
Abstract
This study explores the effect of flooding on the vegetation and soil of the water level fluctuation zone (WLFZ) in the Jiangjin section of the Yangtze River. The vegetation community and physical and chemical soil properties at water level elevations of 188–200 m above mean sea level (a.m.s.l) were studied using the belt transect method and the quadrat method when the section was at its lowest water level, with the nonfluctuating zone above 200 m a.m.s.l as a control. Perennial and flood-resistant species, such as Phalaris arundinacea, were dominant in the herbaceous community from 193 to 200 m, and the apparent lowest elevation of plant growth was from 188 to 193 m. From 193 to 206 m, soil bulk density decreased with increasing elevation but increased with soil layer depth. The soil nitrogen (N), phosphorus (P) and potassium (K) in the nonfluctuating zone were higher than those in the WLFZ zone. However, the soil organic matter from 193 to 200 m was higher than that in the nonfluctuating zone. The soil N, P and K from 188.06 to 188.09 m were higher than those from 188.3 to 189 m, except for hydrolysable N. The lowest elevation of plant growth did not result from a lack of soil fertility due to long-term submergence.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The water level fluctuation zone (WLFZ), which is also called the drawdown area, fluctuation zone, riparian zone, or lakeside zone, refers to the land area between the highest water level and the lowest water level in a river, lake, or reservoir due to seasonal and temporal water level fluctuations (Lu et al. 2015). It is an independent, seasonal wetland ecosystem perennially controlled by moisture gradients and has dual terrestrial and aquatic properties (Cheng et al. 2010). In this area, terrestrial and aquatic ecosystems alternate and form an independent ecological system (Minshall and Rugenski 2007) that plays an important role in water conservation, flood storage, drought control, biodiversity maintenance, ecological balance, and ecological service functions, and it is a natural protective barrier for rivers (Li et al. 2008). Based on different causes of water level fluctuation, the WLFZ can be divided into a natural water level fluctuation area and a reservoir water level fluctuation area. The former, which is formed by natural factors (Lu et al. 2015), refers to the riparian land area that is submerged and exposed. This area is generally submerged during the wet season and exposed during the dry season due to seasonal fluctuations in water levels.
Important substance and energy exchanges occur in soils in various ecosystems (Zhang and Shangguan 2006). Water level fluctuations change the physicochemical properties and nutrient contents of soils within WLFZs, thereby affecting the soil quality, and affect the vegetation in WLFZs, leading to changes in plant community composition, structures, and distribution patterns (Xu et al. 2015). Due to different immersion times, soils at different elevations in WLFZs form seasonal dry and wet backwater zones (Riis and Hawes 2003). Additionally, the soils in WLFZs are characterized by high spatial and temporal variability due to the influence of alternating water inundation and exposure, which in turn affects the entire aquatic and terrestrial ecosystem (Graf 2006). Previous studies have mainly focused on the physicochemical properties of soils affected by soil erosion (Fullen and Brandsma 1995) and changes in soil fertility, soil available nitrogen (N), dry and wet season alternation (Chacón et al. 2008), and soil phosphorus (P) due to natural hydrologic fluctuations (Vicente et al. 2010), especially in areas close to the wetland shoreline (Lishawa et al. 2010). Other studies have analyzed the absorption of P in the WLFZ, the purification mechanisms of N and P in the riparian buffer zone (Vicente et al. 2010) and the changes in the water levels of the Laurentian Great Lakes caused by climate change, which altered coastal wetland ecosystems and soil nutrient concentrations (Lishawa et al. 2010). Although the aforementioned studies addressed soil nutrient changes due to water level fluctuations or dry and wet season alternations, few studies have analyzed vegetation distributions and physical and chemical soil properties over elevation gradients.
