(k_1=k_2=k_3\) in [7]. The blue cure is the invasion curve, which always lies between the lines \(q=q_1\) and \(q=q_1d/d_1\). Moreover, \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable if \((d_2, q_2)\in G_1\), and \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable if \((d_2, q_2)\in G_2\)
4.2 Impact of drift rate
By Propositions 3.4 and 3.13, if the drift rate \(q_1\) is small (\(q_1<{\underline{q}}\)), the invasion curve \(q=q^*_{\varvec{u}}(d)\) is defined on a bounded interval and the species with a smaller random dispersal rate is advantageous; if \(q_1\) is large (\(q_1>{{\overline{q}}}\)), the invasion curve is unbounded with a slant asymptote \(q=\theta d\) for some \(\theta >0\) and larger random dispersal rate is favored. The results for the small drift rate case align with the ones in the seminal works [15, 19], which claim that the species with a smaller random dispersal rate will always out-compete the other one in a spatial heterogeneous environment, when both species randomly move in space and are different only by the movement rate. When the drift rate becomes large, the outcomes of the competition change dramatically, and the species with a larger dispersal rate may win the competition.

The invasion curve for different values of \(q_1\). Here, \(\varvec{k}=(5, 3, 1)\), \(\varvec{r}=(1, 2, 1)\), and \(d_1=1\). The threshold values for the drift rates are \({\underline{q}}=0.4\) and \({{\overline{q}}}=2\)
We numerically explore the impact of the drift rate \(q_1\) on the shape of the invasion curve \(q=q^*_{\varvec{u}}(d)\). Fix \(\varvec{k}=(5, 3, 1)\), \(\varvec{r}=(1, 2, 1)\), and \(d_1=1\). Then, we can compute the threshold values for the drift rates: \({\underline{q}}=0.4\) and \({{\overline{q}}}=2\). In Fig. 7, we plot the invasion curves for \(q_1=0.2, 0.5, 1.2, 4\). If \(q_1=0.2\) or 0.5, the invasion curves seem to be bounded with \(\partial _d q^*_{\varvec{u}}(d_1)<0\), which indicates that a smaller random dispersal rate is favored when \(q_1=q_2\) and \(d_1\approx d_2\). In sharp contrast, if \(d_1=1.2\) or 4, the invasion curves seem to be unbounded with \(\partial _d q^*_{\varvec{u}}(d_1)>0\). This simulation also shows that the invasion curve can be bounded or unbounded for the intermediate drift case (\({\underline{q}}<q_1<{\overline{q}}\)).
4.3 Bistability and coexistence phenomena
Let \(d_1=1, q_1=1.5\), \(\varvec{r}=(3,7,3)\), and \(\varvec{k}=(5,3,1)\). We graph the invasion curve \((d^*(\theta ), q^*(\theta ))\) and \({\tilde{\lambda }}(\theta )\) in Fig. 8. The stability of \((\varvec{0}, \varvec{v}^*)\) when \((d_2, q_2)=(d^*(\theta ), q^*(\theta ))\) is determined by the sign of \({\tilde{\lambda }}(\theta )\). In Fig. 8, we can see that \({\tilde{\lambda }}(\theta )\) changes sign, which means that both bistability and coexistence are possible. Indeed, if we choose \((d_2, q_2)=(3.088, 1.239)\), which is slightly below the invasion curve, then both \((\varvec{u}^*, \varvec{0})\) and \((\varvec{0}, \varvec{v}^*)\) are locally asymptotically stable. As shown in Fig. 9, if the initial data are \((\varvec{u}(0),\varvec{v}(0))=((0.1, 0.1, 0.1), (5, 5, 5))\), then the solution of (1.4) converges to \((\varvec{0}, \varvec{v}^*)\); if the initial data are \((\varvec{u}(0),\varvec{v}(0))=((5, 5, 5), (0.1, 0.1, 0.1))\), then the solution converges to \((\varvec{u}^*, \varvec{0})\). Finally, we choose \((d_2, q_2)=(10.28, 0.03)\), which is slightly above the invasion curve (\((\varvec{u}^*, \varvec{0})\) is unstable). Since \({\tilde{\lambda }}\) is positive, \((\varvec{0}, \varvec{v}^*)\) is also unstable, and the model has at least one stable positive equilibrium. We graph the solution of (1.4) for initial data \((\varvec{u}(0), \varvec{v}(0))=((5, 5, 5), (5, 5, 5))\) and the solution seems to converge to a positive equilibrium, see Fig. 10.

a The invasion curve \((d^*(\theta ), q^*(\theta ))\) when \(d_1=1\), \(q_1=1.5\), \(\varvec{r}=(3,7,3)\), and \(\varvec{k}=(5,3,1)\). b The sign of the curve \(\tilde{\lambda }_1(\theta )\) determines the stability of \((\varvec{0},\varvec{v}^*)\) when \((d_2, q_2)=(d^*(\theta ), q^*(\theta ))\)

Solutions of (1.4) with \(d_1=1\), \(q_1=1.5\), \(d_2=3.088\), \(q_2=1.239\), \(\varvec{r}=(3,7,3)\), \(\varvec{k}=(5,3,1)\). For a, b, the initial data are \(\varvec{u}(0)=(0.1,0.1,0.1)\) and \(\varvec{v}(0)=(5,5,5)\), and species \(\varvec{v}\) wins the competition; for c, d, the initial data are \(\varvec{u}(0)=(5,5,5)\) and \(\varvec{v}(0)=(0.1,0.1,0.1)\), and species \(\varvec{u}\) wins the competition

Solution of (1.4) with \(d_1=1\), \(q_1=1.5\), \(d_2=10.28\), \(q_2=0.03\), \(\varvec{r}=(3,7,3)\), \(\varvec{k}=(5,3,1)\), and the initial data are \(\varvec{u}(0)=(5,5,5)\) and \(\varvec{v}(0)=(5,5,5)\). The two species seem to coexist
4.4 Evolutionarily singular strategies
We formulate a conjecture based on Corollary 3.13 about the existence of an evolutionarily stable strategy (ESS) for the diffusion rate, which may distinguish the 2-patch model from the 3-patch model.
We fix \(q_2=q_1>0\) and view the diffusion rate as an evolutionary strategy of the species. By Corollary 3.13 when \({\underline{q}}<{{\overline{q}}}\), we conjecture that there exists \({\underline{q}}\le q_*< q^*\le {{\overline{q}}}\) such that if \(q<{\underline{q}}\), then the slower diffuser always wins the competition; if \(q>q_*\), then the faster diffuser prevails; if \(q_1\in (q_*, q^*)\), there exists a unique \(d^*(q_1)>0\) such that \(d_1=d^*(q_1)\) is an evolutionarily singular strategy with the asymptotic limits:
or
Moreover, we conjecture that the singular strategy is an ESS in the former case but not in the latter case (Fig. 11).
