
Abstract
Climate change is predicted to reduce Laurentian Great Lakes water levels, altering coastal wetland ecosystems and potentially stimulating invasive macrophytes, like Typha X glauca. Recent prolonged low water levels, which climaxed in 2007, created conditions comparable to those predicted by climate change science. In 2008, we examined ecosystem and plant community properties in 14 intact northern Great Lakes coastal wetlands and compared community data with data from a 1987–1989 high-water period, before T. X glauca invasion. In 2008, T. X glauca occurred in 50% of wetlands and 16% of plots; was associated with reduced Floristic Quality and increased soil organic matter, soil nutrients, and leaf litter (all p < 0.05); and plant community composition had shifted and was more homogeneous than in 1988 (both p < 0.05). Additionally, T. X glauca was more dominant when growing behind barrier beach ridges, which form in high-water conditions and persist in low-water, than in lake-exposed marshes (p < 0.05), revealing a physiographic mechanism for increased dominance. Beach ridges protect T. X glauca from wave and seiche energy, and as water levels decline, these energy-insulating microtopographic features will likely stimulate further invasion and dominance by T. X glauca, even in high quality wetlands.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Laurentian Great Lakes and the freshwater coastal wetlands fringing their shores have experiencing a profound number of invasive species introductions over the last century (Mills et al. 1993; Galatowitsch et al. 1999; Ricciardi 2001; Holeck et al. 2004; Trebitz and Taylor 2007). Furthermore, climate change is predicted to alter several environmental conditions in the Great Lakes over the next century (Magnuson et al. 1997; Lenters 2001; Sousounis and Grover 2002; Angel and Kunkel 2010) resulting in additional pathways of invasive species introduction and establishment (Rahel and Olden 2008). The synergistic impacts of climate change and invasive species on Great Lakes ecosystems will likely result in substantial changes to the ecological structure and function of many ecosystems.
Great Lakes coastal wetlands are profoundly dynamic ecosystems with hydrologic variability at three temporal scales serving as the primary natural disturbances (Keddy and Reznicek 1986; Minc 1997; Gathman et al. 2005; Trebitz 2006). In the short term, storm action, ice flow, and seiche activity cause periodic severe disturbance to nearshore emergent marshes (Trebitz 2006). Annually, Great Lakes wetlands experience cyclical seasonal water-level fluctuations (Minc 1997), with Lake Michigan, Lake Huron, and St. Mary’s River wetlands experiencing 20 to 50 cm annual fluctuations during the 20 years between 1988 and 2008 (USACE 2009). Over the course of years to decades, variable precipitation and evaporation in the Great Lakes basin induces major lake level fluctuations of 1.5–2.5 m (USACE 2009). Water level-initiated fluctuation and disturbance contributes to distinct wetland vegetation zonation, maintains ecosystem dynamism as plant species move lakeward and landward with water level fluctuations, and preserves species diversity in coastal systems (Keddy and Reznicek 1986; Maynard and Wilcox 1997; Wilcox 2004; Kost et al. 2007; Wilcox and Nichols 2008). Over the last 23 years, Lake Michigan and Huron water levels have undergone a 1.8 m peak-to-trough water level change from a high of 177.5 m above sea level in October 1986, the highest in over 90 years, to a low of 175.7 m in December 2007, the lowest water level in 43 years and within 10 cm of the 100 year low water level reached in 1964 (USACE 2009). Between these dates, more moderate oscillation in lake levels also occurred (Fig. 1).

Mean monthly Lake Michigan/Lake Huron water levels 1970–2010. Gray dots represents October 1986, the month in which water level reached its highest level in over 90 years, and December 2007, the month in which water levels reached their lowest water level in 43 years (USACE 2009)
Most climate change models predict reduced Laurentian Great Lakes water levels over the next 50–100 years (Mortsch and Quinn 1996; Magnuson et al. 1997; Lofgren et al. 2002; Kling et al. 2003; Angel and Kunkel 2010) resulting from more winter ice-free days and higher summer temperatures (Sousounis and Grover 2002), leading to net evaporation exceeding the predicted increases in regional precipitation (Magnuson et al. 1997). In the most recent published analyses, average Lake Michigan and Lake Huron water levels are projected to decrease by 0.25–0.41 m relative to the 1970–1999 average by 2080–2094, depending upon the emissions scenario (Angel and Kunkel 2010). Moreover, climate change will likely alter the timing of precipitation and snowmelt, affecting the seasonal pattern of Great Lake water levels (Lenters 2001). The predicted decline in long-term average water levels in concert with altered seasonal water level patterns will result in changes to the timing, duration, and range of Great Lakes water level fluctuations (Mortsch 1998), with consequent impacts on Great Lakes coastal wetland ecosystems. The recent low-water period in the Great Lakes, which lasted through much of the last decade (USACE 2009), mirrors conditions predicted to result from climate change, namely persistent low-water levels (Mortsch 1998; Angel and Kunkel 2010). These conditions provided an opportunity to assess the ecological impacts of predicted climate-change on coastal wetland ecosystems.
Changes in Great lakes water levels have enhanced the establishment and persistence of invasive wetland plant species (Wilcox and Meeker 1991; Boers 2005; Boers et al. 2007; Tulbure et al. 2007; Boers and Zedler 2008; Whyte et al. 2008; Wilcox et al. 2008), specifically hybrid cattail, Typha X glauca, a cross of the native T. latifolia and the invasive T. angustifolia (Smith 1987). In the Great Lakes region, T. X glauca is becoming increasingly abundant (Mills et al. 1993; Trebitz and Taylor 2007) and has been positively correlated with reduced hydrologic variability or otherwise altered hydrology (Wilcox 2004; Boers et al. 2007; Wilcox and Xie 2007; Boers and Zedler 2008; Wilcox and Nichols 2008) as well as elevated nutrients (Woo and Zedler 2002). Furthermore, recent historically low water levels have been linked to new invasions of T. X glauca into human-disturbed Lake Michigan coastal wetlands (Frieswyk and Zedler 2007; Tulbure et al. 2007). Following establishment, T. X glauca is capable of rapid invasion (Tulbure et al. 2007) and can spread vegetatively (Smith 1967) at a rate of up to 4 m in clone diameter per year (Boers and Zedler 2008). Once established, T. X glauca tends to persist by tolerating a wide range of water level conditions (Harris and Marshall 1963; Waters and Shay 1990).
