
Abstract
Concerned in this paper is a discrete predator-prey system with Allee effect and other food resources for the predators. The conditions on the existence and stability of fixed points are obtained. It is shown that the system can undergo fold bifurcation and flip bifurcation by using the center manifold theorem and bifurcation theory. Numerical simulations are provided to illustrate the feasibility of the main results and the influence of Allee effect on the stability of the system. Our study indicates that other food resources for the predator can enrich the dynamical behaviours of the system, including cascades of period-doubling bifurcation in orbits of period-2, 4, 8, and chaotic sets.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The predation relationship between predator and prey is one of the dominant themes in ecology, due to its universal existence and importance. Predator-prey models have been extensively studied with the first one being proposed by Lotka and Volterra. A general Lotka-Volterra predator-prey model can be written as
where \(b_1\) and \(a_{ij}\)’s are positive constants, \(b_2>0\) means that the predator has other food resources and \(b_2<0\) otherwise. According to Ma [1], a positive fixed point must be globally stable when exists.
Allee effect [2] is a crucial phenomenon that has been studied by many scholars [3,4,5,6,7,8,9,10]. It can be regarded as a negative density dependence of the per capita growth rate of a population when its density is smaller than a critical value. It may be caused by many factors including the difficulty in finding a mate, reduced defence against predators at low densities, special trends of social dysfunction, etc. Allee effect may enhance [11, 12] or decrease [13,14,15,16,17,18,19,20,21] the stability of the system. By the biological meanings, the Allee function f(u, x) should satisfy the following requirements:
where u is the Allee constant, x is the population density. Noting that \(f(u,x)=\frac{x}{u+x}\) meets the above requirements.
For a bio-mathematical model, when species have non-overlapping generations or the population densities are too small, discrete models described by difference equations are more realistic than the continuous–time models. The dynamic behaviours of discrete predator-prey systems have been extensively studied over the past decades. To name a few, see [22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37] and references therein. All these works have demonstrated that discrete systems indeed have more complex dynamic behaviours than the continuous ones. In particular, several discrete models with Allee effect are discussed in [11, 12, 38,39,40,41]. Among these investigations, Celik and Duman [11] studied the following discrete predator-prey system with the prey population subject to an Allee effect,
They showed that the Allee effect has a stabilizing effect for system (2) and the positive fixed point arrives stability much faster due to the Allee effect.
The models mentioned above are all based on a one-to-one relationship between the predator and the prey, i.e., the predator species takes the prey species as its unique food resource. Thus the extinction of the prey species will lead to the extinction of the predator species. However, predators are generally polyphagies and do not hunt for only one type of prey. Based on this, Zhu et al. [42] and Chen et al. [36] proposed respectively continuous and discrete models where the predator species has other food resources and the prey species is subject to fear effect. So far, little has been done for discrete predator-prey systems with both Allee effect and other food resources for predators.
Motivated by the above discussion and works, we propose a continuous predator-prey system with Allee effect and other food resources for the predator and study the dynamical behaviours of its discrete version obtained by the forward Euler scheme. The continuous model is
where x and y are the densities of the prey and the predator at time t, respectively. Here r, K, \(u_1\), a, p, e, and h are all positive constants. r and e represent the intrinsic growth rates of the prey and predator, respectively; K is the carrying capacity of the prey in the absence of the predator; a denotes the maximum predation rate of the predator and \(\frac{p}{a}\) stands for the conversion rate of prey’s biomass to predator’s biomass; \(u_1\) is the Allee effect constant of the prey; and h describes the death rate due to intra-species competition of the predator.
For the sake of simplicity, we make the following change of variables,
Denote
After dropping the bars, system (3) becomes
Applying the forward Euler scheme to system (4) and taking the step size \(\delta \rightarrow 1\), we obtain the following discrete system,
The aim of this paper is to study the dynamical behaviours of system (5), which include the existence and stability of fixed points, and the bifurcation phenomena.
The rest of this paper is arranged as follows. Sect. 2 is devoted to the existence and stability of fixed points. Then, in Sect. 3, we show that system (5) can undergo fold bifurcation and flip bifurcation under appropriate conditions on the parameters. Numerical simulations are provided in Sect. 4 to illustrate the feasibility of the main results. The paper ends with a brief conclusion.
2 The existence and stability of fixed points
2.1 The existence of fixed points
The fixed points of system (5) satisfy the following equations,
Obviously, system (5) always admits the boundary fixed points \(E_0(0,0)\), \(E_1(0,m)\), and \(E_2(1,0)\). For the positive fixed points, we only need to consider positive solutions of the following equations,
For positive fixed points, x must satisfy \(0<x<1\). Let \(\Delta \) denote the discriminant of the first equation of (6) and express \(\Delta \) in terms of m, i.e.,
Then \(\Delta (m)\) has two roots,
Note that \(0\le m^*<m^{**}\).
