
Abstract
Cancer cells require higher levels of ATP for their sustained growth, proliferation, and chemoresistance. Mitochondrial matrix protein, C1qbp is upregulated in colon cancer cell lines. It protects the mitochondria from oxidative stress, by inhibiting the Membrane Permeability Transition (MPT) pore and providing uninterrupted synthesis of ATP. This intracellular interaction of C1qbp could be involved in chemoresistance development. Natural chemosensitizing agent, curcumin has been used in the treatment of multiple cancers. In this current study, we elucidate the role of C1qbp during curcumin induced chemosensitization to doxorubicin resistant colon cancer cells. The possible interaction between C1qbp and curcumin was determined using bioinformatics tools—AutoDock, SYBYL, and PyMol. Intracellular doxorubicin accumulation by fluorimetry and dead cell count was carried out to determine development of chemoresistance. Effect of curcumin treatment and cytotoxicity was measured by MTT and lactate dehydrogenase release. Morphological analysis by phase contrast microscopy and colony forming ability by colonogenic assay were also performed. In addition, Cox-2 could mediate P-glycoprotein upregulation via phosphorylation of c-Jun. Thus, the gene level expression of P-glycoprotein and Cox-2 was also investigated using PCR. Through molecular docking we identified possible interaction between curcumin and C1qbp. We observed development of chemoresistance upon 6th day treatment. Concentration dependent alleviation of chemoresistance development by curcumin was confirmed and was found to reduce gene level expression of P-glycoprotein and Cox-2. Hence, curcumin could interact directly with C1qbp protein and this interaction could contribute to the chemosensiting effect to doxorubicin in colon cancer cells.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
C1qbp/g-C1qR/p32 has a doughnut shaped quaternary structure with three monomers; each of which is composed of seven consecutive anti-parallel β-sheets with one N-terminal and two C-terminal α-helices [1]. This protein is predominantly expressed in the mitochondrial matrix (targeted by the amino terminal) [2], is also detected in cytoplasm, and in low levels on the cell surface [3]. The various roles of C1qbp on the cell surface are: pro-inflammatory action by induction of macrophage migration, mediating actions of pro-inflammatory agents like kininogen, prevent cell damage by eliminating C1q from inflammation site, act as entry point for viruses such as HIV, hepatitis C and binding to extracellular matrix protein hyaluronan [4]. Release of the protein into the extracellular matrix has been associated with cell stress, defective apoptosis or necrosis [3]. Enhanced expression of C1qbp/gC1q-R has been observed in a myriad of cancer cells. This increase in expression has been linked to a reversable switch from glycolysis to oxidative phosphorylation and tumorigenesis [5]. In tumors, C1qbp is mainly localized in nutrient deprived/ hypoxia regions and increased levels have been detected in tumor associated macrophage/myeloid subpopulation [6]. mRNA expression levels were higher in breast cancer tissues with increased protein levels in the cytoplasm and is considered as a possible biomarker [7]. Overexpression of C1qbp diminished reactive oxygen species (ROS) induced mitochondrial permeability transition pore (MPTP) activation while knockdown causes sensitization to pore opening [8]. The pore inhibition was observed as a consequence of interaction with cyclin D, maximal at 1:1 ratio and preventing cell death [9]. In addition, C1qbp knockdown resulted in doxorubicin (dox) mediated apoptosis of triple negative breast cancer cells MDA-MB-23 and increased expression was also observed in lung and colon cancer cells compared to normal cells [10].
Commonly used chemotherapeutic drug doxorubicin (dox), has multiple intracellular targets: inhibiting topoisomerase II enzyme [11], promotes apoptosis through the generation of ROS via the activation of NF-κβ [12], formation of dox-DNA adducts [13], induction of immune response [14], ceramide overproduction [15], interaction with iron regulatory proteins [16] and ROS mediated apoptosis through dysregulation of intracellular Ca2+ levels [17]. The rate of success of chemotherapy is higher for colon cancer [18], but the development of chemoresistance is a limitation for chemotherapy in colon cancer [19]. In tumor cell lines, chemoresistance is often associated with the overexpression of ATP–dependent drug efflux proteins belonging to the superfamily of ATP–binding cassette (ABC) transporters: the 170 kDa P-glycoprotein (P-gp, MDR1 gene) and the 190 kDa multidrug resistance-associated protein-1 (MRP-1 gene) [20]. When the function of the membrane permeability transition pore (MPTP) gets inhibited by C1qbp, the mitochondria is protected from damage via oxidative stress, leading to the production of ample amount of ATP. As P-gp is an ATP-dependent efflux pump, it pumps the chemotherapeutic drug out of the cells, making the cancer cell chemoresistant. Curcumin, an anti-inflammatory, anti-cancer compound from Curcuma longa, chemo-sensitizes colorectal cancer cells to 5-FU (5-Fluorouracil) through the inhibition of EMT (Epithelial to Mesenchymal Transition) [21]. Curcumin could enhance the chemotherapeutic effects of irinotecan in colon cancer cell lines by inducing ROS and endoplasmic reticulum stress [22]. Further, curcumin treatment to dox resistant breast cancer cell lines resulted in an increased intracellular levels of dox and reversal of chemoresistance by inhibiting ABCB4 [23]. On the other hand, curcumin induces an increase in rat liver mitochondrial membrane permeability leading to loss of membrane potential, swelling, and ATP synthesis inhibition. This effect was mediated through the opening of MPTP by thiol functional group oxidation, in the presence of low Ca2+ levels [24]. These data suggests that mitochondria might be a target by which curcumin induces apoptosis in tumor cells. In this study, we have evaluated the possible interaction of curcumin with C1qbp and its chemosensitization effect in dox induced chemoresistant coloadenocarcinoma cell line.