Typha X glauca is altering plant and microbial communities (Angeloni et al. 2006; Frieswyk and Zedler 2007; Wilcox et al. 2008), and wetland ecosystem structure and function (Freyman 2008; Farrer and Goldberg 2009) in Great Lakes coastal wetlands. Typha X glauca is often orders of magnitude larger in size than the native species it replaces (Woo and Zedler 2002). Due to its high biomass, the persistent aerial structure of its leaf and stem litter, and recalcitrance of litter to decomposition (Davis and Van der Valk 1978; Freyman 2008), large quantities of partially decomposed litter tend to accumulate in T. X glauca beds. The creation of a dense litter canopy alters microsite temperature and light conditions (Freyman 2008; Farrer and Goldberg 2009) and gradually excludes other aquatic macrophytes, resulting in reduced species richness and diversity in invaded wetlands (Farrer and Goldberg 2009; Tuchman et al. 2009; Vaccaro et al. 2009).
Observations of T. X glauca dominance in Cheboygan Marsh, Cheboygan County, Michigan (Tuchman et al. 2009), led us to investigate whether this aggressive invasive species was invading other northern Michigan coastal wetlands. Apparently unique to the region, the Cheboygan Marsh plant community is dominated by a large stand of T. X glauca (Angeloni et al. 2006; Tuchman et al. 2009) dated to the 1950’s (Vail 2009).
In 1987–1989, the Michigan Natural Features Inventory (MNFI) conducted a series of wetland vegetation and soil surveys with the purpose of identifying coastal wetlands which were large (>40 ha) and minimally disturbed (Albert et al. 1987, 1989). These data provide a unique opportunity to document new invasions into wetlands which did not harbor T. X glauca in the late 1980s and to examine long-term changes to moderate- to high-quality wetland plant communities of the northern Great Lakes.
We hypothesize that climate-change induced water level alterations will result in an expansion of T. X glauca’s range in Great Lakes coastal wetlands and increased dominance within its present range. The goals of this study were to address the following questions: 1) were the low water conditions that reached their minimum in 2007 associated with an expansion of T. X glauca into upper Great Lakes coastal wetlands characterized by broad and contiguous wetland zones and substantial buffering from agriculture and development (Albert and Minc 2004), and if so, 2) are newly formed T. X glauca stands associated with changes in plant communities and soil conditions similar to those that have been documented in more nutrient rich, human disturbed, and long-invaded systems? To answer these questions, we pursued the following objectives: 1) determine the location and spatial extent of T. X glauca stands in a subset of relatively intact northern Lake Huron and Lake Michigan coastal wetlands, 2) evaluate plant species composition and environmental correlates with T. X glauca beds and 3) assess the change in wetland plant community composition and structure from 1987 to 1989, a period of historically high lake levels, to 2008, a period of historically low lake levels (Fig. 1).
Methods
Site Selection
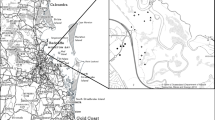
The geographic extent of the study was limited to the northern Lower Peninsula and eastern Upper Peninsula of Michigan to minimize between-wetland variability and to maximize similarity with Cheboygan Marsh, a lacustrine open embayment wetland (Fig. 2). Study sites were randomly selected from wetlands meeting the following criteria: 1) wetlands were of the regional type, Northern Great Lakes Marsh, characterized by extensive intact wetland zones, non-significant human impacts (Albert and Minc 2004), and relatively flat slopes (0.24–0.34%) (Albert et al. 1987, 1989), which limits the impact of ground-water seepage (these wetlands are hereafter called intact); 2) wetlands had open embayment or open shoreline geomorphology (Albert et al. 2005) and sand over clay substrates (Albert, unpublished data); and 3) wetlands were sampled in the 1987 and 1989 MNFI surveys (Albert et al. 1987, 1989). Of the 20 wetlands that met these criteria, we selected a total of 14 for sampling in summer 2008, representing 37% of total regional wetlands and 70% of the geomorphologically qualifying wetlands. At the time of the original MNFI surveys, T. latifolia and/or T. angustifolia were present in 12 of the 14 selected wetlands and T. X glauca was not noted as present in any wetlands (Online Resource 1, Fig. 2).

Map of study sites. Black dot represent the geographic center of each wetland sampled
Aerial Imagery Assessment
Color infrared aerial photography of the selected wetlands from 1978, and color satellite imagery from 1998, 2004, and 2007 were evaluated to determine changes in land-use, anthropogenic disturbance, and vegetation zones through time.
Field Data Collection
The response of Great Lake coastal vegetation demonstrates a lag time which has been effectively related to the previous year’s peak water levels (Frieswyk and Zedler 2007). Thus, the 1987 and 1989 wetland vegetation surveys (hereafter referred to as 1988-high water) corresponded with the 100-year high water point reached in October, 1986 and surveys conducted in summer of 2008 followed the 43-year low-water event reached in December of 2007 (hereafter referred to as 2008-low water).