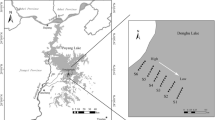
Three Gorges Reservoir is located in the upper reaches of the Yangtze River, which is the longest river in China. This river extends 600 km from Sandhouping, Yichang, Hubei, in the east to Jiangjin, Chongqing, in the west (Diao and Huang 1999). The drawdown area west of Three Gorges Reservoir is the natural WLFZ of the river, which is only affected by the natural flood season during the summer (from May to October). However, after the completion of the Three Gorges Reservoir dam, the hydrological characteristics of the Jiangjin section in the upstream of the dam changed due to effects of flood discharges from the reservoir in summer and autumn under the current schedule of water level management (Fig. 1). The associated hydrological characteristics include a prolonged natural flood time and increased depth of natural water level fluctuations. For example, the highest water level of Mutong Reservoir (which is located in a dual drawdown area) in Chongqing, Changshou-Yubei District, was 180 m above mean sea level (a.m.s.l.) in the 2012 flood season, and the highest impounded water level of the reservoir was 175 m a.m.s.l. Additionally, the highest flood level in Shangdukou, Baisha Town, Jiangjin, which is located in the upper reach of the reservoir, increased from 200 m a.m.s.l. to 208.79 m a.m.s.l. on July 24, 2012, and the flood duration increased from 1 to 2 d to 10–15 d. Therefore, the dam influences soils, spatial distributions of plants, and species compositions in the natural WLFZ. Jiang and Cai (2000) studied the species composition, species diversity, and plant community responses to changes in water levels before and after constructing Three Gorges reservoir. Xu et al. (2015) studied the vegetation at different elevations in the WLFZ to assess species composition, community diversity and spatial distribution. Shen et al. (2013) investigated the distribution and transformation of nutrients in large-scale lakes and reservoir, in which they reviewed the transformation characteristics of nutrients taking the Three Gorges Reservoir as an example. Gao et al. (2016) explored the nutrients, biochemical indexes and heavy metals in the Three Gorges Reservoir to assess the quality of water. However, there is limited literature regarding the association of vegetation with soil nutrients in the natural WLFZ of the Yangtze River upstream of the reservoir. Based on the management strategy of “storing clear water and discharging muddy water” in Three Gorges Reservoir, the stability of the natural WLFZ in the upper reaches of the reservoir plays an important role in the operation of the reservoir. Under this scenario, a low operating water level of 145 m a.m.s.l. is maintained in the summer, and a high operating water level of 175 m a.m.s.l. is maintained in the winter. In a field survey, we found that few plants grew in the natural WLFZ between the lowest water level in the dry season (188 m a.m.s.l.) and a specific threshold height of 193 m a.m.s.l. Although Veronica undulata and some other species were occasionally found, they were sparsely distributed and small. Therefore, this area is defined as the initial lowest elevation of plant growth in the WLFZ.

Lowest elevation of plant growth, sampling sites, and sampling diagram
This study analyzed the Three Gorges Reservoir area in China to explore the effects of flooding on vegetation and soil in the WLFZ. By studying the characteristics of vegetation and soil in the natural WLFZ in the Jiangjin section (the upper reaches of the reservoir) of the Yangtze River, this research investigated whether the lowest elevation of plant growth in the WLFZ is unsuitable for plant growth due to the release of N, P, and K into the water, which decreases the soil fertility, as a result of long-term water level fluctuations (Guo et al. 2012). The findings of this study may shed new light on vegetation restoration and ecological restoration in the WLFZ.
Materials and Methods
Study Area
Jiangjin is located at 28°28′~29°28′ E and 105°49′~106°36’ N at the southeast edge of the Sichuan Basin. It straddles three geomorphological areas: a parallel ridge and valley to the east of the basin, hills to the south of the basin, and a hilly area featuring hills and low mountains on the rim of the basin. In the hilly area, hills, mountains, and flatland account for 65.1%, 31.8%, and 3.1% of the terrain (Jiangjin County Annals Editorial Board 1995). The Yangtze River runs 127 km through the northern part of Jiangjin, extending from Shibatuo, Yangshi Township, in the west to Dazhongba, Luohuang Town, in the east. Flatland and terrace areas consisting of floodplains and dams in the middle of the river were formed by the accumulation of alluvial deposits from the Yangtze River and are mainly distributed along the Yangtze River. Flatland and terrace areas are mainly composed of gravel riverbed layers. Although the soil is fertile, it is easily flooded and eroded by river water in the flooding season and is prone to seasonal damage and erosion. The climate is characterized as a northern hemisphere subtropical monsoon climate zone with an average annual precipitation of 1000 mm. However, the distribution is heterogeneous and mainly concentrated in the summer (from May to October), when 78% of rainfall occurs. Notably, rainfall from June to September accounts for 56% of the annual total, often leading to summer floods.
