
Abstract
Colorectal cancer (CRC) is one of the most common cancers and a major cause of cancer-related mortality worldwide. The efficacy of chemotherapy agents in CRC treatment is often limited due to toxic side effects, heterogeneity of cancer cells, and the possibility of chemoresistance which promotes cancer cell survival through several mechanisms. Combining chemotherapy agents with natural compounds like curcumin, a polyphenol compound from the Curcuma longa plant, has been reported to overcome chemoresistance and increase the sensitivity of cancer cells to chemotherapeutics. Curcumin, alone or in combination with chemotherapy agents, has been demonstrated to prevent chemoresistance by modulating various signaling pathways, reducing the expression of drug resistance-related genes. The purpose of this article is to provide a comprehensive update on studies that have investigated the ability of curcumin to enhance the efficacy of chemotherapy agents used in CRC. It is hoped that it can serve as a template for future research on the efficacy of curcumin, or other natural compounds, combined with chemotherapy agents to maximize the effectiveness of therapy and reduce the side effects that occur in CRC or other cancers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Colorectal cancer (CRC) is the third most prevalent form of malignancy and the second main cause of cancer-related mortality worldwide [1]. Various risk factors have been identified that increase the predisposition of individuals to CRC. These include gene mutations, a family history of gastrointestinal cancers, unhealthy diets, excess body weight, diabetes mellitus, intestinal adenomatous polyps, inflammatory bowel disease, smoking, alcohol consumption, and gut microbiome diversity [2, 3]. The mechanisms involved in the pathogenesis of CRC are highly complex and involve several pathways and the accumulation of numerous genetic and epigenetic alterations of the genome [4]. This development spans several years, starting from small neoplastic foci that are difficult to detect, advancing to adenomas, and eventually developing into malignant carcinomas capable of metastasis [5]. Metastasis, the leading responsible cause of cancer deaths, is present in approximately 25% of CRC patients at first diagnosis, with nearly 40% experiencing recurrence even following curative surgery for the primary tumor [6]. The median overall survival rate now reaches 30 months, underscoring the significance of a comprehensive and continuous approach to advanced disease management [7].
Chemotherapy, which involves the use of cytotoxic antineoplastic drugs such as alkylating agents and antimetabolites, has proven effective to treat cancer. Drugs such as 5-fluorouracil (5FU), doxorubicin (DOX), paclitaxel, irinotecan (CPT-11), and cisplatin (CisPt), are widely used for the treatment of various cancer types [8]. However, such drugs are often associated with significant adverse side effects, limited specificity and tolerability, and a narrow therapeutic index [9, 10]. Their effectiveness and use may be restricted in cases where chemoresistance, including multi-drug resistance (MDR), either inherent or acquired, has developed [11]. Acquired chemoresistance is particularly problematic and arises from genetic and/or epigenetic mutations that occur due to the overexpression of therapeutic targets or activation of cancer-promoting pathways. Moreover, tumors are known to exhibit molecular heterogeneity, which contributes to enhanced chemoresistance [12].
A primary objective of cancer treatment is to discover new agents that exhibit high effectiveness and low toxicity, chemosensitizing properties as well as selectivity. Over the past few decades, scientists have shown increased interest in utilizing natural compounds in cancer therapy [13, 14]. Turmeric, from the rhizomes of the Curcuma longa plant, has been traditionally used against various diseases, such as various cancers. Turmeric powder is estimated to contain approximately 77% curcumin (1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione), along with 17% demethoxycurcumin and 5% bisdemethoxycurcumin [15]. Curcumin is a bright orange-yellow crystalline powder and serves as a valuable food ingredient [16]. It is recognized for its biological effects such as antioxidant, anti-inflammatory, antibacterial, anti-rheumatoid arthritis, antidiabetic, anti-Alzheimer's, and antipsoriatic activities [17, 18].
In preclinical studies, it has been shown to enhance the efficacy of various treatments for CRC by targeting multiple cellular pathways [18]. Curcumin modulates a broad range of signaling pathways, including the signal transducer and activator of transcription (STAT), nuclear factor-kappa B (NF-κB), and the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) pathways. It also exerts an effect on the production of reactive oxygen species (ROSs), cyclooxygenase-2 (COX-2), and angiogenesis factors such as matrix metalloproteinase (MMPs) [19, 20]. Curcumin suppresses the expression of p65, thereby inhibiting the transcriptional activity related to NF-κB and suppressing the expression of anti-apoptotic genes regulated by NF-κB [21]. In CRC cells, evidence suggests that curcumin both suppresses NF-κB activation and induces the free radical-mediated Jun N-terminal kinase (JNK), thereby activating the extrinsic and intrinsic apoptotic pathways [22, 23]. Curcumin inhibits the proliferation of CRC cells by regulating the cell cycle machinery. It is responsible for cell cycle arrest in either the G0/G1 or G2/M phase by overexpressing cyclin-dependent kinases (CDK) inhibitors like p16, p21, and p27, while downregulating CDK2, CDK4, cyclin B, E, and D1 [24, 25]. Curcumin can target cancer stem cells (CSCs) via repression of the Wnt/β-catenin and hedgehog signaling pathways, as well as microRNAs [26]. Additionally, it inhibits angiogenesis and metastasis-related markers like angiopoietin-1, vascular endothelial growth factor, and MMP-3, which can assist in suppressing tumor growth [27,28,29].
There is substantial evidence indicating that curcumin when combined with chemotherapeutic agents can enhance treatment efficacy and reduce side effects in patients with CRC, due to its ability to regulate key signaling pathways involved in MDR. This review aimed to discuss the potential benefits of curcumin in improving the effectiveness of widely used chemotherapy agents with a focus on 5FU, oxaliplatin (OXA), CisPt, 5FU plus OXA plus folinic acid (FOLFOX), dasatinib, CPT-11, capecitabine, DOX, and vincristine (VCR) through diverse mechanisms of action.
Methodology
Online literature databases such as Web of Science, PubMed, and Scopus, were used to search for relevant journal articles using the keyword “Curcumin AND Colorectal Neoplasm OR Colorectal Tumor OR Colorectal OR Colorectal Cancer OR Colorectal Carcinoma OR Colonic Neoplasm OR Colon Cancer OR Colonic Cancer OR Colon Adenocarcinoma OR chemoresistance OR chemotherapy”. Among the 9965 articles found, 134 were initially selected after screening based on their titles and abstracts. Subsequently, the full text of each original article was studied, and 50 articles were retained. Reviews, book chapters, and non-English articles were removed. Studies that reported on the use of synthetic curcumin, nanoformulations and other formulations of curcumin, and cancers other than CRC were also excluded (Fig. 1).

Flow chart of the methodology used. WoS Web of science, 5FU 5-fluorouracil, OXA Oxaliplatin, CisPt Cisplatin, FOLFOX 5FU plus OXA plus folinic acid, CPT-11 Irinotecan, DOX Doxorubicin, VCR Vincristine
Chemoresistance in CRC
Chemoresistance, which refers to the capacity of tumor cells to resist or adapt to therapeutic agents, presents a significant obstacle in oncology research. Extensive efforts have been made to understand and overcome this phenomenon by elucidating the molecular mechanisms that promote cancer cell survival and apoptosis evasion when exposed to conventional chemotherapy agents. These mechanisms encompass diverse signaling transduction pathways activated in response to numerous stimuli, ultimately leading to the development of chemoresistance, including MDR.
Drug efflux transportation
The effectiveness of chemotherapeutic agents relies on their successful penetration into cells, achieving an optimal intracellular concentration. In cancer cells, ATP-binding cassette (ABC) transporters can expel chemotherapeutic agents from the cells, reducing the intracellular accumulation of the drugs and the cytotoxic effects [30]. In chemoresistant CRC cells, ABCA1, ABCA2, and ABCA5 are also upregulated [31]. ABCA2, and ABCA5 located in the lysosomal membrane can induce chemoresistance by increasing drug sequestration in the lysosome and facilitating drug efflux [32]. P-glycoprotein (P-gp), also named multi-drug resistance protein 1 (MDR1), is the product of the ABC subfamily B, member 1 gene, and holds particular significance among ABC transporters in the human gastrointestinal system [33]. Previous studies have shown that P-gp expression is upregulated in CRC, which is associated with increased chemoresistance and chemotherapy failure [34,35,36]. Indeed, cancerous cells can confer expression of P-gp through the upregulation of oncogenic kinases such as NF-κB, mitogen-activated protein kinase (MAPK), and extracellular-signal-regulated kinase (ERK) signaling, thereby enhancing chemoresistance. Du and colleagues demonstrated that interleukin-8 activates the NF-κ B signaling pathway, thereby stimulating MDR1 expression, which induces resistance to DOX [36]. Furthermore, Wnt signaling enhances cytoplasmic Ca2+ concentration and causes nuclear and cell membrane depolarization, leading to the translocation of β-catenin into the nucleus. This process increases ABCA1 gene expression and enhances 5FU resistance in CRC [35]. In addition, by attenuating the intracellular accumulation of chemotherapeutics, these transporters can affect the pharmacokinetics properties of drugs, including biodistribution, metabolism, and cytotoxicity [37, 38].
Apoptosis evasion
Another characteristic feature of cancer is the capacity of cancerous cells to evade physiological cell death or apoptosis, particularly in response to treatments like chemotherapy. Apoptosis is induced following DNA damage and disturbance of cellular organelles, such as mitochondria and the endoplasmic reticulum, through intrinsic or extrinsic pathways [39]. Resistance to apoptosis can arise from the upregulation of anti-apoptotic factors and the downregulation of pro-apoptotic factors, which significantly contribute to resistance to treatment [40]. In DOX-resistant CRC cells, the expression of the anti-apoptotic protein B cell lymphoma-2 (Bcl-2) significantly increased, while the expression of P53 and Bax significantly decreased, indicating resistance to apoptosis [41]. However, the suppression of Bax expression decreased the sensitivity of CRC cells to 5FU and OXA [42].
NF-κB is considered a crucial connection between apoptosis, inflammation, and therapeutic resistance. It controls the expression of several genes, including survivin, Bcl-xL, Bcl-2, COX-2, cyclinD1, P53, and Fas, which play significant roles in the regulation of apoptosis in cancerous cells [43, 44]. Indeed, the activated NF- κB binds to DNA and acts as a transcription factor for the expression of oncogenes, inhibiting apoptosis and facilitating cancer cell proliferation. In colon cancer cells treated with 5FU, Wnt/β-catenin signaling potentially reduced the rate of apoptosis [45]. In fact, β-catenin significantly increased the expression of anti-apoptotic proteins in colon cancer [46]. Furthermore, Wnt1 suppressed the caspase-9 activated by chemotherapeutic agents [47]. Additionally, other downstream genes of Wnt are also involved in chemoresistance through the suppression of apoptosis; MMP-7 inhibited the activation of Fas receptor, an apoptosis stimulator, in colon cancer cells, which led to increased resistance to OXA [48]. Furthermore, P38 MAPK is also involved in chemoresistance by regulating of apoptosis. Inhibition of P38 MAPK reduced drug-induced apoptosis, subsequently increasing 5FU resistance [49]. Numerous strategies are currently being developed to target these pathways and counteract chemoresistance mechanisms in cancer, offering a promising avenue for further investigation.
