
Abstract
Our earlier work in Nguyen et al. (Maximizing metapopulation growth rate and biomass in stream networks. arXiv preprint arXiv:2306.05555, 2023) shows that concentrating resources on the upstream end tends to maximize the total biomass in a metapopulation model for a stream species. In this paper, we continue our research direction by further considering a Lotka–Volterra competition patch model for two stream species. We show that the species whose resource allocations maximize the total biomass has the competitive advantage.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
The impact of resource distributions on the persistence of a single species has been studied extensively (e.g., Cantrell and Cosner 1989, 2004; Berestycki et al. 2005; Lou 2006; Cantrell and Cosner 1998). Lou (2006) observed that if the dynamics of the species is modeled by a reaction-diffusion model with logistic type nonlinearity, the total biomass of the species may exceed the carrying capacity. Later, the ratio of total biomass over carrying capacity was studied both theoretically and experimentally (Bai et al. 2016; DeAngelis et al. 2016; Inoue and Kuto 2021; Liang and Lou 2012; Zhang et al. 2015, 2017). The distributions to maximize the growth rate (Cantrell and Cosner 1989, 1991; Lamboley et al. 2016; Lou and Yanagida 2006) and the total biomass (Ding et al. 2010; Mazari et al. 2020, 2022; Mazari and Ruiz-Balet 2021; Nagahara and Yanagida 2018) have been to shown to be of bang-bang type. Similar maximizing total biomass problems have been studied for patch models with logistic growth and random movement (Liang and Zhang 2021; Nagahara et al. 2021).
Resource allocations may also affect the interactions of multiple species (He and Ni 2013, 2016a, b, 2017; Lin et al. 2014; Gourley and Kuang 2005; Wei and Liu 2021). In a two-patch Lotka–Volterra competition model, it was shown that the species with a more heterogeneous distribution of resources will never lose the competition (Lin et al. 2014; Gourley and Kuang 2005). Similarly, in a two species reaction-diffusion competition model, it was shown that a species with a heterogeneous spatial distribution of resources will outcompete a species with homogeneously distributed resources (He and Ni 2016a). Meanwhile, when both species have a heterogeneous distribution of resources, the slower dispersing species wins competition (He and Ni 2017). For reaction-diffusion models it has also been shown that, when the carrying capacity is proportional to the growth rate, there is no optimal form of resource allocation (Hutson et al. 2003). Meanwhile, Mazari showed that the species whose resource allocation results in the largest total biomass wins the competition when the diffusion rate is large in a competition model with multiple species (Mazari 2019).
Our study is motivated by a series of recent works on metapopulation models in stream environments, where the individuals have both random movement and directed drift. In Nguyen et al. (2023), we considered the impact of the distribution of resources on the persistence of a single stream species. In particular, we showed that to maximize the total biomass one should concentrate the resources on the upstream ends while to maximize the growth rate of the population one may need to concentrate the resources on the downstream ends. In Jiang et al. (2020, 2021), the authors studied the joint impact of the diffusion rate, advection rate and network topology on the competition outcome of two stream species in a three patch Lotka–Volterra competition model whose patches are constructed as shown in Fig. 1. When there are n patches aligned along a line, the results in Chen et al. (2022, 2022, 2023) showed that the magnitude of movement rates, the convexity of the drift rates and the population loss rate at the downstream end can significantly alter the competition outcome of two species. For works on reaction-diffusion-advection models for stream species, we refer the interested readers to Lam et al. (2016); Lou and Lutscher (2014); Lou and Zhou (2015); Lutscher et al. (2006); Pachepsky et al. (2005); Speirs and Gurney (2001); Vasilyeva and Lutscher (2012); Wang et al. (2019); Yan et al. (2022); Zhou et al. (2021) and the references therein.
Motivated by the aforementioned studies, we consider the following two-species n-patch Lotka–Volterra competition model in stream environment:
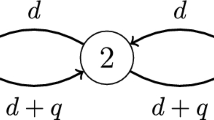
Here the vectors \(\varvec{u}=(u_1, \dots , u_n)\) and \(\varvec{v}=(v_1, \dots , v_n)\) denote the densities of two competing stream species at each patch location. The nonnegative vectors \(\varvec{r}=(r_1, \dots , r_n)\) and \(\varvec{s}=(s_1, \dots , s_n)\) are the growth rates of \(\varvec{u}\) and \(\varvec{v}\), respectively, and the carrying capacity is assumed to be a positive constant k for all the patches. Two \(n\times n\) matrices \(D=(D_{ij})\) and \(Q=(Q_{ij})\) represent the random movement pattern and directed drift pattern of individuals respectively. For the three network configurations with \(n=3\) in Fig. 1 (see Jiang et al. 2020, 2021), the corresponding matrices D and Q are as follows:
-
Case 1:
$$\begin{aligned} D=\begin{bmatrix} -2 &{} 1 &{}1 \\ 1&{}-1&{} 0\\ 1&{}0 &{} -1 \\ \end{bmatrix},\;\;Q=\begin{bmatrix} -2 &{} 0 &{}0 \\ 1&{}0&{} 0\\ 1&{}0 &{} 0\\ \end{bmatrix}; \end{aligned}$$(1.2) -
Case 2:
$$\begin{aligned} D=\begin{bmatrix} -1 &{} 0 &{}1 \\ 0&{}-1&{} 1\\ 1&{}1 &{} -2 \\ \end{bmatrix},\;\;Q=\begin{bmatrix} -1 &{} 0 &{}0 \\ 0&{}-1&{} 0\\ 1&{}1 &{} 0\\ \end{bmatrix}; \end{aligned}$$(1.3) -
Case 3:
$$\begin{aligned} D=\begin{bmatrix} -1 &{} 1 &{}0 \\ 1&{}-2&{} 1\\ 0&{}1 &{} -1 \\ \end{bmatrix},\;\;Q=\begin{bmatrix} -1 &{} 0 &{}0 \\ 1&{}-1&{} 0\\ 0&{}1 &{} 0\\ \end{bmatrix}. \end{aligned}$$(1.4)

A stream with three patches, where d is the random movement rate and q is the directed drift rate. i Patch 1 is the upstream end and patches 2 and 3 are the downstream ends. ii Patches 1 and 2 are the upstream ends and patch 3 is the downstream end. iii Patch 1 is the upstream end and patch 3 is the downstream end
Our objective is to determine how the distribution of resources for each species, as determined by \({\varvec{r}}\) and \({\varvec{s}}\), impact competitive outcomes for model (1.1). We make the assumption that the resources are proportional to the growth rate in each patch and the two species \(\varvec{u}\) and \(\varvec{v}\) have the same amount of resources, i.e.