Sample Sites and Botanical Investigation
Three sample sites were established in Shilanzi (upstream), Shangdukou (midstream), and Xiaoshunba (downstream) in the Shimen-Jingangbei section of the Jiangjin section. Three parallel transects were established in the direction of water flow at elevations of 188–193 m a.m.s.l, 193–197 m a.m.s.l. and 197–200 m a.m.s.l. in the WLFZ. A nonfluctuating transect (elevation above 200 m a.m.s.l.) was used as a control. Each transect was 200 m long. A total of 30 small quadrats of 2–3 × 2–3 m were set in each transect. Specifically, the size of the quadrats was determined using a light stick with red marks at lengths of 2, 2.5, and 3 m, respectively, as well as based on the height of the plants. After the quadrat was determined, it was delineated with a 1-m-long stick with a scale of 5 cm at each corner. The plant species and plant heights in each quadrat were recorded.
Soil Sampling and Measurement
In late March 2013, Shilanzi, Shangdukou, and Xiaoshunba in the Jiangjin section were used as sampling sites. Based on the distribution of plants in the WLFZ between 188 and 200 m a.m.s.l., elevation transects were established from 188 to 193 m, 193–197 m, and 197–200 m. The gradient of the soil profile was measured using a simple homemade slope meter. The 193–197 m transect was sampled from 195 to 195.5 m with a gradient of 5°-40°, the 197–200 m transect was sampled from 198.5 to 199 m with a gradient of 6°- 40°, and the nonfluctuating zone above 200 m was sampled from 205 to 206 m with a gradient of 10° ~ 60°. Soil profiles were dug every 10–15 m. In each soil profile, sampling was performed in the surface layer (A) 0–5 cm, the second layer (B) 40–45 cm (where the plant roots reached), and the third layer (C) (where the plant roots did not reach). Six soil profiles were dug at each sample site and transect (18 total). The lowest elevation of plant growth was 188–193 m a.m.s.l. In this transect, the soil was typical sandy soil with a thickness of 5–12 cm and a gradient of 7°-10°. The highest point of the lowest elevation of plant growth was 2.5–6.0 m above the water slope. A 0–10 cm soil sample was extracted from 15 cm beneath the plant slope as the lowest elevation of plant growth (188.3–189 m). Additionally, a 0–10 cm soil sample was extracted from 50 cm above the water slope as the elevation beneath the lowest elevation of plant growth (188.06–188.09). A soil profile was extracted every 15–20 cm. Six samples were extracted from the lowest elevation of plant growth and the elevation beneath the lowest elevation of plant growth, as shown in Fig. 1. One kilogram of soil was extracted from each layer, placed into a plastic bag, and air dried in the laboratory. The air-dried soil was sieved through a no. 18 mesh and poured into 250-ml wide-mouth bottles for preservation. Then, the soil was quartered. One-quarter of the soil that had been sieved through the no. 18 mesh was sieved through a no. 60 mesh and poured into 60-ml wide-mouth bottles for preservation. The cutting ring method was adopted to measure the soil bulk density, and the alkaline hydrolysis diffusion method (Institute of Soil Science, Chinese Academy of Sciences 1980) was used to measure soil hydrolyzable N (a general term for ammonium N, nitrate N, amino acids, amide, and hydrolytic protein N that can be hydrolyzed by acids or alkalis to produce NH4+ or NH3). Spectrophotometry was used to measure the soil available P (Shimadzu UV spectrophotometer), ammonium acetate extraction-flame photometry was used to measure the soil available potassium (K) (TAS-990 flame atomic absorption spectrometer), and a CHNOS elemental analyzer (Germany) was used to measure the total N and organic matter in the soil (Bao 2008; Yang et al. 2008).
Statistical Analysis
SPSS 16.0. LSD was used for one-way ANOVA and analyses of the soil bulk density, organic matter, N, P and K at different elevations and in different soil layers at the same elevation. Multiple comparisons and the Duncan test were conducted to identify significant differences. Microsoft Excel 2010 was used for statistical analysis and plotting.