In July and August 2008, data were collected as near as possible to the transect locations established and sampled in 1988-high water. Detailed hand drawn maps from the original study (Albert et al. 1987, 1989) and color aerial imagery (NAIP 2007) were used to digitize transect locations and to create wetland transect maps in a Geographic Information System (GIS). Transect locations were located in the field using maps and compass triangulation. Because of the substantial difference in water levels between sample dates, the wetland vegetation zones had shifted water-ward and terrestrial vegetation had invaded much of the drier upper edge of the wetlands. To account for these shifts and to sample vegetation from the wet meadow and emergent marsh vegetation zones, the 2008-low water transects were shifted toward the open water.
Modified vegetation sampling protocols from the Great Lakes Wetland Consortium, Great Lakes Coastal Wetland Monitoring Plan (Albert 2008) and the 1988-high water surveys (Albert et al. 1987, 1989) were used to characterize the vegetation found in the wet meadow and emergent marsh vegetation zones. One-meter square plots were established at 25-m intervals along transects extending through the wetland from the upper edge of the sedge and grass dominated wet meadow zone to the open water edge of vegetation in the emergent marsh zone. Randomly placed plots were added when necessary to ensure a minimum of three plots in each wetland zone. The number of plots sampled in wetlands varied between years based upon the width of wetland vegetation zones encountered.
Plot geographic locations were recorded using a hand held global positioning system unit (Garmin GPSMAP 60Cx Global Positioning System, Garmin International Inc., Olathe KS). Within each plot, all vascular plants were identified to species (taxonomy after Voss 1972, 1985, 1996) and the number of living stems of each species were recorded. In addition, wetland zone, distance to the open water edge of emergent vegetation, and water depth were recorded. To further characterize the environmental conditions of each plot, we recorded litter depth, organic soil depth, and mineral soil texture. Litter depth was measured from the top of the litter to the top of the organic soil surface. To control for variable heights of standing dead litter, depth was measured after compacting the standing detritus to minimize variable air space. Organic soil depth was measured using a 3/4 inch galvanized steel pipe. Mineral soil textures were determined in the field, and included sand, silt, clay, or cobble. Water depth, soil organic depth, and mineral soil textures were determined with the methods used in the original MNFI surveys. In each plot, four soil plugs were collected from the top 10 cm of the organic or mineral soil using a 1.3 cm-diameter tubular soil sampler. Soil samples were homogenized and then used for laboratory nitrate, (NO −3 ), ammonium (NH +4 ), and soil organic content analyses.
Laboratory Soils Preparation
Inorganic soil nitrogen, soil water, and soil organic matter were determined following Angeloni et al. (2006). Soil water content was determined by drying a 10 g soil sample at 105°C for 24 h and then calculated as (wet weight–dry weight)/wet weight (Gardner 1986). Soil organic matter was then determined by loss on ignition in a muffle furnace at 550°C for 2 h (APHA 2005). Soil plugs were gently sieved through a number 10, U.S.A. Standard Testing Sieve with 2 mm nominal opening (Fisher Scientific) to remove coarse root material and stones. NO −3 and NH +4 were extracted by mixing a 10 g subsample of the wet, homogenized, and sieved soil with 40 ml of 2 M KCL solution for 1 h and centrifuged at 134 rotations per minute for five minutes. The supernatant was then filtered through G8 glass fiber filters (Fisher Scientific) and frozen. NO −3 and NH +4 concentration of the extracts were determined using the Cadmium Reduction Method (APHA 2005) and the Automated Phenate Method (APHA 2005), respectively, on an Auto-Analyzer 3 (Bran-Luebbe, Farmington, MI).
Vegetation and Plant Community Analyses
Several measures of plant community quality were determined at both plot and wetland scales. Floristic Quality Assessment (FQA) is a methodology that assigns a Coefficient of Conservatism score (C) to each plant species present at a site, and uses the C scores of each plant species encountered during sampling to generate a Floristic Quality Index (FQI) score for each wetland sampling site (Herman et al. 2001). FQI scores provide a tool for comparing quality and plant diversity at the wetland scale. FQI, C, Shannon-Weiner diversity index (H′), and species richness values were calculated from the 2008 data using FQI: Michigan software (Conservation Research Institute, Elmhurst IL). Additionally, Importance Values (IV; McCune and Grace 2002) were calculated for each species at the plot scale as: (percent of total stems in plot + percent of total biomass in plot)/2.
Plant communities represented by their most dominant species (species which occurred in more than 10% of total plots) were compared between sampling years using several methods. Relative frequency values were calculated for each species (percent of total plots in which a species was present) (McCune and Grace 2002) and compared. In 2008-low water, plant species data were measured in density units (stems per m2) whereas the 1988-high water vegetation data were measured in percent coverage units. In order to compare these data, in July and August 2009 we measured all three values (relative frequency, density, and percent cover) from 88 plots and developed metrics to convert percent coverage and stem densities to above ground biomass per m2 units. These 88 plots represented a range of dominance for each species from four of our northern Great Lakes coastal study wetlands. Species from four genera (Typha, Carex, Juncus, and Salix) were grouped into genera conversion metrics due to morphological similarity among species. Above-ground plant material from each plot was separated by species, dried for 24 h at 105°C, and weighed. Percent coverage to above-ground biomass regressions, and stem density to above-ground biomass regressions were developed to convert 1988-high water and 2008-low water data to equivalent biomass values (Table 1).
Statistical Analyses
To evaluate associations among environmental parameters, and between species and environmental parameters, several univariate and multivariate approaches were used with the 2008-low water data, including data collected at Cheboygan Marsh. All univariate analyses were conducted using SYSTAT statistical software version 11.0 (SYSTAT Software Inc., 2004). All multivariate statistical analyses were performed using R (R Development Core Team 2008). Species density and environmental parameter relationships were evaluated using simple linear regression analyses. Associations between environmental parameters were compared between plots with and without Typha using two-tailed independent samples t-tests. To compare differences between plots with T. X glauca in barrier-beach protected sites with unprotected sites, we used two-tailed independent samples t-tests.