We show that if the resources of species \(\varvec{u}\) are distributed to maximize its biomass, i.e. all resources are distributed in the most upstream patches (see Nguyen et al. 2023), while the resources of species \(\varvec{v}\) are not, then species \(\varvec{u}\) always wins the competition. For example, for configuration (i) in Fig. 1, such a distribution of species \(\varvec{u}\) corresponds to \(\varvec{r}=(r, 0, 0)\) while the distribution of species \(\varvec{v}\) satisfies \(\varvec{s}\ne (r,0,0)\).
Our paper is organized as follows. In Section 2, we present some preliminary results which follow from existing theory. In Section 3, we consider the three-node stream networks shown in Fig. 1. For each of these configurations we show that a species whose resources are distributed so that their total biomass is maximized in the absence of competition is able to out-compete a species whose resources are not optimally distributed. In Section 4, we extend these results to apply to n-patch stream networks.
2 Preliminaries
Let \(\varvec{w}=(w_1, \dots , w_n)\) be a real vector. We write \(\varvec{w}\gg \varvec{0}\) if \(w_i>0\) for all \(i=1, \dots , n\), and \(\varvec{w}>\varvec{0}\) if \(\varvec{w}\ge \varvec{0}\) but \(\varvec{w}\ne \varvec{0}\). Let \(A=(a_{ij})_{n\times n}\) be a real square matrix. Let \(\sigma (A)\) be the set of all eigenvalues of A, and s(A) be the spectral bound of A, i.e.
The matrix A is called irreducible if it cannot be placed into block upper triangular form by simultaneous row and column permutations and essentially nonnegative if \(a_{ij}\ge 0\) for all \(1\le i, j\le n\) such that \(i\ne j\). By the Perron-Frobenius Theorem, if A is irreducible and essentially nonnegative, then \(\lambda _1=s(A)\) is a simple eigenvalue of A. Moreover, \(\lambda _1\) (called the principal eigenvalue of A) is associated with an eigenvector whose components are all positive, which is the unique eigenvalue associated with a nonnegative eigenvector.
Before studying the two species competition model, we revisit the following single species meta-population model:
Here, \(\varvec{u}=(u_1, \dots , u_n)\) is the density of a meta-population living in n-patches; \(\varvec{k}=(k_1, \dots , k_n)\) is the carrying capacity; \(\varvec{r}=(r_1, \dots , r_n)\) is the growth rate. The coefficients \(\ell _{ij}\ge 0\) denote the movement rate of the individuals from patch j to patch i for \(1\le i,j\le n\) and \(i\not = j\); \(l_{ii}=-\sum _{j\ne i} l_{ji}\) is the total movement rate out from patch i. Then the \(n\times n\) connection matrix L is of the form
It is easy to see that \((1,1, \dots , 1)\) is a left eigenvector of L corresponding to eigenvalue 0. We always assume that L is irreducible. By the Perron-Frobenius Theorem, 0 is the principal eigenvalue of L.
We can associate L with a weighted, directed graph (digraph) \({\mathcal {G}}\) consisting of n nodes (each node i in \({\mathcal {G}}\) corresponds to patch i). In \({\mathcal {G}}\), there is a directed edge (arc) from node j to node i if and only if \(\ell _{ij}>0\). The couple \(({\mathcal {G}}, L)\) is called the movement network associated with (2.1).
The global dynamics of (2.1) are well-known:
Lemma 2.1
(Cosner 1996; Li and Shuai 2010; Lu and Takeuchi 1993; Takeuchi 1996) Suppose that L is essentially nonnegative and irreducible matrix that is defined in (2.2). If \(\varvec{r}>\varvec{0}\) and \(\varvec{k}\gg \varvec{0}\), then model (2.1) has a unique positive equilibrium, which is globally asymptotically stable.
By Lemma 2.1, model (1.1) has two semitrivial equilibria \(E_1:=(\varvec{u}^*, \varvec{0})\) and \(E_2:=(\varvec{0}, \varvec{v}^*)\). By the well-known monotone dynamical system theory (Hess 1991; Hsu et al. 1996; Lam and Munther 2016; Smith 1995), the global dynamics of (1.1) is closely related to the local properties of its equilibria. Denote \(X={\mathbb {R}}_+^n\times {\mathbb {R}}_+^n\). Let \(\le _K\) be the order in X induced by the cone \(K={\mathbb {R}}_+^n\times \{-{\mathbb {R}}_+^n\}\). Then if \(\varvec{x}=(\bar{\varvec{u}}, \bar{\varvec{v}}), \varvec{y}=(\tilde{\varvec{u}}, \tilde{\varvec{v}})\in X\), we write \(\varvec{x}\le _K \varvec{y}\) if \(\bar{\varvec{u}}\le \tilde{\varvec{u}}\) and \(\bar{\varvec{v}}\ge \tilde{\varvec{v}}\); \(\varvec{x}<_K \varvec{y}\) if \(\varvec{x}\le _K \varvec{y}\) and \(\varvec{x}\ne \varvec{y}\). We utilize the following result later (this result is proved in Smith (1995) for the case \(n=2\) first but it holds for any \(n\ge 2\) (Smith 1995, Page 70)):
Lemma 2.2
(Smith 1995, Theorem 4.4.2) Suppose that \(E_1\) is linearly unstable. Then one of the following holds:
-
(i)
\(E_2\) attracts all solutions with initial data \((\varvec{u}_0,\varvec{v}_0)\in X\) satisfying \(\varvec{v}_0>0\). In this case, \(E_2\) is linearly stable or neutrally stable;
-
(ii)
There exists a positive equilibrium E satisfying \(E_2\ll _K E\ll _K E_1\) such that E attracts all solutions with initial data \((\varvec{u}_0, \varvec{v}_0)\in X\) satisfying \(E\le _K (\varvec{u}_0, \varvec{v}_0)<_K E_1\).
By Lemma 2.2, if \(E_2\) is linearly unstable and the model has no positive equilibrium, then \(E_1\) is globally attractive. It is easy to see that the stability of \(E_1\) is determined by the sign of \(\lambda _1(\varvec{s}, \varvec{u}^*)\), which is the principal eigenvalue of the matrix \(dD+qQ+\text {diag}(s_i(1-u^*_i/k))\): if \(\lambda _1(\varvec{s}, \varvec{u}^*)<0\), \(E_1\) is locally asymptotically stable; if \(\lambda _1(\varvec{s}, \varvec{u}^*)>0\), \(E_1\) is unstable; if \(\lambda _1(\varvec{s}, \varvec{u}^*)=0\), \(E_1\) is linearly neutrally stable. Similarly, the local stability of \(E_2\) is determined by the sign of \(\lambda _1(\varvec{r}, \varvec{v}^*)\), which is the principal eigenvalue of the matrix \(dD+qQ+\text {diag}(r_i(1-v^*_i/k))\).