Results
Vegetation in the WLFZ
There are 61 vascular plant species in the WLFZ (Flora of China Editorial Board of Chinese Academy of Sciences 1953–2004). Most of them (53 species) are herbaceous plants, with submergence-resistant Phalaris arundinacea, Hemarthria compress, Cynodon dactylon being the dominant species. At a water level elevation of 188–193 m, 9 vascular plant species were observed, with biennial Veronica undulata as the dominant species. There were 36 vascular plant species from 193 to 197 m, including the annuals Polygonum aviculare, Solanum nigrum, and Artemisia annua and the biennial Vicia sativa as the casual species and Phalaris arundinacea, Polypogon fugax, Hemarthria compress and Cynodon dactylon as the dominant species (Phalaris arundinacea, Hemarthria compress, and Cynodon dactylon were resistant to submergence). There were 57 vascular plant species from 197 to 200 m. Perennials Oenanthe sinensis, Centella asiatica, Amaranthus philoxeroides and Solanum lyratum and annuals Bidens pilosa and Artemisia annua were only distributed from 197 to 200 m. Submergence-resistant perennials Cynodon dactylon and Scirpus triqueter and annuals Polypogon fugax and Youngia japonica were distributed from 193 to 200 m. Biennial Veronica undulata was distributed from 188 to 200 m. Amaranthus philoxeroides, Solanum lyratum, Bidens pilosa, and Artemisia annua were only distributed from 197 to 200 m. Above 200 m, a special distribution pattern was formed by arbors (such as Erythrina variegata, Cinnamomum camphora, Broussonetia papyrifera, and Dimocarpus longan), shrubs (such as Zanthoxylum armatum and Morus alba), perennial herbs (such as Selaginella uncinata, Pteridium aquilinum, Anemone hupehensis and Dendranthema indicum), and annual herbs (such as Rostellularia procumbens and Equisetum ravens), with vines such as Pueraria lobata sparsely distributed.
The plant specie growing in river banks at Jiangjin section of Yangtze River from 188 to 200 m (a.s.l) are shown in Tables 1, 2 and 3.
Soil Bulk Density
As shown in Fig. 2, the soil bulk density increased with soil layer depth. Furthermore, the soil bulk density at lower elevations was higher than that at higher elevations, i.e., the soil bulk density from 193 to 197 m was higher than that from 197 to 200 m.

Elevation gradient and soil bulk density values of different soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil Organic Matter
The organic matter at the elevation beneath the lowest elevation of plant growth (188.06–188.09 m) was 55.408 g.kg−1, which was greater than that at the lowest elevation of plant growth (188.3–189 m; 55.361 g.kg−1). However, the organic matter above 193 m decreased with increasing elevation and soil layer depth. From 193 to 197 m, A = 77.952 g.kg−1 > B = 64.097 g.kg−1 > C = 61.124 g.kg−1. From 197 to 200 m, A = 69.156 g.kg−1 > B = 67.999 g.kg−1 > C = 57.952 g.kg−1. In the transect above 200 m, A = 65.652 g.kg−1 > B = 32.222 g.kg−1 > C = 31.559 g.kg−1 (Fig. 3).

Elevation gradients and soil organic matter. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil Total Nitrogen and Soil Hydrolyzable Nitrogen
In Fig. 4, soil total N in the transect above 200 m decreased as the soil layer depth increased, with values of A = 1.892 g.kg−1 > B = 1.293 g.kg−1 > C = 1.160 g.kg−1, and these values were larger than those from 188 to 200 m in the WLFZ. Soil total N from 194 to 197 m decreased with increasing soil layer depth as follows: A = 0.736 g.kg−1 > B = 0.703 g.kg−1 > C = 0.599 g.kg−1. From 197 to 200 m, the soil total N in the second layer was higher than that in the first and third layers, with values of A = 0.842 g.kg−1 < B = 0.924 g.kg−1 > C = 0.792 g.kg−1. Thus, the soil total N at the elevation beneath the lowest elevation of plant growth (0.207 g.kg−1) was slightly higher than that at the lowest elevation of plant growth (0.141 g.kg−1).

Elevation gradients and soil total nitrogen in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
The soil hydrolyzable N at the elevation beneath the lowest elevation of plant growth was 21.366 mg.kg−1, which was greater than that at the lowest elevation of plant growth (21.100 mg.kg−1). From 193 to 197 m, A = 35.463 mg.kg−1 < B = 44.053 mg.kg−1 > C = 38.474 mg.kg−1. From 197 to 200 m, A = 35.463 mg.kg−1 < B = 44.053 mg.kg−1 > C = 38.474 mg.kg−1. Above 200 m, A = 55.507 mg.kg−1 > B = 48.826 mg.kg−1 > C = 41.116 mg.kg−1 (Fig. 5).