Theorem 1
The following statements on positive fixed points of system (5) hold.
-
1.
If either \(m>m^*\) or \(bcu\ge 1\), then there is no positive fixed point.
-
2.
If \(m=m^*\) and \(bcu<1\), then there is a unique positive fixed point \(E_{31}(x_{31},y_{31})\), where \(x_{31}=\sqrt{\frac{u(u+1)}{bc+1}}-u\) and \(y_{31}=cx_{31}+m\).
-
3.
If \(0<m<m^*\) and \(bcu<1\), then there are two distinct positive fixed points \(E_{32}(x_{32},y_{32})\) and \(E_{33}(x_{33},y_{33})\), where \(x_{32}=\frac{1-bcu-bm-\sqrt{\Delta (m)}}{2(bc+1)}\), \(x_{33}=\frac{1-bcu-bm+\sqrt{\Delta (m)}}{2(bc+1)}\), \(y_{32}=cx_{32}+m\), and \(y_{33}=cx_{33}+m\).
Proof
Let \(f(x)=(bc+1)x^2+(bcu+bm-1)x+bmu\). We only need to show when f has positive zeros in (0, 1). Note that \(f'(x)=2(bc+1)x+bcu+bm-1\). It follows that \(f'({\bar{x}})=0\) with \({\bar{x}}=\frac{1-bm-bcu}{2(bc+1)}\). If \(bcu\ge 1\), then \({\bar{x}} {\le }-\frac{bm}{2(bc+1)}<0\). This, combined with \(f(0)=bmu>0\), implies that f(x) has no positive zeros when \(bcu\ge 1\). So in the following, we assume that \(bcu< 1\), which implies that \(m^*\ne 0\).
If \(m^*<m<m^{**}\), then \(\Delta (m)<0\), which means that f(x) has no real zeros.
If \(m\ge m^{**}\), then \({\bar{x}} =\frac{1-bm-bcu}{2(bc+1)}\le \frac{1-bm^{**}-bcu}{2(bc+1)} =\frac{-2u(bc+1)-2\sqrt{u(bc+1)(u+1)}}{2(bc+1)}<0\). It follows from the argument at the beginning of the proof that f(x) has no positive zeros.
If \(m=m^{*}\), then \(\Delta (m)=0\). In addition, \( {\bar{x}}=\frac{1-bm^*-bcu}{2(bc+1)}=\sqrt{\frac{u(u+1)}{bc+1}}-u >0\) since \(bcu<1\). It is easy to see that \({\bar{x}}<1\). Therefore, \({\bar{x}}\) is the only positive real zero of f. It follows that system (5) has a unique positive fixed point \(E_{31}(x_{31},y_{31})\) when \(m=m^*\) and \(bcu<1\), where \(x_{31}={\bar{x}}\) and \(y_{31}=cx_{31}+m\).
If \(0<m<m^{*}\), we have \(\Delta (m)>0\). It follows from \(bcu<1\) that \({\bar{x}}>\frac{1-bm^*-bcu}{2(bc+1)}>0\). Note that \(f(0)>0\), \(f(1)=b(1+m)(1+u)>0\), and \(f'(1)=1+2bc+bcu+bm>0\). Thus \(f(x)=0\) has two distinct positive roots \(x_{32}=\frac{1-bcu-bm-\sqrt{\Delta (m)}}{2(bc+1)}\) and \(x_{33}=\frac{1-bcu-bm+\sqrt{\Delta (m)}}{2(bc+1)}\), both in (0, 1). This shows that system (5) has two distinct positive fixed points \(E_{32}(x_{32},y_{32})\) and \(E_{33}(x_{33},y_{33})\), where \(y_{32}=cx_{32}+m\) and \(y_{33}=cx_{33}+m\). \(\square \)
2.2 The stability of fixed points
In this subsection, we use linearization to discuss the stability of the fixed points obtained in the previous subsection.
The Jacobian matrix of system (5) evaluated at a fixed point E(x, y) is given by
where
Write the characteristic equation of J(E) as \(F(\lambda )=\lambda ^2+B\lambda +C=0\). Assume that \(\lambda _1\) and \(\lambda _2\) are the two roots of \(F(\lambda )=0\). Then E is classified as follows.
Definition 1
The fixed point E of (5) is
-
1.
locally asymptotically stable if \(\max \{\vert \lambda _1\vert , \vert \lambda _2\vert \}<1\) and it is called a sink;
-
2.
unstable if \({\max }\{\vert \lambda _1\vert , \vert \lambda _2\vert \}>1\);
-
3.
non-hyperbolic if either \(\vert \lambda _1\vert =1\) or \(\vert \lambda _2\vert =1\).