3 Stream networks of three nodes
In this section, we consider model (1.1) for the three-node stream networks shown in Fig. 1. Here we provide detailed analysis for configuration (i), with analogous results for configurations (ii) and (iii) provided in the appendix. Our results state that a species whose resources are concentrated on the upstream end will have the competitive advantage. In particular, for configuration (i) we show that if \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}\ne \varvec{r}\), then species \(\varvec{u}\) always wins the competition.
We first prove the following lemma which is used to show that model (1.1) does not have a positive equilibrium.
Lemma 3.1
Suppose that D and Q are given by (1.2). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). If \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1), then \(u_1+v_1<k\), \(u_2+v_2>k\) and \(u_3+v_3>k\).
Proof
Suppose that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Then \((\varvec{u}, \varvec{v})\) satisfies
Let \(w_i:=u_i+v_i\) for \(i=1, 2, 3\). Adding each corresponding pair of equations above, we have
We prove that \(w_2=u_2+v_2 >k\) by contradiction. Assume to the contrary that \(w_2\le k\). Then by the second equation of (3.2), we have
This implies
By \(r_1=r>0\) and the first equation of (3.2), we have
Since we have shown that \((d+q)w_1 - dw_2 \le 0\) and \(w_1 <k\), we must have \((d+q)w_1-dw_3>0\). Thus
This further implies
which contradicts the third equation of (3.2). Therefore, we must have \(u_2+v_2>k\). Similarly, we have \(u_3+v_3>k\).
Since \(\varvec{s}\ne \varvec{r}\), either \(s_2\ne 0\) or \(s_3\ne 0\). Without loss of generality, say \(s_2\ne 0\). Then by the second equation of (3.2) and \(w_2>k\), \((d+q)w_1-dw_2>0\). By the third equation of (3.2) and \(w_3>k\), we have \((d+q)w_1-dw_3\ge 0\). Finally by the first equation of (3.2) and \(s_1>0\), we have \(w_1=u_1+v_1<k\). \(\square \)
Next, in Lemma 3.3 we make use of the following well-known result (e.g., see (Berman and Plemmons 1994, Corollary 2.1.5)) to prove the non-existence of a positive equilibrium.
Lemma 3.2
Suppose that P and Q are \(n\times n\) real-valued matrices, P is essentially nonnegative, Q is nonnegative and nonzero, and \(P+Q\) is irreducible. Then, \(s(P+Q)>s(P)\).
Lemma 3.3
Suppose that D and Q are given by (1.2). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). Then model (1.1) has no positive equilibrium.
Proof
Suppose to the contrary that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Then \((\varvec{u}, \varvec{v})\) satisfies (3.1). By the first equation of (3.1), \(\varvec{u}\) is a positive eigenvector of matrix \(M_1:=dD+qQ+\text {diag}(r_i(1-(u_i+v_i)/k))\) corresponding with eigenvalue 0. By the Perron-Frobenius Theorem, we must have \(s(M_1)=0\). Similarly, \(\varvec{v}\) is a positive eigenvector of matrix \(M_2:=dD+qQ+\text {diag}(s_i(1-(u_i+v_i)/k))\) corresponding with eigenvalue 0 and \(s(M_2)=0\). By the assumptions on \(\varvec{r}\) and \(\varvec{s}\) and Lemma 3.1, we have
and
Therefore, by Lemma 3.2, we must have \(s(M_1)>s(M_2)\), which is a contradiction. This proves the result. \(\square \)
In the following two lemmas, we show that the semitrivial equilibrium \(E_2\) is always unstable.
Lemma 3.4
Suppose that D and Q are given by (1.2). Let \(\varvec{s}=(s_1, s_2, s_3)\ge \varvec{0}\) with \(s_2>0\) or \(s_3>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) satisfies \(v^*_1<k\).
Proof
We observe that \(\varvec{v}^*\) must satisfy
That is
Assume to the contrary that \(v_1^*\ge k\). By the first equation of (3.3), either \((d+q)v^*_1-dv^*_2\le 0\) or \((d+q)v^*_1-dv^*_3\le 0\). Without loss of generality, say \(s_2>0\). If \((d+q)v^*_1-dv^*_2\le 0\), then by the second equation of (3.3), we have \(v^*_2\le k\). This implies
which is a contradiction. Hence, \((d+q)v^*_1-dv^*_3\le 0\). If \(s_3>0\), the third equation of (3.3) implies \(v^*_3\le k\). Then,
which is a contradiction. If \(s_3=0\), then \((d+q)v^*_1-dv^*_3=0\). Again by \(v_1^*\ge k\) and the first equation of (3.3), \((d+q)v^*_1-dv^*_2\le 0\). This leads to contradiction by the second equation of (3.3) and (3.4). \(\square \)
Lemma 3.5
Suppose that D and Q are given by (1.2). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) is unstable and the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is stable for model (1.1).
Proof
The stability of \(E_2\) is determined by the sign of \(\lambda _1(\varvec{r}, \varvec{v}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(r_i(1-v^*_i/k))\). By the assumptions on \(\varvec{r}\) and Lemma 3.4, we have
and
Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{r}, \varvec{v}^*)>s(dD+qQ)=0\). Hence, \(E_2\) is unstable.
The stability of \(E_1\) is determined by the sign of \(\lambda _1(\varvec{s}, \varvec{u}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(s_i(1-u^*_i/k))\). By Nguyen et al. (2023), we have \(\varvec{u}^*=(k, (d+q)k/d, (d+q)k/d)\). So,
and
with at least one strict sign by the assumption on \(\varvec{s}\). Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{s}, \varvec{u}^*)<s(dD+qQ)=0\). Hence, \(E_1\) is stable. \(\square \)
By the theory of monotone dynamical systems (Lemma 2.2) and Lemmas 3.3 and 3.5, we obtain the following result:
Theorem 3.6
Suppose that D and Q are given by (1.2). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). Then the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is globally asymptotically stable for model (1.1).
4 Stream networks of n nodes
In this section, we generalize the results in Sect. 3 to a certain type of network of n nodes. We recall the definition of stream networks of n nodes in Nguyen et al. (2023).
Definition 4.1
Let G be a directed graph, and denote the set of nodes of G by V. Consider a function \(f: V\rightarrow {\mathbb {Z}}_{\ge 0}\). For each node i, we call f(i) the level of the node and (G, f) a leveled graph if the following assumptions are satisfied
-
(i)
For each \(0\le k\le \max _{i\in V}\{f(i)\}\), there exists a node j such that \(f(j)=k\).
-
(ii)
For each pair of nodes i and j, there is no edge between i and j if \(|f(i)-f(j)|\ne 1\).
We use level graphs to describe a type of stream network, where the nodes in further downstream positions have larger levels. The left digraph in Fig. 2 is a leveled graph while the right digraph is not.