Materials and methods
Materials
HT-29 cells (RRID:CVCL_0320) were obtained from National Centre for Cell Science, Pune, India. Doxorubicin hydrochloride was purchased from MP Biomedicals, Japan. McCoy’s 5A Media, Fetal bovine serum (FBS), Antibiotic/Antimycotic Solution, dimethyl sulphoxide (DMSO), tryphan blue dye, agarose, bovine serum albumin (BSA), bromophenol blue, tris base, sodium dodecyl sulphate (SDS), acrylamide, bis-acrylamide, Ammonium persulphate (APS) and Sodium deoxycholate were purchased from Himedia. DL-dithiothreitol, phenylmethylsulphonylfluoride (PMSF), Taq DNA polymerase and TRIzol S reagent were purchased from Sisco Research Laboratory. Leupeptin hemisulphate and dNTP mix were purchased from Merck. Curcumin and diethyl pyrocarbonate (DEPC) were purchased from Sigma. Primescript 1st strand cDNA synthesis kit from Takara Bio. PCR primers for MDR1 (P-gp), COX-2, and β-actin from Xcelris Genomics.
Methodology
Bioinformatics study—to find interaction between C1qbp and curcumin
3-D structures of C1qbp and curcumin, its derivatives and metabolites, were retrieved from Protein Data Bank (PDB) and Zinc Database respectively. Since the structures from PDB, contain neither water molecules nor polar hydrogen bonds, they were added using PyMol and AutoDock (version 4.5) softwares, respectively. Since curcumin’s active site on C1qbp protein is unknown, SYBYL software was used for this purpose. On analysis, 11 binding pockets were deduced and curcumin’s interaction with each pocket was performed through AutoDock. After the binding site was obtained, the interaction of the derivatives and metabolites of curcumin to the protein was also further examined by AutoDock.
Cell culture and treatment
HT-29 cells were cultured in McCoy’s 5A media with 10% FBS and 1% antibiotic–antimycotic solution. The experiments were carried out with 60% confluent cells with passages between 34 to 49. HT-29 cells were treated with increasing concentrations of dox (0.23 μM, 0.46 μM, 0.69 μM, 0.92 μM, 1.15 μM, 1.38 μM and 1.61 μM) [25] each day for the 7-day treatment. HT-29 cells after 7th day were labelled as MDR-HT-29 cells. Curcumin was co-treated at 1 μM and 10 μM each day.
Morphology study
Cells were seeded in 6 well plates at a seeding density of 4 × 105 cells/well. The cells were subjected to dox treatment and dox- curcumin co-treatment. Images of the cells were taken on the 0th, 1st, 3rd and 7th days respectively using a phase contrast microscope (Q-Capture Pro 7 Software) at 10x and 40x magnifications.
Doxorubicin accumulation assay
Cells were seeded in 6 well plates with a seeding density of 20,000 cells/well. Dox accumulation assay was performed for the cells treated with dox, each day up to 7th day [26]. The medium was aspirated, after 1X PBS wash, cells were scrapped using lysis solution (1:1 ratio, Ethanol:0.3 N HCl) and sonicated for two 10 s bursts at 100 W, on ice. 200 μl of each lysate was analysed using a fluorescence microplate reader at excitation and emission wavelengths, 475 nm and 553 nm respectively. A calibration curve in the range of 0.2 to 7 nmol/ml was used to elucidate dox concentrations. 50 μl of lysate was utilized to determine the protein concentration by Bradford’s method [27]. Protein concentrations of the samples were determined using BSA standard and was used to normalize the dox concentration.
Dead cell count
Cells were seeded at 20,000 cells/well in 6-well plates. The medium was centrifuged and the pellet resuspended using 1X PBS, 10 µl of this suspension was mixed with equal volume of 0.4% tryphan blue and used for counting.
Lactate dehydrogenase (LDH) assay
1 mL of media after treatment was centrifuged at 1500 rpm for 3 min at 37 °C. 50 μl of supernatant was added to 2 mL of 56 mM Tris-5.6 mM EDTA-170 µM NADH buffer and incubated at room temperature for 15 min. The absorbance of the samples was taken at 340 nm, immediately after the addition of 200 μl of freshly prepared 14 mM sodium pyruvate [28]. Readings were obtained every 15 s for 3 min. The LDH activity was calculated for each day.
Cell proliferation assay
The cells were seeded at a density of 5000 cells/well in 96 well plate. After the respective treatment period, the media was aspirated, 100 μl of fresh media and 50 μl of MTT (5 mg/ml in 1x PBS) was added to each well and incubated at 37 °C for 4 h. 100 μl DMSO was added to each well to dissolve the formazan crystals and after few minutes the plates were read at 570 nm [29].