The following results tell us how to determine the type of the fixed point E.
Lemma 1
([43, Lemma 2]) Assume that \(F(1)>0\). Then
-
1.
\(|\lambda _1|<1\) and \(|\lambda _2|<1\) if and only if \(F(-1)>0\) and \(C<1\);
-
2.
\(|\lambda _1|>1\) and \(|\lambda _2|>1\) if and only if \(F(-1)>0\) and \(C>1\);
-
3.
(\(|\lambda _1|>1\) and \(|\lambda _2|<1\)) or (\(|\lambda _1|<1\) and \(|\lambda _2|>1\)) if and only if \(F(-1)<0\);
-
4.
\(\lambda _1=-1\) and \(|\lambda _2|\ne 1\) if and only if \(F(-1)=0\) and \(B\ne 0,2\);
-
5.
\(\lambda _1\) and \(\lambda _2\) are conjugate complex roots and \(|\lambda _1|=|\lambda _2|=1\) if and only if \(B^2-4C<0\) and \(C=1\).
Note that \(F(1)>0\) and \(F(-1)=0\) imply that \(B\ne 0\). Hence \(B\ne 0\) is redundant in (iv) of Lemma 1, which will be ignored in the coming discussion.
The following result can be proved in the same manner as Lemma 1 and hence the detail is omitted here.
Lemma 2
Assume that \(F(1)<0\). Then
-
1.
\(|\lambda _1|>1\) and \(|\lambda _2|>1\) if and only if \(F(-1)<0\);
-
2.
(\(|\lambda _1|>1\) and \(|\lambda _2|<1\)) or (\(|\lambda _1|<1\) and \(|\lambda _2|>1\)) if and only if \(F(-1)>0\);
-
3.
\(\lambda _1=-1\) and \(|\lambda _2|\ne 1\) if and only if \(F(-1)=0\).
For the boundary fixed points \(E_0(0,0)\), \(E_1(0,m)\), and \(E_2(1,0)\), we have \( J(E_0)= \begin{pmatrix} 1 &{}\quad 0 \\ 0 &{}\quad 1+m \end{pmatrix}\), \(J(E_1)= \begin{pmatrix} 1-bm &{}\quad 0 \\ cm &{}\quad 1-m \end{pmatrix}\), and \(J(E_2)= \begin{pmatrix} \frac{u}{u+1} &{}\quad -b\\ 0 &{}\quad 1+c+m \end{pmatrix}\), respectively. Then we can easily get their stability, which is summarized below.
Theorem 2
For the three boundary fixed points \(E_0\), \(E_1\), and \(E_2\) of system (5),
-
1.
\(E_0(0,0)\) is always non-hyperbolic;
-
2.
\(E_1(0,m)\) is
-
(a)
stable if \(m<\min \{\frac{2}{b},2\}\);
-
(b)
non-hyperbolic if \(m=\frac{2}{b}\) or \(m=2\);
-
(c)
unstable for the other cases;
-
(a)
-
3.
\(E_2(1,0)\) is always unstable.
Now, we turn to the positive fixed points of (5). Recall that the positive fixed points \(E_{3i}\) (\(i=1\), 2, 3) satisfy
Substitute (8) into (7) to simplify the Jacobian matrix evaluated at \(E_{3i}\) as
where \(\alpha _i=\frac{u(1-x_{3i})}{(u+x_{3i})^2}-\frac{x_{3i}}{u+x_{3i}}\). Thus the characteristic equation of \(J(E_{3i})\) is
where \(P=-2-\alpha _i x_{3i}+y_{3i}\) and \(Q=1+\alpha _i x_{3i}-y_{3i}+\left( bc-\alpha _i\right) x_{3i}y_{3i}\). In particular,
Theorem 3
Under the conditions on the existence of positive fixed points of (5) in Theorem 1,
-
1.
\(E_{31}\) is always non-hyperbolic;
-
2.
\(E_{32}\) is
-
(a)
non-hyperbolic if \((bc-\alpha _2)cx_{32}^2+2(\alpha _2-c)x_{32}+4>0\) and
$$\begin{aligned} m=\frac{(bc-\alpha _2)cx_{32}^2 +2(\alpha _2-c)x_{32}+4}{2-(bc-\alpha _2)x_{32}}; \end{aligned}$$ -
(b)
unstable for the other cases;
-
(a)
-
3.
The properties of \(E_{33}\) are listed in Table 1.