Elevation gradients and soil hydrolyzable nitrogen in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil Total Phosphorus and Available Phosphorus
Soil total P slightly changed based on the soil layer. Soil total P at the elevation beneath the lowest elevation of plant growth was 0.882 g.kg−1, which was less than the value of 0.885 g.kg−1 at the lowest elevation of plant growth. From 193 to 197 m, A = 0.923 g.kg−1 > B = 0.883 g.kg−1 > C = 0.818 g.kg−1. From 197 to 200 m, A = 0.951 g.kg−1 > B = 0.827 g.kg−1 < C = 0.831 g.kg-1. In the transect above 200 m, total phosphorous displayed the following trend: A = 0.908 g.kg−1 > B = 0.752 g.kg−1 > C = 0.721 g.kg−1, as shown in Fig. 6.

Elevation gradients and soil total phosphorus in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil available P at the elevation beneath the lowest elevation of plant growth was 3.4 mg.kg−1, which was greater than 2.7 mg.kg−1 at the lowest elevation of plant growth. From 193 to 197 m, A = 5.0 mg.kg−1 > B = 4.0 mg.kg−1 > C = 2.9 mg.kg−1. From 197 to 200 m, A = 5.0 mg.kg−1 > B = 3.0 mg.kg−1 > C = 2.8 mg.kg−1. Above 200 m, A = 11.4 mg.kg−1 > B = 0.9 mg.kg−1 = C = 0.9 mg.kg−1. The available P contents in the second and third soil layers were very low (less than 1.0 mg.kg−1), as shown in Fig. 7.

Elevation gradients and soil available phosphorus in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil Total Potassium and Rapidly Available Potassium
Soil total K at the elevation beneath the lowest elevation of plant growth was 1.504 g.kg−1, which was greater than that at the lowest elevation of plant growth (1.483 g.kg−1). From 193 to 197 m, A = 1.523 g.kg−1 < B = 1.566 g.kg-1 > C = 1.531 g.kg−1. From 197 to 200 m, A = 1.543 g.kg-1 < B = 1.581 g.kg−1 > C = 1.566 g.kg−1. In the transect above 200 m, A = 1.609 g.kg−1 < B = 1.657 g.kg−1 > C = 1.648 g.kg−1 (Fig. 8).

Elevation gradients and soil total potassium in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Soil available K at the elevation beneath the lowest elevation of plant growth was 28.5 mg.kg−1, which was greater than that at the lowest elevation of plant growth (27.6 mg.kg−1). From 193 to 197 m, A = 33.8 mg.kg−1 > B = 32.1 mg.kg−1 > C = 27.5 mg.kg−1. From 197 to 200 m, A = 34.8 mg.kg−1 > B = 33.1 mg.kg−1 > C = 31.2 mg.kg−1. In the transect above 200 m, A = 41.7 mg.kg−1 > B = 30.3 mg.kg−1 < C = 31.8 mg.kg−1 (Fig. 9).

Elevation gradients and soil available potassium in soil layers. The soil thickness from 188 to 193 m was 5–10 cm. At this elevation, the soil sample was from the 0 to 10 cm layer (indicated by A). At other elevations, soil samples were from three layers, namely, 1–10 cm (a), 40–45 cm (b), and 55–60 cm (c). Combinations of different letters a, b, c, d, e, f, and g indicate significant differences between elevation zones and between soil layers according to the Duncan test (P < 0.05)
Discussion
There are 61 vascular plant species in the natural WLFZ in the Jiangjin section of the Yangtze River, most of which are herbaceous plants (53 species), with flood-resistant P. arundinacea, H. compress, and C. dactylon being the dominant species. The number of plant types increased with the increase in elevation. From 188 to 193 m, only 9 species were observed; compared with 193–200 m, this elevation showed noticeable differences in plant type, plant height and density. From 193 to 197 m, 36 species were observed, with Phalaris arundinacea, Polypogon fugax, Hemarthria compress, and Cynodon dactylon being the dominant species and the annuals Polygonum aviculare and Solanum nigrum being rare species. From 197 to 200 m, 57 species were observed. Although most of the species were herbaceous plants, shrubs such as Salix variegate, Debregeasia longifolia, Buddleja lindleyana, Morus alba, and Amorpha fruticosa were scattered. The vegetation distribution in WLFZs is controlled to some extent by natural water-level fluctuation (Carmignani and Roy 2017). Flooding is the most serious interference to vegetation distribution in WLFZs; it affects vegetation composition, species diversity, and plant growth (Stromberg et al. 2007). Large-scale fluctuations in water level control the characteristics of the WLFZ and the associated ecological processes (Battaglia and Collins 2006). The seasonal fluctuation zone changes environmental factors such as illumination, pressure, and oxygen contents in the area and inhibits plant photosynthesis, respiration, growth, reproduction, and phenology. Additionally, these fluctuations alter the original vegetation distribution (Urbanc-Bereic and Gaberscik 2004); species composition, abundance, and coverage; and ecological functions. Aquatic environment changes can also lead to changes in species life forms, with a higher proportion of annual and biennial plants (Nilsson et al. 1991) and a lower proportion of perennials. In addition, water level variations can weaken ecosystem functions in the WLFZ and cause the original vegetation to be replaced by invasive species (Holmes et al. 2008). Magee and Kentula (2005) found that minor changes in average water levels (10 cm) could promote a shift from assemblages dominated by natives to those dominated by invasive or alien taxa in the Pacific Northwest of the United States.