The left digraph is a leveled graph with level function \(f(1)=0, f(2)=f(3)=1, f(4)=2\). The right digraph cannot be a leveled graph for any choice of level function
Definition 4.2
(Nguyen et al. 2023) Consider a graph G with level function f and connection matrix L. We say that (G, f, L) is a homogeneous flow stream network if the following assumptions are satisfied:
-
(i)
The matrix L is irreducible.
-
(ii)
If there is an edge from node i to node j, then there is also an edge from node j to node i.
-
(iii)
If there is an edge from node i to node j, then the weight is \(\ell _{ij} = d+q\) if \(f(j)-f(i)=1\) (i.e. the edge is from an upstream to a downstream node) and \(\ell _{ij}=d\) if \(f(i)-f(j)=1\) (i.e. the edge is from a downstream to an upstream node). Here, d and q are positive constants.
The connection matrix L of a homogeneous flow stream network can be written as \(L=dD+qQ\). We recall the following result about the positive eigenvector of L proved in Nguyen et al. (2023).
Lemma 4.3
Let (G, f, L) be a homogeneous flow stream network. Let \(\varvec{v}\) be the solution to \(L\varvec{v}=(dD+qQ)\varvec{v} = 0\). Then the eigenvector \(\varvec{v}\) writes, up to a constant multiple, as
Let \(\varvec{u}^*=(u_1^*, \dots , u_n^*)\) be the positive equilibrium of
The total biomass \({\mathcal {K}}\) of \(\varvec{u}^*\) is defined as \({\mathcal {K}}:=\sum _{i=1}^n u_i^*\). We also recall the following theorem in Nguyen et al. (2023) about the total biomass \({\mathcal {K}}\):
Theorem 4.4
Let (G, f, L) be a homogeneous flow stream network, where \(L=dD+qQ\). Suppose that \(\varvec{r}=(r_1,\dots , r_n)>\varvec{0}\) with \(\sum _{i=1}^nr_i=r>0\) and \(k>0\). Then the total biomass \({\mathcal {K}}\) of the positive equilibrium of (4.1) has the upper bound
Moreover, the maximum is achieved as the upper bound when \(r_i=0\) for any node i with positive level, i.e. \(f(i)>0\). In this case, \(u_i^*=k(\frac{d+q}{d})^{f(i)}\).
The main result we prove in this section is the following, which states that to gain a competitive advantage in a homogeneous flow stream network one needs to distribute all the resources to the upstream ends, i.e. nodes with level 0.
Theorem 4.5
Let (G, f, L) be a homogeneous flow stream network, where \(L=dD+qQ\). Let \(k>0\) and \(\varvec{r}, \varvec{s}>\varvec{0}\) such that \(\sum _{i=1}^n r_i=\sum _{i=1}^n s_i=r>0\). Suppose that \(r_i=0\) for any node i with \(f(i)>0\) and there exists at least one node \(i_0\) with \(f(i_0)>0\) such that \(s_{i_0}>0\). Then the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is globally asymptotically stable for (1.1).
4.1 Proof of Theorem 4.5
Suppose to the contrary that \((\varvec{u},\varvec{v})\) is a positive equilibrium of (1.1). Let \(w_i^*=u_i+v_i\) for each \(i=1, \dots , n\). To show the non-existence of a positive equilibrium \(E^*\), we recall the sign pattern approach used in Nguyen et al. (2023). We associate the stream network with a sign pattern graph. The nodes of the sign pattern graph are the same as the nodes in the stream network, but additionally we assign each node i in the sign pattern graph with a sign based on the value of \(w_i^*\):
Next, if there is an edge between node i and an adjacent, downstream node j in the stream network, we draw an edge between node i and node j in the sign pattern graph as follows:
-
1.
There is a directed edge from node i to node j (adjacent, downstream of node i), denoted \(i\rightarrow j\), if
$$\begin{aligned} (d+q)w_i^* > dw_j^*. \end{aligned}$$ -
2.
There is a directed edge from node j to node i (adjacent, downstream of node i), denoted \(j\rightarrow i\), if
$$\begin{aligned} (d+q)w_i^* < dw_j^*. \end{aligned}$$ -
3.
There is an undirected edge between node i and node j, denoted \(i - j\), if
$$\begin{aligned} (d+q)w_i^* = dw_j^*. \end{aligned}$$
The edges in the sign pattern graph describe the net flow between adjacent nodes in the stream network.
Lemma 4.6
For any \((+)\) node, there must be at least one directed edge out of the node. For any \((-)\) node, there must be at least one directed edge into the node. For any \((0),(0^+),(0^-)\) node, either all edges connected to the node are undirected, or there must be at least one edge into and one edge out of the node.
Proof
Suppose node i has sign \((+)\). We add the equations of \(u_i\) and \(v_i\) to obtain
Since \(w_i^* < k\), there must be a negative term in the sum above corresponding to a node j adjacent to node i. It is easy to check that whether j is upstream or downstream of i we always have an edge \(i\rightarrow j\) in the sign pattern graph. We can repeat the same argument for the nodes with sign \((-), (0), (0^+), (0^-)\). \(\square \)
Lemma 4.7
If \(i-j\) or \(i\rightarrow j\), then
Proof
If node i is upstream of node j, we have \(f(i)-f(j)=-1\). Since \(i-j\) or \(i\rightarrow j\), by the way we assign edges to the sign pattern graph we must have
If node i is downstream of node j, we have \(f(i)-f(j)=1\). Since \(i-j\) or \(i\rightarrow j\), again we have
\(\square \)
The following corollary follows directly from Lemma 4.7.
Corollary 4.8
If node i is downstream of node j (i.e. \(f(i)-f(j)=1\)) and \(i-j\) or \(i\rightarrow j\) then \(w_i^* > w_j^*\).
Corollary 4.9
If there is a path from node i to node j, i.e. there exist nodes \(i_1,i_2,\dots ,i_h\) such that \(i - (or\rightarrow ) i_1 - (or\rightarrow ) \dots - (or\rightarrow ) i_h - (or\rightarrow ) j\), then
The equality happens when all edges are undirectedly connected.
Proof
We apply Lemma 4.7 repeatedly
Since each equality happens when the corresponding edge is −, the overall equality happens when all the edges in the path are −. \(\square \)
Corollary 4.10
If there is a cycle \(i - (or\rightarrow ) i_1 - (or\rightarrow ) \dots - (or\rightarrow ) i_h - (or\rightarrow ) i\), then all the edges in the cycle must be −.
Proof
The proof follows directly from Corollary 4.9 where we set \(j=i\). \(\square \)
Lemma 4.11
For any i such that \(f(i)=0\) (i.e. the most upstream nodes) and \(r_i>0\), we have \(w_i^* \le k\).
Proof
Assume by contradiction that there exists a node i such that \(f(i)=0\), \(r_i>0\) and \(w_i^* >k\). Then node i has \((-)\) sign and by Lemma 4.6, there must exist a node \(i_1\) such that \(i_1\rightarrow i\). Since node i is a most upstream node, node \(i_1\) must be downstream of it and thus by Corollary 4.8, we must have \(w_{i_1}^*> w_i^* >k\), thus node \(i_1\) has either sign \((-)\) or \((0^-)\).