Proof
Note that \(H(x_{3i})=\frac{h(x_{3i})}{(u+x_{3i})^2}\), where
Then the sign of F(1) is determined by that of \(h(x_{3i})\).
-
(i)
At \(E_{31}\), \(x_{31}=\sqrt{\frac{u(u+1)}{bc+1}}-u\). A simple calculation gives \(h(x_{31})=0\) and hence \(F(1)=0\). Therefore, \(E_{31}\) is always non-hyperbolic.
Recall from Theorem 1 that one of the conditions on the existence of \(E_{32}\) and \(E_{33}\) is \(bcu<1\). Then \(h(0)=bcu^2-u<0\). Since the vertex of h(x) is at the left of the y-axis, h(x) is monotonically increasing for \(x>0\). Moreover, since \(0<m<m^*\), it follows from
$$\begin{aligned} f(x_{31})= & {} (bc+1)x^2_{31}+(bcu+bm-1)x_{31}+bmu \\= & {} [(bc+1)x^2_{31}+(bcu+bm^*-1)x_{31}+bm^*u]+b(m-m^*)(x_{31}+u) \\< & {} 0 \end{aligned}$$that \(x_{32}<x_{31}<x_{33}\).
-
(ii)
For \(E_{32}\), by the above discussion, \(h(x_{32})<h(x_{31})=0\) and hence \(F(1)=(bc-\alpha _2)x_{32}y_{32}<0\), which implies that \(bc-\alpha _2<0\). By Lemma 2, if \(F(-1)=0\) then \(E_{32}\) is non-hyperbolic and otherwise it is unstable. Noting
$$\begin{aligned} F(-1)=(bc-\alpha _2)x_{32}^2+2(\alpha _2-c)x_{32}+4-m[2-(bc-\alpha _2)x_{32}], \end{aligned}$$we easily see that \(F(-1)=0\) if and only if \((bc-\alpha _2)x_{32}^2+2(\alpha _2-c)x_{32}+4>0\) and \(m=\frac{(bc-\alpha _2)x_{32}^2+2(\alpha _2-c)x_{32}+4}{2-(bc-\alpha _2)x_{32}}\). Then 2(a) and 2(b) follow immediately.
-
(iii)
For \(E_{33}\), we have \(h(x_{33})>h(x_{31})=0\) and hence \(F(1)>0\). Express \(F(-1)\) and \(Q-1\) as
$$\begin{aligned} F(-1)= & {} \left[ (bc-\alpha _3)x_{33}-2\right] m +(bc-\alpha _3)cx_{33}^2+2(\alpha _3-c)x_{33}+4 \\\triangleq & {} \left[ (bc-\alpha _3)x_{33}-2\right] m+{\widetilde{m}} \end{aligned}$$and
$$\begin{aligned} Q-1= & {} \left[ (bc-\alpha _3)x_{33}-1\right] m +(bc-\alpha _3)cx_{33}^2+(\alpha _3-c)x_{33} \\\triangleq & {} \left[ (bc-\alpha _3)x_{33}-1\right] m+{\overline{m}}, \end{aligned}$$respectively. Then the result on stability of \(E_{33}\) follows easily from Lemma 1. \(\square \)
3 Bifurcation analysis
In this section, we investigate the possible bifurcations occurring at the fixed points of system (5) by using the center manifold theorem [44] and bifurcation theory [45, 46]. We start with the fold bifurcation.
3.1 Fold bifurcation
Recall from Theorem 1(ii) that if
then system (5) has only one positive fixed point \(E_{31}(x_{31},y_{31})\) and the eigenvalues of the Jacobian matrix \(J(E_{31})\) are \(\lambda _1=1\) and \(\lambda _2=1+\alpha _1x_{31}-(cx_{31}+m_1)\). Suppose that
Then \(|\lambda _2 |\ne 1\).
Let \(w=x-x_{31}\), \(v=y-y_{31}\), and \(\eta =m-m_1\). Then system (5) can be rewritten as
where \(\beta =\frac{u^3+u^2}{(u+x_{31})^3}-1\). We choose
which is invertible. Then with the transformation
we transform (11) into
where
By the center manifold theory, in a small neighborhood of \(\eta _1=0\), there exists a center manifold \(W^c(0)\) of (12) at the fixed point \(({\tilde{x}},{\tilde{y}})=(0,0)\), which can be represented as
where \({\tilde{x}}\) and \(\eta _1\) are sufficiently small. Suppose that the expression of h is
which must satisfy
Substituting (13) into (14) and comparing the coefficients of the like terms \({\tilde{x}}^k\eta _1^l\), we get
Therefore, the map (12) restricted to the center manifold \(W^c(0)\) can be written as
where
Since \(F_1(0,0)=0\), \(\frac{\partial F_{1}}{\partial {\tilde{x}}}(0,0)=1\), \(\frac{\partial F_{1}}{\partial \eta _1}(0,0)=1\), and \(\frac{\partial ^2 F_{1}}{\partial {\tilde{x}}^2}(0,0)=2k_1\ne 0\), we obtain the following result.