Cell cycle progression
Tumor cells have a propensity to accumulate changes in the components of the cell-division machinery, disrupting their ability to DNA damage response. This disruption often involves arresting the progression of the cell cycle, which represents a characteristic feature of cancer. To evade cancer treatments, cancer cells frequently overactivate cyclins specifically altering cyclin D, which accelerates cancer cell proliferation [50]. Simultaneously, this uncontrolled cell cycle progression is closely linked to the downregulation of CDK inhibitors such as p21 and p27 [51]. Yu and colleagues demonstrated that disruption of cell cycle regulation is associated with resistance to OXA. The increased expression of Cell division cycle 7, attributed to the dysregulation of the polo-like kinase1 -MYC signaling pathway, results in the activation of CSCs and oncogenic transformation, thereby enhancing resistance to OXA in CRC [52]. In another study, the deficiency of smad4 in SW620 cells activated the PI3K/Akt/CDC2/surviving cascade, leading to reduce G2-M cell cycle arrest, and subsequently induced 5FU resistance [53]. The dysregulation of signaling pathways that stimulate growth, including the transforming growth factor-β (TGF-β), PI3K/ Akt/ mTOR signaling pathway, the epidermal growth factor receptor (EGFR), and NF-κB, during cell cycle progression can lead to chemoresistance [54]. Taking into account the significance of NF-κB, particularly in mechanisms related to MDR, it is crucial to consider its role in relation to cyclin/CDK complexes [55]. Indeed, NF-κB contributes to resistance by regulating the genes involved in the cell cycle and expression of anti-apoptotic proteins such as Bcl-2, X-linked inhibitor of apoptosis protein, and Bcl-xL.
Epithelial-to-mesenchymal transition (EMT)
EMT is a dynamic reversible phenomenon, which the cells experience a phenotypic shift from an epithelial state to a mesenchymal state, which is marked by the disruption of intercellular connections such as tight junctions, adherence junctions, desmosomes, and gap junctions [56]. This is associated with a reduced expression of genes coding for epithelial markers like zonula occludens-1, claudins, E-cadherin, and desmoplakin and overexpression of mesenchymal markers like vimentin, fibronectin, α-smooth muscle actin, and N-cadherin [57, 58]. There is substantial evidence that the induction of the EMT phenotype contributes to chemotherapy resistance. In fact, EMT has the capacity to induce changes in the TME, subsequently creating conditions that promote chemotherapy resistance in CRC cells [59]. Chemoresistant CRC cells exhibit high expression levels of ZEB1, vimentin, MMP2, and MMP9, while E-cadherin expression shows a significant decrease [60]. In this context, resistant cells to CPT-11 demonstrate decreased E-cadherin, and zonula occludens-1 expression alongside increased N-cadherin, and vimentin expression. Additionally, there is a significant increase in Twist, a transcription factor, expression, indicating the induction of the EMT phenotype [61]. In chemoresistant CRC, the Wnt signaling pathway is able to induce EMT phenotype through the activation of MDR1 [34]. Furthermore, Snail, the transcription factor regulating EMT, binds to the ABCB1 and increases its expression, thereby contributing to increased chemotherapy resistance [62]. AXL, a tyrosine kinase receptor, contributed in chemotherapy resistance in CRC by upregulating MDR1 and inducing the EMT phenotype through the increased expression of Twist1 [63]. Furthermore, EMT can be induced by TGF-β, and intracellular signaling pathways like PI3K and NF-κB in CRC [64, 65]. Cancerous cells that have undergone EMT not only acquire the capacity to migrate and invade surrounding tissues but also exhibit increased MDR. However, the precise underlying mechanisms for this resistance are not completely understood.
Cancer stem cell generation
According to the CSC theory, tumor initiation is predominantly driven by a subpopulation of cells called CSCs, which may contribute to cancer initiation, progression, metastasis, treatment resistance, recurrence, and worse prognosis [66]. Similar to non-stem-derived tumor cells, CSCs have a unique capacity for self-renewal and generate non-stem daughter cells that constitute the bulk of the tumor mass [67].
There is considerable evidence that CSCs are implicated in the metastasis and recurrence of colorectal tumors, contributing to increased resistance to apoptosis and chemotherapy [68]. Many chemotherapeutic agents target dividing cells; therefore, CSCs, due to their quiescent nature, evade the effects of these drugs [68]. CSCs increase the expression of anti-apoptotic proteins and, in addition, significantly enhance the rates of mitophagy and oxidative phosphorylation in mitochondria, consequently correlating with increased chemoresistance in CRC [69]. One study found that intervention with diHEP-DPA, combined with 5FU, reduced CSCs activation and suppressed tumor-associated macrophages infiltration, thereby improving 5FU resistance in CRC [70]. Exosomes secreted by fibroblasts enhance chemotherapy resistance in CRC patients by stimulating a CSCs-like phenotype through Wnt signaling activation [71]. Furthermore, the neurogenic locus notch homolog protein-1 (Notch-1) signaling pathway is implicated in chemotherapy resistance in CRC, increasing the number of colonospheres, generating chemoresistant cells, and inducing a CSCs-like phenotype [72]. PI3K activation is associated with chemoresistance in CRC. Metastasis-associated colon cancer 1 (MACC1) related to PI3K/AKT in terms of enhancing of chemoresistance which in turn significantly increases the expression of MDR1, CSCs markers include CD44, CD133, and Nanog, and sphere formation, thereby enhancing resistance to 5FU in colon cells [73]. Given the leading edge of CSCs in tumor maintenance and MDR, targeting and eliminating these cells represents a promising strategy to combat the development of drug resistance in the context of treating cancer.
Efficacy of curcumin combined with chemotherapy agents in CRC treatment
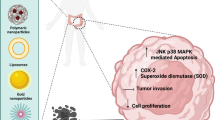
Despite the wide use of chemotherapy agents such as 5FU, OXA, and FOLFOX in CRC treatment, the effectiveness of these drugs is hindered by significant cytotoxic side effects and the increasing emergence of MDR in metastatic patients. To address these limitations, researchers and clinicians are now exploring the co-administration of conventional chemotherapy drugs with curcumin (Fig. 2). This approach seeks to improve the therapeutic outcomes by harnessing the synergistic antitumor effects of combining these agents (Table 1).

Summarized the effect of curcumin on different mechanisms of chemoresistance in CRC. Abbreviations CDK2 Cyclin-dependent kinases2, Notch-1 Neurogenic locus notch homolog protein-1, EMT Epithelial-to-mesenchymal transition, MACC1 Metastasis-associated colon cancer1, EZH2 Enhancer of zeste homolog 2, SUZ12, Suppressor of zeste 12, BMI1 B cell-specific Moloney murine leukemia virus integration site1, CSCs Cancer stem cells, TETs Ten–eleven translocation methylcytosine dioxygenases, Oct ¾, CD44 Cluster of differentiation 44, ALDH1 Aldehyde dehydrogenase1, CD133 Cluster of differentiation 133, Bcl-2 B cell lymphoma-2, Bax Bcl-2-associated protein x, Bcl-xl B-cell lymphoma-extra-large, PARP Poly-ADP ribose polymerase, Nrf2 Nuclear factor erythroid2–related factor2, P-gp P-glycoprotein, HER Human epidermal growth factor receptor, EGFR Epidermal growth factor receptor, IGF-1 Insulin-like growth factor-1, IGFBP3 IGF-binding protein3, NF-κB Nuclear factor-kappa B, CXCL CXC chemokine ligand, COX-2 Cyclooxygenase-2, TNF-α Tumor necrosis factor-α, STAT1 Signal transducer and activator of transcription1, ERK Extracellular signal-regulated kinase
Curcumin enhances the efficacy of 5FU in CRC treatment
5FU was discovered in the late 1950s and is used as an anticancer drug that functions as an antimetabolite by mimicking the nucleobase uracil with a fluorine atom at the C-5 position. After entry into cells, it is transformed into biologically active metabolites that inhibit cancer cell growth by disrupting DNA synthesis through the suppression of thymidylate synthase, an enzyme involved in nucleotide synthesis. By inhibiting the formation of thymidine, 5FU inhibits cell cycle progression, by targeting the S phase. When 5FU is incorporated into RNA, instead of uracil nucleotides, this leads to the inhibition of RNA transcription necessary for protein synthesis. The induction of apoptosis is also an important mechanism underlying the toxic effects of 5FU [124,125,126].
Previous studies have revealed that curcumin in combination with 5FU could potentially reduce resistance to 5FU. In particular, pre-treatment with curcumin (5 µM) raised the sensitivity of 5FU (0.1 M) and reversed MDR in DNA mismatch repair (MMR)-deficient CRC models [74]. As the activation of the NF-κB pathway has been shown to enhance chemoresistance to drugs, NF-κB inhibitors have the potential to increase the efficacy of chemotherapy. It has been demonstrated that curcumin boosts 5FU sensitivity in CRC cells resistant to 5FU by inhibiting NF-κB signaling and NF-κB-regulated genes expression. Indeed, the activation of inflammatory pathways, such as the NF-kB and PI-3K/Src pathways, by 5FU stimulates survival responses that lead to MDR and potentially contribute to the development of resistance. Curcumin has been reported to counteract this effect by increasing apoptosis in CRC cells resistant to 5FU, ultimately enhancing the efficacy of 5FU. In addition, curcumin can synergize the effect of 5FU on apoptosis via upregulation of pro-apoptotic proteins including Bax, caspase-8, caspase-9, caspase-3, and poly ADP ribose polymerase (PARP), while downregulating the anti-apoptotic protein Bcl-xL and the proliferative protein cyclin D1 [75, 76]. The activation of apoptotic pathways was achieved by hampering the activation of IKK and IκBa phosphorylation [75]. In 5FU-resistant HCT-8 cells, a combination of 5FU (10 mM) with curcumin (10 µM) has been shown to reduce the mRNA expression of nuclear factor erythroid2–related factor 2 (Nrf2), a transcription factor related to cell survival and chemoresistance, inhibit the Bcl-2/Bax ratio, and induce apoptosis, ultimately reversing resistance to 5FU and MDR [78]. Additionally, curcumin (5 µM) has been reported to increase ROSs generation, thereby facilitating apoptosis and consequently promoting chemosensitivity in CRC cells with resistance to 5FU [79]. Recently, Zheng et al. showed that low concentration of curcumin (10 µM) in combination with 5FU (5 µM) could inhibit cell proliferation and induce apoptosis in SW620 cells via suppression of pERK and STAT1 and reduced expression of L1 [90].