To evaluate shifts in community composition through time, several multivariate statistical methods were employed. To determine which species (as centroids) and environmental variables (as vectors) affected community composition and structure, we applied nonmetric multidimensional scaling (NMDS) ordination to the 1988-high water and 2008-low water datasets independently. To evaluate community change through time, we applied NMDS ordination of the entire data set (1988-high water and 2008-low water). NMDS is a multivariate ordination method appropriate for both linear and non-linear responses and robust for non-normal data (McCune and Grace 2002). ANOSIM (analysis of similarity), a nonparametric permutation procedure, was used to test for differences in community composition between time periods (McCune and Grace 2002). A paired two-sample t-test was used to test for changes in plant species richness through time at the wetland scale. To test for plant community homogenization, similarity between wetlands was evaluated within each time period by calculating the mean wetland similarity to the other 13 wetlands using the Bray-Curtis measure of similarity. Within a group, more homogeneous community composition would be represented by higher average Bray-Curtis similarity values. Changes in average similarity between time periods were evaluated using an independent samples t-test.
Results
Aerial Imagery Interpretation of Study Wetlands
Aerial imagery revealed that Cheboygan Marsh, the T. X glauca invaded reference wetland, has experienced the most severe human disturbance including ditching, historic shipping docks, and nearby urban land-use, including a wastewater treatment facility. All other study sites have negligible management histories, indicated by broad undeveloped buffer zones and clear wetland community zonation from emergent marsh to swamp forest. Human disturbance is evident near several wetlands: historical ditching for hay harvest occurred outside of the Pickford Point and Hursley Creek wetlands, and vacation cabins are present within 1 km of the Mismer Bay and Hursley Creek wetlands. A comparison of aerial imagery from 1978, 1998, 2004, and 2007 revealed no land-use change in any study wetlands. The principal change in conditions between sampling dates appeared to be solely due to water level changes. Aerial imagery revealed a general lakeward movement of all plant community zones, with shrubs invading the upper edge of the wet meadow zone and the wet meadow and emergent marsh zones expanding lakeward.
Location and Extent of Typha X glauca in Study Wetlands
We found that since 1989, when T. X glauca was absent from all study wetlands, T. X glauca has invaded 7 of the 14 open embayment and barrier beach lagoon wetlands included in this study (Online Resource 1), and T. X glauca was found in 16% of all plots sampled in 2008-low water conditions (Table 2).
Plant Community Composition and Environmental Correlates with Typha X glauca
In 2008, plots where T. X glauca was present shared significantly greater litter depth, soil organic matter depth, soil organic matter (%), soil water content (%), distance to open water, and NH +4 (p < 0.05, Table 3). These T. X glauca plots also shared significantly lower water depth, FQI, C, H′, species richness, and native species richness (p < 0.05, Table 3).
Within plots with Typha spp. present, significant positive linear relationships existed between Typha spp. density (stems/m−2) and litter depth and organic soil depth (p < 0.05). However, T. X glauca and T. angustifolia (narrow-leaved cattail) exhibited different environmental associations. Typha X glauca density was positively correlated with organic soil depth and distance from open water, whereas T. angustifolia density was positively correlated with water depth (p < 0.05; Fig. 3).

Correlations between Typha species densities (stems/m2) and environmental variables from 2008: Typha spp. and litter depth (cm) (a), Typha spp. and organic depth (cm) (b), Typha X glauca and litter depth (cm) (c), Typha X glauca and organic depth (cm) (d), Typha X glauca and distance to open water (m) (e), Typha angustifolia and distance to open water (m) (f), Typha X glauca and water depth (cm) (g), and Typha angustifolia and water depth (h)
In several wetlands, we observed the presence of small (<1 m tall) barrier beach ridges, which were not noted in 1988-high water (Online Resource 1). We assessed the difference in T. X glauca density, importance value, organic depth, water depth, species richness, and FQI in stands located behind newly created barrier beaches and in stands open to the lake. We found, T. X glauca was significantly more abundant (stems m−2) and had greater importance value when occurring behind barrier beaches (p < 0.05). Additionally, we found significantly greater organic soil depth (p < 0.05), significantly lower water depth (p < 0.05), and lower FQI (p < 0.05) in barrier beach-protected stands than in unprotected stands (Table 4).
An NMDS ordination of 2008 plant community data and environmental variable vectors revealed several clear patterns (Fig. 4a). The wet meadow and emergent marsh vegetation zones, represented by standard deviation ellipses, clearly separated in species space with Carex stricta, Calamagrostis canadensis, and Schoenoplectus pungens correlated with the wet meadow zone and E. smallii, S. acutus, T. angustifolia, and T. latifolia correlated with the emergent marsh zone. Typha latifolia and T. angustifolia clearly fell in the deep-water end of the emergent marsh zone, whereas T. X glauca was distributed in moderate water depths and between the wet meadow and emergent marsh zones. Additionally, compared to the adjacent wet meadow and emergent marsh vegetation zones, T. X glauca was strongly correlated with significant differences in ecologically important litter depth, organic soil depth, NH +4 , and distance to open water.