Theorem 4
The system (5) undergoes a fold bifurcation at \(E_{31}\) if conditions (9) and (10) hold. Moreover, the fixed points \(E_{32}\) and \(E_{33}\) bifurcate from \(E_{31}\) for \(m<m_1\), coalesce at \(E_{31}\) for \(m=m_1\), and disappear for \(m>m_1\).
3.2 Flip bifurcation
Now we discuss the flip bifurcations of system (5).
System (5) can undergo flip bifurcation at the boundary fixed point \(E_1(0,m)\) when parameters vary in a small neighborhood of \(m=2\) or \(m=\frac{2}{b}\). Since a center manifold of system (5) at \(E_1\) is \(x=0\) and system (5) restricted to it is the logistic model,
Its nontrivial fixed point is \(y_1=m\). If \(g'(y_1)=1-m\ne 0\) when parameters vary in a small neighborhood of \(m=2\) or \(m=\frac{2}{b}\), then flip bifurcation can occur (see Fig. 2). In this case, the prey species becomes extinct and, by choosing m as the bifurcation parameter, the predator species undergoes the flip bifurcation to chaos due to the other food resources.
Since \(E_{32}\) is always unstable, with the biological significance in mind, in the following we focus on the flip bifurcation at \(E_{33}\). Here we again choose m as the bifurcation parameter.
Rewrite the conditions in rows 8 to 10 of Table 1 as the following three subsets,
We show that flip bifurcation may undergo when parameters vary in one of \(F_{A1}\), \(F_{A2}\) and \(F_{A3}\).
Take parameter values \((b, c, u, m_2)\) arbitrarily from \(F_{A1}\), \(F_{A2}\), or \(F_{A3}\). Then the eigenvalues of \(J(E_{33})\) are \(\lambda _1=-1\) and \(\lambda _2\ne \pm 1\). Let \(w=x-x_{33}\), \(v=y-y_{33}\), and \(\mu =m-m_2\). Then system (5) can be rewritten as
where \(\beta _1=\frac{u^3+u^2}{(u+x_{33})^3}-1\). We choose
which is invertible. With the transformation
the map (15) becomes
where
Now we determine the center manifold \(W^c(0)\) of (16) at the fixed point \((X,Y)=(0,0)\) in a small neighborhood of \(\mu _1=0\), which can be expressed as
for X and \(\mu _1\) sufficiently small. h must satisfy
We suppose that h has the form
Substituting (18) into (17) and comparing the corresponding coefficients of the like terms in the left-hand and right-hand sides of the resultant, we can obtain
Therefore, the restricted map of (16) on the center manifold \(W^c(0)\) is
where
In order for map (19) to undergo a flip bifurcation, we require that the two discriminatory quantities \(\gamma _1\) and \(\gamma _2\) are not zero, where
In summary, from the above discussion and theory in [45, 46], we have derived the following result.
Theorem 5
If \(\gamma _1\ne 0\) and \(\gamma _2\ne 0\), then system (5) undergoes a flip bifurcation at the fixed point \(E_{33}\) when the parameter m varies in a small neighborhood of \(m_2\). Moreover, if \(\gamma _2>0\) (resp., \(\gamma _2<0\)), then the period-2 orbits that bifurcate from \(E_{33}\) are stable (resp., unstable).
4 Numerical simulations
This section presents the bifurcation diagrams and phase portraits of system (5) to confirm the feasibility of the main results. Further, numerical simulations are provided to investigate the influence of Allee effect on the stability of system (5).
Example 1
(Fold bifurcation at the positive fixed point \(E_{31}\)) We choose m as the bifurcation parameter. With
one obtains the bifurcation value \(m_1\approx 0.759\) and system (5) only has one positive fixed point \(E_{31}(0.257,0.836)\). It is easy to verify (9) and (10). Further, the eigenvalues of \(J(E_{31})\) are \(\lambda _1=1\) and \(\lambda _2=0.203\). All the conditions of Theorem 4 hold and hence fold bifurcation occurs at \(E_{31}\). Fig. 1 agrees very well with Theorem 4. Moreover, we can see that \(E_{32}\) is unstable while \(E_{33}\) is stable when \(m<m_1\) and they disappear when \(m>m_1\).