We provide some numerical evidence to support the conjecture below. Since
the sign of
is crucial to determine which strategy is favored when \(d_2\approx d_1\): if \({\mathcal {S}}(d_1,q_1)<0\) the slow diffuser is favored; if \({\mathcal {S}}(d_1,q_1)>0\) the faster diffuser is favored. By Corollary 3.13, \({\mathcal {S}}(d_1,q_1)\) changes signs. In particular, if \(q_1<{\underline{q}}\), \({\mathcal {S}}(d_1,q_1)<0\) and if \(q_1>{\underline{q}}\), \({\mathcal {S}}(d_1,q_1)>0\). We numerically solve \({\mathcal {S}}(d_1,q_1)=0\) and plot the solution in Fig. 11, which consists with a curve \(d_1=d^*(q_1)\), where \(q_1\in (q_*, q^*)\). In the left figure, the sign of \({\mathcal {S}}(d_1,q_1)\) changes from negative to positive when moving from above to below the curve. This suggests that the diffusion rate \(d_1=d^*\) may be an ESS for \(q\in (q_*, q^*)\). In the right figure, opposite phenomenon appears when crossing the curve and we suspect that the singular strategy is not an ESS in this case.

The sign of \({\mathcal {S}}(d_1,q_1)\), where \({\mathcal {S}}(d_1,q_1)<0\) in the dark blue region and \({\mathcal {S}}(d_1,q_1)>0\) in the light blue region. Left figure: \(\varvec{r}=(3,7,3)\), \(\varvec{k}=(5,3,1)\); right F \(\varvec{r}=(50,3,3)\), \(\varvec{k}=(5,3,1)\)
We remark such an intermediate diffusion rate \(d_1=d^*\) as an ESS does not appear in the corresponding 2-patch model. For the 2-patch model, as proved in [18, 45] (see [23, Theorem 1]), there exists a critical value \(q_*\) such that if \(q_1=q_2<q_*\) then \(d_1=0\) is an ESS; if \(q_1=q_2>q_*\) then \(d_1=\infty \) is an ESS. We also note that if \({\underline{q}}={{\overline{q}}}\) in the 3-patch model, then the curve \(d_1=d^*(q_1)\) is a vertical line and an intermediate ESS also does not exists (which is similar to the 2-patch case).
Finally, we conjecture that the results in this paper hold for the N-patch model. Our results for 3-patch model are based on the monotonicity of the semitrivial equilibrium (see Lemma 5.1 (iii)–(iv)), which we cannot prove for the N-patch model. Similarly, if the movement rates among patches are not homogeneous (i.e., the off-diagonal entries of D and Q are not 1 s), it is also not trivial to show how the movement rates affect the monotonicity of the semitrivial equilibrium.
5 Proofs for the invasion curve
In this section, we present the proofs of the results on the invasion curve \(q_{\varvec{u}}^*(d)\). We begin with an analysis on \(\varvec{u}^*\). A similar result of the following lemma when \(r_1=r_2=r_3\) except for the sign of \(\sum _{i=1}^3r_i\left( 1-{u^*_i}/{k_i}\right) \) can be found in [23].
Lemma 5.1
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), \(d_1>0\), and \(q_1\ge 0\). Then, the following statements on \(\varvec{u}^*\) hold:
-
(i)
\(d_1u_{i+1}^*-(d_1+q_1)u_i^*<0\) for \(i=1,2\);
-
(ii)
\(u^*_1<k_1\) and \(u^*_3>k_3\);
-
(iii)
If \(q_1>{{\overline{q}}}\), then \(u^*_1<u^*_2<u^*_3\) and \(\sum _{i=1}^3r_i\left( 1-\frac{u^*_i}{k_i}\right) >0\);
-
(iv)
If \(q_1<{\underline{q}}\), then \(u^*_1>u^*_2>u^*_3\) and \(\sum _{i=1}^3r_i\left( 1-\frac{u^*_i}{k_i}\right) <0\).
Proof
By (1.4), we have
Suppose to the contrary that \(d_1u^*_2-(d_1+q_1)u^*_1\ge 0\). Then, by the first equation of (5.1), we have \(u_2^*\ge u_1^*\ge k_1\). This, together with assumption \((\textbf{H})\) and the second equation of (5.1), implies that \(d_1u_3^*-(d_1+q_1)u^*_2>0\) and \(u_3^*>k_3\), which contradicts the third equation of (5.1). Therefore, we have \(d_1u^*_2-(d_1+q_1)u^*_1<0\). Similarly, we can prove \(d_1u_3^*-(d_1+q_1)u^*_2<0\). This proves (i). By (i) and the first and third equations of (5.1), we can easily obtain (ii).
The proof of (iv) is similar to that of (iii), so we only prove (iii) here. Suppose \(q_1>{{\overline{q}}}\). We rewrite (5.1) as follows:
Suppose to the contrary that \(u^*_1\ge u^*_2\). Then, by the first equation of (5.2), we have \(k_1-\displaystyle \frac{q_1k_1}{r_1}-u^*_1\ge 0\). Since \(q_1>\displaystyle \frac{r_1}{k_1}(k_1-k_2)\), we obtain
Then, by the second equation of (5.2), we get \(u^*_2>u^*_3\). This, combined with \(q_1>\displaystyle \frac{r_3}{k_3}(k_2-k_3)\), yields
which contradicts the last equation of (5.2). This proves \(u^*_1< u^*_2\).
Suppose to the contrary that \(u^*_2\ge u^*_3\). Then, by the last equation of (5.2), we have \(u_3^*\ge k_3\left( 1+\displaystyle \frac{q_1}{r_3}\right) \). By \(q_1>{{\overline{q}}}\), we obtain
Then, by the second equation of (5.2), we have \(u_1^*>u_2^*\). By the first equation of (5.2), we get
which is a contradiction. This proves \(u_2^*<u_3^*\).
Dividing the ith equation of (5.2) by \(u_i^*\), we have
Adding up the equations in (5.3), we obtain
Then, by (i) and \(u_1^*<u_2^*<u_3^*\), we have \(\sum _{i=1}^3r_i\left( 1-\frac{u^*_i}{k_i}\right) >0\). This proves (iii). \(\square \)
5.1 Proof of Theorem 3.1
We prove the existence of the invasion curve \(q=q_{\varvec{u}}^*(d)\) in this subsection.