Overexpression of the COX-2 protein contributes to the upregulation of Bcl-2, resulting in the suppression of tumor cell apoptosis and the development of chemoresistance [127]. The combination of curcumin (20 µM) with 5FU (50 µM) and halogenated hydrocarbon curcumin (HHC) (25 µM) with 5FU (5 µM) have been demonstrated to synergistically inhibit the growth of HT-29 cells. These combination remarkably downregulated COX-2 expression by almost sixfold compared to each agent alone. As a result, it enhanced the anticancer effects of 5FU in HT-29 cells [82, 83]. In an in vivo study, orally treatment of HHC (50 mg/kg) in combination with intraperitoneal injection of 5FU (50 mg/kg) significantly reduced the number and size of aberrant crypt foci. Furthermore, HHC enhanced the inhibitory effects of 5FU on CRC progression by increasing apoptosis and downregulating of COX-2 [84]. NF‐κB through activation of COX‐2 leads to produce prostaglandins and promotes apoptosis resistance on cancerous cells. Furthermore, curcumin in combination with 5FU potentially led to downregulation of P-gp and heat shock protein 27 (HSP-27) expression [86]. The ability of this combination to sensitize resistance to 5FU may be attributed to the increasing expression of ATP-dependent topoisomerase II and the reduction in survivin, p-AKT in the NF-κB pathway, and Bcl-2 expression, which induce apoptosis [77]. The toxicity profile was improved when co-treating with curcumin, as it reduced the concentration of 5FU required to inhibit the proliferation of tumor cells in a 3D model which was achieved via inhibiting arresting the cell cycle at G0/G1 phase through degradation of cyclin D1 [86, 89]. Another study focusing on the cell cycle observed that combination of curcumin (5 µM) with 5FU (1 µM) arrested cell cycle at S phase and subsequent activated both the extrinsic and the intrinsic apoptotic pathways in 5FU-resistant HCT116 cells [75]. The combined treatment of curcumin with 5FU inhibited cell proliferation and induced apoptosis in 5FU-resistant and parental SW480 and HCT116 cells. Moreover, curcumin (10 µM) augmented the anticancer effects of 5FU (10 µM) in cancerous cells by attenuating EMT due to downregulation of transcript factors involved in EMT such as B lymphoma Mo-MLV insertion region 1 homolog, suppressor of zeste12 and Enhancer of zeste homolog2 [87]. A recent study showed that curcumin was a potential agent to suppress CRC cell growth and increased sensitivity to 5FU by inhibiting EMT through the reduction of MACC1 expression, which was associated with the activation of the PI3K/AKT/mTOR pathway [88].
Curcumin significantly inhibited multiple pathways associated with CSCs, suggesting its potential as an adjunctive treatment to enhance the effectiveness of existing therapies while reducing adverse side effects [128]. The administration of curcumin (5 µM) prior to 5FU (0.1 µM) treatment resulted in the modification of the interaction between tumor cells and fibroblasts. This modification led to the inhibition of EMT and the promotion of chemosensitivity in CSCs by reducing the expression of NF-κB, MMP-13, TGF-β, and EMT markers [91]. This created a favorable environment for increasing chemotherapeutic action of 5FU, particularly targeting CSCs resistance to 5FU. Curcumin (5 µM) also suppressed the CSC sub-population and downregulation of CRC markers including CD133, CD44, and aldehyde dehydrogenase 1 (ALDH1) expression, leading to enhanced sensitivity to 5FU in DNA of MMR-deficient CRC cells [74].
Curcumin can induce epigenetic alterations, like modulating methylation of DNA and miRNA expression, leading to enhanced chemosensitivity of CRC cells to 5FU treatment [87, 129]. Lu et al. revealed that curcumin effectively halted the progression of EMT and inhibited Wnt signaling by hindering the expression of ten–eleven translocation methylcytosine dioxygenase1 (TET1), naked cuticle homolog2 (NKD2), and vimentin. Furthermore, curcumin decreased the levels of β-catenin, E-cadherin, transcription factor 4, and axin in HCT116 cells, which are implicated in chemoresistance. As a result, curcumin has the potential to counteract resistance to 5FU in HCT116 cells by regulating the TET1-NKD2-WNT signaling pathway and reversing the advancement of EMT [89]. In particular, It has been found that curcumin (10 µM) exerted the chemosensitve effects on CRC cell resistance to 5FU through regulation of miRNAs involved in suppression of the EMT process, including miR-200b, miR-200c, miR-141, miR-429, and miR-101 [87]. Additionally, curcumin degraded miR-21, which leads to inhibition of the CRC tumor’s ability to invade and spread. It was also able to inhibit miR-130a expression to block Wnt/β-Catenin signaling [130]. The presence of curcuminoids resulted in a decrease in drug resistance and hindered the growth of SW480. This was achieved by suppressing the Sp, pro-oncogenic transcription factors, and their associated genes, like MDR1. The downregulation of Sp factors was facilitated by disrupting the miR-27a:ZBTB10 axis, and this impact was reliant on the presence of ROS [85]. Thus, it seems that epigenetic modifications might be a crucial mechanism for curcumin-mediated chemoresistance.
It has been shown showed that curcumin (20 µM) inhibits the proliferation and migration of chemoresistant CRC cells. This effect was attributed to a substantial downregulation of insulin receptor, insulin-like growth factor-1 receptor (IGF-1R), and MYC. Consequently, curcumin was able to reverse the chemoresistance to 5FU in SW480 cell lines [81]. New evidence suggests that Nicotinamide N-methyltransferase (NNMT) plays a significant role in cancer progression, apoptosis inhibition, and drug resistance. Recently, Li et al. demonstrated that curcumin inhibits cell proliferation, arrests the cell cycle at the G2 phase, and increases the production of free radicals, leading to increased drug sensitivity in CRC cell lines with high NNMT expression [80]. Therefore, emerging evidence strongly supports the therapeutic potential of combining curcumin and 5FU to improve CRC treatment.
Curcumin enhances the efficacy of platinum-based compounds in CRC treatment
Platinum-based agents like CisPt, and OXA, carboplatin are predominantly used for the treatment of solid neoplasms, including CRC. These drugs exert their effects by forming covalent bonds with DNA purine bases, resulting in the formation of platinum–DNA adducts. As such, the replication and transcription of DNA is inhibited, ultimately causing apoptosis [131, 132].
OXA is commonly used as a first-line treatment for CRC and various other malignancies [99, 133]. Its mechanism of action involves the generation of ROS in physiological solutions, which leads to the formation of covalent bonds between platinum and DNA. As a result, inter- and intra-strand platinum DNA adducts are formed, primarily between neighboring guanines or adenines and guanines [134]. These adducts disrupt crucial cellular processes, ultimately leading to cell death related to DNA damage. The treatment of cancer with OXA is usually associated with the emergence of chemoresistant-related relapses [105]. To enhance the effectiveness of OXA in cancer chemotherapy, it would be advantageous to identify a safe adjuvant that can improve therapeutic outcomes without causing additional toxicity [135]. Several studies have revealed that a combination of OXA and curcumin has synergistic effects and can improve the anticancer effects of OXA and overcome resistance in CRC both in vivo and in vitro. It has been revealed that curcumin enhanced the effects of OXA in damaging DNA and inducing apoptosis in cancer cells, attributed to the augmentation of platinum–DNA binding [92]. Indeed, curcumin increased the cytotoxic effects of platinum (II)-based agents, including OXA, CisPt, and carboplatin on chemoresistant CRC cells. Its combination with platinum agents reversed chemoresistance. The increase in apoptosis rate, arrested cell cycle progression in the G2/M phase, as well as inhibited the expression of platinum (II)-based agents-induced nuclear NF-kB were attributed to this effect [93, 94]. The combination of OXA (2 µM) and curcumin (5 µM) enhanced the efficacy of OXA by downregulating Ki-67, Notch-1 pathway in HT-29 and HCT116 cells. Such a combination makes tumors OXA-sensitive [95]. In addition, this combination exerted synergistic apoptotic effects by increasing p53 and Bax expression and decreasing Bcl-2 expression [94, 96]. Additionally, the intraperitoneal injection of OXA (50 mg/kg) and curcumin (25 mg/kg) reduced the proliferative capacity of OXA- resistant CRC cells. In a HCT116 nude mouse xenograft tumor model, curcumin significantly enhanced the antitumor effectiveness of OXA. This combination caused a notable enhanced apoptosis in tumor cells, along with upregulation of Bax, caspase-3, and PARP expression, while suppressing Bcl-2, survivin, HSP70, pro-caspase-3, and pro-PARP expression in colorectal tumors [97].
Several investigations have clarified that curcumin could reverse MDR via hampering NF-κB signaling responsible for the downregulation of genes related to chemoresistance and anti-apoptosis [98, 136]. Moreover, chemokines such as CXC motif chemokine ligand1 (CXCL1) and CXCL8 are considered as autocrine tumor-promoting factors and can enhance NF-κB transcriptional activity [137]. Curcumin in combination with OXA has been reported to downregulate the expression of CXCL1 and CXCL8 genes in an OXA-resistant HTOXAR3 cell line, resulting in a substantial reduction in NF-κB signaling, and decrease in the IC50 of OXA, thereby sensitizing the cells to OXA. These findings suggest a potential role for these chemokines in acquired resistance to OXA through activation of the AKT/NF-κB pathway. Therefore, the levels of chemokines, specifically CXCL1, were suggested as a potential predictive indicator for the efficacy of this combined therapy in CRC [98]. Moreover, a co-treatment with curcumin (8 µM) reversed OXA (4 µM) resistance in CRC cells through the inhibition of EMT markers such as TGF-β, Smad-2, and N-cadherin [99].
The excision repair cross-complementing gene (ERCC1) plays a vital role in the nucleotide repair process, indicating its involvement in tumor resistance to platinum drugs [138]. Additionally, there is evidence indicating that the miR-409-3p-mediated regulation of ERCC1 might be associated with resistance to OXA in CRC cells. Recently, Han et al. demonstrated that curcumin (30 µM) effectively reduces the expression of ERCC1, Bcl-2, glutathione s-transferase-π (GST-π), multidrug resistance-related protein, P-gp, and survivin. Additionally, curcumin increases the expression of miR-409-3p and enhances the apoptotic rate of resistant cells. These findings suggest that curcumin has the ability to reverse drug resistance to OXA by modulating miR-409-3p-mediated ERCC1 expression [100].
CisPt is another common cancer chemotherapy drug based on platinum that has been proven to improve survival rates in patients undergoing chemotherapy [139]. CisPt is a cytotoxic substance that primarily acts by interacting with proteins containing a HMG domain and cross-link to tumor cell DNA. The formation of adducts between the drug and cancer cells disrupts cell division. Consequently, the DNA is irreversibly damaged, leading to cell cycle arrest and apoptosis [140]. The ability of cell apoptosis was significantly enhanced after treatment with curcumin and CisPt, as evidenced by the increased expression of apoptotic proteins such as Bax, while the anti-apoptotic proteins like Bcl-2 and the Notch-1 pathways were suppressed [102]. There is evidence indicating that long non-coding RNAs (lncRNAs) are involved in chemotherapeutic resistance. The expression of one such lncRNA, KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1), is significantly increased in CRC tissues [141, 142]. It has been proposed that KCNQ1OT1 is also implicated in resistance to CisPt, something that is attributed to enhanced proliferation and inhibited apoptosis through the miR-497/Bcl-2 axis, thereby exacerbating resistance to CisPt [103]. Zheng demonstrated that curcumin (10 µM) reverses resistance to CisPt by downregulating KCNQ1OT1 expression [103]. Furthermore, curcumin increased the expression of miR-137. Subsequently, miR inhibits the metabolism of glutamine by targeting glutaminase, resulting in suppression of the proliferation of chemoresistant CRC cells and increasing sensitivity to CisPt [104].
Curcumin enhances the efficacy of FOLFOX in CRC treatment
The standard chemotherapeutic regimen for CRC, called FOLFOX, combines folinic acid with 5FU and OXA. This treatment is associated with severe side effects, and often results in poor outcomes, leading to cancer recurrence [105]. Studies have shown that almost half of all diagnosed CRC patients treated with first-line FOLFOX chemotherapy will experience cancer recurrence, and around 40% of non-resectable patients with colorectal liver metastases do not respond to the administration of FOLFOX [134]. According to a study involving colon cancer cells, the percentage of CD44-positive cells, which indicates the presence of CSCs, was found to be 0.04% in wild-type cells. However, in CRC cells resistant to FOLFOX, the percentage of CD44-positive cells increased by tenfold [105]. Curcumin, whether used alone or in conjunction with FOLFOX, shows potential for effectively eliminating CSCs in CRC. Curcumin in conjunction with FOLFOX was found to reduce the expression of transcription factors related to CSC-like features, such as octamer-binding transcription factor3/4, Nanog, Otx2, and AFP [121].