Since there is already an edge out of node \(i_1\), this means there exists a node \(i_2\) such that \(i_2 \rightarrow i_1\). This gives us a path from \(i_2\) to i and thus by Corollary 4.9 we must have
since \(f(i)=0\). Thus again we must have \(i_2\) has either sign \((-)\) or \((0^-)\).
For each index \(h\ge 3\), we can repeat the argument to obtain node \(i_h\) such that \(i_h \rightarrow i_{h-1}\) and node \(i_h\) has either sign \((-)\) or \((0^-)\). Since the number of nodes is finite, the above process must stop after a finite number of steps. This is only possible if we have a cycle. However, since all edges in this cycle are directed, we reach a contradiction based on Corollary 4.10. \(\square \)
Lemma 4.12
Let (G, f, L) be a homogeneous flow stream network, where \(L=dD+qQ\). Let \(k>0\) and \(\varvec{r}, \varvec{s}>\varvec{0}\) such that \(\sum _{i=1}^n r_i=\sum _{i=1}^n s_i=r>0\). Suppose that \(r_i=0\) for any node i with \(f(i)>0\) and there exists at least one node \(i_0\) with \(f(i_0)>0\) such that \(s_{i_0}>0\). Then a positive equilibrium \(E^*\) does not exist.
Proof
Suppose by contradiction that a positive equilibrium \(E^*\) exists. From the assumption on \(\varvec{r}\), there must exists a node i with \(f(i)=0\) and \(r_i>0\). Taking the sum of all \(du_i/dt\), at the positive equilibrium, we must have
From Lemma 4.11, for any i such that \(f(i)=0\) and \(r_i>0\) we must have \(w_i^*\le k\). From this fact and equation (4.2), we have \(w_i^*=k\) for any i such that \(f(i)=0\) and \(r_i>0\).
Without loss of generality, suppose that \(f(1)=0\) and \(r_1>0\). From the argument above, \(w_1^*=k\) and node 1 has sign (0). If there is a node j adjacent to node 1 such that \(j\rightarrow 1\), then repeating the argument in Lemma 4.11, we have a cycle which leads to a contradiction. Since there is no directed edge into node 1, from Lemma 4.6, all edges connected to node 1 must be undirected. Let \(j_1,\dots ,j_h\) denote the nodes adjacent to node 1. Then from Corollary 4.8, they must have sign \((-)\) or \((0^-)\).
We will show again that all edges connected to node \(j_1\) must be undirected, and a similar argument can be applied to show all edges connected to nodes \(j_1,\dots ,j_h\) are undirected. Assume by contradiction that not all edges connected to node \(j_1\) are undirected. Since node \(j_1\) has sign \((-)\) or \((0^-)\), from Lemma 4.6 there must be a directed edge into node \(j_1\) from another node \(j'\). However, that means there is a path \(j' \rightarrow j_1 - 1\). From Corollary 4.9 we have
and thus node \(j'\) has sign \((-)\) or \((0^-)\) and there is a directed edge into it. We repeat the argument in Lemma 4.11, which leads to a cycle and thus a contradiction.
The same argument can be repeated, and since the stream network is strongly connected, we have all edges in the sign pattern graph must be undirected. Thus all nodes aside from the most upstream nodes must have sign \((-)\) or \((0^-)\). Since there exists \(i_0\) such that \(f(i_0)>0\) and \(s_{i_0}>0\), there must be at least one node with sign \((-)\). Taking the sum of all equations for \(u_i\) and \(v_i\), we have
However, since there is at least one node with sign \((-)\) and no node with sign \((+)\), the right hand side of the equation above must be strictly negative, which is a contradiction. Thus a positive equilibrium \(E^*\) does not exists. \(\square \)
The proof of the following result is similar to that of Theorem 4.4. We include it here for the sake of completeness.
Lemma 4.13
Let (G, f, L) be a homogeneous flow stream network, where \(L=dD+qQ\). Suppose \(\varvec{k}=(k, \dots , k)\) with \(k>0\) and \(\varvec{s}>\varvec{0}\). Let \(\varvec{v}^*=(v^*_1, \dots , v^*_n)\) be the positive equilibrium of (2.1). If there exists at least one node \(i_0\) with \(f(i_0)>0\) such that \(s_{i_0}>0\), then \(v^*_i<k\) for all node i with \(f(i)=0\).
Proof
Since \(L=dD+qQ\) is essentially nonnegative and irreducible, by Smith (1995, Theorem 4.1.1), the solutions of (1.1) induce a strongly monotone dynamical system: if \(\varvec{u}_1(0)>\varvec{u}_2(0)\) then the corresponding solutions satisfy \(\varvec{u}_1(t)\gg \varvec{u}_2(t)\) for all \(t>0\). By Lemma 4.3, \(dD+qQ\) has a positive eigenvector \(\varvec{v}=(v_1, \dots , v_n)\) such that \(v_i=1\) if \(f(i)=0\) for all \(i=1, \dots , n\). Moreover, \(v_i>1\) if \(f(i)>0\). Define \(\bar{\varvec{u}}=k\varvec{v}\). Since \(\bar{\varvec{u}}\) is an eigenvector of \(dD+qQ\) corresponding to eigenvalue 0, we have
Moreover, the \(i_0\)-th inequality is strict since \(s_{i_0}>0\) and \(v_{i_0}>1\). Hence, the solution \(\varvec{u}(t)\) of (1.1) with initial condition \(\varvec{u}(0)=\varvec{{\bar{u}}}\) is strictly decreasing and converges to an equilibrium (Smith 1995, Proposition 3.2.1), which is the positive equilibrium \(\varvec{v}^*\) by Lemma 2.1. Hence, \(\varvec{v}^*\ll \bar{\varvec{u}}\). In particular, \(v_i^*<k\) if \(f(i)=0\). \(\square \)
We are now ready to prove that \(E_2\) is unstable.
Lemma 4.14
Let (G, f, L) be a homogeneous flow stream network, where \(L=dD+qQ\). Let \(k>0\) and \(\varvec{r}, \varvec{s}>\varvec{0}\) such that \(\sum _{i=1}^n r_i=\sum _{i=1}^n s_i=r>0\). Suppose that \(r_i=0\) for any node i with \(f(i)>0\) and there exists at least one node \(i_0\) with \(f(i_0)>0\) such that \(s_{i_0}>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) of (1.1) is unstable and the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) of (1.1) is stable.