Lemma 5.2
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, the following statements hold about the semitrivial equilibrium \((\varvec{u}^*, \varvec{0})\) of (1.4):
-
(i)
If \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) \ge 0\), then for any \(d>0\) there exists \(q_{\varvec{u}}^*(d)>0\) such that \(\lambda _1\left( d,q_{\varvec{u}}^*(d), \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\), \(\lambda _1\left( d,q, \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) <0\) for all \(q>q_{\varvec{u}}^*(d)\), and \(\lambda _1\left( d,q, \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) >0\) for all \(q<q_{\varvec{u}}^*(d)\);
-
(ii)
If \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) < 0\), then there exists \(d_0>0\) such that \(\lambda _1(d_0,0,\varvec{1}-{\varvec{u}^*}/{\varvec{k}})=0\), \(\lambda _1(d,0,\varvec{1}-{\varvec{u}^*}/{\varvec{k}})<0\) for all \(d>d_0\), and \(\lambda _1(d,0,\varvec{1}-{\varvec{u}^*}/{\varvec{k}})>0\) for all \(d<d_0\). Moreover, the following results hold:
- \({\mathrm{(ii_1)}}\):
-
For any \(d\in (0,d_0)\), there exists \(q_{\varvec{u}}^*(d)>0\) such that the statement in \(\mathrm{(i)}\) holds;
- \({\mathrm{(ii_2)}}\):
-
For any \(d\in [d_0, \infty )\), we have \(\lambda _1(d,q,\varvec{1}-{\varvec{u}^*}/{\varvec{k}})< 0\) for all \(q>0\).
Proof
For simplicity, we denote \(\lambda _1(d,q):=\lambda _1\left( d,q,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) \). An essential step of the proof is to show the following claim.
Claim 1: Fixing \(d>0\), equation \(\lambda _1(d, q)=0\) has at most one root for \(q\in [0, \infty )\).
Proof of Claim: Let \(\varvec{\psi }\) be the positive eigenvector corresponding to \(\lambda _1(d,q)\) with \(\sum _{i=1}^3\psi _i=1\). Then, we have
Differentiating (5.5) with respect to q and denoting \('=\partial /\partial q\), we obtain
Multiplying (5.5) by \(\psi _i'\) and (5.6) by \(\psi _i\) and taking the difference of them, we have
Motivated by [7, Eq. (3.7)], we introduce \((\beta _1,\beta _2,\beta _3)=\left( 1,{d}/(d+q), d^2/(d+q)^2\right) \). Multiplying (5.7) by \(\beta _i\) and summing up in i, we obtain
Suppose \(\lambda _1\left( d,{{\tilde{q}}}\right) =0\) for some \({{\tilde{q}}}\ge 0\). By Lemma 5.1 (ii), we see that
Therefore, by (5.8), we have \(\lambda '(d, {{\tilde{q}}})<0\). This proves the claim.
According to the claim, whether the equation \(\lambda _1(d, q)=0\) has a root in q is determined by the sign of \(\lambda _1(d, 0)\) and \(\lim _{q\rightarrow \infty }\lambda _1(d,q)\).
Claim 2: \(\lim _{q\rightarrow \infty }\lambda _1(d,q)<0\).
Proof of claim: Adding up all the equations of (5.5), we have
which implies that \(\lambda _1(d,q)\) is bounded for \(d,q>0\). So up to a subsequence, we may assume \(\lim _{q\rightarrow \infty }\varvec{\psi }=\tilde{\varvec{\psi }}\). Dividing (5.5) by q and taking \(q\rightarrow \infty \), we obtain
which yields \(\tilde{\varvec{\psi }}=(0,0,1)^T\). Then, taking \(q\rightarrow \infty \) in (5.9), we have
where we have used Lemma 5.1 (ii) in the last step. This proves the claim.
By Lemma 2.1, \(\lambda _1(d, 0)\) is strictly decreasing in d with
where we have used Lemma 5.1 (ii) again to see that \(1-u_1^*/k_1>0\). Now, the desired results follow from this and Claims 1 and 2. \(\square \)
We are ready to prove Theorem 3.1.
Proof of Theorem 3.1
Let \(d_0\) be defined in Lemma 5.2, \(d^*\) be defined by (3.2), and \(q=q^*_{\varvec{u}}(d): (0, d^*)\rightarrow {\mathbb {R}}_+\) be defined in Lemma 5.2. Then, Theorem 3.1 (i)–(ii) follows from Lemmas 5.1–5.2 and the fact that the stability/instability of \((\varvec{u}^*, \varvec{0})\) is determined by the sign of \(\lambda _1\left( d,q, \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) \). The continuity of \(q=q^*_{\varvec{u}}(d)\) follows from \(\partial _q \lambda _1\left( d,q^*_{\varvec{u}}(d), \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) <0\) (Claim 1 of Lemma 5.2) and the implicit function theorem. \(\square \)
5.2 Proof of Propositions 3.3 and 3.4
First, we prove the following useful result:
Lemma 5.3
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, we have
if one of the following conditions holds:
-
(i)
\((d_2,q_2)\in G_{11}\cup G_{21}\);
-
(ii)
\(q_1>{{\overline{q}}}\) and \((d_2,q_2)\in G_{12}\cup G_{22}\);
-
(iii)
\(q_1<{\underline{q}}\) and \((d_2,q_2)\in G_{13}\cup G_{23}\).
Proof
Suppose to the contrary that \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\), and let \(\varvec{\phi }\gg 0\) be a corresponding eigenvector. Note that \(\lambda _1\left( d_1,q_1,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\) with a corresponding eigenvector \(\varvec{u}^*\). Let
and
Then, we have
Using similar arguments as in [7, Lemma 5.7], we can show
and
Indeed, multiplying (5.11a) by \(\left( \displaystyle \frac{d_1}{d_1+q_1}\right) ^j\phi _j\), and summing up from \(j=1\) to \(j=3\), we have
where we have used \({{\tilde{f}}}_3={{\tilde{f}}}_0=0\) in the last step. Similarly, multiplying (5.11b) by \(\left( \displaystyle \frac{d_1}{d_1+q_1}\right) ^ju^*_j\) and summing up from \(j=1\) to \(j=3\), we obtain
Taking the difference of (5.14) and (5.15), we obtain (5.12). Similarly, we can prove (5.13).
By Lemma 5.1 (i)–(ii) and (5.11b), we have \({{\tilde{f}}}_j, {{\tilde{g}}}_j<0\) for \(j=1,2\). Now we obtain a contradiction for each of (i)–(iii).