Soil is the basis of plant growth and development in the WLFZ, and N, P, and K are three elements essential for plant growth and development. In the WLFZ, the transfer of solutes into water is partly controlled by hydrographic features (Ledesma et al. 2016). Repeated and periodic exposure, submergence, and erosion due to water level fluctuations and sedimentation can cause a series of changes in topography and soil and water conditions (Urbanc-Bereic and Gaberscik 2004). Additionally, soil is washed away via long-term erosion, peeling, and scouring (Yu et al. 2017). Based on erosion monitoring data from the WLFZ within the 156-m water level cycle of Three Gorges Reservoir, the soil erosion intensity of the mainstream WLFZ is 71 mm/a, while the soil erosion intensity in the reservoir bay is only 11 mm/a (Bao and He 2011). The annual soil erosion in Three Gorges Reservoir is 18,476. 27 × 104 t/a (Liu et al. 2009), and sedimentation occurs downstream. Fluctuations in water level can also change the physicochemical properties of soil in the WLFZ and cause variations in soil nutrients, which affect the soil quality. Within the range of 188–200 m a.m.s.l. in the WLFZ in the Jiangjin section of the Yangtze River, the 188–193 m transect features gravel or sandy soil with a thickness of 5–12 cm, the 193–200 m transect features sandy soil and sandy loam, and the soil in the nonfluctuating zone above 200 m a.m.s.l. is loam. Water and seed resources are the most important conditions for vegetation restoration within a short time. Even if the water content is sufficient, disseminule shortages, poor nutrition, and harsh living conditions can inhibit large-scale, spontaneous vegetation restoration (Sliva and Pfadanhauer 1999).
From 188 to 200 m in the WLFZ, total N at the elevation beneath the lowest elevation of plant growth (0.207 g.kg−1) was higher than that at the lowest elevation of plant growth (0.141 g.kg−1), and hydrolyzable N at the elevation beneath the lowest elevation of plant growth was 21.366 mg.kg−1, which was higher than that at the lowest elevation of plant growth (21.100 mg.kg−1). Soil available phosphorous and available K at the elevation beneath the lowest elevation of plant growth were 28.5 mg.kg−1 and 3.4 mg.kg−1, respectively, which were higher than those at the lowest elevation of plant growth, which were 27.6 mg.kg−1 and 2.7 mg.kg−1, respectively. Therefore, N, P, and K concentrations at the elevation beneath the lowest elevation of plant growth (188.06–188.09 m) were not lower than those at the lowest elevation of plant growth (188.3–189 m). Although the sandy soil was only 5–12 cm thick from 188 to 193 m, N, P, and K concentrations were lower than those from 193 to 200 m in the WLFZ. The contents of total N, hydrolyzable N, soil available phosphorous, and available K were higher at the elevation beneath the lowest elevation of plant growth than those at the lowest elevation of plant growth. Therefore, the reason why the lowest elevation of plant growth is unsuitable for plant growth is not long-term submergence in the flood season, which has been generally assumed to decrease soil nutrient contents to levels lower than those necessary for plant growth. However, the boundary may exist due to a shortage of plant seeds or a lack of propagules. In a field survey, rare plant propagules were found from 188 to 193 m a.m.s.l. in the WLFZ, and plant seedlings of the grass family 5~10 cm in height were occasionally observed. While the lowest elevation of plant growth within a certain elevation range in the WLFZ cannot be attributed to the decline in soil fertility due to long-term submergence, submergence may wash away plant seeds in the WLFZ. Furthermore, the soil layer in this area is thin and unfavorable for seed accumulation. Because the lowest elevation of plant growth likely resulted from a shortage of plant seeds, further studies are necessary. In addition, this study suffers from another limitation. Plant growth is influenced by multiple factors, such as climatic (e.g., water, temperature, light illumination, etc.), soil, and biological factors. This study focused on the influence of soil nutrition on the formation and development of plant communities over different altitudinal gradients within the WLFZ. Therefore, the influence of other factors remains to be explored.