Proof
To see that \(E_2\) is unstable, it suffices to show \(\lambda _1:=\lambda _1(\varvec{r}, \varvec{v}^*)>0\), where \(\lambda _1(\varvec{r}, \varvec{v}^*)\) is the principal eigenvalue of the matrix \(dD+qQ+\text {diag}(r_i(1-v^*_i/k))\). Let \(\varvec{\varphi }=(\varphi _1, \dots , \varphi _n)\) be a positive eigenvector corresponding with \(\lambda _1\). Then,
Adding up all the equations and noticing that each column sum of \(dD+qQ\) is zero, we obtain
where we used the assumption that \(r_i=0\) if \(f(i)>0\) in the last step. By Lemma 4.13, \(v_i^*<k\) if \(f(i)=0\). Therefore, we have \(\lambda _1>0\).
The stability of \(E_1\) is determined by the sign of \(\lambda _1(\varvec{s}, \varvec{u}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(s_i(1-u^*_i/k))\). By Theorem 4.4, we have \( u_i^*=k(\frac{d+q}{d})^{f(i)}\). So we have
and
with at least one strict sign due to the assumption on \(\varvec{s}\). Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{s}, \varvec{u}^*)<s(dD+qQ)=0\). Hence, \(E_1\) is stable. \(\square \)
Finally, Theorem 4.5 follows from Lemmas 2.2, 4.12, and 4.14.
References
Bai X, He X, Li F (2016) An optimization problem and its application in population dynamics. Proc Am Math Soc 144(5):2161–2170
Berestycki H, Hamel F, Roques L (2005) Analysis of the periodically fragmented environment model: I-species persistence. J Math Biol 51(1):75–113
Berman A, Plemmons RJ (1994) Nonnegative matrices in the mathematical sciences, volume 9 of classics in applied mathematics. Society for Industrial and Applied Mathematics (SIAM), Philadelphia, PA
Cantrell RS, Cosner C (1989) Diffusive logistic equations with indefinite weights: population models in disrupted environments. Proc R Soc Edinb Sect A Math 112(3–4):293–318
Cantrell RS, Cosner C (1991) Diffusive logistic equations with indefinite weights: population models in disrupted environments II. SIAM J Math Anal 22(4):1043–1064
Cantrell RS, Cosner C (1998) On the effects of spatial heterogeneity on the persistence of interacting species. J Math Biol 37(2):103–145
Cantrell RS, Cosner C (2004) Spatial ecology via reaction-diffusion equations. Wiley, Hoboken
Chen S, Liu J, Wu Y (2022) Evolution of dispersal in advective patchy environments with varying drift rates. Submitted
Chen S, Liu J, Wu Y (2022) Invasion analysis of a two-species Lotka–Volterra competition model in an advective patchy environment. Stud Appl Math 149(3):762–797
Chen S, Shi J, Shuai Z, Wu Y (2023) Evolution of dispersal in advective patchy environments. J Nonlinear Sci 33:40(40):1–35
Cosner C (1996) Variability, vagueness and comparison methods for ecological models. Bull Math Biol 58(2):207–246
DeAngelis DL, Ni W-M, Zhang B (2016) Dispersal and spatial heterogeneity: single species. J Math Biol 72(1):239–254
Ding W, Finotti H, Lenhart S, Lou Y, Ye Q (2010) Optimal control of growth coefficient on a steady-state population model. Nonlinear Anal Real World Appl 11(2):688–704
Gourley SA, Kuang Y (2005) Two-species competition with high dispersal: the winning strategy. Math Biosci Eng 2(2):345
He X, Ni W-M (2013) The effects of diffusion and spatial variation in Lotka–Volterra competition-diffusion system II: the general case. J Differ Equ 254(10):4088–4108
He X, Ni W-M (2016) Global dynamics of the Lotka–Volterra competition-diffusion system with equal amount of total resources, II. Calc Var Partial Differ Equ 55(2):25
He X, Ni W-M (2016) Global dynamics of the Lotka–Volterra competition-diffusion system: diffusion and spatial heterogeneity I. Commun Pure Appl Math 69(5):981–1014
He X, Ni W-M (2017) Global dynamics of the Lotka–Volterra competition-diffusion system with equal amount of total resources, iii. Calc Var Partial Differ Equ 56(5):132
Hess P (1991) Periodic-parabolic boundary value problems and positivity, vol 247. Pitman research notes in mathematics series. Longman Scientific & Technical, Harlow
Hsu SB, Smith HL, Waltman P (1996) Competitive exclusion and coexistence for competitive systems on ordered Banach spaces. Trans Am Math Soc 348(10):4083–4094
Hutson V, Martinez S, Mischaikow K, Vickers GT (2003) The evolution of dispersal. J Math Biol 47(6):483–517
Inoue J, Kuto K (2021) On the unboundedness of the ratio of species and resources for the diffusive logistic equation. Discrete Contin Dyn Sys-Ser B 26(5):2441–2450
Jiang H, Lam KY, Lou Y (2020) Are two-patch models sufficient? The evolution of dispersal and topology of river network modules. Bull. Math. Biol., 82(10):Paper No. 131, 42
Jiang H, Lam K-Y, Lou Y (2021) Three-patch models for the evolution of dispersal in advective environments: Varying drift and network topology. Bull Math Biol 83(10):1–46
Lam K-Y, Lou Y, Lutscher F (2016) The emergence of range limits in advective environments. SIAM J Appl Math 76(2):641–662
Lam K-Y, Munther D (2016) A remark on the global dynamics of competitive systems on ordered Banach spaces. Proc Am Math Soc 144(3):1153–1159
Lamboley J, Laurain A, Nadin G, Privat Y (2016) Properties of optimizers of the principal eigenvalue with indefinite weight and robin conditions. Calc Var Partial Differ Equ 55(6):1–37
Li MY, Shuai Z (2010) Global-stability problem for coupled systems of differential equations on networks. J Differ Equ 248(1):1–20
Liang S, Lou Y (2012) On the dependence of population size upon random dispersal rate. Discrete Contin Dyn Syst-B 17(8):2771–2788
Liang X, Zhang L (2021) The optimal distribution of resources and rate of migration maximizing the population size in logistic model with identical migration. Discrete Contin Dyn Syst-B 26(4):2055–2065
Lin K-H, Lou Y, Shih C-W, Tsai T-H (2014) Global dynamics for two-species competition in patchy environment. Math Biosci Eng 11(4):947
Lou Y (2006) On the effects of migration and spatial heterogeneity on single and multiple species. J Differ Equ 223(2):400–426
Lou Y, Lutscher F (2014) Evolution of dispersal in open advective environments. J Math Biol 69(6–7):1319–1342
Lou Y, Yanagida E (2006) Minimization of the principal eigenvalue for an elliptic boundary value problem with indefinite weight, and applications to population dynamics. Jpn J Ind Appl Math 23(3):275–292
