
Abstract
The present study was conducted to test the efficacy of Serendipita indica in Brassica juncea against cadmium (Cd) stress. Cd is a trace element that enters into plants through contaminated soil. Serendipita indica is a fungal endophyte which colonizes and benefits exceptionally large group of plants. Roots of 3-days old B. juncea seedlings were inoculated with S. indica and raised in toxic Cd concentrations (10 µM, 30 µM and 50 µM Cd) till 12 days after inoculation in plant growth chamber with conditions of 16-h light/8-h dark photoperiod, temperature 25 °C (± 2 °C) and 80% relative humidity. The seedlings were tested for various morpho-physiological parameters like seed germination, plant biomass and %age heavy metal tolerance index. Also, various non-enzymatic antioxidants like vitamin A, vitamin C, phenols, flavonoids, total soluble sugars, reducing and non-reducing sugars were analyzed. Cd led to reduction in the % age germination of B. juncea seeds. Serendipita indica inoculation incremented the growth parameters like fresh weight, dry weight, root length, shoot length which were diminished by Cd stress. Enhancement of 20.66% in root length was observed in inoculated and Cd stressed seedlings. The content of phenols increased under unstressed conditions by 26.6% and by 55.29% under Cd stressed conditions in S. indica inoculated seedlings along with increasing contents of vitamin A and C. Overall, the content of carbohydrates was enhanced with S. indica inoculation. The study suggests that S. indica endophyte has great potential in boosting growth and physiological parameters which help in managing Cd stress in B. juncea.
Graphical Abstract

Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Plants develop mutualistic relationships with a wide array of microorganisms residing internally or externally in their vicinity [1]. Fungal root endophytes are microorganisms that harbor inside the plant roots without causing any apparent damage, thereby forming a symbiotic type of association [2, 3]. These symbiotic fungal root endophytes have gained significant attention because of their potential role in promoting the growth of the plants they reside in, along with inculcating stress tolerance against various environmental stressors [4,5,6]. Among the various known endophytes, S. indica (also known as Piriformopsora indica) has gained significant attention as it has emerged as an intriguing candidate to promote plant growth by enhancing nutrient acquisition, hormonal production, and shielding against environmental adversities [7, 8]. Serendipita indica has been a popular endophyte ever since its discovery in the rhizospheric zone of the woody shrubs Prosopis juliflora and Zizyphus nummularia in the Thar Desert of northwestern India [9]. Serendipita indica is characterized by producing pear-shaped chlamydospores and belongs to the order Sebacinales (Basidiomycota) [9]. It tends to produce axenic cultures and colonize a broad spectrum of plant species [7,8,9,10]. Owing to the remarkable versatility of S. indica, its symbiotic association with the roots of plants underscores its potential to help plants combat environmental stress conditions and fortify plants’ capacity to retaliate against stressors, thereby presenting itself as a potential sustainable candidate for agricultural productivity enhancement.
Plants in their natural environment are posed with numerous conditions of environmental stress and heavy metal stress is one of the major condition of stress that plant experiences [11]. Elevation in anthropogenic activities like mining, coal burning, use of chemical fertilizers, pesticides, paint industries, automobile industries, and many more has posed a significant threat to all living organisms by increasing the levels of environmental pollution by infiltrating the soil–water system with an immense amount of pollutants [11, 12]. Heavy metal pollution has become a pressing issue in the environmental and agricultural sciences, as it poses serious implications for plants because of their sessile nature [13]. Cadmium (Cd) is one such heavy metal that holds no biological significance and is known to cause toxicity symptoms even at minute concentrations like 0.001–0.1 mg/L [14]. Cd is a trace element with a high density that does not undergo decomposition and thus persists in the main sink of contamination, which is soil, thereby posing serious health implications to organisms at almost all trophic levels [15,16,17]. In plants, Cd toxicity disrupts the metabolic machinery of plants by impacting enzyme activities, photosynthesis, mineral nutrition, and redox homeostasis, leading to oxidative stress due to imbalance in reactive oxygen species (ROS), DNA damage, and finally impacting yield by reducing the overall growth parameters [11, 18, 19]. Therefore, Cd toxicity is a serious threat to plants as they cannot escape contaminated soils and have to develop a shielding tendency against such stressors, and this defensive property can be built with the help of root endophytes like S. indica.