Total RNA isolation
Total RNA was isolated from the untreated and treated cells using TRIzol S reagent, according to the manufacturer’s recommendations. The medium was removed and 1 ml of TRIzol was added. Cells were lysed by pipetting several times and incubated for 5 min at room temperature. 0.2 ml of chloroform was added, agitated vigorously for 15 s and centrifuged at 11,000 rpm for 15 min at 4 °C. The aqueous phase was transferred to a fresh eppendrof, 0.5 ml of isopropanol was added and centrifuged at 11,000 rpm for 10 min at 4 °C. The pellet was washed with 1 ml of 75% ethanol and centrifuged at 8000 rpm for 5 min at 4 °C. The pellet obtained was air-dried and resuspended in 100 μl of RNase free water. The sample was subjected to purity analysis and integrity was detected using 1.2% agarose gel electrophoresis.
cDNA synthesis
cDNA was synthesized using Primescript 1st strand synthesis kit, as per the manufacturer’s instructions. A template mix was prepared per reaction (1 μl Random Hexamer (50 μM), 1 μl dNTP mixture (10 mM each), 1.6 μl Total RNA (2 μg) and 6.4 μl of RNase free water); incubated at 65 °C for 5 min and immediately cooled on ice. The RT (Reverse Transcription) mix (4 μl of 5× Primescript buffer, 0.5 μl of RNase inhibitor (40 U/μl), 1 μl of Primescript RTase (200U/μl) and 4.5 μl of RNase free water) was prepared. 10 μl of the RT mix was added to template mix and placed in thermal cycler programmed for incubation at 30 °C for 10 min, 42 °C for 60 min, 70 °C for 15 min and 4 °C (Hold).
Polymerase chain reaction
PCR was performed for MDR1, Cox-2 and β-actin expression. The primer sequences are.
β-actin Forward primer: 5′-TGGCACCCAGCACAATGAA-3′.
β-actin Reverse primer: 5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′.
MDR1 Forward primer: 5′-AGGCCAACATACATGCCTTCATC-3′.
MDR1 Reverse primer: 5′-GCTGACGTGGCTTCATCCAA-3′.
COX-2 Forward primer: 5′-CTGTAACCAAGATGGATGCAAAGA-3′.
COX-2 Reverse primer: 5′-GTCAGTGACAATGAGATGTGGAA-3′.
All reactions were carried out at a total volume of 50 μl. A master mix was prepared for each reaction (5 μl of 10X PCR buffer, 0.4 μl of dNTP mix (25 mM each), 2.5 μl each of the forward and reverse primers (10 μM each), 3 μl of 25 mM MgCl2, 2 μl of Taq polymerase (1 U/μl), 2 μl of cDNA and 32.6 μl of water). The tubes were mixed and spun briefly. The tubes were placed in the thermal cycler and the following PCR conditions were used for both the targets: 1 cycle of initial denaturation at 94 °C for 3 min, 40 cycles each of denaturation at 94 °C for 30 s, annealing at 60 °C for 30 s, extension at 72 °C for 1 min and 1 cycle of final extension at 72 °C for 10 min. Hold at 4 °C. The PCR products were analysed using 3% Agarose gel electrophoresis. The band intensities were quantified using Image J software. MDR1 expression was normalized against β-actin expression.
Colonogenic assay
The colony forming ability of the cells after treatment were performed by seeding 2 × 105 cells/well in a 6-well plate. After 7 days treatment, cells were washed with 1× PBS and fixed using methanol: acetic acid (3:1 ratio) solution for 5 min. The cells were then stained in 0.5% crystal violet (in methanol) for 15 min. Excess stain was washed off with water till the colonies were visible [30].
Statistical analysis
All the experiments were performed as independent triplicates and were expressed as Mean ± SD. The data obtained were analyzed using SPSS v.24 software by one-way ANOVA with Bonferroni post hoc analysis and p-value of less than 0.05 was considered significant.
Results
Doxorubicin accumulation assay
The intracellular dox accumulation was determined for each day of treatment (Fig. 1a). The intracellular dox accumulation was found to increase up to 5th day of treatment, followed by a significant decrease on 6th and 7th days of treatment. This indicates that the cells might have acquired multi drug resistance phenotype upon 6th day of dox treatment. The concentrations of dox was elucidated using the dox calibration curve.

a Intracellular doxorubicin accumulation (nmol/mg protein) for 7 days of dox treatment. *Significant difference from 5th day, p < 0.05. b Number of dead cells after each day of treatment for 7 days. *Significance with respect to day 1, #significance with respect to day 2. $Significance with respect to day 3, &significance with respect to day 5, p < 0.05. b Dead cell count over the 7 days of dox treatment
Dead cell count
The number of dead cells in the medium aspirated was counted after each day of dox treatment. The number of dead cells were found to increase up to the 5th day of treatment followed by a decrease after the 6th and 7th days of treatment (Fig. 1b). This correlates with the decrease in the intracellular dox levels after 6th and 7th days, indicating that the cells might have acquired chemoresistance.
Interaction between C1qbp and curcumin
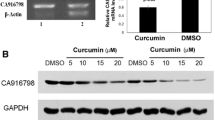
Higher the negative value of the binding energy (generally − 6 to − 8 kcal/mol), more is the possibility and strength of the interaction between the protein and the ligand. The third binding pocket shows the highest energy (Supplementary File A), which could be the possible active site for curcumin to bind to the protein. Binding Energies of the derivatives and metabolites of curcumin were obtained through AutoDock (Demethoxycurcumin = − 2.38 kcal/mol; Bisdemethoxycurcumin = − 1.96 kcal/mol; Tetrahydrocurcumin = − 2.29 kcal/mol; Hexahydrocurcumin = − 2.38 kcal/mol; Octahydrocurcumin = − 1.97 kcal/mol). These results show that only curcumin interacts with C1qbp (Fig. 2).