When used alone or in combination with FOLFOX, curcumin not only downregulated several markers associated with CSCs and EGFR – but also remarkably reduced colony and tumor sphere formation of CRC cells resistant to FOLFOX. In fact, reduced EGFR promoter methylation was responsible for upregulation of EGFR in CRC cells resistant to FOLFOX. However, treatment with curcumin (20 µM) in 5FU-resistant cells exerted opposite effects which was correlated with DNA methyltransferase1 expression [105]. These findings suggest that the induction of specific growth factor receptors such as EGFR can assist the growth and survival of CSC, leading to the increased possibility of developing malignancies [105]. Extensive research has provided substantial evidence regarding the significance of the IGF signaling pathway in the initiation and progression of various cancers which has emerged as a crucial prognostic factor. It activates pathways within colorectal epithelial cells that facilitate transformation, including both apoptosis and chemoresistance signaling pathways [143, 144]. Curcumin (10 µM) in combination with FOLFOX was reported to considerably suppress cell growth and induce apoptosis in HCT116 and HT-29 cells. This was accompanied by a downregulation of EGFR, human epidermal growth factor receptor-2 (HER-2), HER-3, IGF-1R, PI3K/Akt, and COX-2 and accompanied with a fivefold increase in the expression of IGF-binding protein-3 (IGFBP-3). This increase in IGFBP-3 facilitated the sequestration of IGF, making IGF-1 unavailable for binding and activation of IGF-1R [106]. Furthermore, curcumin suppressed protein inhibitor of activated STAT3, a crucial factor in maintaining of IGF-1R in the nucleus and preventing translocation of IGF-1R to the nucleus [101]. Consequently, these co-treatment can be associated with the prohibition of signaling pathways involving EGFRs and IGF-1R [101, 106]. In this context, the IGF-1/IGF-1R axis could represent promising therapeutic targets for the prevention and treatment of CRC [18]. Notably, FOLFOX when combined with curcumin (25 µM) greatly attenuated the survival of CRC cells, followed by a simultaneous decrease of EGFR, HER-2, IGF-1R, AKT activation, as well as downregulation of COX-2 and cyclin-D1 expression [107].
Curcumin enhances the efficacy of other chemotherapy agents in CRC treatment
Dasatinib was discovered as a potent Src/Abl kinases inhibitor, potentially exhibiting anti-proliferative effects against various cancer cell lines [145]. Dasatinib (1 μM) in conjunction with curcumin (10 μM) also inhibits the activation of HER-2, HER-3 and IGF-1R signaling, resulting in a decrease in the phosphorylation levels of AKT and Erks (1/2). Further results on the downregulation of anti-apoptotic proteins such as Bcl-XL and of COX-2 indicated its possible efficacy in CRC treatment. [108]. The synergistic combination of dasatinib (1 μM) and curcumin (10 μM) effectively inhibits cell growth, migration, invasion, and formation of colonospheres. This combination was also reported to reduce the population of CSCs, as indicated by downregulation of CSC markers including CD133, CD44, CD166, and ALDH [109].
CPT-11 and its active metabolite, 7-ethyl-10-hydroxycamptothecin, exert their anticancer effects by inhibiting DNA topoisomerase, leading to DNA double-strand breaks and triggering a cellular response to DNA damage. However, CPT-11-based chemotherapy for CRC is associated with the development of enterotoxigenesis and chemoresistance [146]. Studies on the potential synergistic effect of curcumin with CPT-11 showed increased therapeutic efficacy of CPT-11 in human CRC cells. The combination of CPT-11 with curcumin inhibited cell proliferation, arrested the cell cycle, and promoted apoptosis. The latter was attributed, at least in part, to the generation of ROS, the stimulation of the endoplasmic reticulum-based cellular stress response and the upregulation of binding immunoglobulin protein and CCAAT/enhancer-binding protein homologous protein [110]. In addition to reducing growth, curcumin significantly reduced the colon CSCs’ population. The co-treatment of curcumin (5 μM) with CPT-11 (100 μM) noticeably suppressed the expression of CSC-associated markers, including CD44, CD133, CD24 and EpCAM, leading to substantial induction of apoptosis in colon CSCs and reversal of chemoresistance [111].
Zhang et al. demonstrated that co-treatment of curcumin (5 μg) and CPT-11 (20 μg) significantly reduced cell proliferation, induced apoptosis and reversed drug resistance in CPT-11-resistant CRC cells by inhibiting the EMT process via increased expression of E-cadherin, and reduced expression of vimentin and N-cadherin [112]. Antioxidant enzymes such as GSTs can scavenge ROSs and detoxify toxic agents. In oxidative stress situations, GSTs are activated to protect the body's cell DNA against ROSs. However, evidence suggests that in chemoresistant cells, the level of GSTs is increased, potentially playing a protective role against chemotherapy agents [147]. Furthermore, the formation of Glutathione (GSH)-chemotherapeutic agents enhances intracellular agent solubility, leading to drug efflux from the cell, thereby inducing resistance to chemotherapy in cancer cells. It has been revealed that curcumin reduces the expression of GST M5 and prevents CPT-11 efflux from the cell, consequently reversing chemotherapy resistance by increasing cellular apoptosis [113].
Curcumin (1 g/kg, orally) in combination with capecitabine (60 mg/kg, orally)effectively suppressed metastasis in nude mice by attenuating the activation of the NF-κB pathway and NF-κB-regulated genes such as cyclin D1, c-MYC, Bcl-2, Bcl-xL, COX-2, and MMP-9, which resulted in increased susceptibility of CRC cells to the anticancer and antimetastatic activities of capecitabine [114].
Curcumin was reported to reverse resistance to DOX, a common chemotherapy agent used for a wide range of cancers including CRC, by decreasing cellular viability [115] via suppressing caspase-3, -8, and -9 activation in CRC cells [115, 148]. The treatment of polyphenols, including curcumin, tannic acid, and epigallocatechin gallate with DOX attenuated chemoresistance. This attenuation was associated with the suppression of P-gp activity, which could render CRC cells sensitive to DOX [118]. Curcumin, in combination with DOX, was able to inhibit COX-2-induced P-gp expression, and by increasing intracellular DOX concentration, led to decreased cellular proliferation and increased apoptosis. Additionally, curcumin (10 μM) directly interacts with certain mitochondrial matrix proteins like C1qbp, disrupting oxidative phosphorylation and ATP synthesis, thus preventing the advancement of chemoresistance in CRC [116]. Alteration of metabolic processes are implicated in the onset and progression of MDR [149]. The concentration of polyamines such as spermine, spermidine, and putrescine significantly augments in chemoresistant cells. This increase leads to heightened activation of tricarboxylic acid cycle cycles, generating more ATP, thereby providing the energy required for P-gp function. Additionally, polyamines enhance the synthesis of antioxidant enzymes like GSH, which protects DNA against ROSs and promotes chemoresistance [150, 151]. Besides, D-glutamine metabolism induces chemoresistance by increasing P-gp activation and GSH expression. Recently, Zhang and colleagues indicated that the combination of curcumin (5.5 μM) with DOX (0.5 μM) was capable of inhibiting polyamine biosynthesis and D-glutamine metabolism, subsequently reducing P-gp function due to decreased ATP availability. Thus, DOX accumulation in cells and cytotoxic effects of it against cancer cells are strengthened [117].
Curcumin (˃25 μM) enhances the inhibitory effect of VCR on the growth of CRC cells. It suppressed the expression of the P-gp gene and its activity, significantly reducing the efflux of VCR from the cells while increasing its intracellular concentration. Moreover, the combination of curcumin and VCR prevented tumor growth and reduced the expression of the anti-apoptotic protein such as survivin. Consequently, curcumin diminished chemoresistant CRC cells and augmented their sensitivity to 5FU, VCR, and CisPt [119]. As mentioned, chemoresistant cells utilize the host's antioxidant defenses to mitigate the cytotoxic effects of chemotherapy. Curcumin (25 μM) reduced resistance to VCR by downregulating pyridoxine 6 and GSTP1 while increasing apoptosis [120].
Clinical trials
Several clinical trials have investigated the effects of curcumin on advanced CRC. Sharma and colleagues investigated the safety and Pharmacokinetic/Pharmacodynamics of curcumin in advanced CRC patients. No toxic doses were observed after 16 weeks. Although the intervention with 36 mg of curcumin for 4 weeks resulted in a significant reduction in GST activity, a similar reduction was not observed at higher doses per 28 weeks. Furthermore, curcumin or its metabolites were not detected in urine and plasma after 8 weeks of intervention. Low doses of curcumin approximately 519 nmol/g and high doses of curcumin 1054 nmol/g of curcumin were detected in dry feces [152]. In another study, the administration of 3.6 g/day curcumin in patients with colon cancer reduced Tumor necrosis factor-α (TNF-α) concentration while increasing the expression of P53 and Bax in cancer cells, which was associated with an increase in apoptosis [153]. However, clinical studies investigating the effectiveness of combining curcumin with chemotherapy agents in CRC patients have been limited to date. Curcumin (2 g/day, orally) has been reported to enhance the effectiveness of FOLFOX in CRC patients with liver metastasis [121]. Additionally, It was found to be safe and well-tolerated [121]. It has been established that abdominal pain, dyspepsia, and nausea were the most common side effects of curcumin treatment. In this context, the combination of curcumin (2 g/day, orally) with FOLFOX significantly improved overall survival (Hazard ratio = 0.34), although it had no effect on progression-free survival and quality of life [122].Another phase II study using curcumin in CRC patients undergoing chemoradiotherapy demonstrated that curcumin (4 g, orally for 6 weeks) did not increase disease progression [123].
However, current chemotherapeutic agents exhibit high toxicity and significant side effects; therefore, finding safe natural compounds with minimal side effects as adjunctive therapies for chemotherapy is essential. The efficacy of curcumin in combination with chemotherapy in humans have been conducted, or still ongoing. Currently, clinical trials are exploring the effects of curcumin formulations in conjunction with chemotherapeutic agents in CRC. Jeon et al. indicated that nanoformulations of curcumin combined with bevacizumab/FOLFIRI improved progression-free survival in CRC patients with metastases. Additionally, this combination was found to be safe and well-tolerated [154]. NCT01859858 assessed the safety and pharmacokinetics of CPT-11 and curcumin in colon cancer patients. Another clinical trial, NCT02724202 is examining the safety of 5FU in combination with BCM-95, synthetic curcuminoid, in patients with metastatic colon cancer. However, the conducted studies have several limitations, including small sample sizes, different doses of curcumin and its derivatives, short follow-up durations, and the lack of control for confounding factors.