Brassica juncea (Indian Mustard) is an essential oilseed crop belonging to Brassicaceae and is widely utilized as an as an agricultural crop across the globe. Although B. juncea, being a hyperaccumulator, has a high tolerance for Cd, the growth parameters decline significantly [20]. Therefore, there is a need to add to the shielding capacity of B. juncea against Cd stress. S. indica is known to shield a variety of plants against different heavy metal stressors like zinc (Zn), lead (Pb), Cd, nickel (Ni), arsenic (As), and copper (Cu) [21,22,23]. S. indica helps plants fight against HM stress by inhibiting the uptake of metals, effective sequestration, enhancing the efficiency of antioxidant defense system to overcome oxidative stress, improving accumulation of osmolytes, improving morpho-physiological characters, photosynthetic efficiency and strengthening the non-enzymatic antioxidant defense mechanism of plants thereby improving overall growth and metabolism [21,22,23,24,25]. A large amount of contaminants, like Cd, in the soil challenges food security by directly targeting the vitality of the crops. Also, the use of synthetic fertilizers and other yield-promoting chemicals will deteriorate soil health. Therefore, it has become a pressing issue that demands an urgent search for a sustainable solution. Therefore, it will be interesting to explore the role of S. indica as a root endophyte in B. juncea against Cd stress.
The study endeavors to present a sustainable solution to enhance B. juncea’s resilience against Cd toxicity, which is in coherence with the United Nations Sustainable Development Goals (SDGs), specifically SDG 2 (zero hunger) and SDG 3 (good health and well-being) by promoting food security through a sustainable approach like S. indica inoculation. Furthermore, it also relates to SDG 15 (Life on Land), which emphasizes protecting the ecosystem where soil and plants are integral parts. Therefore, utilizing S. indica can be beneficial in protecting plants from Cd toxicity and promoting their growth sustainably, thereby aligning with the objectives of the SDGs. The present study seeks to add to the existing reservoir of knowledge on the potential role of fungal root endophytes (S. indica) in developing resilience in B. juncea exposed to Cd toxicity and present a sustainable opportunity to shield crops against stress by employing root endophytes. The present research aims to test efficacy of S. indica inoculation in roots of B. juncea for improving various morpho-physiological parameters along with enhancing various non-enzymatic defense parameters like phenols, flavonoids, sugars and vitamin contents which prove to be beneficial in overcoming Cd stress conditions.
Materials and Methods
Culturing and Inoculation of Fungal Endophyte, Preparation of Plant Material
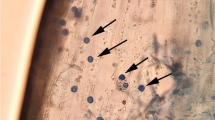
Culturing of S. indica was done using composition of Hill and Kafer medium and pH was set at 6.5 [26]. Liquid cultures used for inoculation were obtained at temperature conditions of 28 °C at 120 rpm for 3–5 days in a shaking incubator (Fig. 1A). Seeds of B. juncea cv RLC-3 were procured from Department of Plant Breeding and Genetics, Punjab Agricultural University, Ludhiana. Seeds were surface sterilised using 0.05% sodium hypochlorite for about 15 min followed by washing with double distilled water (DDW). The seeds were sowed over petri-plates lined with whatman filter paper containing different concentrations of Cd (10, 30, 50 µM CdCl2).The seeds were raised for three days and three days old seedlings were inoculated with liquid cultures of S. indica and then transferred to germinating towel sheets (14.5 × 42.5 cm) following cigar roll method and double distilled water was taken as physiological control (CN). The germinating towel rolls were transferred to beakers containing 500 ml solutions of aforementioned Cd concentrations. The seedlings were raised as three biological replicates in plant growth chamber with conditions of 16-h light/8-h dark photoperiod, temperature 25 °C (± 2 °C) and 80% relative humidity. The harvesting of seedlings of B. juncea was done 12 days after inoculation (DAI). The inoculation in seedlings was checked through microscopic studies using light microscope. For staining, the roots of B. juncea were harvested and washed with DDW and stored in 10% potassium hydroxide (KOH) followed by normalization with 1N hydrochloric acid and the roots were stained using 0.02% trypan blue later observed under light microscope at at 40 × magnification (Fig. 1B).

Schematic representation of culturing and colonisation of S. indica in B. juncea (A) Cultures of S. indica (B) Root colonisation in B. juncea (C) Morphology of inoculated and non-inoculated roots (visible enhanced differentiation)
The following combinations were studied Double distilled water Control (CN), 10 µM Cd, 30 µM Cd, 50 µM Cd, S. indica inoculated, S. indica + 10 µM Cd, S.indica + 30 µM Cd and S.indica + 50 µM Cd.