a PDB ID:1P32–C1qbp, an acidic mitochondrial matrix protein, consisting of three chains—A (Yellow), B (Pink) and C (Green); b Zinc Database ID: CUR—Curcumin. c AutoDock Result—Interaction of Curcumin with the third binding pocket in C1qbp. Curcumin (Yellow) binds to C1qbp (Grey) by forming two Hydrogen Bonds (Green Dots, white arrows) with the amino acid residues, Asparagine (ASN I67—B Chain, top white circle) and Aspartic Acid (ASP 221—A Chain, lower white circle)
Morphology analysis
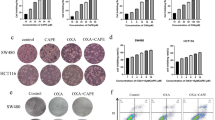
The cells begin to lose their morphology from the third day of dox, 1 μM curcumin + dox and 10 μM curcumin + dox treatment. More cell death was observed in 10 μM curcumin + dox treatment when compared to 1 μM curcumin + dox. Significant morphological changes were not observed in cells treated with 10 μM curcumin alone (Fig. 3).

Phase contrast microscopy images of the treated HT-29 cells in the magnifications 10x and 40x
Lactate dehydrogenase assay
The release of lactate dehydrogenase enzyme during non-apoptotic cell death was determined by the addition of substrate, pyruvate and β-NADH cofactor. From the 3rd day, dox treatment had significantly increased LDH activity compared to the control (Fig. 4a). In addition, curcumin co-treatment at both the concentrations, resulted in a decreased level of enzyme activity in the medium compared to dox treatment. Thus, indicating that curcumin plays a role in dox induced cell death.

a Lactate dehydrogenase release from the cells over the 7 days treatment period. *Significant difference between control and doxorubicin, #significant difference between dox and curcumin co-treatments: dox + 1 μM curcumin and dox + 10 μM curcumin, $significant difference between control and 10 μM curcumin, %significant difference between dox + 1 μM curcumin and dox + 10 μM curcumin, p < 0.05. b Cell proliferation assay of HT-29 cells treated with dox in the presence of 1 μM and 10 μM curcumin for 1, 3, and 7 days. *Significant when compared to control, #significant when compared to 10 µM curcumin with doxorubicin treatment, $significant when compared to 10 µM curcumin treatment, p < 0.05
Cell proliferation assay
The cell proliferation rate upon dox treatment showed a significant reduction by 10%, 38%, and 41% after day 1, day 3, and day 7 treatment respectively. 1 µM curcumin co-treatment with dox also showed a similar effect with no comparable difference with dox treatment alone. However, 10 µM curcumin co-treatment resulted in cell proliferation rate lower than that of dox and 10 µM curcumin treatment controls. The 10 µM curcumin co-treatment resulted in cell proliferation reduction by 20%, 50%, and 85% after day 1, day 3, and day 7 treatment respectively. While, 10 µM curcumin treatment alone resulted in reduction by 13%, 38%, and 70% after day 1, day 3, and day 7 treatment respectively (Fig. 4b).
Polymerase chain reaction
The band intensity of MDR1 and Cox-2 expression was normalized with that of β-Actin and the relative mRNA expression levels were calculated. The MDR1 and Cox-2 expression was found to have significantly increased after the 7th day of dox treatment compared to that of the control. This confirms that the cells have acquired resistance phenotype through MDR1 overexpression, following 7th day of dox treatment, consistent with the dox accumulation. The MDR1 expression was found to have significantly reduced in the 1 μM and 10 μM curcumin co-treated cells, in a concentration dependent manner (Fig. 5a, b). In addition, Cox-2 expression levels were also found to be higher in MDR-HT-29 cells while a reduction was observed in curcumin co-treatment in concentration dependent manner (Fig. 5c).

a MDR1, Cox-2, and β-Actin PCR products. Lane 1: control (untreated) HT-29 cells, Lane 2: MDR HT29 cells (7 days dox treated), Lane 3: 1 μM curcumin co-treated cell, Lane 4: 10 μM curcumin co-treated cells. b Relative mRNA expression of MDR1. *Significant difference when compared to control at p < 0.05; # significant difference when compared to MDR-HT29 at p < 0.05). c Relative mRNA expression of Cox-2. *Significant difference when compared to control at p < 0.05; #significant difference when compared to MDR-HT29 at p < 0.05). d Colony formation after 7 days treatment, visualized by crystal violet staining
Colonogenic assay
Colonogenic assay was performed to determine the colony forming ability after treatment with curcumin and dox combinations. Compared to the control, dox treatment had a significantly visible increase in the number of colonies. This might be attributed to the development of chemoresistance of HT-29 cells (Fig. 5d). However, when curcumin was co-treated with dox, this effect was reversed. The percentage of cells were decreased with increasing concentrations of curcumin. Thus, curcumin has a dose dependent effect in preventing dox chemoresistance development. No observable increase in the cell colonies were detected in 10 µM curcumin treatment alone compared to control.