Curcumin and immunotherapy
Immune cells exhibit variations in their phenotypes and functions over time, thereby facilitating the immune microenvironment to possess both a "tumor- suppressing" and "tumor-promoting" effect as a whole [155]. However, cancer cells can hinder the functions of anti-tumor T cells, which reduces tumor infiltration and compromised cytotoxicity and proliferation [156]. One of the most significant challenges in cancer treatment is the evasion tumor cells from the immune response. In fact, they can escape the host immune response by increasing the population of regulatory T (Treg) cells and disrupting the function of CD8+ T and natural killer cells [157]. Therefore, the suppression of Treg cells is considered a potential strategy to enhance the efficacy of immunotherapy. It has been suggested that the combination of natural compounds such as curcumin with immunotherapy is associated with improved immune responses against cancer cells. Curcumin downregulated programmed cell death ligand 1 expression in a manner that enhances the effects of immunotherapy against cancer [158, 159]. It significantly augmented the expression of CD8+ T cells while downregulating Treg cells [160]. In a clinical study, supplementation with curcumin in 40 patients with colon cancer reduced the population of Treg cells, while increasing the T Helper1 cell population. Furthermore, it increased the interferon-γ produced by Treg cells which facilitates the conversion of Treg cells to T Helper1 cells [161]. Based on these findings, curcumin appears to be an adjunctive compound for immunotherapy; however, additional preclinical and clinical studies are necessary to evaluate the efficacy of this combination.
Conclusion and future prospects
Finding an effective chemotherapeutic approach to combat cancer remains challenging. The toxicity to both cancerous and non-cancerous cells of conventional chemotherapy agents and the presence of chemoresistance are two main contributing factors. Combining chemotherapy agents with curcumin has been identified as a promising approach for the treatment of CRC. Furthermore, Chemotherapy-related toxicity is considered one of the most main limiting factors in cancer treatment, leading to treatment inefficacy and discontinuation. For instance, fluoropyrimidine agents are associated with side effects such as stomatitis, diarrhea, nausea, and neuropathy [162]. Gastrointestinal symptoms, hematological abnormalities, and hepatotoxicity are among the most important adverse effects of OXA [162]. There is substantial evidence indicating that, in addition to increasing sensitivity to chemotherapy agents, curcumin has potential effects on certain chemotherapy-related side effects. It has been shown to protect against OXA-induced liver damage by reducing oxidative stress through the activation of Nrf2 [163]. The suppression of NF-κB activation by curcumin alleviated intestinal and cardiovascular damage [164]. It has been observed curcumin could attenuate gastrointestinal symptoms induced by chemotherapy agents [162].
Although many in vitro and in vivo studies have demonstrated the beneficial effects of curcumin in CRC, its bioavailability poses a significant challenge in clinical studies. In other words, its low water solubility and conversion into inactive metabolites result in reduced Pharmacokinetic/Pharmacodynamics, leading to potential limitations of curcumin in clinical settings. Therefore, structural modifications of the curcumin scaffold and the use of various nanocarrier formulations for drug delivery have already been explored to overcome the low oral bioavailability and enhance its Pharmacokinetic/Pharmacodynamics and improve its efficacy of curcumin [165,166,167]. Future work should aim to evaluate the optimal safe dose of curcumin required to exert activity, the optimal formulation to ensure good oral bioavailability, and the long-term toxicity of combinations of curcumin with selected chemotherapy agents. This will provide better insights into the role of curcumin in limiting the undesirable side effects of conventional anticancer drugs, reversing chemoresistance, and its potential for improving the quality of life for CRC patients.
Data availability
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Abbreviations
- CRC:
-
Colorectal cancer,
- 5FU:
-
5-Fluorouracil
- DOX:
-
Doxorubicin
- CPT-11:
-
Irinotecan
- CisPt:
-
Cisplatin
- MDR:
-
Multi-drug resistance
- STAT:
-
Signal transducer and activator of transcription
- NF-κB:
-
Nuclear factor-kappa B
- PI3K/ Akt:
-
Phosphoinositide 3-kinase/protein kinase B
- mTOR:
-
Mammalian target of rapamycin
- ROSs:
-
Reactive oxygen species
- COX-2:
-
Cyclooxygenase-2
- MMP:
-
Matrix metalloproteinase
- JNK:
-
Jun N-terminal kinase
- CDKs:
-
Cyclin-dependent kinases
- CSCs:
-
Cancer stem cells
- OXA:
-
Oxaliplatin
- FOLFOX:
-
5FU plus OXA plus folinic acid
- VCR:
-
Vincristine
- ABC:
-
ATP-binding cassette
- P-gp:
-
P-glycoprotein
- MDR1:
-
Multi-drug resistance protein 1
- MAPK:
-
Mitogen-activated protein kinase
- ERK:
-
Extracellular-signal-regulated kinase
- Bcl-2:
-
B cell lymphoma-2, Bcl-xl: B-cell lymphoma-extra-large
- Bax:
-
Bcl-2-associated protein x
- TGF-β:
-
Transforming growth factor-β
- EGFR:
-
Epidermal growth factor receptor
- EMT:
-
Epithelial-to-mesenchymal transition
- Notch:
-
Neurogenic locus notch homolog protein-1
- MACC1:
-
Metastasis-associated colon cancer 1
- MMR:
-
Mismatch repair
- PARP:
-
Poly ADP ribose polymerase
- Nrf2:
-
Nuclear factor erythroid2–related factor 2
- HHC:
-
Halogenated hydrocarbon curcumin
- HSP:
-
Heat shock protein
- ALDH1:
-
Aldehyde dehydrogenase1
- TET1:
-
Ten–eleven translocation methylcytosine dioxygenase1
- NKD2:
-
Naked cuticle homolog2
- IGF-1R:
-
Insulin-like growth factor-1 receptor
- NNMT:
-
Nicotinamide N-methyltransferase
- CXCL1:
-
CXC motif chemokine ligand 1
- ERCC1:
-
Excision repair cross-complementing gene
- lncRNAs:
-
Long non-coding RNAs
- KCNQ1OT1:
-
KCNQ1 opposite strand/antisense transcript 1
- HER:
-
Human epidermal growth factor receptor
- IGFBP-3:
-
IGF-binding protein-3
- GST:
-
Glutathione S-transferase
- GSH:
-
Glutathione
- TNF-α:
-
Tumor necrosis factor-α
- Treg cell:
-
Regulatory T cell
References:
Morgan E, Arnold M, Gini A, Lorenzoni V, Cabasag C, Laversanne M, et al. Global burden of colorectal cancer in 2020 and 2040: Incidence and mortality estimates from GLOBOCAN. Gut. 2023;72(2):338–44.
Rawla P, Sunkara T, Barsouk A. Epidemiology of colorectal cancer: incidence, mortality, survival, and risk factors. Prz Gastroenterol. 2019;14(2):89–103.
Keum N, Giovannucci E. Global burden of colorectal cancer: emerging trends, risk factors and prevention strategies. Nat Rev Gastroenterol Hepatol. 2019;16(12):713–32.
Fatemi N, Tierling S, Es HA, Varkiani M, Mojarad EN, Aghdaei HA, et al. DNA methylation biomarkers in colorectal cancer: Clinical applications for precision medicine. Int J Cancer. 2022;151(12):2068–81.
Kumar Pandurangan A, Divya T, Kumar K, Dineshbabu V, Velavan B, Sudhandiran G. Colorectal carcinogenesis: Insights into the cell death and signal transduction pathways: a review. World J Gastrointest Oncol. 2018;10(9):244.
Van Cutsem E, Oliveira J. Advanced colorectal cancer: ESMO clinical recommendations for diagnosis, treatment and follow-up. Ann Oncol. 2009;20:61–3.
Venook AP, Niedzwiecki D, Lenz H-J, Innocenti F, Fruth B, Meyerhardt JA, et al. Effect of first-line chemotherapy combined with cetuximab or bevacizumab on overall survival in patients with KRAS wild-type advanced or metastatic colorectal cancer: a randomized clinical trial. JAMA. 2017;317(23):2392–401.
Tan BL, Norhaizan ME. Curcumin combination chemotherapy: The implication and efficacy in cancer. Molecules. 2019;24(14):2527.
Khan AQ, Ahmed EI, Elareer N, Fathima H, Prabhu KS, Siveen KS, et al. Curcumin-mediated apoptotic cell death in papillary thyroid cancer and cancer stem-like cells through targeting of the JAK/STAT3 signaling pathway. Int J Mol Sci. 2020;21(2):438.
Negrette-Guzman M. Combinations of the antioxidants sulforaphane or curcumin and the conventional antineoplastics cisplatin or doxorubicin as prospects for anticancer chemotherapy. Eur J Pharmacol. 2019;859: 172513.
Hsu HH, Chen MC, Baskaran R, Lin YM, Day CH, Lin YJ, et al. Oxaliplatin resistance in colorectal cancer cells is mediated via activation of ABCG2 to alleviate ER stress induced apoptosis. J Cell Physiol. 2018;233(7):5458–67.
Sa G, Das T. Anti cancer effects of curcumin: cycle of life and death. Cell Div. 2008;3:1–14.
Chan W-JJ, Adiwidjaja J, McLachlan AJ, Boddy AV, Harnett JE. Interactions between natural products and cancer treatments: Underlying mechanisms and clinical importance. Cancer Chemother Pharmacol. 2023.
Nan Y, Su H, Zhou B, Liu S. The function of natural compounds in important anticancer mechanisms. Front Oncol. 2023;12:1049888.
Anderson AM, Mitchell MS, Mohan RS. Isolation of curcumin from turmeric. J Chem Educ. 2000;77(3):359.
Lestari ML, Indrayanto G. Curcumin. Profiles of drug substances, excipients and related methodology. 2014;39:113–204.
Patel SS, Acharya A, Ray R, Agrawal R, Raghuwanshi R, Jain P. Cellular and molecular mechanisms of curcumin in prevention and treatment of disease. Crit Rev Food Sci Nutr. 2020;60(6):887–939.
Selvam C, Prabu SL, Jordan BC, Purushothaman Y, Umamaheswari A, Zare MSH, et al. Molecular mechanisms of curcumin and its analogs in colon cancer prevention and treatment. Life Sci. 2019;239: 117032.
Shishodia S. Molecular mechanisms of curcumin action: gene expression. BioFactors. 2013;39(1):37–55.
Oliviero F, Scanu A, Zamudio-Cuevas Y, Punzi L, Spinella P. Anti-inflammatory effects of polyphenols in arthritis. J Sci Food Agric. 2018;98(5):1653–9.
Collett GP, Campbell FC. Overexpression of p65/RelA potentiates curcumin-induced apoptosis in HCT116 human colon cancer cells. Carcinogenesis. 2006;27(6):1285–91.
Thayyullathil F, Chathoth S, Hago A, Patel M, Galadari S. Rapid reactive oxygen species (ROS) generation induced by curcumin leads to caspase-dependent and-independent apoptosis in L929 cells. Free Radic Biol Med. 2008;45(10):1403–12.
Agarwal A, Kasinathan A, Ganesan R, Balasubramanian A, Bhaskaran J, Suresh S, et al. Curcumin induces apoptosis and cell cycle arrest via the activation of reactive oxygen species–independent mitochondrial apoptotic pathway in Smad4 and p53 mutated colon adenocarcinoma HT29 cells. Nutr Res. 2018;51:67–81.
Dasiram JD, Ganesan R, Kannan J, Kotteeswaran V, Sivalingam N. Curcumin inhibits growth potential by G1 cell cycle arrest and induces apoptosis in p53-mutated COLO 320DM human colon adenocarcinoma cells. Biomed Pharmacother. 2017;86:373–80.
Choudhuri T, Pal S, Das T, Sa G. Curcumin selectively induces apoptosis in deregulated cyclin D1-expressed cells at G2 phase of cell cycle in a p53-dependent manner. J Biol Chem. 2005;280(20):20059–68.
Zhu JY, Yang X, Chen Y, Jiang Y, Wang SJ, Li Y, et al. Curcumin suppresses lung cancer stem cells via inhibiting Wnt/β-catenin and sonic hedgehog pathways. Phytother Res. 2017;31(4):680–8.