(i) We only consider the case \((d_2,q_2)\in G_{21}\), since the case \((d_2,q_2)\in G_{11}\) can be studied similarly. Suppose \((d_2,q_2)\in G_{21}\). Then, we have \(d_2\le d_1, q_2\le q_1d_2/d_1\), and \((d_1, q_1)\ne (d_2, q_2)\). If \(d_1\ne d_2\), then it is easy to check that
(This inequality can be found in [53, Lemma 2.4].) This, together with \({{\tilde{g}}}_1,{{\tilde{g}}}_2<0\), yields
If \(d_1=d_2\), then \(q_1>q_2\) and (5.16) also holds. Then, by \({{\tilde{f}}}_1,{{\tilde{f}}}_2<0\) and (5.12), we have
which is a contradiction.
(ii) We only need to obtain a contradiction for the case \((d_2,q_2)\in G_{22}\), since the case \((d_2, q_2)\in G_{12}\) can be studied similarly. Suppose \((d_2,q_2)\in G_{22}\). Then, we have \(d_2>d_1\) and \(q_2\le q_1\). By Lemma 5.1 (i), we have \(u_1^*<u_2^*<u^*_3\), which implies that
This, combined with \({{\tilde{g}}}_1, {{\tilde{g}}}_2<0\) and (5.13), gives a contradiction.
(iii) We only obtain a contradiction for the case \((d_2,q_2)\in G_{23}\), since the case \((d_2,q_2)\in G_{13}\) can be studied similarly. Suppose \((d_2,q_2)\in G_{23}\). Then, we have \(d_2\le d_1\), \(q_2\le q_1\), and \((d_1, q_1)\ne (d_2, q_2)\). By Lemma 5.1 (ii), we have \(u_1^*>u_2^*>u^*_3\), which implies that
This combined with \({{\tilde{g}}}_1, {{\tilde{g}}}_2<0\) and (5.13) gives a contradiction. \(\square \)
We are ready to prove Propositions 3.3 and 3.4.
Proof of Proposition 3.3
(i) We only prove the case \(G_{21}\subset S_2\), (i.e., \((\varvec{u}^*,\varvec{0})\) is unstable for \((d_2,q_2)\in G_{21}\)), since the case \( G_{11}\subset S_1\) can be proved similarly. To avoid confusion, we denote \(\varvec{u}^*\) by \(\varvec{u}^*_{\varvec{k}}\). It is easy to see that \(\varvec{u}^*_{\varvec{k}}\) depends continuously on \(\varvec{k}\).
Suppose \((d_2, q_2)\in G_{21}\). We need to prove that \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}}}/{\varvec{k}}\right) >0\). Suppose to the contrary that \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}}}/{\varvec{k}}\right) \le 0\). By Lemma 5.3, we must have \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}}}/{\varvec{k}}\right) < 0\). By [7, Theorem 4.2], we have \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}'}}/{\varvec{k}'}\right) >0\), where \(\varvec{k}'=(k_3,k_3,k_3)\).
Let \(\Lambda (s):=\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}_1(s)}}/{\varvec{k}_1(s)}\right) \), where \(\varvec{k}_1(s)=s\varvec{k}+(1-s)\varvec{k}'\) satisfies (H) for any \(s\in [0, 1]\). Since
there exists \(s_0\in (0,1)\) such that \(\Lambda (s_0)=0\), which contradicts Lemma 5.3.
(ii) By [7, Theorem 4.2], if \((d_2,q_2)\in G_{22}\), then \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}'}}/{\varvec{k}'}\right) >0\); and if \((d_2,q_2)\in G_{12}\), then \(\lambda _1\left( d_2,q_2,\varvec{1}-{\varvec{u}^*_{\varvec{k}'}}/{\varvec{k}'}\right) <0\), where \(\varvec{k}'=(k_3,k_3,k_3)\). Then, using similar arguments as (i), we can prove (ii).
(iii) Let \(\varvec{\psi }\) be the positive eigenvector corresponding to \(\lambda _1:=\lambda _1\left( d_2,q_1,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) \) with \(\sum _{i=1}^3\psi _i=1\). Then, we have
Differentiating (5.5) with respect to \(d_2\) and denoting \('=\partial /\partial d_2\), we obtain
Multiplying (5.17) by \(\psi _i'\) and (5.18) by \(\psi _i\) and taking the difference of them, we have
Similar to the proof of Lemma 5.2, let \((\beta _1,\beta _2,\beta _3)=\left( 1,{d_2}/(d_2+q_1), d_2^2/(d_2+q_1)^2\right) \). Multiplying (5.19) by \(\beta _i\) and adding up them in i, we obtain
Note that \(\lambda _1\left( d_1,q_1,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\) with a corresponding eigenvector \((u_1^*, u^*_2, u_3^*)^T\). Moreover, by Lemma 5.1, we have \(d_1u_{i+1}^*-(d_1+q_1)u_i^*<0\) for \(i=1, 2\) and \(u_1^*>u^*_2>u_3^*\). Then, it follows from (5.20) that
This implies that \(\lambda _1\left( d_2,q_1,\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) >0\) if \(0<d_1-d_2\ll 1\) and \(\lambda _1\left( d_2,q_1,\varvec{1}-\frac{\varvec{u}^*}{\varvec{k}}\right) <0\) if \(0<d_2-d_1\ll 1\). Then, by Lemma 5.3, we have \(G_{13}\subset S_1\) and \(G_{23}\subset S_2\). \(\square \)
Remark 5.4
A similar inequality of (5.21) is proved in [23], and we include the proof for completeness here.