Discussion
197 intracellular protein targets for C1qbp has been reported of which, interaction with protein kinase C Z type (PRKCZ) resulted in its activation, translocation, cofilin mediated actin polymerization, mobilization and hence cell chemotaxis in the presence of EGF. Furthermore, the C1qbp was observed to be overexpressed in breast cancer cells and higher expression was observed to be linked to metastasis and TNM stage of the cancer [31]. Cleaved form of C1qbp (14 kDa), has been reported to be elevated in colon cancer compared to normal cells [32]. Through CoIP-MS, C1qbp was found to interact with the mitochondrial protein DLAT, core component of pyruvate dehydrogenase enzyme, and regulate oxidative phosphorylation. The interaction resulted in higher activity of the enzyme by 30% and depletion of which resulted in 45% reduced activity in renal cancer cell lines [33]. In pancreatic cancer cells, C1qbp was found to be translocated to the lipid rafts through IGF-1 (Insulin like growth factor 1) and upon interaction with CD44v6 resulted in phosphorylation of IGF-1R, activation of PI3K, and MAPK pathways. Thus, being responsible for hepatic metastasis of pancreatic cancer cells [34]. In lung carcinoma cell line A549, growth factor induced lamellipodia formation and metastasis was significantly reduced when the receptor for c1q (gC1q-R) in lipid rafts was depleted [35]. In renal carcinoma cell lines (786-0, ACHN, and HEK-293T), knockdown of C1qbp increased the expression of adhesion molecules such as L1CAM and is said to induce lung and liver metastasis by GSK3/β-Catenin/L1CAM signaling pathway [36]. In contrast, in renal cell carcinoma, C1qbp has been found to interact with YBX1, suppressing cell migration and invasion [37]. Among the metabolites of curcumin, we were able to identify through AutoDock that curcumin could interact directly with C1qbp through hydrogen bonds with asparagine 167-B chain and aspartic acid 221-A chain. The strength and possibility of interaction depends on higher negative binding energy. Here we show that the 3rd pocket interaction has a binding energy of − 6.02 kcal/mol. Hence, we conclude that curcumin alone and not its metabolites can interact with C1qbp through autodocking. Furthermore, membrane type-1 matrix metalloproteases involved in locomotion, matrix degradation, and activation of soluble secretary matrix metalloproteases, binds directly with p32 through the cytoplasmic tail and is involved in its presentation on the tumor cell surface [38]. Taken together, we infer that differential expression of C1qbp might be associated with tumorigenesis and metastasis.
Chemosensitizing effect of curcumin was found to be through direct interaction with P-gp, modulating its functionality and expression in primary rat hepatocytes [39]. Further, curcumin and its analogues could reverse P-gp by inhibiting either its function, expression or both in dox resistant K562 leukemic cells [40] and paclitaxel resistant human breast cancer cells [41]. Similarly, curcumin treatment was found to target P-gp and S100A8 calcium binding protein, reversing dox resistance by inhibiting the expression and functionality of P-gp, increasing dox accumulation, downregulation S100A8 expression, calcium ion imbalance, increasing endoplasmic reticulum (ER) stress and apoptosis [42]. Previously, it has been reported that cardiotoxic side effect of dox through excessive opening of MPTP was alleviated when curcumin was co-treated to the cells, by downregulating mitochondrial phosphate carrier, and preventing cellular apoptosis [43]. At two to three-fold lower concentration, dox-curcumin liposomal formulation reduced the anti-proliferative effect of C26 murine colon adenocarcinoma cell line than free dox and curcumin. The synergistic cytotoxic effect of the encapsulated drugs was curcumin concentration dependent [44]. PEGylated micelles consisting of d-tocopheryl PEG1000 succinate (TPGS) have been synthesized for the delivery of dox and curcumin to dox resistant A549 lung cancer cells. TPGS a known inhibitor of P-gp; curcumin a sensitizer and inhibitor of MDR proteins, synergistically reversed dox resistance [45]. To determine the development of chemoresistance we treated the cells with increasing concentrations of dox over a 7-day period. We observed that, after 6th day treatment the intracellular dox accumulation level significantly reduced by approximately 5.5 nmol/mg protein. Thus, indicating that the cells might have developed a mechanism to pump the chemotherapeutic drug out. Further, this conclusion was supported by the decrease in the dead cell count after day 6 treatment. To determine curcumin’s role in inhibiting resistance development, we co-treated dox with curcumin at two different concentrations. We found that when 10 µM curcumin was treated to the cells along with increasing concentrations of dox, higher cell death and reduced proliferation rate was observed. In contrast, with dox treatment alone, the cells had acquired chemoresistance by day 7. Further, we observed that with co-treatment, LDH release into the medium was significantly reduced compared to that of dox alone. This might be an indication that curcumin induces apoptotic cell death in contrast to dox. We then conducted PCR to determine the altered gene level expression of P-glycoprotein after 7 days treatment. We observed that MDR1 expression was increased in dox treated group, while curcumin co-treatment significantly reduced the expression in a concentration dependent manner. Thus, the upregulation of P-gp might be responsible for the resistance development and reduced dox accumulation observed from 6th day of treatment. Curcumin could sensitize the cells to dox by downregulating the expression of P-gp. It has been reported that increased expression of Cox-2 results in increase in P-gp in rat glomerular mesangial cells [46]. Colorectal polyp formation was significantly reduced when celecoxib, an inhibitor of cyclooxygenase-2 was treated at 400 mg twice daily to patients with familial adenomatous polyposis [47]. In colon cancer cell HCT8 and vincristine resistant HCT8/v, Cox-2 contributes to multidrug resistance through P-gp upregulation by phosphorylation of c-Jun [48]. The prostaglandin production by Cox-2 may induce the protein kinase c (PKC), expression of c-Jun and in turn increase the expression of MDR1 [49]. Similarly, we have also found increased expression of cyclooxygenase-2 gene levels upon dox treatment and a reduction in the expression with curcumin co-treatment. Thus, Cox-2 pathway might be involved in MDR1 gene expression and development of multidrug resistance. Furthermore, the colony forming ability of dox resistant cells were diminished when curcumin co-treatment was performed. Thus curcumin treatment could sensitize colon cancer cell line HT-29 to dox in a concentration dependent manner.