Li W, Jiang Z, Xiao X, Wang Z, Wu Z, Ma Q, et al. Curcumin inhibits superoxide dismutase-induced epithelial-to-mesenchymal transition via the PI3K/Akt/NF-κB pathway in pancreatic cancer cells. Int J Oncol. 2018;52(5):1593–602.
Shakeri A, Ward N, Panahi Y, Sahebkar A. Anti-angiogenic activity of curcumin in cancer therapy: a narrative review. Curr Vasc Pharmacol. 2019;17(3):262–9.
Ramezani M, Hatamipour M, Sahebkar A. Promising anti-tumor properties of bisdemethoxycurcumin: a naturally occurring curcumin analogue. J Cell Physiol. 2018;233(2):880–7.
Ivanyuk A, Livio F, Biollaz J, Buclin T. Renal drug transporters and drug interactions. Clin Pharmacokinet. 2017;56:825–92.
Ravindranathan P, Pasham D, Goel A. Oligomeric proanthocyanidins (OPCs) from grape seed extract suppress the activity of ABC transporters in overcoming chemoresistance in colorectal cancer cells. Carcinogenesis. 2019;40(3):412–21.
Pasello M, Giudice AM, Scotlandi K. The ABC subfamily A transporters: multifaceted players with incipient potentialities in cancer. Semin Cancer Biol. 2020.
Krishna R, Mayer LD. Multidrug resistance (MDR) in cancer: mechanisms, reversal using modulators of MDR and the role of MDR modulators in influencing the pharmacokinetics of anticancer drugs. Eur J Pharm Sci. 2000;11(4):265–83.
Zhou H, Lin C, Zhang Y, Zhang X, Zhang C, Zhang P, et al. miR-506 enhances the sensitivity of human colorectal cancer cells to oxaliplatin by suppressing MDR 1/P-gp expression. Cell Prolif. 2017;50(3): e12341.
Wang T, Chen Z, Zhu Y, Pan Q, Liu Y, Qi X, et al. Inhibition of transient receptor potential channel 5 reverses 5-Fluorouracil resistance in human colorectal cancer cells. J Biol Chem. 2015;290(1):448–56.
Du J, He Y, Li P, Wu W, Chen Y, Ruan H. IL-8 regulates the doxorubicin resistance of colorectal cancer cells via modulation of multidrug resistance 1 (MDR1). Cancer Chemother Pharmacol. 2018;81:1111–9.
Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, Snyder N, et al. Drug resistance in cancer: an overview. Cancers. 2014;6(3):1769–92.
Robinson K, Tiriveedhi V. Perplexing role of P-glycoprotein in tumor microenvironment. Front oncol. 2020;10:265.
Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495–516.
Razaghi A, Heimann K, Schaeffer PM, Gibson SB. Negative regulators of cell death pathways in cancer: perspective on biomarkers and targeted therapies. Apoptosis. 2018;23:93–112.
Hu T, Wang L, Zhang L, Lu L, Shen J, Chan RL, et al. Sensitivity of apoptosis-resistant colon cancer cells to tanshinones is mediated by autophagic cell death and p53-independent cytotoxicity. Phytomedicine. 2015;22(5):536–44.
Hu T, Li Z, Gao C-Y, Cho CH. Mechanisms of drug resistance in colon cancer and its therapeutic strategies. World J Gastroenterol. 2016;22(30):6876.
Cao F, Liu T, Xu Y, Xu D, Feng S. Curcumin inhibits cell proliferation and promotes apoptosis in human osteoclastoma cell through MMP-9, NF-κB and JNK signaling pathways. Int J Clin Exp Pathol. 2015;8(6):6037–45.
Tsai J-R, Liu P-L, Chen Y-H, Chou S-H, Cheng Y-J, Hwang J-J, et al. Curcumin inhibits non-small cell lung cancer cells metastasis through the adiponectin/NF-κb/MMPs signaling pathway. PLoS ONE. 2015;10(12): e0144462.
Wu X, Luo F, Li J, Zhong X, Liu K. Tankyrase 1 inhibitior XAV939 increases chemosensitivity in colon cancer cell lines via inhibition of the Wnt signaling pathway. Int J Oncol. 2016;48(4):1333–40.
Zhang T, Otevrel T, Gao Z, Gao Z, Ehrlich SM, Fields JZ, et al. Evidence that APC regulates survivin expression: a possible mechanism contributing to the stem cell origin of colon cancer. Cancer Res. 2001;61(24):8664–7.
Chen S, Guttridge DC, You Z, Zhang Z, Fribley A, Mayo MW, et al. Wnt-1 signaling inhibits apoptosis by activating β-catenin/T cell factor–mediated transcription. J Cell Biol. 2001;152(1):87.
Almendro V, Ametller E, García-Recio S, Collazo O, Casas I, Augé JM, et al. The role of MMP7 and its cross-talk with the FAS/FASL system during the acquisition of chemoresistance to oxaliplatin. PLoS ONE. 2009;4(3): e4728.
Micallef I, Baron B. The mechanistic roles of ncRNAs in promoting and supporting chemoresistance of colorectal cancer. Non-coding RNA. 2021;7(2):24.
Otto T, Sicinski P. Cell cycle proteins as promising targets in cancer therapy. Nat Rev Cancer. 2017;17(2):93–115.
Abukhdeir AM, Park BH. P21 and p27: roles in carcinogenesis and drug resistance. Expert Rev Mol Med. 2008;10: e19.
Yu Z, Deng P, Chen Y, Liu S, Chen J, Yang Z, et al. Inhibition of the PLK1-coupled cell cycle machinery overcomes resistance to oxaliplatin in colorectal cancer. Adv Sci. 2021;8(23):2100759.
Zhang B, Leng C, Wu C, Zhang Z, Dou L, Luo X, et al. Smad4 sensitizes colorectal cancer to 5-fluorouracil through cell cycle arrest by inhibiting the PI3K/Akt/CDC2/survivin cascade. Oncol Rep. 2016;35(3):1807–15.
Panda M, Biswal BK. Cell signaling and cancer: a mechanistic insight into drug resistance. Mol Biol Rep. 2019;46:5645–59.
Yang L, Zhou Y, Li Y, Zhou J, Wu Y, Cui Y, et al. Mutations of p53 and KRAS activate NF-κB to promote chemoresistance and tumorigenesis via dysregulation of cell cycle and suppression of apoptosis in lung cancer cells. Cancer Lett. 2015;357(2):520–6.
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial–mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–96.
Liu Y. Epithelial to mesenchymal transition in renal fibrogenesis: pathologic significance, molecular mechanism, and therapeutic intervention. J Am Soc Nephrol. 2004;15(1):1–12.
Thiery JP, Sleeman JP. Complex networks orchestrate epithelial–mesenchymal transitions. Nat Rev Mol Cell Biol. 2006;7(2):131–42.
Ni Q, Li M, Yu S. Research progress of epithelial-mesenchymal transition treatment and drug resistance in colorectal cancer. Technol Cancer Res Treat. 2022;21:15330338221081220.
Guo C, Ma J, Deng G, Qu Y, Yin L, Li Y, et al. ZEB1 promotes oxaliplatin resistance through the induction of epithelial-mesenchymal transition in colon cancer cells. J Cancer. 2017;8(17):3555.
Yang Y, Wang G, Zhu D, Huang Y, Luo Y, Su P, et al. Epithelial-mesenchymal transition and cancer stem cell-like phenotype induced by Twist1 contribute to acquired resistance to irinotecan in colon cancer. Int J Oncol. 2017;51(2):515–24.
Wang H, Li JM, Wei W, Yang R, Chen D, Ma XD, et al. Regulation of ATP-binding cassette subfamily B member 1 by snail contributes to chemoresistance in colorectal cancer. Cancer Sci. 2020;111(1):84–97.
Solanes-Casado S, Cebrian A, Rodríguez-Remírez M, Mahíllo I, García-García L, Rio-Vilarino A, et al. Overcoming PLK1 inhibitor resistance by targeting mevalonate pathway to impair AXL-TWIST axis in colorectal cancer. Biomed Pharmacother. 2021;144: 112347.
Joyce T, Cantarella D, Isella C, Medico E, Pintzas A. A molecular signature for Epithelial to Mesenchymal transition in a human colon cancer cell system is revealed by large-scale microarray analysis. CClin Exp Metastasis. 2009;26:569–87.
Suman S, Kurisetty V, Das TP, Vadodkar A, Ramos G, Lakshmanaswamy R, et al. Activation of AKT signaling promotes epithelial–mesenchymal transition and tumor growth in colorectal cancer cells. Mol Carcinog. 2014;53(S1):E151–60.
Mondal S, Bhattacharya K, Mandal C. Nutritional stress reprograms dedifferention in glioblastoma multiforme driven by PTEN/Wnt/Hedgehog axis: a stochastic model of cancer stem cells. Cell Death Discov. 2018;4(1):110.
Zhao J. Cancer stem cells and chemoresistance: The smartest survives the raid. Pharmacol Ther. 2016;160:145–58.
Ashique S, Bhowmick M, Pal R, Khatoon H, Kumar P, Sharma H, et al. Multi drug resistance in colorectal cancer-approaches to overcome, advancements and future success. Adv Cancer Biol: Metastasis. 2024.
de Almeida RM, Siqueira PB, de Amorim ÍSS, Mencalha AL, Thole AA. Mitochondria in colorectal cancer stem cells-a target in drug resistance. Cancer Drug Resist. 2023;6(2):273.
Su Y, Choi HS, Choi JH, Kim H-S, Jang Y-S, Seo J-W. 7 S, 15 R-Dihydroxy-16 S, 17 S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model. Mar Drugs. 2023;21(2):80.
Hu Y-B, Yan C, Mu L, Mi YL, Zhao H, Hu H, et al. Exosomal Wnt-induced dedifferentiation of colorectal cancer cells contributes to chemotherapy resistance. Oncogene. 2019;38(11):1951–65.
Huang R, Wang G, Song Y, Tang Q, You Q, Liu Z, et al. Colorectal cancer stem cell and chemoresistant colorectal cancer cell phenotypes and increased sensitivity to Notch pathway inhibitor. Mol Med Rep. 2015;12(2):2417–24.
Wang J, Wang W, Cai H, Du B, Zhang L, Ma W, et al. MACC1 facilitates chemoresistance and cancer stem cell-like properties of colon cancer cells through the PI3K/AKT signaling pathway. Mol Med Rep. 2017;16(6):8747–54.
Shakibaei M, Buhrmann C, Kraehe P, Shayan P, Lueders C, Goel A. Curcumin chemosensitizes 5-fluorouracil resistant MMR-deficient human colon cancer cells in high density cultures. PLoS ONE. 2014;9(1): e85397.
Shakibaei M, Mobasheri A, Lueders C, Busch F, Shayan P, Goel A. Curcumin enhances the effect of chemotherapy against colorectal cancer cells by inhibition of NF-κB and Src protein kinase signaling pathways. PLoS ONE. 2013;8(2): e57218.
Shakibaei M, Kraehe P, Popper B, Shayan P, Goel A, Buhrmann C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer. 2015;15(1):1–15.
Fan Y, Abulimiti P, Zhang H, Zhou Y, Zhu L. Mechanism of reversal of multidrug resistance by curcumin in human colorectal cancer cell line HCT-8/5-FU. Genet Mol Res. 2017.