Fold bifurcation diagram of system (5) in the mx-plane where parameter values are given in (20) with the initial value (0.5, 0.6). The dash curve corresponds to the unstable fixed point \(E_{32}\) and the solid curve corresponds to the stable fixed point \(E_{33}\). The fold bifurcation value is \(m_1\approx 0.759\)

Bifurcation diagram of the logistic model \(y\rightarrow (1+m)y-y^2\) with \(m\in [1,3]\)
Example 2
(Flip bifurcation at the boundary fixed point \(E_{1}\)) System (5) always has a boundary fixed point \(E_{1}(0,m)\). Take \(b=0.5\), \(c=0.3\), and \(u=0.2\). From sect 3.2, we know that flip bifurcation emerges from the fixed point \(E_1\) at \(m=2\) (see Fig. 2).
Example 3
(Flip bifurcation at the positive fixed point \(E_{33}\)) Now we choose
According to Theorem 1(3.), system (5) has two distinct positive fixed points \(E_{32}\) and \(E_{33}\) when \(m<m^*=5.238\). After some simple calculations, we can find that the flip bifurcation emerges from \(E_{33}\) at \(m\approx 1.96\). With \(m\in [0,3]\), the discriminatory quantities \(\gamma _1\ne 0\) and \(\gamma _2>0\), and \((b,c,u,m)=(0.08,0.05,0.2,1.96)\in F_{A1}\). Fig. 3 shows the feasibility of Theorem 5. It is easy to see that \(E_{33}\) is stable for \(m<1.96\). When m reaches 1.96, with the increase of m, two points with a period-2 cycle are bifurcated, and then points with period-4 and period-8 are bifurcated in sequence. Some phase portraits related to Fig. 3 are displayed in Fig. 4, which include orbits of periods 2, 4, and 8. When \(m=2.9\), we can see chaotic sets in Fig. 4f.


Phase portraits for various values of m corresponding to Fig. 3
Example 4
(Effect of Allee effect) We now investigate the influence of Allee effect on the stability of system (5) through numerical simulations. Take the parameter values as
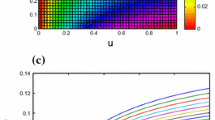
Figure 5 shows the graphs of the prey densities and the predator densities for various u. We considered the cases where system (5) has Allee effect (\(u\ne 0\)) and no Allee effect (\(u=0\)), and set up the control groups \(u=0.5\) and \(u=1\) as a way to see the effect of the magnitude of the Allee constant on properties of system (5). From Fig. 5, we observe that the Allee effect has little influence on the predator species while the local stability of the prey species decreases and its density arrives equilibrium value more slowly as the Allee constant u increases. What’s more, it is clear to see that the larger u is, the lower is the prey level at the fixed point.

Effect of Allee effect on the prey species and the predator species
5 Conclusion
In this paper, a discrete predator-prey system with Allee effect (on the prey species) and other food resources for predator has been proposed and studied. Conditions on the existence and the stability of fixed points are obtained. Moreover, taking the ratio of the intrinsic growth rates of predator to prey (m) as the bifurcation parameter, the model can undergo fold and flip bifurcations. For fold bifurcation, the number of the positive fixed points changes from two to one and eventually to 0 as m increases (see Fig. 1). According to theoretical analysis and Example 2, we obtain that flip bifurcation can occur at the boundary fixed point \(E_1(0,m)\), which means that the prey species is driven to extinct while the predator species first remains stable when m is small and gradually becomes chaotic as m increases through flip bifurcation (see Fig. 2). Further, it is shown that flip bifurcation will occur at the positive fixed point \(E_{33}\), which includes orbits of period-2, 4, 8 (see Fig. 3). This means that the positive fixed point \(E_{33}\) is stable if m is small, and when m is large enough, system (5) becomes unstable and even chaotic. Such a result is contrary to the conclusion drawn by Ma [1]. In other words, when the prey species is subject to Allee effect, a positive fixed point is likely to be unstable under certain conditions, rather than globally stable, that is, the Allee effect would decrease the stability of system (5), at least for the discrete systems.
In [11], Celik showed that Allee effect has a stabilizing force to system (2) and the fixed points reach stable steady state much faster when the prey species is subject to Allee effect. In this paper, however, we find that Allee effect reduces the population density of prey species at the stable steady state and it takes a longer time to reach the stable steady state when the Allee effect constant increases in the range of low values. Namely, the trajectories of system (5) take more time to arrive at the constant solution as the Allee effect increases in the range of low values, which is different from the results of Celik. Therefore, the discrete predator-prey system where the predator has other food resources presents more complex dynamical behaviors than systems in which the predator only takes the prey species as its unique food resource. When the Allee effect constant is large enough, the prey species becomes extinct because of low reproduction rate. Moreover, the larger the Allee effect constant is, the faster the prey species becomes extinct (see Fig. 5a). It is shown that Allee effect has little impact on predator species (stable steady state levels decrease by only 0.04) (see 5b).