In conclusion, 61 vascular plants were observed in the Jiangjin section of the Yangtze River in China, with flood-resistant herbaceous plants, including perennials such as P. arundinacea, H. compress, and C. dactylon and annuals such as P. fugax, as dominant species, forming herbaceous plant communities. Due to long-term flooding stress, the lowest elevation of plant growth was observed from 188 to 193 m a.m.s.l. The thickness of the sandy soil at this elevation was 5–12 cm, which was far thinner than that from 193 to 200 m a.m.s.l., and the contents of N, P, and K at this elevation were lower than those from 193 to 200 m a.m.s.l. However, at the elevation beneath the lowest elevation of plant growth (188.06–188.09 m a.m.s.l.), these concentrations were not lower than those at the lowest elevation of plant growth (188.3–189 m a.m.s.l.). The existence of the lowest elevation of plant grow in the WLFZ is not caused by a decrease in soil fertility because long-term submergence does not cause a significant release of soil nutrients into the water. Rather, its existence is very possibly a result of other factors such as a lack of plant disseminules. If this is the case, other factors, such as plant seed storage and plant disseminules, may be taken into consideration for the restoration of vegetation in the WLFZ.
References
Bao SD (2008) Soil agricultural and chemical analysis, 3rd edn. China Agriculture Press, Beijing
Bao YH, He XB (2011) Preliminary Study on Soil Erosion at the Water-Level-Fluctuating Zone of the Three-Gorges Reservoir Research of Soil and Water Conservation 18:190–195
Battaglia LL, Collins BS (2006) Linking hydroperiod and vegetation response in Carolina bay wetlands. Plant Ecol 184:173–185
Carmignani JR, Roy AH (2017) Ecological impacts of winter water level drawdowns on lake littoral zones: a review. Aquat Sci 79:1–22
Chacón N, Dezzeo N, Range M, Flores S (2008) Seasonal changes in soil phosphorus dynamics and root mass along a flooded tropical forest gradient in the lower Orinoco River, Venezuela. Biogeochemistry 87:157–168
Cheng RM, Wang XR, Xiao WF, Guo QS (2010) Advances in studies on water-Leve1-fluctuation zone. Sci Silvae Sin 46:111–119
Diao CT, Huang JH (1999) A preliminary study on land resources of the water-level-fluctuating zone in the three gorges reservoir. Resour Environ Yangtze Basin 8:75–80
Flora of China Editorial Board of Chinese Academy of Sciences (1953–2004) Flora of China (Vol. 2–80). Science Press, Beijing
Fullen MA, Brandsma RT (1995) Property changes by erosion of loamy sand soils in east Shropshire, UK. Soil Technol 8:l–15
Gao Q, Li Y, Cheng Q, Yu M, Hu B, Wang Z, Yu Z (2016) Analysis and assessment of the nutrients, biochemical indexes and heavy metals in the three gorges reservoir, China, from 2008 to 2013. Water Res 92:262–274
Graf WL (2006) Downstream hydrologic and geomorphic effects of large dams on American rives. Geomorphology 79:336–360
Guo QS, Kang Y, Zhao YJ, Hong M, Kong QQ, Nie BH (2012) Changes in the contents of N, P, K, pH and organic matter of the soil which experienced the hydro-fluctuation in the three gorges reservoir. Sci Silvae Sin 48:7–10
Holmes PM, Esler KJ, Richardson DM, Witkowski ETF (2008) Guidelines for improved management of riparian zones invaded by alien plants in South Africa. S Afr J Bot 74:538–552
Institute of Soil Science, Chinese Academy of Sciences (1980) Analysis of Soil Physico-Chemical Properties. Shanghai Science and Technology Press, Shanghai
Jiang MX, Cai QH (2000) Preliminary studies on the riparian plant communities of main channel in the three gorges area, the Yangtze River. Acta Hydrobiol Sin 24:458–463
Jiangjin County Annals Editorial Board (1995) Jiangjin County annals. Sichuan Science and Technology Press, Chengdu