Zhang C, He LJ, Ye HZ, Liu DF, Zhu YB, Miao DD, et al. Nrf2 is a key factor in the reversal effect of curcumin on multidrug resistance in the HCT-8/5-Fu human colorectal cancer cell line. Mol Med Rep. 2018;18(6):5409–16.
Yang J, He C, Liu N. Proteomic analysis of the chemosensitizing effect of curcumin on CRC cells treated with 5-FU. Front Med. 2022.
Li G, Fang S, Shao X, Li Y, Tong Q, Kong B, et al. Curcumin reverses NNMT-induced 5-fluorouracil resistance via increasing ROS and cell cycle arrest in colorectal cancer cells. Biomolecules. 2021;11(9):1295.
Hosseini SA, Zand H, Cheraghpour M. The influence of curcumin on the downregulation of MYC, insulin and IGF-1 receptors: a possible mechanism underlying the anti-growth and anti-migration in chemoresistant colorectal cancer cells. Medicina. 2019;55(4):90.
Du B, Jiang L, Xia Q, Zhong L. Synergistic inhibitory effects of curcumin and 5-fluorouracil on the growth of the human colon cancer cell line HT-29. Chemotherapy. 2005;52(1):23–8.
Srimuangwong K, Tocharus C, Chintana PY, Suksamrarn A, Tocharus J. Hexahydrocurcumin enhances inhibitory effect of 5-fluorouracil on HT-29 human colon cancer cells. World J Gastroenterol. 2012;18(19):2383.
Srimuangwong K, Tocharus C, Tocharus J, Suksamrarn A, Chintana PY. Effects of hexahydrocurcumin in combination with 5-fuorouracil on dimethylhydrazine-induced colon cancer in rats. World J Gastroenterol. 2012;18(47):6951–9.
Noratto GD, Jutooru I, Safe S, Angel-Morales G, Mertens-Talcott SU. The drug resistance suppression induced by curcuminoids in colon cancer SW-480 cells is mediated by reactive oxygen species-induced disruption of the micro RNA-27a-ZBTB 10-S p axis. Mol Nutr Food Res. 2013;57(9):1638–48.
He W-t, Zhu Y-h, Zhang T, Abulimiti P, Zeng F-y, Zhang L-p, et al. Curcumin reverses 5-fluorouracil resistance by promoting human colon cancer HCT-8/5-FU cell apoptosis and down-regulating heat shock protein 27 and P-glycoprotein. Chin J Integr Med. 2019.
Toden S, Okugawa Y, Jascur T, Wodarz D, Komarova NL, Buhrmann C, et al. Curcumin mediates chemosensitization to 5-fluorouracil through miRNA-induced suppression of epithelial-to-mesenchymal transition in chemoresistant colorectal cancer. Carcinogenesis. 2015;36(3):355–67.
Ma X, Sun X, Wang R, Guo Y, Xu M. Effects of curcumin on 5-fluorouracil resistance of colon cancer cells through the PI3K/AKT/mTOR pathway via MACC1. Eur J Integr Med. 2022;56: 102202.
Lu Y, Zhang R, Zhang X, Zhang B, Yao Q. Curcumin may reverse 5-fluorouracil resistance on colonic cancer cells by regulating TET1-NKD-Wnt signal pathway to inhibit the EMT progress. Biomed Pharmacother. 2020;129: 110381.
Zheng X, Yang X, Lin J, Song F, Shao Y. Low curcumin concentration enhances the anticancer effect of 5-fluorouracil against colorectal cancer. Phytomedicine. 2021;85: 153547.
Buhrmann C, Kraehe P, Lueders C, Shayan P, Goel A, Shakibaei M. Curcumin suppresses crosstalk between colon cancer stem cells and stromal fibroblasts in the tumor microenvironment: potential role of EMT. PLoS ONE. 2014;9(9): e107514.
Bali H, Yu JQ, Beale P, Huq F. Dose Dependent Synergism from Combination of Platinum Drugs with Curcumin against Colorectal Cancer Cell Lines. J Angiother. 2020;4(1):E176–93.
Wang YT, Liu HS, Su CL. Curcumin-enhanced chemosensitivity of FDA-approved platinum (II)-based anti-cancer drugs involves downregulation of nuclear endonuclease G and NF-κB as well as induction of apoptosis and G2/M arrest. Int J Food Sci Nutr. 2014;65(3):368–74.
Howells LM, Mitra A, Manson MM. Comparison of oxaliplatin-and curcumin-mediated antiproliferative effects in colorectal cell lines. Int J Cancer. 2007;121(1):175–83.
Howells LM, Sale S, Sriramareddy SN, Irving GR, Jones DJ, Ottley CJ, et al. Curcumin ameliorates oxaliplatin-induced chemoresistance in HCT116 colorectal cancer cells in vitro and in vivo. Int J Cancer. 2011;129(2):476–86.
Zhang N, Hao YF, Liu HS, Yu QQ, Bo B, Liang JW. Combined anti-cancer effects of curcumin and oxaliplatin on colon carcinoma colo205 cells using transplanted nude mice. Pak J Pharm Sci. 2021;34(5):2021–5.
Guo Ld, Yq S, Zhao Xh, Guo Lj Yu, Zj WD, et al. Curcumin combined with oxaliplatin effectively suppress colorectal carcinoma in vivo through inducing apoptosis. Phytother Res. 2015;29(3):357–65.
Ruiz de Porras V, Bystrup S, Martínez-Cardús A, Pluvinet R, Sumoy L, Howells L, et al. Curcumin mediates oxaliplatin-acquired resistance reversion in colorectal cancer cell lines through modulation of CXC-Chemokine/NF-κB signalling pathway. Sci Rep. 2016.
Yin J, Wang L, Wang Y, Shen H, Wang X, Wu L. Curcumin reverses oxaliplatin resistance in human colorectal cancer via regulation of TGF-β/Smad2/3 signaling pathway. Onco Targets Ther. 2019;12:3893.
Han W, Yin H, Ma H, Wang Y, Kong D. Curcumin regulates ERCC1 expression and enhances oxaliplatin sensitivity in resistant colorectal cancer cells through its effects on miR-409-3p. Evid Based Complement Alternat Med. 2020;2020:8394574.
Codony-Servat J, Cuatrecasas M, Asensio E, Montironi C, Martínez-Cardús A, Marín-Aguilera M, et al. Nuclear IGF-1R predicts chemotherapy and targeted therapy resistance in metastatic colorectal cancer. Br J Cancer. 2017;117(12):1777–86.
Chen GP, Zhang Y, Xu ZY, Yu JF, Wei X. Curcumin combined with cis-platinum promote the apoptosis of human colorectal cancer HT29 cells and mechanism. Int J Clin Exp Pathol. 2017;10(12):11496–505.
Zheng Z-h, You H-y, Feng Y-j, Zhang Z-t. LncRNA KCNQ1OT1 is a key factor in the reversal effect of curcumin on cisplatin resistance in the colorectal cancer cells. Mol Cell Biochem. 2021;476:2575–85.
Fan WH, Wang FC, Jin Z, Zhu L, Zhang JX. Curcumin synergizes with cisplatin to inhibit colon cancer through targeting the microRNA-137-glutaminase axis. Curr Med Sci. 2022;42(1):108–17.
Yu Y, Kanwar SS, Patel BB, Nautiyal J, Sarkar FH, Majumdar AP. Elimination of colon cancer stem-like cells by the combination of curcumin and FOLFOX. Transl Oncol. 2009;2(4):321–8.
Patel BB, Sengupta R, Qazi S, Vachhani H, Yu Y, Rishi AK, et al. Curcumin enhances the effects of 5-fluorouracil and oxaliplatin in mediating growth inhibition of colon cancer cells by modulating EGFR and IGF-1R. Int J Cancer. 2008;122(2):267–73.
Patel BB, Gupta D, Elliott AA, Sengupta V, Yu Y, Majumdar AP. Curcumin targets FOLFOX-surviving colon cancer cells via inhibition of EGFRs and IGF-1R. Anticancer Res. 2010;30(2):319–25.
Nautiyal J, Banerjee S, Kanwar SS, Yu Y, Patel BB, Sarkar FH, et al. Curcumin enhances dasatinib-induced inhibition of growth and transformation of colon cancer cells. Int J Cancer. 2011;128(4):951–61.
Nautiyal J, Kanwar SS, Yu Y, Majumdar AP. Combination of dasatinib and curcumin eliminates chemo-resistant colon cancer cells. J Mol Signal. 2011;6(1):1–11.
Huang Y-F, Zhu D-J, Chen X-W, Chen Q-K, Luo Z-T, Liu C-C, et al. Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget. 2017;8(25):40264.
Su PF, Yang Y, Wang GX, Chen XW, Ju YL. Curcumin attenuates resistance to irinotecan via induction of apoptosis of cancer stem cells in chemoresistant colon cancer cells. Int J Oncol. 2018;53(3):1343–53.
Zhang C, Xu Y, Wang H, Li G, Yan H, Fei Z, et al. Curcumin reverses irinotecan resistance in colon cancer cell by regulation of epithelial–mesenchymal transition. Anticancer Drugs. 2018;29(4):334–40.
Zhu DJ, Chen XW, Wang JZ, Ju YL, Yang MZO, Zhang WJ. Proteomic analysis identifies proteins associated with curcumin-enhancing efficacy of irinotecan-induced apoptosis of colorectal cancer LOVO cell. Int J Clin Exp Pathol. 2014;7(1):1–15.
Kunnumakkara AB, Diagaradjane P, Anand P, Kuzhuvelil HB, Deorukhkar A, Gelovani J, et al. Curcumin sensitizes human colorectal cancer to capecitabine by modulation of cyclin D1, COX-2, MMP-9, VEGF and CXCR4 expression in an orthotopic mouse model. Int J Cancer. 2009;125(9):2187–97.
Khameneh ZR, Mohammadian M, Rasouli MA, Moradi Z, Ahmadi Z, Khiyavi AA. Effects of curcumin in combination with doxorubicin in human colorectal cancer cell line. Asian Pac j cancer biol. 2018;3(4):89–92.
Jayarajan J, Angandoor S, Vedulla SH, Sritharan S, Ganesan K, War AR, et al. Curcumin induces chemosensitization to doxorubicin in Duke’s type B coloadenocarcinoma cell line. Mol Biol Rep. 2020;47(10):7883–92.
Zhang N, Gao M, Wang Z, Zhang J, Cui W, Li J, et al. Curcumin reverses doxorubicin resistance in colon cancer cells at the metabolic level. J Pharm Biomed Anal. 2021;201: 114129.
Li HM, Krstin S, Wink M. Modulation of multidrug resistant in cancer cells by EGCG, tannic acid and curcumin. Phytomedicine. 2018;50:213–22.
Lu WD, Qin Y, Yang C, Li L, Fu ZX. Effect of curcumin on human colon cancer multidrug resistance <i>in vitro</i> and <i>in vivo</i>. Clinics. 2013;68(5):694–701.
Li L, Yu L, Cao X, Zhang C, Liu Q, Chen J. Comparative analysis of proteomic of curcumin reversing multidrug resistance in HCT-8/VCR Cells. J Oncol. 2022;2022(1):3605436.
James MI, Iwuji C, Irving G, Karmokar A, Higgins JA, Griffin-Teal N, et al. Curcumin inhibits cancer stem cell phenotypes in ex vivo models of colorectal liver metastases, and is clinically safe and tolerable in combination with FOLFOX chemotherapy. Cancer Lett. 2015;364(2):135–41.