Conclusion
MDR1 is most commonly implicated in dox chemoresistance and is said to be regulated by various growth factors, but the mechanism is not clearly understood. The mitochondrial matrix protein C1qbp, has been implicated in regulation of oxidative phosphorylation and uninterrupted ATP synthesis when overexpressed in cancer cells. We are the first to report a direct interaction between curcumin and C1qbp through hydrogen bonds at asparagine and aspartic acid residues, identified using bioinformatics tools. We have confirmed that when curcumin was co-treated with dox, the development of chemoresistance was significantly reduced and it could be through Cox-2 pathway. Curcumin’s interaction with C1qbp might be a possible mechanism by which development of dox-induced chemoresistance can be regulated. Further studies on confirming this interaction through in vitro and in vivo models and their regulation could serve as a novel molecular target for overcoming development of dox chemoresistance in colon cancer cell lines.
Data availability
Yes, can be furnished upon requirement.
References
Jiang J, Zhang Y, Krainer AR, Xu RM (1999) Crystal structure of human p32, a doughnut-shaped acidic mitochondrial matrix protein. Proc Natl Acad Sci USA 96(7):3572–3577. https://doi.org/10.1073/pnas.96.7.3572
Dedio J, Jahnen-Dechent W, Bachmann M, Muller-Esterl W (1998) The multiligand-binding protein gC1qR, putative C1q receptor, is a mitochondrial protein. J Immunol 160(7):3534–3542
Eggleton P, Tenner AJ, Reid KB (2000) C1q receptors. Clin Exp Immunol 120(3):406–412
Barna J, Dimen D, Puska G, Kovacs D, Csikos V, Olah S, Udvari EB, Pal G, Dobolyi A (2019) Complement component 1q subcomponent binding protein in the brain of the rat. Sci Rep 9(1):4597. https://doi.org/10.1038/s41598-019-40788-z
Fogal V, Richardson AD, Karmali PP, Scheffler IE, Smith JW, Ruoslahti E (2010) Mitochondrial p32 protein is a critical regulator of tumor metabolism via maintenance of oxidative phosphorylation. Mol Cell Biol 30(6):1303–1318. https://doi.org/10.1128/MCB.01101-09
Fogal V, Zhang L, Krajewski S, Ruoslahti E (2008) Mitochondrial/cell-surface protein p32/gC1qR as a molecular target in tumor cells and tumor stroma. Cancer Res 68(17):7210–7218. https://doi.org/10.1158/0008-5472.CAN-07-6752
Scully OJ, Yu Y, Salim A, Thike AA, Yip GW, Baeg GH, Tan PH, Matsumoto K, Bay BH (2015) Complement component 1, q subcomponent binding protein is a marker for proliferation in breast cancer. Exp Biol Med (Maywood) 240(7):846–853. https://doi.org/10.1177/1535370214565075
Elrod JW, Molkentin JD (2013) Physiologic functions of cyclophilin D and the mitochondrial permeability transition pore. Circ J 77(5):1111–1122
McGee AM, Baines CP (2011) Complement 1q-binding protein inhibits the mitochondrial permeability transition pore and protects against oxidative stress-induced death. Biochem J 433(1):119–125. https://doi.org/10.1042/BJ20101431
McGee AM, Douglas DL, Liang Y, Hyder SM, Baines CP (2011) The mitochondrial protein C1qbp promotes cell proliferation, migration and resistance to cell death. Cell Cycle 10(23):4119–4127. https://doi.org/10.4161/cc.10.23.18287
Tewey KM, Rowe TC, Yang L, Halligan BD, Liu LF (1984) Adriamycin-induced DNA damage mediated by mammalian DNA topoisomerase II. Science 226(4673):466–468
Wang S, Kotamraju S, Konorev E, Kalivendi S, Joseph J, Kalyanaraman B (2002) Activation of nuclear factor-kappaB during doxorubicin-induced apoptosis in endothelial cells and myocytes is pro-apoptotic: the role of hydrogen peroxide. Biochem J 367(Pt 3):729–740. https://doi.org/10.1042/BJ20020752
Cutts SM, Nudelman A, Rephaeli A, Phillips DR (2005) The power and potential of doxorubicin-DNA adducts. IUBMB Life 57(2):73–81. https://doi.org/10.1080/15216540500079093
Bracci L, Schiavoni G, Sistigu A, Belardelli F (2014) Immune-based mechanisms of cytotoxic chemotherapy: implications for the design of novel and rationale-based combined treatments against cancer. Cell Death Differ 21(1):15–25. https://doi.org/10.1038/cdd.2013.67
Yang F, Teves SS, Kemp CJ (1845) Henikoff S (2014) Doxorubicin, DNA torsion, and chromatin dynamics. Biochim Biophys Acta 1:84–89. https://doi.org/10.1016/j.bbcan.2013.12.002
Gammella E, Maccarinelli F, Buratti P, Recalcati S, Cairo G (2014) The role of iron in anthracycline cardiotoxicity. Front Pharmacol 5:25. https://doi.org/10.3389/fphar.2014.00025