Lou Y, Zhou P (2015) Evolution of dispersal in advective homogeneous environment: the effect of boundary conditions. J Differ Equ 259(1):141–171
Lu ZY, Takeuchi Y (1993) Global asymptotic behavior in single-species discrete diffusion systems. J Math Biol 32(1):67–77
Lutscher F, Lewis MA, McCauley E (2006) Effects of heterogeneity on spread and persistence in rivers. Bull Math Biol 68(8):2129–2160
Mazari I (2019) Trait selection and rare mutations: the case of large diffusivities. Discrete Contin Dyn Syst-Ser B
Mazari I, Nadin G, Privat Y (2020) Optimal location of resources maximizing the total population size in logistic models. Journal de mathématiques pures et appliquées 134:1–35
Mazari I, Nadin G, Privat Y (2022) Optimisation of the total population size for logistic diffusive equations: bang-bang property and fragmentation rate. Commun Partial Differ Equ 47(4):797–828
Mazari I, Ruiz-Balet D (2021) A fragmentation phenomenon for a nonenergetic optimal control problem: optimization of the total population size in logistic diffusive models. SIAM J Appl Math 81(1):153–172
Nagahara K, Lou Y, Yanagida E (2021) Maximizing the total population with logistic growth in a patchy environment. J Math Biol 82(1):1–50
Nagahara K, Yanagida E (2018) Maximization of the total population in a reaction-diffusion model with logistic growth. Calc Var Partial Differ Equ 57(3):1–14
Nguyen TD, Wu Y, Veprauskas A, Tang T, Zhou Y, Beckford C, Chau B, Chen X, Rouhani BD, Wu Y, Yang Y, Shuai Z (2023) Maximizing metapopulation growth rate and biomass in stream networks. arXiv preprint arXiv:2306.05555
Pachepsky E, Lutscher F, Nisbet R, Lewis MA (2005) Persistence, spread and the drift paradox. Theor Popul Biol 67(1):61–73
Smith HL (1995) Monotone dynamical systems: an introduction to the theory of competitive and cooperative systems. American Mathematical Society, Providence
Speirs DC, Gurney WSC (2001) Population persistence in rivers and estuaries. Ecology 82(5):1219–1237
Takeuchi Y (1996) Global dynamical properties of Lotka–Volterra systems. World Scientific, Singapore
Vasilyeva O, Lutscher F (2012) How flow speed alters competitive outcome in advective environments. Bull Math Biol 74(12):2935–2958
Wang Y, Shi J, Wang J (2019) Persistence and extinction of population in reaction-diffusion-advection model with strong Allee effect growth. J Math Biol 78(7):2093–2140
Wei J, Liu B (2021) Coexistence in a competition-diffusion-advection system with equal amount of total resources. Math Biosci Eng 18(4):3543–3558
Yan X, Nie H, Zhou P (2022) On a competition-diffusion-advection system from river ecology: mathematical analysis and numerical study. SIAM J Appl Dyn Syst 21(1):438–469
Zhang B, Kula A, Mack KM, Zhai L, Ryce AL, Ni W-M, DeAngelis DL, Van Dyken JD (2017) Carrying capacity in a heterogeneous environment with habitat connectivity. Ecol Lett 20(9):1118–1128
Zhang B, Liu X, DeAngelis DL, Ni W-M, Wang GG (2015) Effects of dispersal on total biomass in a patchy, heterogeneous system: analysis and experiment. Math Biosci 264:54–62
Zhou P, Tang D, Xiao D (2021) On Lotka–Volterra competitive parabolic systems: exclusion, coexistence and bistability. J Differ Equ 282:596–625
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Dedicated to Professor Glenn Webb in honor of his 80th birthday.
Appendix
Appendix
1.1 Results on configuration (ii)
For configuration (ii) we show that if \(\varvec{r}=(r_1, r_2, 0)\) with \(r_1+r_2=r\) and \(\varvec{s}=(s_1, s_2, s_3)\) with \(\sum _{i=1}^3 s_i=r\) and \(s_3\ne 0\) then \(E_1\) is globally asymptotically stable.
Lemma 5.1
Suppose that D and Q are given by (1.3). Let \(\varvec{r}=(r_1, r_2, 0)>\varvec{0}\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) such that \(\sum _{i=1}^2 r_i=\sum _{i=1}^3 s_i=r\) and \(s_3>0\). If \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1), then \(u_1+v_1<k\), \(u_2+v_2<k\), and \(u_3+v_3>k\).
Proof
Suppose that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Let \(w_i:=u_i+v_i\) for \(i=1, 2, 3\). Then, we have
Assume to the contrary that \(w_3\le k\). Then by the third equation of (5.1), we have either \((d+q)w_1-dw_3\le 0\) or \((d+q)w_2-dw_3\le 0\). Without loss of generality, we may assume \((d+q)w_1-dw_3\le 0\). This implies that \(w_1\le dw_3/(d+q)<k\). If \(r_1>0\) or \(s_1>0\), then
which contradicts the first equation of (5.1). If \(r_1=s_1=0\), then \(r_2>0\) and \((d+q)w_1-dw_3= 0\) by the first equation of (5.1). Then by \(w_3\le k\) and the third equation of (5.1) again, we have \((d+q)w_2-dw_3\le 0\). By \(r_2>0\) and the second equation of (5.1), we have \(w_2\ge k\). Therefore,
which is a contradiction. Hence, \(w_3=u_3+v_3>k\).
By \(w_3>k\), either \((d+q)w_1-dw_3>0\) or \((d+q)w_2-dw_3>0\). Without loss of generality, say \((d+q)w_1-dw_3>0\). Then by the first equation of (5.1), we have \(w_1=u_1+v_1<k\). Suppose to the contrary that \(w_2\ge k\). Then by the second equation of (5.1), \((d+q)w_2-dw_3\le 0\). Since \(w_3<(d+q)w_1/d<(d+q)k/d\),
which is a contradiction. Therefore, \(w_2=u_2+v_2<k\). \(\square \)
Then we show the non-existence of a positive equilibrium.
Lemma 5.2
Suppose that D and Q are given by (1.3). Let \(\varvec{r}=(r_1, r_2, 0)>\varvec{0}\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) such that \(\sum _{i=1}^2 r_i=\sum _{i=1}^3 s_i=r>0\) and \(s_3>0\). Then model (1.1) has no positive equilibrium.
Proof
Suppose to the contrary that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Since \(r_3=0\), \((\varvec{u}, \varvec{v})\) satisfies
Adding up the equations in (5.2), we obtain
By Lemma 5.1 and \(r_1+r_2>0\), the left hand side of (5.3) is positive, which is a contradiction. \(\square \)
In the following two lemmas, we show that the semitrivial equilibrium \(E_2\) is unstable.
Lemma 5.3
Suppose that D and Q are given by (1.3). Let \(\varvec{s}=(s_1, s_2, s_3)\ge \varvec{0}\) with \(s_3>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) satisfies \(v^*_1<k\) and \(v^*_2<k\).