In summary, our results show that the Allee effect and ratio of intrinsic growth rates m combined play an important role on the dynamic behaviors of the proposed model.
References
Ma, Z.: Mathematical Modelling and Study of Population Ecology. Anhui Education Press, (1996)
Dennis, B.: Allee effects: population growth, critical density, and the chance of extinction. Nat. Resour. Model. 3(4), 481–538 (1989)
Xiao, Z., Li, Z.: Stability and bifurcation in a stage-structured predator-prey model with Allee effect and time delay. Int. J. Appl. Math. 49(1), 6–13 (2019)
Zhu, Z., He, M., Li, Z., Chen, F.: Stability and bifurcation in a Logistic model with Allee effect and feedback control. Int. J. Bifurcation and Chaos 30(15), 2050231 (2020)
Lv, Y., Chen, L., Chen, F., Li, Z.: Stability and bifurcation in an SI epidemic model with additive Allee effect and time delay. Int. J. Bifurcation and Chaos 31(04), 2150060 (2021)
Chen, F., Guan, X., Huang, X., Deng, H.: Dynamic behaviors of a Lotka-Volterra type predator-prey system with Allee effect on the predator species and density dependent birth rate on the prey species. Open Math. 17(1), 1186–1202 (2019)
Rebelo, C., Soresina, C.: Coexistence in seasonally varying predator-prey systems with Allee effect. Nonlinear Anal. Real World Appl. 55, 103140 (2020)
Lai, L., Zhu, Z., Chen, F.: Stability and bifurcation in a predator-prey model with the additive Allee effect and the fear effect. Mathematics 8(8), 1280 (2020)
Lin, Q.: Allee effect increasing the final density of the species subject to the Allee effect in a Lotka-Volterra commensal symbiosis model. Adv. Diff. Equ. 2018(1), 1–9 (2018)
Lei, C.: Dynamic behaviors of a Holling type commensal symbiosis model with the first species subject to Allee effect. Commun. Math. Biol. Neurosci. 2019, 3 (2019)
Celik, C., Duman, O.: Allee effect in a discrete-time predator-prey system. Chaos Solitons Fractals 40(4), 1956–1962 (2009)
Chen, X., Fu, X., Jing, Z.: Dynamics in a discrete-time predator-prey system with Allee effect. Acta Math. Appl. Sin. Engl. Ser. 29(1), 143–164 (2013)
Ye, Y., Liu, H., Wei, Y., Ma, M., Zhang, K.: Dynamic study of a predator-prey model with weak Allee effect and delay. Adv. Math. Phys. 2019, 7296461 (2019)
Huang, Y., Zhu, Z., Li, Z.: Modeling the Allee effect and fear effect in predator-prey system incorporating a prey refuge. Adv. Diff. Equ. 2020(1), 1–13 (2020)
Guan, X., Chen, F.: Dynamical analysis of a two species amensalism model with Beddington-Deangelis functional response and Allee effect on the second species. Nonlinear Anal. Real World Appl. 48, 71–93 (2019)
Song, D., Song, Y., Li, C.: Stability and turing patterns in a predator-prey model with hunting cooperation and Allee effect in prey population. Int. J. Bifurcation and Chaos 30(09), 2050137 (2020)
Wu, R., Li, L., Lin, Q.: A Holling type commensal symbiosis model involving Allee effect. Commun. Math. Biol. Neurosci. 2018, 6 (2018)
Lin, Q.: Stability analysis of a single species Logistic model with Allee effect and feedback control. Adv. Diff. Equ. 2018(1), 1–13 (2018)
Chen, B.: Dynamic behaviors of a commensal symbiosis model involving Allee effect and one party can not survive independently. Adv. Diff. Equ. 2018(1), 1–12 (2018)
Lv, Y., Chen, L., Chen, F.: Stability and bifurcation in a single species Logistic model with additive Allee effect and feedback control. Adv. Diff. Equ. 2020(1), 1–15 (2020)
Xiao, Z., Xie, X., Xue, Y.: Stability and bifurcation in a Holling type II predator-prey model with Allee effect and time delay. Adv. Diff. Equ. 2018(1), 1–21 (2018)
Liu, X., Xiao, D.: Bifurcations in a discrete time Lotka-Volterra predator-prey system. Discrete Contin. Dyn. Syst. Ser. B 6(3), 559 (2006)
Liu, X., Xiao, D.: Complex dynamic behaviors of a discrete-time predator-prey system. Chaos Solitons Fractals 32(1), 80–94 (2007)
He, Z., Lai, X.: Bifurcation and chaotic behavior of a discrete-time predator-prey system. Nonlinear Anal. Real World Appl. 12(1), 403–417 (2011)