%age Seed Germination
The seeds were deemed to have germinated when the radical was at least 1 cm long. The seed germination rate was calculated 72 h after treatment. The germination rate (%) was equal to the number of germinated seeds at 72 h/total number of seeds (%). It was calculated as: Number of seeds that germinated/Total number of seeds × 100.
Estimation of Seedling Biomass
For the purpose of evaluating fresh weight (FW), dry weight (DW), shoot length (SL), root length (RL), and other parameters of 50 seedlings from each treatment in three biological replicates were chosen. The RL and SL were manually measured in centimeters (cm) and their FW was calculated using a weighing balance. Seedlings were dried in an oven at 70 °C for 48 h before being weighed to determine their DW. The units of DW and FW were recorded in milligrams (mg).
Total Soluble Sugar, RS and NRS
Total soluble sugar (TSS) content was determined following the method of Ref. [27]. A 0.5 g dried sample was homogenized in 80% ethanol and centrifuged at 3000 × g for 15 min. After that, ethanol extract (50 µL) was mixed with 0.95 mL distilled water, and 3 mL cold anthrone reagent was added with continuous shaking. The reaction mixture was heated in boiling water for 10 min, cooled rapidly, and the optical density was measured at 630 nm. The estimation of reducing sugars (RS) was performed using the [28] method and determining absorbance at 620 nm using a spectrophotometer (Beckman 640 D, USA). Non-reducing sugar (NRS) content was determined by subtracting the amount of reducing sugars from the total soluble sugars, and the results were expressed as mg/g DW tissue.
Vitamin C and A
Ascorbic acid content in fresh tissue was determined following [29] by homogenizing 500 mg of tissue in metaphosphoric Citrate Buffer (pH 5.4) and centrifuging at 3000 × g for 5 min. The resulting extract (2 mL) was mixed with 8 mL of 2, 6-Dichlorophenol Indophenols dye, and ascorbic acid concentration was quantified at 530 nm, reported in mg g−1 FW. Vitamin A estimation was performed using the method described by Ref. [30]. For extraction, 1 g of fresh shoot tissue was homogenized in 1.0 mL of 2 N KOH in 90% alcohol followed by heating and later cooling. The extraction was done using 10 ml of petroleum ether in separating funnel and the 5 ml of aliquot of the extract of petroleum ether was evaporated at 60 °C followed by addition of 1 ml of chloroform and for estimation 2 mL of trichloroacetic acid was added and absorbance was taken at 620 nm and results were calculated using standard vitamin A.
Total Phenol Content and Total Flavonoid Content
The method for extracting and measuring total phenol content (TPC) and total flavonoid content (TFC) followed the procedure outlined by Jaafar et al. 2010. In brief, 0.5 g of plant tissue was homogenized in 3 mL of methanol. After centrifuging the homogenate at 5000 rpm for 10 min, used the supernatant for quantification. For total phenolics, 200 µl of the extract was mixed with Folin–Ciocalteu reagent (1.5 mL) and left at 22 °C for 5 min before adding sodium carbonate (1.5 mL). The final mixture was incubated in the dark at room temperature for 60 min, and measured absorbance at 725 nm. Results were expressed as μg g−1 DW and gallic acid was used as stamdard. For total flavonoid determination, a sample of 1 mL was mixed with NaNO3 (0.3 mL) in a covered test tube, followed by the addition of 10% AlCl3 (0.3 mL) and 1 M NaOH (2 mL). The absorbance of this mixture was measured at 510 nm using rutin as a standard, and the results were expressed as μg g−1 DW.
Statistical analysis
Statistical analyses were performed using Graph Pad Prism v8, with one-way ANOVA and Tukey’s test (p < 0.05) for multiple comparisons. Standard deviation is represented by the error bar (SD ≤ 0.05). A significant difference has been marked by the stars above the bars. The experiment was conducted in three biological duplicates.