Proof
We observe that \(\varvec{v}^*\) must satisfy
That is
Assume to the contrary that \(v_1^*\ge k\). Then by the first equation of (5.4), \((d+q)v^*_1-dv^*_3\le 0\). This implies \(v_3^*\ge (d+q)k/d>k\). By \(s_3>0\) and the third equation of (5.4), either \((d+q)v_1^*-dv^*_3>0\) or \((d+q)v_2^*-dv^*_3>0\). Hence, \((d+q)v_2^*-dv^*_3>0\). Then by the second equation of (5.4), we have \(v_2^*<k\) and
which is a contradiction. Therefore, \(v_1^*<k\). Similarly, \(v_2^*<k\). \(\square \)
Lemma 5.4
Suppose that D and Q are given by (1.3). Let \(\varvec{r}=(r_1, r_2, 0)>\varvec{0}\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) such that \(\sum _{i=1}^2 r_i=\sum _{i=1}^3 s_i=r>0\) and \(s_3>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) is unstable and the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is stable for model (1.1).
Proof
The stability of \(E_2\) is determined by the sign of \(\lambda _1(\varvec{r}, \varvec{v}^*)\), which is the principal eigenvalue of matrix \(dD+qQ+\text {diag}(r_i(1-v^*_i/k))\). By the assumptions on \(\varvec{r}\) and Lemma 5.3, we have
with at least one strict inequality and
Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{r}, \varvec{v}^*)>s(dD+qQ)=0\). Hence, \(E_2\) is unstable.
The stability of \(E_1\) is determined by the sign of \(\lambda _1(\varvec{s}, \varvec{u}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(s_i(1-u^*_i/k))\). By Nguyen et al. (2023), we have \(\varvec{u}^*=(k, k, (d+q)k/d)\). So,
and
since \(s_3>0\). Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{s}, \varvec{u}^*)<s(dD+qQ)=0\). Hence, \(E_1\) is stable. \(\square \)
By the theory of monotone dynamical systems (see Lemma 2.2) and Lemmas 5.2 and 5.4, we obtain the following result:
Theorem 5.5
Suppose that D and Q are given by (1.3). Let \(\varvec{r}=(r_1, r_2, 0)>\varvec{0}\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) such that \(\sum _{i=1}^2 r_i=\sum _{i=1}^3 s_i=r>0\) and \(s_3>0\). Then the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is globally asymptotically stable for model (1.1).
1.2 Results on configuration (iii)
Finally, for configuration (iii) we show that if \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}\ne \varvec{r}\), \(E_1\) is always globally asymptotically stable.
In the following two lemmas, we show that the model has no positive equilibrium.
Lemma 5.6
Suppose that D and Q are given by (1.4). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). If \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1), then \(u_1+v_1<k\) and \(u_3+v_3>k\).
Proof
Suppose that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Let \(w_i=u_i+v_i\) for \(i=1, 2, 3\). Then, we have
Suppose to the contrary that \(w_1\ge k\). Then by the first equation of (2), \((d+q)w_1-dw_2\le 0\). So \(w_2\ge (d+q)k/d>k\). So by the second equation (2), \((d+q)w_2-dw_3\le 0\) and the inequality is strict if \(s_2>0\). Hence, \(w_3\ge (d+q)k/d>k\). If \(s_3>0\), then \((r_3u_3+s_3v_3)(1-{w_3}/{k} )<0\) and the third equation of (2) leads to a contradiction. If \(s_3=0\), by the assumptions on \(\varvec{s}\), \(s_2\ne 0\) and \((d+q)w_2-dw_3< 0\). Then \((r_3u_3+s_3v_3)(1-{w_3}/{k} )\le 0\) and the third equation of (2) gives a contradiction. Therefore, \(w_1=u_1+v_1<k\). A similar argument can be used to prove that \(u_3+v_3>k\).
\(\square \)
Lemma 5.7
Suppose that D and Q are given by (1.4). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). Then model (1.1) has no positive equilibrium.
Proof
Suppose to the contrary that \((\varvec{u}, \varvec{v})\) is a positive equilibrium of (1.1). Since \(r_2=r_3=0\), \((\varvec{u}, \varvec{v})\) satisfies
Adding up the equations in (5.6), we obtain \(ru_1(1-(u_1+v_1)/k)=0\), which implies \(u_1+v_1=k\). This contradicts Lemma 5.6. \(\square \)
In the following two lemmas, we show that semitrivial equilibrium \(E_2\) is unstable.
Lemma 5.8
Suppose that D and Q are given by (1.4). Let \(\varvec{s}=(s_1, s_2, s_3)\ge \varvec{0}\) with \(s_2>0\) or \(s_3>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) satisfies \(v^*_1<k\).
Proof
We observe that \(\varvec{v}^*\) must satisfy
That is
The rest proof is similar to that of Lemma 5.6, so we omit it here. \(\square \)
Lemma 5.9
Suppose that D and Q are given by (1.4). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). Then the semitrivial equilibrium \(E_2=(\varvec{0}, \varvec{v}^*)\) is unstable and the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is stable for model (1.1).
Proof
The stability of \(E_2\) is determined by the sign of \(\lambda _1(\varvec{r}, \varvec{v}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(r_i(1-v^*_i/k))\). By the assumptions on \(\varvec{r}\) and Lemma 5.8, we have
and
Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{r}, \varvec{v}^*)>s(dD+qQ)=0\). Hence, \(E_2\) is unstable.
The stability of \(E_1\) is determined by the sign of \(\lambda _1(\varvec{s}, \varvec{u}^*)\), which is the principal eigenvalue of \(dD+qQ+\text {diag}(s_i(1-u^*_i/k))\). By Nguyen et al. (2023), we have \(\varvec{u}^*=(k, (d+q)k/d, (d+q)^2k/d^2)\). So,
and
with at least one strict sign by the assumption on \(\varvec{s}\). Therefore, by Lemma 3.2, we must have \(\lambda _1(\varvec{s}, \varvec{u}^*)<s(dD+qQ)=0\). Hence, \(E_1\) is stable. \(\square \)
By the theory of monotone dynamical systems (see Lemma 2.2) and Lemmas 5.7 and 5.9, we obtain the following result:
Theorem 5.10
Suppose that D and Q are given by (1.4). Let \(\varvec{r}=(r, 0, 0)\) and \(\varvec{s}=(s_1, s_2, s_3)>\varvec{0}\) with \(\varvec{s}\ne \varvec{r}\) and \(\sum _{i=1}^3 s_i=r>0\). Then the semitrivial equilibrium \(E_1=(\varvec{u}^*, \varvec{0})\) is globally asymptotically stable for model (1.1).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nguyen, T.D., Wu, Y., Tang, T. et al. Impact of resource distributions on the competition of species in stream environment. J. Math. Biol. 87, 62 (2023). https://doi.org/10.1007/s00285-023-01978-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00285-023-01978-6