Proof of Proposition 3.4
For any \(0<d<d^*\), let \(\varvec{\psi }\) be the eigenvector corresponding to \(\lambda _1\left( d,q_{\varvec{u}}^*(d),\varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\) with \(\varvec{\psi }\gg 0\) and \(\sum _{i=1}^3\psi _i=1\). Then,
(i) Up to a subsequence, we may assume \(\lim _{d\rightarrow 0}\varvec{\psi }=\tilde{\varvec{\psi }}\) for some \(\tilde{\varvec{\psi }}\ge \varvec{0}\) and \(\sum _{i=1}^3{\tilde{\psi }}_i=1\). We first claim that \(q_{\varvec{u}}^*(d)\) is bounded for \(d\in (0,\delta )\) with \(\delta \ll 1\). If it is not true, then dividing (5.22) by \(q_{\varvec{u}}^*(d)\) and taking \(d\rightarrow 0\), we have
which yields \(\tilde{\varvec{\psi }}=(0,0,1)^T\). Adding up all the equations of (5.22) and taking \(d\rightarrow 0\) in (5.24), we have
and consequently, \(k_3-u_3^*=0\), which contradicts Lemma 5.1 (ii). This proves the claim. By the claim, up to a subsequence, we may assume \(\lim _{d\rightarrow 0}q^*(\theta )={{\tilde{q}}}_0\in [0,\infty )\). Consequently, for sufficiently small \(\epsilon >0\), there exists \({{\bar{d}}}>0\) such that \(q_{\varvec{u}}^*(d)<{{\tilde{q}}}_0+\epsilon \) for all \(0<d<{{\bar{d}}}\). It follows from Lemma 5.2 that
for all \(0<d<{{\bar{d}}}\). Hence,
Since \(k_3-u_3^*=0\) and \(\epsilon >0\) was arbitrary,
Therefore, we have \({{\tilde{q}}}_0\ge q_0>0\). Similarly, we can prove \({{\tilde{q}}}_0\le q_0\). This proves (i).
Now we prove (ii)–(iv). If we show that (3.7) holds when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) < 0\), (3.8) holds with \(\theta \in (0,q_1/d_1)\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) > 0\), and (3.8) holds with \(\theta =0\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) =0\). Then, (iv) holds and (ii)–(iii) follow from Theorem 5.1 (iii)–(iv).
By Lemma 5.2 (i), the function \(q=q_{\varvec{u}}^*(d)\) is defined for \(d\in (0,\infty )\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) \ge 0\). We claim that \({q_{\varvec{u}}^*(d)}/{d}\) is bounded for \(d\in (\delta ,\infty )\) for any fixed \(\delta >1\). If it is not true, up to a subsequence, we may assume
for some \(\hat{\varvec{\psi }}\ge \varvec{0}\) and \(\sum _{i=1}^3{\hat{\psi }}_i=1\). Then, dividing (5.22) by \(q_{\varvec{u}}^*(d)\) and taking \(d\rightarrow \infty \), we can obtain a contradiction using similar arguments as in the proof of (i). Therefore, \({q_{\varvec{u}}^*(d)}/{d}\) is bounded for \(d\in (\delta ,\infty )\). Then, using similar arguments as in the proof of [7, Proposition 4.4], we can show that (3.8) holds with \(\theta \in (0,q_1/d_1)\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) > 0\), and (3.8) holds with \(\theta =0\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) =0\).
By Lemma 5.2 (ii), the function \(q=q_{\varvec{u}}^*(d)\) is defined for \(d\in (0,d_0)\) when \(\sum _{i=1}^3 r_i\left( 1-{u_i^*}/{k_i}\right) < 0\). Using similar arguments as in (i), we can show that \(q_{\varvec{u}}^*(d)\) is bounded for \(d\in (d_0-\delta ,d_0)\) for some \(\delta \ll 1\). Then, up to a subsequence, we may assume
for some \({\varvec{\psi }^*}\ge \varvec{0}\) and \(\sum _{i=1}^3\psi ^*_i=1\). Taking \(d\rightarrow d_0\) in (5.22), we see that
which yields \(\lambda _1\left( d_0,\eta , \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\). By the proof of Lemma 5.2, \(\lambda _1\left( d_0,q, \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\) has at most one root for \(q\in [0,\infty )\). Since \(\lambda _1\left( d_0,0, \varvec{1}-{\varvec{u}^*}/{\varvec{k}}\right) =0\), we must have \(\eta =0\). This proves (3.7). \(\square \)
6 Proofs for the competitive exclusion results
Let \((\varvec{u},\varvec{v})\) be a positive equilibrium of model (1.4). Define
Clearly, we have
and
Then, we have the following result about the sign of \(f_j,g_j\), \(j=1,2\).
Lemma 6.1
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1, q_1, d_2, q_2>0\). If \((\varvec{u},\varvec{v})\) is a positive equilibrium of model (1.4), then we have \(f_1,g_1,f_2,g_2<0\).
Proof
First we prove \(f_1<0\). Suppose to the contrary that \(f_1\ge 0\). By (6.2) and (6.3), we have \(k_1-u_1-v_1\le 0\) and \(g_1\ge 0\). Since \(f_1,g_1\ge 0\), we have \(u_2>u_1\) and \(v_2>v_1\). This combined with \((\textbf{H})\) implies that \(k_2-u_2-v_2<0\). Then, by (6.2) and (6.3) again, we obtain that \(f_2,g_2>0\) and \(k_3-u_3-v_3<0\), which contradicts (6.2) with \(j=3\). Therefore, we have \(f_1<0\). Consequently, by (6.2) and (6.3) with \(j=1\), we have \(g_1<0\). Using similar arguments, we can prove \(f_2,g_2<0\). \(\square \)
The following result is similar to [7, Lemma 5.7] with \(j=1\) and \(j=n=3\) (see also the proof of Lemma 5.3). Thus, we omit the proof.
Lemma 6.2
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1, q_1, d_2, q_2>0\). If \((\varvec{u},\varvec{v})\) is a positive equilibrium of model (1.4), then the following equations hold:
and
An essential step to prove the competitive exclusion results for model (1.4) is to show the nonexistence of positive equilibrium:
Lemma 6.3
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Let \(G^*_{12}\) and \(G_{23}^*\) be defined by (3.9) and (3.10), respectively. Then, model (1.4) admits no positive equilibrium, if one of the following conditions holds:
-
(i)
\((d_2,q_2)\in G_{11}\cup G_{21}\);
-
(ii)
\(q_1>{{\overline{q}}}\) and \((d_2,q_2)\in G_{12}\cup G_{22}\cup G_{23}^*\);
-
(iii)
\(q_1<{\underline{q}}\) and \((d_2,q_2)\in G_{13}\cup G_{23}\cup G^*_{12}\).
-
(iv)
\({\underline{q}}\le q_1\le {{\overline{q}}}\) and \((d_2,q_2)\in G^*_{12}\cup G_{23}^*\);
Proof
Suppose to the contrary that model (1.4) admits a positive equilibrium \((\varvec{u},\varvec{v})\). Then, we will obtain a contradiction for each of the cases (i)–(iv).
(i) We only consider the case \((d_2,q_2)\in G_{21}\). Since the nonlinear terms of (1.4) are symmetric, the case \((d_2,q_2)\in G_{11}\) can be proved similarly. Suppose \((d_2,q_2)\in G_{21}\). Then, we have \(d_2\le d_1, q_2\le q_1d_2/d_1\) and \((d_1, q_1)\ne (d_2, q_2)\). First, we claim that
Indeed if \(d_1=d_2\), then \(q_1>q_2\) and (6.6) holds. If \(d_1>d_2\), then it is easy to check that
(This inequality is in [53, Lemma 2.4]). This, combined with \(g_1,g_2<0\), proves (6.6). Then, by \(f_1, f_2<0\) and (6.4), we have
which is a contradiction.