NMDS ordination of 2008-low water species composition data (a) and 1988-high water species composition (b) in biomass units. Dissimilarity was based on Bray-Curtis distances and plots were constructed using two dimensions. Fitted vector arrows are significant (p < 0.05, by permutation procedure) and their length is proportional to their explanatory strength. 2008–low water explanatory variables tested were water depth (p < 0.001), litter depth (p < 0.001), organic depth (p < 0.001), distance to open water (p < 0.001), NH +4 (ammonium concentration in soil sample) (p < 0.001), and Typha density (stems/m2) (p < 0.001). Substrate centroids (p < 0.01) are also displayed. 1988–high water explanatory variables tested were water depth (p < 0.001), organic depth (p < 0.001), and Typha presence/absence (p < 0.001). Species were limited to those found in more than 10% of total plots. Species codes: Calcan = Calamagrostis canadensis, Cxlacu = Carex lacustris, Cxstri = Carex stricta, Elesma = Eleocharis smallii, Junalp = Juncus alpinoarticulatus, Salspp = Salix spp., Schacu = Schoenoplectus acutus, Schpun = Schoenoplectus pungens, Typang = Typha angustifolia, Typgla = Typha X glauca, Typlat = Typha latifolia
In 1988-high water, plots containing Typha spp. also exhibited a series of common environmental traits: significantly greater water depth (p < 0.001), organic depth (p = 0.001), and species richness (p = 0.004) (Table 5). The NMDS ordination of 1988 species and environmental data revealed clear separation in species space between the wet meadow and emergent marsh vegetation zones, represented by standard deviation ellipses, with C. stricta, C. lacustris, C. canadensis, and Salix spp. dominating the wet meadow and S. acutus, S. pungens, and E. smallii dominating the emergent marsh zone. The water depth vector was strongly associated with the emergent marsh zone. Additionally, T. latifolia was highly correlated with the organic depth vector (Fig. 4b).
Plant Community Change Analyses
The relative frequency of plots containing the nine most commonly encountered species and three species of Typha revealed species change between study years. Carex lacustris and C. stricta decreased in relative frequency in 2008-low water, while E, smallii, J. alpinoarticulatus, Salix spp., S. acutus, S. pungens, and T. angustifolia all increased in relative frequency. In 1988-high water T. X glauca was not encountered, whereas in 2008-low water it was present in 16% of plots sampled (Table 2).
At the wetland scale, despite the increase in T. X glauca between study years, the average total vascular plant species richness did not differ significantly (Online Resource 1; paired t-test, mean 1988 = 25.4; mean 2008 = 30.1, p = 0.116). However, ANOSIM analyses revealed that 1988-high water and 2008-low water plant communities differed significantly in species composition (Global R = 0.080, p < 0.001). Furthermore, t-tests of Bray-Curtis similarity revealed that 2008-low water plant communities were significantly more similar to one another than 1988-high water plant communities overall (p < 0.001) and within those wetlands invaded by T. X glauca (p < 0.001; Fig. 5).

Mean wetland-scale Bray-Curtis plant community similarity (percent similarity) by year for all wetlands (a; N = 28; p < 0.0001) and for only those wetlands invaded by Typha X glauca since 1988 (b; N = 14; p < 0.0001). Error bars ±1 S.E
NMDS ordination of 1988-high water and 2008-low water plots demonstrated a directional shift in community composition through time. Sample-year plant community composition, represented by standard deviation ellipses, have clearly moved in the direction of the significant Typha presence/absence vector and toward the T. X glauca centroid between 1988-high water and 2008-low water (Fig. 6).

NMDS ordination of species composition data in biomass units. Dissimilarity was based on Bray-Curtis distances and plot was constructed using two dimensions. Fitted vector arrows are significant (p < 0.05, by permutation procedure) and their length is proportional to their explanatory strength. Explanatory variables tested were water depth (p < 0.001), organic depth (p < 0.001), and Typha presence or absence (p < 0.01). 1988-high water and 2008-low water ellipses represent the standard deviation of the mean plot location in ordination space. Species were limited to those found in more than 10% of total plots. Species codes: Calcan = Calamagrostis canadensis, Cxlacu = Carex lacustris, Cxstri = Carex stricta, Elesma = Eleocharis smallii, Junalp = Juncus alpinoarticulatus, Salspp = Salix spp., Schacu = Schoenoplectus acutus, Schpun = Schoenoplectus pungens, Typang = Typha angustifolia, Typgla = Typha X glauca, Typlat = Typha latifolia
Discussion
In the upper Great Lakes, was the low-water period that reached its minimum in 2007 associated with an expansion of T. X glauca into coastal wetlands characterized by low disturbance and low nutrient conditions?
A major expansion of T. X glauca has occurred throughout the northern Lake Michigan/Huron region over the last 20 years. In emergent marsh and wet meadow communities in our study wetlands, the presence of T. X glauca was positively correlated with low-water and distance from the open lake, indicating that this invasive species can dominate relatively dry portions of Great Lakes coastal wetlands under extremely low-water conditions. Similarly, increased dominance by T. X glauca has been associated with water level declines in the Great Lakes (McDonald 1955) and in western Canada (Shay and Shay 1986). Tolerance for low water conditions by T. X glauca (Waters and Shay 1990) differentiated the hybrid species from its parent species T. angustifolia and T. latifolia (McDonald 1955; Wilcox et al. 2008). Correspondingly, we found the Typha species in different habitats, with the parent species dominating deeper water sites in both years and T. X glauca dominating drier sites. We conclude that the wet meadows in the Great Lakes region, which are typically found upslope of deeper coastal emergent marshes, are at high risk of invasion by T. X glauca under low-water conditions, even in relatively unspoiled wetlands.