Octavia Y, Tocchetti CG, Gabrielson KL, Janssens S, Crijns HJ, Moens AL (2012) Doxorubicin-induced cardiomyopathy: from molecular mechanisms to therapeutic strategies. J Mol Cell Cardiol 52(6):1213–1225. https://doi.org/10.1016/j.yjmcc.2012.03.006
Weeks JC, Catalano PJ, Cronin A, Finkelman MD, Mack JW, Keating NL, Schrag D (2012) Patients' expectations about effects of chemotherapy for advanced cancer. N Engl J Med 367(17):1616–1625. https://doi.org/10.1056/NEJMoa1204410
Bose D, Zimmerman LJ, Pierobon M, Petricoin E, Tozzi F, Parikh A, Fan F, Dallas N, Xia L, Gaur P, Samuel S, Liebler DC, Ellis LM (2011) Chemoresistant colorectal cancer cells and cancer stem cells mediate growth and survival of bystander cells. Br J Cancer 105(11):1759–1767. https://doi.org/10.1038/bjc.2011.449
Tang XQ, Bi H, Feng JQ, Cao JG (2005) Effect of curcumin on multidrug resistance in resistant human gastric carcinoma cell line SGC7901/VCR. Acta Pharmacol Sin 26(8):1009–1016. https://doi.org/10.1111/j.1745-7254.2005.00149.x
Toden S, Okugawa Y, Jascur T, Wodarz D, Komarova NL, Buhrmann C, Shakibaei M, Boland CR, Goel A (2015) Curcumin mediates chemosensitization to 5-fluorouracil through miRNA-induced suppression of epithelial-to-mesenchymal transition in chemoresistant colorectal cancer. Carcinogenesis 36(3):355–367. https://doi.org/10.1093/carcin/bgv006
Huang YF, Zhu DJ, Chen XW, Chen QK, Luo ZT, Liu CC, Wang GX, Zhang WJ, Liao NZ (2017) Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget 8(25):40264–40275. https://doi.org/10.18632/oncotarget.16828
Wen C, Fu L, Huang J, Dai Y, Wang B, Xu G, Wu L, Zhou H (2019) Curcumin reverses doxorubicin resistance via inhibition the efflux function of ABCB4 in doxorubicinresistant breast cancer cells. Mol Med Rep 19(6):5162–5168. https://doi.org/10.3892/mmr.2019.10180
Morin D, Barthelemy S, Zini R, Labidalle S, Tillement JP (2001) Curcumin induces the mitochondrial permeability transition pore mediated by membrane protein thiol oxidation. FEBS Lett 495(1–2):131–136
Liu Z, Duan ZJ, Chang JY, Zhang ZF, Chu R, Li YL, Dai KH, Mo GQ, Chang QY (2014) Sinomenine sensitizes multidrug-resistant colon cancer cells (Caco-2) to doxorubicin by downregulation of MDR-1 expression. PLoS ONE 9(6):e98560. https://doi.org/10.1371/journal.pone.0098560
Riganti C, Miraglia E, Viarisio D, Costamagna C, Pescarmona G, Ghigo D, Bosia A (2005) Nitric oxide reverts the resistance to doxorubicin in human colon cancer cells by inhibiting the drug efflux. Cancer Res 65(2):516–525
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Sivalingam N, Basivireddy J, Pulimood AB, Balasubramanian KA, Jacob M (2009) Activation of phospholipase A2 is involved in indomethacin-induced damage in Caco-2 cells. Toxicol In Vitro 23(5):887–896. https://doi.org/10.1016/j.tiv.2009.05.008
Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65(1–2):55–63. https://doi.org/10.1016/0022-1759(83)90303-4
Franken NA, Rodermond HM, Stap J, Haveman J, van Bree C (2006) Clonogenic assay of cells in vitro. Nat Protoc 1(5):2315–2319. https://doi.org/10.1038/nprot.2006.339
Zhang X, Zhang F, Guo L, Wang Y, Zhang P, Wang R, Zhang N, Chen R (2013) Interactome analysis reveals that C1QBP (complement component 1, q subcomponent binding protein) is associated with cancer cell chemotaxis and metastasis. Mol Cell Proteomics 12(11):3199–3209. https://doi.org/10.1074/mcp.M113.029413
Kim K, Kim MJ, Kim KH, Ahn SA, Kim JH, Cho JY, Yeo SG (2017) C1QBP is upregulated in colon cancer and binds to apolipoprotein A-I. Exp Ther Med 13(5):2493–2500. https://doi.org/10.3892/etm.2017.4249
Chen R, Xiao M, Gao H, Chen Y, Li Y, Liu Y, Zhang N (2016) Identification of a novel mitochondrial interacting protein of C1QBP using subcellular fractionation coupled with CoIP-MS. Anal Bioanal Chem 408(6):1557–1564. https://doi.org/10.1007/s00216-015-9228-7
Shi H, Fang W, Liu M, Fu D (2017) Complement component 1, q subcomponent binding protein (C1QBP) in lipid rafts mediates hepatic metastasis of pancreatic cancer by regulating IGF-1/IGF-1R signaling. Int J Cancer 141(7):1389–1401. https://doi.org/10.1002/ijc.30831
Kim KB, Yi JS, Nguyen N, Lee JH, Kwon YC, Ahn BY, Cho H, Kim YK, Yoo HJ, Lee JS, Ko YG (2011) Cell-surface receptor for complement component C1q (gC1qR) is a key regulator for lamellipodia formation and cancer metastasis. J Biol Chem 286(26):23093–23101. https://doi.org/10.1074/jbc.M111.233304