(ii) We first consider the case \((d_2,q_2)\in G_{22}\). Since the nonlinear terms of (1.4) are symmetric, the case \((d_2,q_2)\in G_{12}\) can be proved similarly. Suppose \((d_2, q_2)\in G_{22}\). Then, \(d_2>d_1\) and \(q_2\le q_1\). By (1.4), we have
Then, we show that \(u_1<u_2<u_3\). Suppose to the contrary that \(u_1\ge u_2\). Then, by (6.7a), we see that
where we have used \(q_2\le q_1\) in the last inequality. This, combined with (6.7b), implies that \(v_1\ge v_2\). Noticing that
we have
and consequently \(u_3<u_2\) and \(v_3<v_2\) by (6.7c) and (6.7d). This, combined with \(q_1>{{\overline{q}}}\), implies that
which contradicts (6.7e). Similarly, we can show that \(u_2<u_3\). So, \(u_1<u_2<u_3\), which leads to
Then, by Lemma 6.1 and (6.5), we have
which is a contradiction.
Now suppose that \((d_2,q_2)\in G_{23}^*\). Then, \(q_2<{\underline{q}}\) and \((d_1,q_1)\in {\hat{G}}_{13}\), where
Since the nonlinear terms of (1.4) are symmetric, this case can be proved similarly as the case \((d_2,q_2)\in G_{13}\) (the proof is immediately below).
(iii) Suppose that \((d_2,q_2)\in G_{13}\). Then, we have \(d_2\ge d_1\), \(q_1\le q_2\le q_1d_2/d_1\), and \((d_1, q_1)\ne (d_2, q_2)\). We show that \(u_1>u_2>u_3\). Suppose to the contrary that \(u_1\le u_2\). Then, by (6.7a) and \(q_2\ge q_1\), we have
This, combined with (6.7b), yields \(v_1\le v_2\). Noting that
we have
Then, by (6.7c) and (6.7d), we have \(u_3>u_2\) and \(v_3>v_2\). Since \(q_1<{\underline{q}}\), we have
which contradicts (6.7e). Similarly, we can show \(u_2>u_3\). Therefore, we have \(u_1>u_2>u_3\) and
Then, it follows from Lemma 6.1 and (6.5) that
which is a contradiction.
For the case \((d_2,q_2)\in G_{23}\), using similar arguments as above, we can obtain \(v_1>v_2>v_3\), which leads to
This, combined with Lemma 6.1 and (6.4), implies that
which is a contradiction.
Next suppose that \((d_2,q_2)\in G_{12}^*\). Then, \(q_2>{{\overline{q}}}\) and \((d_1,q_1)\in {\hat{G}}_{22}\), where
Since the nonlinear terms of (1.4) are symmetric, this case can be proved similarly the case \((d_2,q_2)\in G_{22}\) in (ii).
(iv) If \((d_2,q_2)\in G_{23}^*\), the proof is similar to the corresponding case in (ii). If \((d_2,q_2)\in G_{12}^*\), the proof is similar to the corresponding case in (iii). \(\square \)
We are ready to prove Theorems 3.6, 3.8 and 3.10.
Proof of Theorem 3.6
(i) Suppose that \(q_1<{\underline{q}}\) and \((d_2,q_2)\in G_{21}\cup G_{23}\). By Lemma 6.3 (i) and (iii), model (1.4) admits no positive equilibrium. By Theorem 3.3 (i) and (iii), \((\varvec{u}^*, \varvec{0})\) is unstable. Then, it follows from the monotone dynamical system theory [20, 21, 29, 46] that \((\varvec{u}^*,\varvec{0})\) is globally asymptotically stable.
(ii) Suppose that \(q_1<{\underline{q}}\) and \((d_2,q_2)\in G_{11}\cup G_{12}^*\). By Lemma 6.3 (i) and (iii), model (1.4) admits no positive equilibrium. By the monotone dynamical system theory [20, 21, 29, 46], it suffices to show that \((\varvec{0},\varvec{v}^*)\) is unstable. If \((d_1,q_1)\in G_{12}^*\), then \(q_2>{{\overline{q}}}\) and \((d_1,q_1)\in {{\tilde{G}}}_{22}:=\{(d,q):d>d_2,0<q\le q_2\}\). Since the nonlinear terms of model (1.4) are symmetric, it follows from Proposition 3.3 (ii) that \((\varvec{0},\varvec{v}^*)\) is unstable. If \((d_2,q_2)\in G_{11}\), then \((d_1,q_1)\in {{\tilde{G}}}_{21}\), where
Similarly, it follows from Proposition 3.3 (i) that \((\varvec{0},\varvec{v}^*)\) is unstable.