Furthermore, we found that T. X glauca reached the highest densities and was the most dominant when growing behind recently created barrier-beach ridges. We posit that the creation of barrier-protected wetlands during a period of reduced water levels, may be a primary physiographic driver for the increased dominance of T. X glauca in northern Great Lakes coastal wetlands. Low barrier beaches form during periods of high water and are sometimes associated with specific storm events. When water levels drop, these low barrier beaches isolate formerly exposed wetlands from the wave action and seiche disturbances that are associated with the open lake (Minc 1997). The reduced energy conditions in barrier-protected wetlands are favorable to the persistence of Typha species, which root more shallowly than other emergent wetland species, do not tolerate extreme wave action, and can be severely affected or even eliminated from coastal wetlands by rapid increases in water levels (Minc 1997; Gathman et al. 2005). Small barrier beaches and other micro-topographic features, which would otherwise be destroyed by periodic high water levels (McDonald 1955), will likely become more persistent and ecologically important with reduced future Great Lakes water levels. Furthermore, our data reveal that barrier beaches appear to affect Great Lakes coastal wetland conditions similarly to dikes built within wetlands, which have also been associated with increased Typha spp. and invasive species dominance, greater soil organic matter content, and higher nutrient levels than in un-diked wetlands (Herrick and Wolf 2005).
Anthropogenically introduced nutrients and/or altered hydrology have been demonstrated to be the principal drivers for T. X glauca invasion (Galatowitsch et al. 1999; Woo and Zedler 2002; Wilcox et al. 2008), but up to this point there has been little evidence of invasions occurring without either of these human disturbances. Our data suggest that even in sites with minimal recent human disturbance, when both of the parental species are present, naturally occurring reductions in water levels by themselves are sufficient to promote the spread of T. X glauca into intact Great Lakes coastal wetlands.
Are newly establish T. X glauca stands associated with changes in plant communities and soils similar to those that have been documented in more nutrient rich, human disturbed, and long-established stands?
We found that the intact wetlands of the northern Great Lakes that were recently invaded by T. X glauca exhibited environmental and plant community characteristics comparable to highly disturbed, cattail dominated wetlands in urbanized or agricultural watersheds in the southern Great Lakes, namely dense litter accumulation (Vaccaro et al. 2009), low species diversity (Frieswyk and Zedler 2007; Vaccaro et al. 2009), elevated soil nutrients (Galatowitsch et al. 1999; Farrer and Goldberg 2009), and thick organic soils (Tuchman et al. 2009). These data support the hypothesis that T. X glauca not only responds to human degradation of environmental conditions, but may also be responsible for the alteration or degradation of ecological conditions (Farrer and Goldberg 2009; Tuchman et al. 2009).
Did plant communities change between 1988-high water and 2008-low water?
In Great Lakes coastal wetlands, fluctuating water levels drive changes in plant community composition (Wilcox and Nichols 2008). Under conditions of decreasing water levels, species richness is expected to increase due to regeneration of species from the soil seed bank (Keddy and Reznicek 1986). As expected, dominant species composition differed between sampling years in our study. However, wetland-scale plant communities were more homogenous and species richness did not change significantly between study dates despite an increase in emergent species such as Juncus spp., which rapidly colonize newly exposed mud flats on the open lake fringe during low water conditions (Albert unpublished data; Table 2). These unexpected results may be explained in part, by localized T. X glauca dominance, and increased woody plant (Alnus incana, Salix spp., and Thuja occidentalis) occurrence and dominance, which reduced the extent and diversity of the wet meadow community.
Typha X glauca affected plant communities at the plot scale, clearly reducing plant species diversity and altering community composition and was associated with greater community homogenization at the wetland scale. Because T. X glauca is highly competitive (Galatowich 1999) and tolerates variable water levels (Waters and Shay 1990), we expect the newly established T. X glauca stands (since 1988-high water) will maintain or increase their coverage with naturally fluctuating water levels, resulting in reduced plant community variation and decreased wetland-scale species richness over the long-term.
Implications
Human-initiated environmental changes, including increased nutrient levels, altered hydrology, and exposed mineral and organic sediments (seed beds), have been demonstrated to be responsible for the establishment of invasive wetland plants (Woo and Zedler 2002; Kercher and Zedler 2004; Herrick and Wolf 2005; Boers et al. 2007; Boers and Zedler 2008). However, other recent studies have alleged that invasive plants are responsible for altering ecosystem structure and function without previous human-initiated environmental degradation (e.g., Ehrenfeld 2003; Farrer and Goldberg 2009). Our study demonstrates that in ecosystems with no or little recent anthropogenic alteration, natural variation in environmental conditions alone can create circumstances appropriate for invasive plant species to establish. In such a case, invasive species establishment alone could be linked to changes in ecosystem structure and function, even where there was little direct human disturbance.
Climate change models consistently predict decreases in Lake Michigan and Lake Huron water levels over the next 50–100 years (Kling et al. 2003; Angel and Kunkel 2010). If these predictions hold true, environmental conditions which facilitate the spread of T. X glauca will likely be enhanced, resulting in further widespread establishment and spread of T. X glauca, concomitant plant community homogenization, long-term loss of plant diversity, and compromised ecosystem function (Farrer and Goldberg 2009; Tuchman et al. 2009), even in systems sheltered from direct human disturbance.