Wang Y, Fu D, Su J, Chen Y, Qi C, Sun Y, Niu Y, Zhang N, Yue D (2017) C1QBP suppresses cell adhesion and metastasis of renal carcinoma cells. Sci Rep 7(1):999. https://doi.org/10.1038/s41598-017-01084-w
Yue D, Wang Y, Sun Y, Niu Y, Chang C (2017) C1QBP regulates YBX1 to suppress the androgen receptor (AR)-enhanced RCC cell invasion. Neoplasia 19(2):135–144. https://doi.org/10.1016/j.neo.2016.12.003
Rozanov DV, Ghebrehiwet B, Ratnikov B, Monosov EZ, Deryugina EI, Strongin AY (2002) The cytoplasmic tail peptide sequence of membrane type-1 matrix metalloproteinase (MT1-MMP) directly binds to gC1qR, a compartment-specific chaperone-like regulatory protein. FEBS Lett 527(1–3):51–57
Romiti N, Tongiani R, Cervelli F, Chieli E (1998) Effects of curcumin on P-glycoprotein in primary cultures of rat hepatocytes. Life Sci 62(25):2349–2358. https://doi.org/10.1016/s0024-3205(98)00216-1
Mapoung S, Pitchakarn P, Yodkeeree S, Ovatlarnporn C, Sakorn N, Limtrakul P (2016) Chemosensitizing effects of synthetic curcumin analogs on human multi-drug resistance leukemic cells. Chem Biol Interact 244:140–148. https://doi.org/10.1016/j.cbi.2015.12.001
Gao L, Zhao P, Li Y, Yang D, Hu P, Li L, Cheng Y, Yao H (2020) Reversal of PGlycoprotein-mediated multidrug resistance by novel curcumin analogues in paclitaxel-resistant human breast cancer cells. Biochem Cell Biol. https://doi.org/10.1139/bcb-2019-0377
Yang L, Li D, Tang P, Zuo Y (2020) Curcumin increases the sensitivity of K562/DOX cells to doxorubicin by targeting S100 calcium-binding protein A8 and P-glycoprotein. Oncol Lett 19(1):83–92. https://doi.org/10.3892/ol.2019.11083
Junkun L, Erfu C, Tony H, Xin L, Sudeep KC, Mingliang Z, Yanqin W, XiangQian Q (2016) Curcumin downregulates phosphate carrier and protects against doxorubicin induced cardiomyocyte apoptosis. Biomed Res Int 2016:1980763. https://doi.org/10.1155/2016/1980763
Tefas LR, Sylvester B, Tomuta I, Sesarman A, Licarete E, Banciu M, Porfire A (2017) Development of antiproliferative long-circulating liposomes co-encapsulating doxorubicin and curcumin, through the use of a quality-by-design approach. Drug Des Dev Ther 11:1605–1621. https://doi.org/10.2147/DDDT.S129008
Gu Y, Li J, Li Y, Song L, Li D, Peng L, Wan Y, Hua S (2016) Nanomicelles loaded with doxorubicin and curcumin for alleviating multidrug resistance in lung cancer. Int J Nanomed 11:5757–5770. https://doi.org/10.2147/IJN.S118568
Patel VA, Dunn MJ, Sorokin A (2002) Regulation of MDR-1 (P-glycoprotein) by cyclooxygenase-2. J Biol Chem 277(41):38915–38920. https://doi.org/10.1074/jbc.M206855200
Steinbach G, Lynch PM, Phillips RK, Wallace MH, Hawk E, Gordon GB, Wakabayashi N, Saunders B, Shen Y, Fujimura T, Su LK, Levin B, Godio L, Patterson S, Rodriguez-Bigas MA, Jester SL, King KL, Schumacher M, Abbruzzese J, DuBois RN, Hittelman WN, Zimmerman S, Sherman JW, Kelloff G (2000) The effect of celecoxib, a cyclooxygenase-2 inhibitor, in familial adenomatous polyposis. N Engl J Med 342(26):1946–1952. https://doi.org/10.1056/NEJM200006293422603
Sui H, Zhou S, Wang Y, Liu X, Zhou L, Yin P, Fan Z, Li Q (2011) COX-2 contributes to P-glycoprotein-mediated multidrug resistance via phosphorylation of c-Jun at Ser63/73 in colorectal cancer. Carcinogenesis 32(5):667–675. https://doi.org/10.1093/carcin/bgr016
Ratnasinghe D, Daschner PJ, Anver MR, Kasprzak BH, Taylor PR, Yeh GC, Tangrea JA (2001) Cyclooxygenase-2, P-glycoprotein-170 and drug resistance; is chemoprevention against multidrug resistance possible? Anticancer Res 21(3C):2141–2147
Acknowledgements
The authors would like to acknowledge the Department of Biotechnology, School of Bioengineering, College of Engineering and Technology, Faculty of Engineering and Technology, SRM Institute of Science and Technology, Kattankulathur-603203, Tamil Nadu, India, for the financial support.
Author information
Authors and Affiliations
Contributions
NS conceived the project and designed the experiments. Experiments were performed by JJ, SHV, SA, ARW and KG. Result analysis was performed by JJ, SHV, SA, SS, NS and KG. Manuscript was prepared was by SS and NS. The authors have read and have approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Jayarajan, J., Angandoor, S., Vedulla, S. et al. Curcumin induces chemosensitization to doxorubicin in Duke’s type B coloadenocarcinoma cell line. Mol Biol Rep 47, 7883–7892 (2020). https://doi.org/10.1007/s11033-020-05866-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-020-05866-w