Finally, suppose that \(q_1<{\underline{q}}\) and \((d_2,q_2)\in G_{13}\). By Proposition 3.3 (iii), \((\varvec{u}^*, \varvec{0})\) is locally asymptotically stable. By Lemma 6.3 (iv), model (1.4) admits no positive equilibrium. If \((\varvec{0}, \varvec{v}^*)\) is locally asymptotically stable, then model (1.4) admits one unstable positive steady state, which is a contradiction. If \((\varvec{0}, \varvec{v}^*)\) is unstable, then the monotone dynamical system theory [20, 21, 29, 46] implies that \((\varvec{u}^*, \varvec{0})\) is globally asymptotically stable. If \((\varvec{0}, \varvec{v}^*)\) is neutrally stable, by [29, Theorem 1.4], \((\varvec{u}^*, \varvec{0})\) is globally asymptotically stable. This proves (ii). \(\square \)
Proof of Theorems 3.8 and 3.10
We only need to prove the case \((d_2,q_2)\in G_{23}^*\), since the other cases can be proved using similar arguments in the proof of Theorem 3.6. If \((d_2,q_2)\in G_{23}^*\), then \(q_2<{\underline{q}}\) and \((d_1,q_1)\in {{\hat{G}}}_{13}\), where
Since the nonlinear terms of model (1.4) are symmetric, it follows from Theorem 3.6 (ii) that \((\varvec{0},\varvec{v}^*)\) is globally asymptotically stable. \(\square \)
References
Altenberg, L.: Resolvent positive linear operators exhibit the reduction phenomenon. Proc. Natl. Acad. Sci. USA 109(10), 3705–3710 (2012)
Cantrell, R.S., Cosner, C., Deangelis, D.L., Padron, V.: The ideal free distribution as an evolutionarily stable strategy. J. Biol. Dyn. 1(3), 249–271 (2007)
Cantrell, R.S., Cosner, C., Lou, Y.: Movement toward better environments and the evolution of rapid diffusion. Math. Biosci. 204(2), 199–214 (2006)
Cantrell, R.S., Cosner, C., Lou, Y.: Evolution of dispersal in heterogeneous landscapes. In: Cantrell, S., Cosner, C., Ruan, S. (eds.) Spatial Ecology, pp. 213–229. CRC Press, Boca Raton (2010)
Cantrell, R.S., Cosner, C., Lou, Y.: Evolutionary stability of ideal free dispersal strategies in patchy environments. J. Math. Biol. 65(5), 943–965 (2012)
Cantrell, R.S., Cosner, C., Lou, Y., Schreiber, S.J.: Evolution of natal dispersal in spatially heterogeneous environments. Math. Biosci. 283, 136–144 (2017)
Chen, S., Liu, J., Wu, Y.: Invasion analysis of a two-species Lotka–Volterra competition model in an advective patchy environment. Stud. Appl. Math. 149, 762–797 (2022)
Chen, S., Shi, J., Shuai, Z., Wu, Y.: Global dynamics of a Lotka–Volterra competition patch model. Nonlinearity 35(2), 817 (2022)
Chen, S., Shi, J., Shuai, Z., Wu, Y.: Two novel proofs of spectral monotonicity of perturbed essentially nonnegative matrices with applications in population dynamics. SIAM J. Appl. Math. 82(2), 654–676 (2022)
Chen, S., Shi, J., Shuai, Z., Wu, Y.: Evolution of dispersal in advective patchy environments. J. Nonlinear Sci. 33, 40 (2023)
Chen, X., Lam, K.-Y., Lou, Y.: Dynamics of a reaction–diffusion–advection model for two competing species. Discrete Contin. Dyn. Syst. 32(11), 3841 (2012)
Cheng, C.-Y., Lin, K.-H., Shih, C.-W.: Coexistence and extinction for two competing species in patchy environments. Math. Biosci. Eng. 16(2), 909–946 (2019)
Cosner, C.: Variability, vagueness and comparison methods for ecological models. Bull. Math. Biol. 58(2), 207–246 (1996)
Dieckmann, U., Law, R.: The dynamical theory of coevolution: a derivation from stochastic ecological processes. J. Math. Biol. 34(5), 579–612 (1996)
Dockery, J., Hutson, V., Mischaikow, K., Pernarowski, M.: The evolution of slow dispersal rates: a reaction diffusion model. J. Math. Biol. 37(1), 61–83 (1998)
Geritz, S., Kisdi, E., Mesze, G., Metz, J.A.J.: Evolutionarily singular strategies and the adaptive growth and branching of the evolutionary tree. Evol. Biol. 12(1), 35–57 (1998)
Gourley, S.A., Kuang, Y.: Two-species competition with high dispersal: the winning strategy. Math. Biosci. Eng. 2(2), 345–362 (2005)
Hamida, Y.: The Evolution of Dispersal for the Case of Two-Patches and Two-Species with Travel Loss. PhD thesis, The Ohio State University (2017)
Hastings, A.: Can spatial variation alone lead to selection for dispersal? Theor. Popul. Biol. 24(3), 244–251 (1983)
Hess, P.: Periodic–Parabolic Boundary Value Problems and Positivity, volume 247 of Pitman Research Notes in Mathematics Series. Longman Scientific & Technical, Harlow (1991)
Hsu, S.B., Smith, H.L., Waltman, P.: Competitive exclusion and coexistence for competitive systems on ordered Banach spaces. Trans. Am. Math. Soc. 348(10), 4083–4094 (1996)
Huang, Q.-H., Jin, Y., Lewis, M.A.: \(R_0\) analysis of a Benthic-drift model for a stream population. SIAM J. Appl. Dyn. Syst. 15(1), 287–321 (2016)
Jiang, H., Lam, K.-Y., Lou, Y.: Are two-patch models sufficient? The evolution of dispersal and topology of river network modules. Bull. Math. Biol. 82(10), 131, 42 (2020)
Jiang, H., Lam, K.-Y., Lou, Y.: Three-patch models for the evolution of dispersal in advective environments: varying drift and network topology. Bull. Math. Biol. 83(10), 109, 46 (2021)
Jin, Y., Lewis, M.A.: Seasonal influences on population spread and persistence in streams: critical domain size. SIAM J. Appl. Math. 71(4), 1241–1262 (2011)
Johnson, M.L., Gaines, M.S.: Evolution of dispersal: theoretical models and empirical tests using birds and mammals. Ann. Rev. Ecol. Syst. 21, 449–480 (1990)
Kirkland, S., Li, C.-K., Schreiber, S.J.: On the evolution of dispersal in patchy landscapes. SIAM J. Appl. Math. 66(4), 1366–1382 (2006)
Lam, K.Y., Lou, Y., Lutscher, F.: Evolution of dispersal in closed advective environments. J. Biol. Dyn. 9(suppl. 1), 188–212 (2015)
Lam, K.-Y., Munther, D.: A remark on the global dynamics of competitive systems on ordered Banach spaces. Proc. Am. Math. Soc. 144(3), 1153–1159 (2016)
Levin, S.A., Cohen, D., Hastings, A.: Dispersal strategies in patchy environments. Theor. Popul. Biol. 26(2), 165–191 (1984)
Li, M.Y., Shuai, Z.: Global-stability problem for coupled systems of differential equations on networks. J. Differ. Equ. 248(1), 1–20 (2010)
Lin, K.-H., Lou, Y., Shih, C.-W., Tsai, T.-H.: Global dynamics for two-species competition in patchy environment. Math. Biosci. Eng. 11(4), 947–970 (2014)