Ledesma JLJ, Futter MN, Laudon H, Evans CD, Kohler SJ (2016) Boreal forest riparian zones regulate stream sulfate and dissolved organic carbon. Sci Total Environ 560–561:110–122
Li DL, Han L, Ruan HH, Zhang JL (2008) Analysis of soil physical and chemical properties of riparian zone along Qinhuai River. J Nanjing For Univ (Nat Sci Ed) 32:17–22
Lishawa SC, Albert DA, Tuchman NC (2010) Water level decline promotes Typha X glauca establishment and vegetation change in Great Lakes coastal wetlands. Wetlands 30:1085–1096
Liu A, Wang J, Liu ZJ (2009) Remote sensing quantitative monitoring of soil erosion in three gorges reservoir area: a GIS /RUSLE - based research. J Nat Dis 18:25–30
Lu MQ, Wu SJ, Chen CD, Jiang Y, Wen ZF, Chen JL et al (2015) A review of studies on water level fluctuating zone (WLFZ) of the three gorges reservoir(TGR)based on bibliometric perspective. Acta Ecol Sin 35:3504–3518
Magee TK, Kentula ME (2005) Response of wetland plant species to hydrologic condition. Wetl Ecol Manag 13:163–181
Minshall GW, Rugenski A (2007) Riparian processes and interactions. Methods Stream Ecol 2:721–742
Nilsson C, Ekblad A, Gardfjell M, Carlberg B (1991) Long-term effects of river regulation on river margin vegetation. J Appl Ecol 28:963–987
Riis T, Hawes I (2003) Effect of wave exposure on vegetation abundance,richness and depth distribution of shallow water plants in a New Zealand lake. Freshw Biol 48:75–88
Shen Z, Niu J, Wang X, Wang H, Zhao X (2013) Distribution and transformation of nutrients in Large-Scale Lakes and reservoirs: the three gorges reservoir. Springer Science & Business Media. http://www.springer.com/us/book/9783642349638
Sliva J, Pfadanhauer J (1999) Restoration of cut-over raised bogs in southern Germany: a comparison of methods. Appl Veg Sci 2:137–148
Stromberg JC, Beauchamp VB, Dixon MD, Lite SJ, Paradzick C (2007) Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in arid South-Western United States. Freshw Biol 52:651–679
Urbanc-Bereic O, Gaberscik A (2004) The relationship of the processes in the rhizosphere of common reed phragmites australis(Cav.),TRIN. Ex STEDEUL to water fluctuation. Int Rev Hydrobiol 89:500–507
Vicente ID, Andersen FØ, Hansen HCB, Cruz-Pizarro L (2010) Water level fluctuations may decrease phosphate adsorption capacity of the sediment in oligotrophic high mountain lakes. Hydrobiologia 651:253–264
Xu JX, Peng GY, Wang JZ (2015) Diversity and spatial distribution of vegetation from the water-level fluctuation zone of the Xiangxi River in the three gorges reservoir area. Resour Environ Yangtze Basin 24:1345–1350
Yang JH, Wang CQ, Dai HL (2008) Soil agricultural and chemical analysis and environment monitoring. China Land Press, Beijing
Yu C, Chen S, Zhang L, Wang ZD, Shen QS, Gao Q (2017) Phosphorus fractions and their spatial distribution in surface sediments of inflow rivers in the northeastern Lake Tanganyika. J Lake Sci 29:334–342
Zhang XB, Shangguan ZP (2006) Effect of human-induced disturbance on physical properties of soil in artificial Pinus tabulaeformis Carr. Forests of the Loess Plateau. Acta Ecol Sin 26:3685
Acknowledgements
The work was supported by the National Key R&D Program of China (2017YFC0505301), the National Natural Science Foundation of China (grant number 31770465), and Ministry of Finance & State Council Executive Office of Three Gorges Project Construction Committee of China (grant numbers 5000002013BB5200001, 5000002013BB5200002).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interests.
Rights and permissions
About this article

Cite this article
Liu, M., Liu, Y., Zeng, B. et al. Lowest Elevation of Plant Growth and Soil Characteristics in Natural Drawdown Areas in the Jiangjin Section of the Yangtze River. Wetlands 39, 381–391 (2019). https://doi.org/10.1007/s13157-018-1087-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-018-1087-5