Howells LM, Iwuji CO, Irving GR, Barber S, Walter H, Sidat Z, et al. Curcumin combined with FOLFOX chemotherapy is safe and tolerable in patients with metastatic colorectal cancer in a randomized phase IIa trial. J Nutr. 2019;149(7):1133–9.
Gunther JR, Chadha AS, Guha S, Raju GS, Maru DM, Munsell MF, et al. A phase II randomized double blinded trial evaluating the efficacy of curcumin with pre-operative chemoradiation for rectal cancer. J Gastrointest Oncol. 2022;13(6):2938.
Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003;3(5):330–8.
Deveci HA, Nazıroğlu M, Nur G. 5-Fluorouracil-induced mitochondrial oxidative cytotoxicity and apoptosis are increased in MCF-7 human breast cancer cells by TRPV1 channel activation but not Hypericum perforatum treatment. Mol Cell Biochem. 2018;439:189–98.
Shirasaka T, Taguchi T. Timeline from discovery of 5-FU to development of an oral anticancer agent S-1 and its drug concept. Gan to Kagaku ryoho Cancer & Chemotherapy. 2006;33:4–18.
Sun Y, Tang XM, Half E, Kuo MT, Sinicrope FA. Cyclooxygenase-2 overexpression reduces apoptotic susceptibility by inhibiting the cytochrome c-dependent apoptotic pathway in human colon cancer cells. Cancer Res. 2002;62(21):6323–8.
Ramasamy TS, Ayob AZ, Myint HHL, Thiagarajah S, Amini F. Targeting colorectal cancer stem cells using curcumin and curcumin analogues: insights into the mechanism of the therapeutic efficacy. Cancer Cell Int. 2015;15(1):1–15.
Link A, Balaguer F, Shen Y, Lozano JJ, Leung H-CE, Boland CR, et al. Curcumin modulates DNA methylation in colorectal cancer cells. PLoS ONE. 2013.
Mudduluru G, George-William JN, Muppala S, Asangani IA, Kumarswamy R, Nelson LD, et al. Curcumin regulates miR-21 expression and inhibits invasion and metastasis in colorectal cancer. Biosci Rep. 2011;31(3):185–97.
Sottile ML, Cuello-Carrión FD, Gómez LC, Semino S, Ibarra J, García MB, et al. DNA damage repair proteins, HSP27, and phosphorylated-hsp90α as predictive/prognostic biomarkers of platinum-based cancer chemotherapy: an exploratory study. Appl Immunohistochem Mol Morphol. 2022;30(6):425–34.
Makovec T. Cisplatin and beyond: molecular mechanisms of action and drug resistance development in cancer chemotherapy. Radiol Oncol. 2019;53(2):148–58.
Jardim D, Rodrigues C, Novis Y, Rocha V, Hoff P. Oxaliplatin-related thrombocytopenia. Ann Oncol. 2012;23(8):1937–42.
Raymond E, Faivre S, Chaney S, Woynarowski J, Cvitkovic E. Cellular and molecular pharmacology of oxaliplatin. Mol Cancer Ther. 2002;1(3):227–35.
Irving GR, Iwuji CO, Morgan B, Berry DP, Steward WP, Thomas A, et al. Combining curcumin (C3-complex, Sabinsa) with standard care FOLFOX chemotherapy in patients with inoperable colorectal cancer (CUFOX): study protocol for a randomised control trial. Trials. 2015;16:1–10.
Zhang X, Guan Z, Wang X, Sun D, Wang D, Li Y, et al. Curcumin alleviates oxaliplatin-induced peripheral neuropathic pain through inhibiting oxidative stress-mediated activation of NF-κB and mitigating inflammation. Biol Pharm Bull. 2020;43(2):348–55.
Wilson C, Purcell C, Seaton A, Oladipo O, Maxwell PJ, O’Sullivan JM, et al. Chemotherapy-induced CXC-chemokine/CXC-chemokine receptor signaling in metastatic prostate cancer cells confers resistance to oxaliplatin through potentiation of nuclear factor-κB transcription and evasion of apoptosis. J Pharmacol Exp Ther. 2008;327(3):746–59.
Marteijn JA, Lans H, Vermeulen W, Hoeijmakers JH. Understanding nucleotide excision repair and its roles in cancer and ageing. Nat Rev Mol Cell Biol. 2014;15(7):465–81.
Oun R, Moussa YE, Wheate NJ. The side effects of platinum-based chemotherapy drugs: a review for chemists. Dalton Trans. 2018;47(19):6645–53.
Rezaee R, Momtazi AA, Monemi A, Sahebkar A. Curcumin: a potentially powerful tool to reverse cisplatin-induced toxicity. Pharmacol Res. 2017;117:218–27.
Nakano S, Murakami K, Meguro M, Soejima H, Higashimoto K, Urano T, et al. Expression profile of LIT1/KCNQ1OT1 and epigenetic status at the KvDMR1 in colorectal cancers. Cancer Sci. 2006;97(11):1147–54.
Qi X, Yu X-J, Wang X-M, Song T-N, Zhang J, Guo X-Z, et al. RETRACTED: knockdown of kcnq1ot1 suppresses cell invasion and sensitizes osteosarcoma cells to CDDP by upregulating DNMT1-mediated Kcnq1 expression. Mol Ther Nucleic Acids. 2019;17:804–18.
Vigneri PG, Tirrò E, Pennisi MS, Massimino M, Stella S, Romano C, et al. The insulin/IGF system in colorectal cancer development and resistance to therapy. Front oncol. 2015;5:230.
Cheraghpour M, Askari M, Tierling S, Shojaee S, Sadeghi A, Ketabi Moghadam P, et al. A systematic review and meta-analysis for the association of the insulin-like growth factor1 pathway genetic polymorphisms with colorectal cancer susceptibility. Front oncol. 2023;13:1168942.
Lombardo LJ, Lee FY, Chen P, Norris D, Barrish JC, Behnia K, et al. Discovery of N-(2-chloro-6-methyl-phenyl)-2-(6-(4-(2-hydroxyethyl)-piperazin-1-yl)-2-methylpyrimidin-4-ylamino) thiazole-5-carboxamide (BMS-354825), a dual Src/Abl kinase inhibitor with potent antitumor activity in preclinical assays. J Med Chem. 2004;47(27):6658–61.
Ouyang M, Luo Z, Zhang W, Zhu D, Lu Y, Wu J, et al. Protective effect of curcumin against irinotecan-induced intestinal mucosal injury via attenuation of NF-κB activation, oxidative stress and endoplasmic reticulum stress. Int J Oncol. 2019;54(4):1376–86.
Sheehan D, Meade G, Foley VM, Dowd CA. Structure, function and evolution of glutathione transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem J. 2001;360(1):1–16.
Sadzuka Y, Nagamine M, Toyooka T, Ibuki Y, Sonobe T. Beneficial effects of curcumin on antitumor activity and adverse reactions of doxorubicin. Int J Pharm. 2012;432(1–2):42–9.
Zhang R, Zhuang X, Zong L, Liu S, Liu Z, Song F. Investigations on the cell metabolomics basis of multidrug resistance from tumor cells by ultra-performance liquid chromatography–mass spectrometry Analytical and bioanalytical chemistry. Anal Bioanal Chem. 2016.
Cort A, Ozben T, Saso L, De Luca C, Korkina L. Redox control of multidrug resistance and its possible modulation by antioxidants. Oxid Med Cell Longev. 2016.
Matés JM, Di Paola FJ, Campos-Sandoval JA, Mazurek S, Márquez J. Therapeutic targeting of glutaminolysis as an essential strategy to combat cancer. Semin Cell Biol. 2020;98:34–43.
Sharma RA, McLelland HR, Hill KA, Ireson CR, Euden SA, Manson MM, et al. Pharmacodynamic and pharmacokinetic study of oral Curcuma extract in patients with colorectal cancer. Clin Cancer Res. 2001;7(7):1894–900.
He Z-Y, Shi C-B, Wen H, Li F-L, Wang B-L, Wang J. Upregulation of p53 expression in patients with colorectal cancer by administration of curcumin. Cancer Investig. 2011;29(3):208–13.
Jeon Y, Sym SJ, Yoo BK, Baek JH. Long-term survival, tolerability, and safety of first-line bevacizumab and folfiri in combination with ginsenoside-modified nanostructured lipid carrier containing curcumin in patients with unresectable metastatic colorectal cancer. Integr Cancer Ther. 2022;21:15347354221105498.
Guo L, Wang C, Qiu X, Pu X, Chang P. Colorectal cancer immune infiltrates: significance in patient prognosis and immunotherapeutic efficacy. Front immunol. 2020;11:1052.
Gregory AD, McGarry HA. Tumor-associated neutrophils: new targets for cancer therapy. Cancer Res. 2011;71(7):2411–6.
Tanaka A, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Cell Res. 2017;27(1):109–18.
Dent P, Booth L, Roberts JL, Poklepovic A, Hancock JF. (Curcumin+ sildenafil) enhances the efficacy of 5FU and anti-PD1 therapies in vivo. J Cell Physiol. 2020;235(10):6862–74.
Xiao Z, Su Z, Han S, Huang J, Lin L, Shuai X. Dual pH-sensitive nanodrug blocks PD-1 immune checkpoint and uses T cells to deliver NF-κB inhibitor for antitumor immunotherapy. Sci Adv. 2020.
Weng W, Goel A. Curcumin and colorectal cancer: an update and current perspective on this natural medicine. Semin Cancer Biol. 2022.
Xu B, Yu L, Zhao L-Z. Curcumin up regulates T helper 1 cells in patients with colon cancer. Am J Transl Res. 2017;9(4):1866.
Layos L, Martínez-Balibrea E, Ruiz de Porras V. Curcumin: a novel way to improve quality of life for colorectal cancer patients? Int J Mol Sci. 2022.
Lu Y, Wu S, Xiang B, Li L, Lin Y. Curcumin attenuates oxaliplatin-induced liver injury and oxidative stress by activating the Nrf2 pathway. Drug Des Devel Ther. 2020.
Belcaro G, Hosoi M, Pellegrini L, Appendino G, Ippolito E, Ricci A, et al. A controlled study of a lecithinized delivery system of curcumin (Meriva®) to alleviate the adverse effects of cancer treatment. Phytother Res. 2014;28(3):444–50.
Mundekkad D, Cho WC. Applications of curcumin and its nanoforms in the treatment of cancer. Pharmaceutics. 2023;15(9):2223.
Wu X, Yang Y. Research progress on drug delivery systems for curcumin in the treatment of gastrointestinal tumors. World J Gastrointest Oncol. 2023;15(8):1342.
Zhang Y, Li Z, Huang Y, Xu Y, Zou B. Nanotechnology and curcumin: a novel and promising approach in digestive cancer therapy. Nanomedicine. 2023;18(27):2081–99.
Acknowledgements
None
Funding
None.
Author information
Authors and Affiliations
Contributions
Shadnoush M, Momenan M, Seidel V, Tierling S, Fatemi N, Nazemalhosseini-Mojarad E, Tayefeh Norooz M and Cheraghpour M contributed to the writing and final approval of the manuscript.
Corresponding author
Ethics declarations
Conflict of interests
The authors declare that they have no conflicts of interest.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Shadnoush, M., Momenan, M., Seidel, V. et al. A comprehensive update on the potential of curcumin to enhance chemosensitivity in colorectal cancer. Pharmacol. Rep (2024). https://doi.org/10.1007/s43440-024-00652-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s43440-024-00652-y