Lou, Y.: Ideal free distribution in two patches. J. Nonlinear Model Anal. 2, 151–167 (2019)
Lou, Y., Lutscher, F.: Evolution of dispersal in open advective environments. J. Math. Biol. 69(6–7), 1319–1342 (2014)
Lou, Y., Nie, H., Wang, Y.: Coexistence and bistability of a competition model in open advective environments. Math. Biosci. 306, 10–19 (2018)
Lou, Y., Xiao, D.-M., Zhou, P.: Qualitative analysis for a Lotka–Volterra competition system in advective homogeneous environment. Discrete Contin. Dyn. Syst. 36(2), 953–969 (2016)
Lou, Y., Zhao, X.-Q., Zhou, P.: Global dynamics of a Lotka–Volterra competition–diffusion–advection system in heterogeneous environments. J. Math. Pures Appl. 9(121), 47–82 (2019)
Lou, Y., Zhou, P.: Evolution of dispersal in advective homogeneous environment: the effect of boundary conditions. J. Differ. Equ. 259(1), 141–171 (2015)
Lu, Z.Y., Takeuchi, Y.: Global asymptotic behavior in single-species discrete diffusion systems. J. Math. Biol. 32(1), 67–77 (1993)
Lutscher, F., Lewis, M.A., McCauley, E.: Effects of heterogeneity on spread and persistence in rivers. Bull. Math. Biol. 68(8), 2129–2160 (2006)
Lutscher, F., McCauley, E., Lewis, M.A.: Spatial patterns and coexistence mechanisms in systems with unidirectional flow. Theor. Popul. Biol. 71(3), 267–277 (2007)
Lutscher, F., Pachepsky, E., Lewis, M.A.: The effect of dispersal patterns on stream populations. SIAM Rev. 47(4), 749–772 (2005)
Ma, L., Tang, D.: Evolution of dispersal in advective homogeneous environments. Discrete Contin. Dyn. Syst. 40(10), 5815–5830 (2020)
McPeek, M.A., Holt, R.D.: The evolution of dispersal in spatially and temporally varying environments. Am. Nat. 140(6), 1010–1027 (1992)
Noble, L.: Evolution of Dispersal in Patchy Habitats. PhD thesis, The Ohio State University (2015)
Smith, H.L.: Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems. American Mathematical Society, Providence (1995)
Speirs, D.C., Gurney, W.S.C.: Population persistence in rivers and estuaries. Ecology 82(5), 1219–1237 (2001)
Vasilyeva, O., Lutscher, F.: Population dynamics in rivers: analysis of steady states. Can. Appl. Math. Q. 18(4), 439–469 (2010)
Vasilyeva, O., Lutscher, F.: How flow speed alters competitive outcome in advective environments. Bull. Math. Biol. 74(12), 2935–2958 (2012)
Xiang, J.-J., Fang, Y.: Evolutionarily stable dispersal strategies in a two-patch advective environment. Discrete Contin. Dyn. Syst. B 24(4), 1875 (2019)
Yan, X., Nie, H., Zhou, P.: On a competition–diffusion–advection system from river ecology: mathematical analysis and numerical study. SIAM J. Appl. Dyn. Syst. 21(1), 438–469 (2022)
Zhao, X.-Q., Zhou, P.: On a Lotka–Volterra competition model: the effects of advection and spatial variation. Calc. Var. Partial Differ. Equ. 55(4), 73 (2016)
Zhou, P.: On a Lotka–Volterra competition system: diffusion vs advection. Calc. Var. Partial Differ. Equ. 55(6), 137 (2016)
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Shanshan Chen is supported by National Natural Science Foundation of China (Nos. 12171117, 11771109) and Shandong Provincial Natural Science Foundation of China (No. ZR2020YQ01).
Appendix
Appendix
In the Appendix, we study the relations of \({{\overline{q}}}\), \({\underline{q}}\) and \(q_0\). For convenience, we recall the definition of \({{\overline{q}}}\), \({\underline{q}}\) and \(q_0\):
Lemma 6.4
Suppose that \((\textbf{H})\) holds, \(\varvec{r}\gg \varvec{0}\), and \(d_1,q_1>0\). Then, the following statements hold:
-
(i)
If \(q_1<{\underline{q}}\), then \(q_0>q_1\);
-
(ii)
If \(q_1>{{\overline{q}}}\), then \(q_0<q_1\);
-
(iii)
If \(q_1>{\underline{q}}\), then \(q_0>{\underline{q}}\);
-
(iv)
If \(q_1<{{\overline{q}}}\), then \(q_0<{{\overline{q}}}\).
Proof
By (5.10) and (5.11) and Lemma 5.1 (i), we have
which will be used in the proof below.
(i) By Lemma 5.1 (iv), we have \(u^*_1>u^*_2>u^*_3\). This, together with (6.9c) and (6.10a), implies that
(ii) By Lemma 5.1 (iii), we have \(u^*_1<u^*_2<u^*_3\). Then, by (6.10a) again, we obtain
By (6.10c), we obtain that
where we have used (6.10c) and \(u_2^*<u_3^*\) in the last step. It follows from (6.9c), (6.12) and (6.13) that \(q_0<q_1\).
(iii) We divide the proof into three cases:
For case (A1), we see from (6.9b) and (6.9c) that
For case (A2), we see from (6.9c) and (6.10a) that
Now we consider (A3). Suppose to the contrary that \(q_0\le {\underline{q}}\). This, combined with (6.9b) and (6.9c), yields
Noticing that \(u_2^*>k_2\), we see from (6.10c) that
Since \(u^*_1< u^*_2\), we see from (6.15) that \(u^*_2< u^*_3\). Then, we have
which yields
This, together with (6.16), (6.10a) and (6.10b), implies that
which contradicts (6.14). Therefore, \(q_0>{\underline{q}}\) for case (A3).
(iv) We first show that
and the proof is divided into three cases:
For case (B1), we have
For case (B2), we see from (6.10a) that
For case (B3), using similar arguments as the above case (A3), we have
This, combined with (6.10a) and (6.10b), implies that
Then, we show that
and the proof is also divided into three cases:
For case (C1), we see from (6.13) that
For case (C2), we have
For case (C3), we see from (6.10) that
By (6.18) and (6.20), we see that (iv) holds. \(\square \)
Remark 6.5
By \({\underline{q}}\le {{\overline{q}}}\) and Lemma 6.4, we see that if \(q_1< {\underline{q}}\), then \(q_1<q_0<{{\overline{q}}}\); if \(q_1> {{\overline{q}}}\), then \({\underline{q}}<q_0<q_1\); and if \({\underline{q}}<q_1<{{\overline{q}}}\), then \({\underline{q}}<q_0<{{\overline{q}}}\).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chen, S., Liu, J. & Wu, Y. On the impact of spatial heterogeneity and drift rate in a three-patch two-species Lotka–Volterra competition model over a stream. Z. Angew. Math. Phys. 74, 117 (2023). https://doi.org/10.1007/s00033-023-02009-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00033-023-02009-6
Keywords
- Lotka–Volterra competition model
- Patch environment
- Evolution of dispersal
- Directed drift
- Random movement