References
Albert DA (2008) Vegetation community indicators. In Burton TM, Brazner JC, Ciborowksi JJH, Grabas GP, Hummer J, Schneider J, Uzarski DG (eds) Great Lakes coastal wetlands monitoring plan. Great Lakes Coastal Wetlands Consortium
Albert D, Minc L (2004) Plants as regional indicators of Great Lakes coastal wetland health. Aquatic Ecosystem Health & Management 7:233–247
Albert DA, Crispin SR, Reese G, Wilsmann LA, Ouwinga SJ (1987) A survey of great lakes marshes in Michigan’s Upper Peninsula. Michigan Natural Features Inventory, Lansing, Technical Report 1987–02
Albert DA, Reese G, Penskar MR, Wilsmann LA, Ouwinga SJ (1989) A survey of great lakes marshes in the northern half of Michigan’s Lower Peninsula and throughout Michigan’s Upper Peninsula. Michigan Natural Features Inventory, Lansing, Technical Report 1989–01
Albert DA, Wilcox DA, Ingram JW, Thompson TA (2005) Hydrogeomorphic classification for Great Lakes coastal wetlands. Journal of Great Lakes Research 31:129–146
Angel JR, Kunkel KE (2010) The response of Great Lakes water levels to future climate scenarios with an emphasis on Lake Michigan-Huron. Journal of Great Lakes Research 36:51–58
Angeloni NL, Jankowski KJ, Tuchman NC, Kelly JJ (2006) Effects of an invasive cattail species (Typha x glauca) on sediment nitrogen and microbial community composition in a freshwater wetland. FEMS Microbiology Letters 263:86–92
APHA (2005) Standard methods for the examination of water and wastewater, 21st edn. American Public Health Association, Washington, DC
Boers A (2005) The effects of stabilized water levels on invasion by hybrid cattail (Typha x glauca). PhD Dissertation, University of Wisconsin-Madison
Boers AM, Zedler JB (2008) Stabilized water levels and Typha invasiveness. Wetlands 28:676–685
Boers AM, Veltman RLD, Zedler JB (2007) Typha x glauca dominance and extended hydroperiod constrain restoration of wetland diversity. Ecological Engineering 29:232–244
Davis C, Van der Valk A (1978) The decomposition of standing and fallen litter of Typha glauca and Scirpus fluviatilis. Canadian Journal of Botany 56:662–675
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503–523
Farrer E, Goldberg D (2009) Litter drives ecosystem and plant community changes in cattail invasion. Ecological Applications 19:398–412
Freyman M (2008) The effect of litter accumulation of the invasive cattail Typha x glauca on a great lakes coastal marsh. Masters Thesis, Loyola University Chicago
Frieswyk CB, Zedler JB (2007) Vegetation change in great lakes coastal wetlands: deviation from the historical cycle. Journal of Great Lakes Research 33:366–380
Galatowitsch SM, Anderson NO, Ascher PD (1999) Invasiveness in wetland plants in temperate North America. Wetlands 19:733–755
Gardner W (1986) Water content. In: Klute A (ed) Methods of soil analysis. Soil Science Society of America, Madison, pp 493–544
Gathman JP, Albert DA, Burton TM (2005) Rapid plant community response to a water level peak in northern Lake Huron coastal wetlands. Journal of Great Lakes Research 31:160–170
Harris S, Marshall W (1963) Ecology of water-level manipulations on a northern marsh. Ecology 44:331–343
Herman K, Masters L, Penskar MR, Reznicek AA, Wilhelm G, Brodowicz W, Gardiner K (2001) Floristic quality assessment with wetland categories and examples of computer applications for the state of Michigan. Michigan Department of Natural Resources, Wildlife Division, Natural Heritage Program, Lansing, Report 2001–17
Herrick B, Wolf A (2005) Invasive plant species in diked vs. undiked Great Lakes wetlands. Journal of Great Lakes Research 31:277–287
Holeck K, Mills E, MacIsaac H, Dochoda M, Colautti R, Ricciardi A (2004) Bridging troubled waters: biological invasions, transoceanic shipping, and the Laurentian Great Lakes. Bioscience 54:919–929
Keddy PA, Reznicek AA (1986) Great Lakes vegetation dynamics: the role of fluctuating water levels and buried seeds. Journal of Great Lakes Research 12:25–36
Kercher SM, Zedler JB (2004) Multiple disturbances accelerate invasion of reed canary grass (Phalaris arundinacea L.) in a mesocosm study. Oecologia 138:455–464
Kling GW, Hayhoe K, Johnson LB, Magnuson JJ, Polasky S, Robinson SK, Shuter BJ, Wander MM, Wuebbles DJ, Zak DR, Lindroth RL, Moser SC, Wilson ML (2003) Confronting climate change in the Great Lakes region: impacts on our communities and ecosystems. Union of Concerned Scientists, Cambridge Massachusetts, and Ecological Society of America, Washington, DC
Kost M, Albert D, Cohen J, Slaughter B, Schillo R, Weber C, Chapman K (2007) Natural communities of Michigan: classification and description. Michigan Natural Features Inventory, Lansing, Report 2007–21
Lenters J (2001) Long-term trends in the seasonal cycle of Great Lakes water levels. Journal of Great Lakes Research 27:342–353
Lofgren B, Quinn F, Clites A, Assel R, Eberhardt A, Luukkonen C (2002) Evaluation of potential impacts on Great Lakes water resources based on climate scenarios of two GCMs. Journal of Great Lakes Research 28:537–554
Magnuson J, Webster K, Assel R, Bowser C, Dillon P, Eaton J, Evans H, Fee E, Hall R, Mortsch L (1997) Potential effects of climate changes on aquatic systems: Laurentian Great Lakes and Precambrian shield region. Hydrological processes 11:825–871
Maynard L, Wilcox D (1997) Coastal wetlands of the Great Lakes: background paper for the State of the Lake conference. Environment Canada; US Environmental Protection Agency, Chicago, IL, USA, and Toronto Canada. EPA 905-D-96-001c
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
McDonald ME (1955) Cause and effects of a die-off of emergent vegetation. Journal of Wildlife Management 19:24–35
Mills EL, Leach JH, Carlton JT, Secor CL (1993) Exotic species in the Great-Lakes—a history of biotic crises and anthropogenic introductions. Journal of Great Lakes Research 19:1–54