Chen, B.: Global attractivity of a discrete competition model. Adv. Diff. Equ. 2016(1), 1–11 (2016)
Cheng, L., Cao, H.: Bifurcation analysis of a discrete-time ratio-dependent predator-prey model with Allee effect. Commun Nonlinear Sci Numer Simul 38, 288–302 (2016)
Cui, Q., Zhang, Q., Qiu, Z., Hu, Z.: Complex dynamics of a discrete-time predator-prey system with Holling IV functional response. Chaos Solitons Fractals 87, 158–171 (2016)
Huang, T., Zhang, H.: Bifurcation, chaos and pattern formation in a space-and time-discrete predator-prey system. Chaos Solitons Fractals 91, 92–107 (2016)
Salman, S., Yousef, A., Elsadany, A.: Stability, bifurcation analysis and chaos control of a discrete predator-prey system with square root functional response. Chaos Solitons Fractals 93, 20–31 (2016)
Banerjee, R., Das, P., Mukherjee, D.: Stability and permanence of a discrete-time two-prey one-predator system with Holling type-III functional response. Chaos Solitons Fractals 117, 240–248 (2018)
Huang, J., Liu, S., Ruan, S., Xiao, D.: Bifurcations in a discrete predator-prey model with nonmonotonic functional response. J. Math. Anal. Appl. 464(1), 201–230 (2018)
Zhao, J., Yan, Y.: Stability and bifurcation analysis of a discrete predator-prey system with modified Holling-Tanner functional response. Adv. Diff. Equ. 2018(1), 1–18 (2018)
Rana, S.S.: Bifurcations and chaos control in a discrete-time predator-prey system of Leslie type. J. Appl. Anal. Comput 9(1), 31–44 (2019)
Santra, P., Mahapatra, G., Phaijoo, G.: Bifurcation and chaos of a discrete predator-prey model with Crowley–Martin functional response incorporating proportional prey refuge. Math. Probl. Eng. 2020, 5309814 (2020)
Singh, A., Deolia, P.: Dynamical analysis and chaos control in discrete-time prey-predator model. Commun Nonlinear Sci Numer Simul 90, 105313 (2020)
Chen, J., He, X., Chen, F.: The influence of fear effect to a discrete-time predator-prey system with predator has other food resource. Mathematics 9(8), 865 (2021)
Singh, A., Malik, P.: Bifurcations in a modified Leslie-Gower predator-prey discrete model with Michaelis-Menten prey harvesting. J. Appl. Math. Comput 67, 1–32 (2021)
Wang, W., Zhang, Y., Liu, C.: Analysis of a discrete-time predator-prey system with Allee effect. Ecol. Complex. 8(1), 81–85 (2011)
Işık, S.: A study of stability and bifurcation analysis in discrete-time predator-prey system involving the Allee effect. Int. J. Biomath. 12(01), 1950011 (2019)
Cheng, L., Cao, H.: Bifurcation analysis of a discrete-time ratio-dependent predator-prey model with Allee effect. Commun. Nonlinear. Sci. 38, 288–302 (2016)
Pal, S., Sasmal, S.K., Pal, N.: Chaos control in a discrete-time predator-prey model with weak Allee effect. Int. J. Biomath. 11(07), 1850089 (2018)
Zhu, Z., Wu, R., Lai, L., Yu, X.: The influence of fear effect to the Lotka-Volterra predator-prey system with predator has other food resource. Adv. Diff. Equ. 2020(1), 1–13 (2020)
Chen, G., Teng, Z.: On the stability in a discrete two-species competition system. J. Appl. Math. Comput. 38(1), 25–39 (2012)
Liaw, D.: Application of center manifold reduction to nonlinear system stabilization. Appl. Math. Comput. 91(2–3), 243–258 (1998)
Wiggins, S.: Introduction to Applied Nonlinear Dynamical Systems and Chaos, vol. 2. Springer, NY (2003)
Robinson, C.: Dynamical Systems: Stability, Symbolic Dynamics, and Chaos. CRC press, (1998)
Acknowledgements
This work was supported partially by the Natural Science Foundation of Fujian Province (2020J01499).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chen, J., Chen, Y., Zhu, Z. et al. Stability and bifurcation of a discrete predator-prey system with Allee effect and other food resource for the predators. J. Appl. Math. Comput. 69, 529–548 (2023). https://doi.org/10.1007/s12190-022-01764-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12190-022-01764-5