Minc L (1997) Great Lakes coastal wetlands: an overview of controlling abiotic factors, regional distribution, and species composition. Michigan Natural Features Inventory, Lansing, Report 1997–12
Mortsch L (1998) Assessing the impact of climate change on the Great Lakes shoreline wetlands. Climatic Change 40:391–416
Mortsch L, Quinn F (1996) Climate change scenarios for Great Lakes Basin ecosystem studies. Limnology and Oceanography 41:903–911
NAIP (2007) Aerial photographs of Northern Michigan. In: National Agriculture Imagery Program. United States Geological Survey. Available via web. http://seamless.usgs.gov/index.php. Accessed 5 May 2008
Rahel F, Olden J (2008) Assessing the effects of climate change on aquatic invasive species. Conservation Biology 22:521–533
R Development Core Team (2008) R: a language and environment for statistical computing. In Journal of Computational and Graphical Statistics. R Foundation for Statistical Computing, Vienna, Austria
Ricciardi A (2001) Facilitative interactions among aquatic invaders: is an “invasional meltdown” occurring in the Great Lakes? Canadian Journal of Fisheries and Aquatic Sciences 58:2513–2525
Shay J, Shay C (1986) Prairie marshes in western Canada, with specific reference to the ecology of five emergent macrophytes. Canadian Journal of Botany 64:443–454
Smith S (1967) Experimental and natural hybrids in North American Typha (Typhaceae). American Midland Naturalist 78:257–287
Smith S (1987) Typha: its taxonomy and the ecological significance of hybrids. Archiv für Hydrobiologie 27:129–138
Sousounis P, Grover E (2002) Potential future weather patterns over the Great Lakes region. Journal of Great Lakes Research 28:496–520
Trebitz AS (2006) Characterizing seiche and tide-driven daily water level fluctuations affecting coastal ecosystems of the Great Lakes. Journal of Great Lakes Research 32:102–116
Trebitz AS, Taylor DL (2007) Exotic and invasive aquatic plants in great lakes coastal wetlands: distribution and relation to watershed land use and plant richness and cover. Journal of Great Lakes Research 33:705–721
Tuchman NC, Jankowski KJ, Geddes P, Wildova R, Larkin D, Goldberg D (2009) Patterns of environmental change associates with Typha X glauca invasion in a great lakes coastal wetland. Wetlands 29:964–975
Tulbure MG, Johnston CA, Auger DL (2007) Rapid invasion of a Great Lakes coastal wetland by non-native Phragmites australis and Typha. Journal of Great Lakes Research 33:269–279
USACE (2009) Historic great lakes water levels. In: United States Army Corps of Engineers, Detroit, MI. Available via web. http://www.lre.usace.army.mil/greatlakes/hh/greatlakeswaterlevels/historicdata/. Accessed 2 Feb 2009
Vaccaro L, Bedford B, Johnston C (2009) Litter accumulation promotes dominance of invasive species of cattails (Typha spp.) in Lake Ontario wetlands. Wetlands 29:1036–1048
Vail L (2009) Soil nutrient changes following a Typha x glauca invasion in a Great Lakes coastal wetland. Masters Thesis, Loyola University Chicago
Voss E (1972) Michigan flora: a guide to the identification and occurrence of the native and naturalized seed-plants of the state: Part 1. Gymnosperms and monocots. Cranbrook Institute of Science, Bloomfield Hills
Voss E (1985) Michigan flora: a guide to the identification and occurrence of the native and naturalized seed-plants of the state. Part 2. Dicots (Saururaceae-Cornaceae). Cranbrook Institute of Science, Bloomfield Hills
Voss E (1996) Michigan flora: a guide to the identification and occurrence of the native and naturalized seed-plants of the state. Part 3, Dicots (Pyrolaceae-Compositae). Cranbrook Institute of Science, Bloomfield Hills
Waters I, Shay J (1990) A field study of the morphometric response of Typha glauca shoots to a water depth gradient. Canadian Journal of Botany 68:2339–2343
Whyte R, Trexel-Kroll D, Klarer D, Shields R, Francko D (2008) The invasion and spread of Phragmites australis during a period of low water in a Lake Erie coastal wetland. Journal of Coastal Research 55:111–120
Wilcox D (2004) Implications of hydrologic variability on the succession of plants in Great Lakes wetlands. Aquatic Ecosystem Health & Management 7:223–231
Wilcox D, Meeker J (1991) Disturbance effects on aquatic vegetation in regulated and unregulated lakes in northern Minnesota. Canadian Journal of Botany 69:1542–1551
Wilcox D, Nichols J (2008) The effects of water-level fluctuations on vegetation in a Lake Huron wetland. Wetlands 28:487–501
Wilcox DA, Xie YC (2007) Predicting wetland plant community responses to proposed water-level-regulation plans for Lake Ontario: GIS-based modeling. Journal of Great Lakes Research 33:751–773
Wilcox DA, Kowalski KP, Hoare HL, Carlson ML, Morgan HN (2008) Cattail invasion of sedge/grass meadows in Lake Ontario: photointerpretation analysis of sixteen wetlands over five decades. Journal of Great Lakes Research 34:301–323
Woo I, Zedler JB (2002) Can nutrients alone shift a sedge meadow towards dominance by the invasive Typha x glauca? Wetlands 22:509–521
Acknowledgements
This research was supported by an award from the Loyola University Chicago Office of the Provost to N. Tuchman. We also thank the University of Michigan Biological Station for lab facilities. Particular thanks are due to K. Koch for assistance in the field; M. Grant for laboratory analytical services; D. Larkin for field and statistical assistance; and M. Mitchell, C. Smith, and D. Miceli for assistance in the field.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
(PDF 48 kb)
Rights and permissions
About this article
Cite this article
Lishawa, S.C., Albert, D.A. & Tuchman, N.C. Water Level Decline Promotes Typha X glauca Establishment and Vegetation Change in Great Lakes Coastal Wetlands. Wetlands 30, 1085–1096 (2010). https://doi.org/10.1007/s13157-010-0113-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-010-0113-z