Results
Inoculation with S. indica Helps to Improve the Growth Parameters of B. juncea Seedlings Under Cd Stress
Seed germination, seedling vitality, and optimum biomass are considered to be essential traits that determine the efficient productivity of the plant in the field. Therefore, any condition of stress during the initial phase of growth proves to be deteriorating for the plants. The seed germination percentage was calculated in the present study to underpin the effect of different concentrations of Cd on seed germination, and the results were compared with CN. %age seed germination was found to be reduced in all three Cd concentrations (10 µM, 30 µM and 50 µM Cd), and the highest decline in seed germination was found in 50 µM Cd-stressed B. juncea seeds, which presented a decline of 31.52% as compared to that of CN seedlings. The seedlings after root emergence were inoculated with S. indica, and the remaining parameters were analyzed. The plant biomass parameters decline significantly (Table 1).
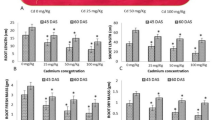
Furthermore, a noticeable decline in RL and SL was observed in the case of metal-stressed seedlings, where it was found that a significant decrease in RL of 27.44%, 23.17%, and 33.24% was found in 10 µM, 30 µM, and 50 µM Cd-exposed seedlings, respectively, as compared to CN (Fig. 2A). However, it was observed that inoculating seedlings with S. indica helped in mitigating the impacts of Cd stress in B. juncea seedlings. RL was found to increase in the case of inoculated seedlings as compared to Cd-stressed seedlings, and maximum enhancement was found in the case of 50 µM Cd-exposed inoculated seedlings, which was 20.66% higher as compared to metal treatment of 50 µM Cd. Also, a visibly more differentiation in root morphology was evident in S. indica inoculated roots (Fig. 1C). Similarly, in case of SL of B. juncea seedlings, a maximum decline of 40.42% was observed at 50 µM Cd, which was the highest decline in SL as compared to CN seedlings (Fig. 2B). Again, an increasing trend was observed in SL when inoculated with S. indica which suggests the mitigation potential of this fungal endophyte against Cd stress in maintaining the morphology of the seedlings that were exposed to Cd stress right from germination.

Effect of S. indica on seedling growth parameters under different Cd concentrations. Growth parameters: A RL (root length), B SL (shoot length), C FW (fresh weight), and D DW (dry weight)
FW and DW are other essential parameters of growth as they give an idea about the healthy biomass of the seedlings. Cd stress impacted the FW and DW of B. juncea seedlings, which is evident from the results. A declining trend in both FW and DW was observed when the concentration of Cd was increased from 10 to 50 µM, signifying a more deleterious impact of a higher concentration of the heavy metal Cd (Fig. 2C & D). Nevertheless, S.indica + 30 µM and S. indica + 50 µM showed an increase in DW of 22.96% and 22.67%,respectively, as compared to the respective Cd concentrations of 30 µM Cd and 50 µM Cd exposed seedlings of B. juncea (Fig. 2D). Maximum enhancement of FW was found at S.indica + 50 µM Cd treatment, which was 9.13% higher as compared to only 50 µM Cd treatment (Fig. 2C). Owing to the improved DW by S. indica inoculation, the percentage heavy metal tolerance index was found to be improved as compared to the respective Cd stress conditions.
Carbohydrate Machinery Improved with S. indica Inoculation
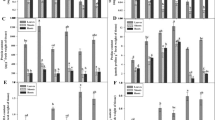
TSS is the ultimate source of energy for plants and is readily available for utilization. Under Cd-stressed seedlings, it was evident from the results that a reduction in TSS was found as compared to CN seedlings, and the maximum reduction was at 50 µM Cd, which was an 18.55% decline as compared to Cd, which signifies the deleterious impact of Cd on photosynthesis and ultimately TSS (Fig. 3A). Furthermore, the S. indica inoculated seedlings demonstrated an increment of 16.67% in TSS as compared to CN seedlings, along with improving the content of TSS in Cd-stressed seedlings, thereby giving a clear indication of an endophyte-mediated improved carbon energy source. An increase of 4.7%, 15.89%, and 25.08% was found in S. indica inoculated in seedlings experiencing stress of 10 µM Cd, 30 µM Cd, and 50 µM Cd, respectively (Fig. 3A). Incoulation with S. indica helped in increasing the contents of RS, and an increment of 40.48% in RS was found as compared to CN seedlings (Fig. 3B). The content of RS was reduced under Cd-stressed seedlings, and the maximum decline was at 30 µM Cd, and this was increased by 10.06% when inoculated with the endophyte S. indica. A similar decrease in NRS content was evident in Cd-stressed seedlings of B. juncea, but the maximum decline was at 50 µM Cd, which was a 35.14% decrease as compared to CN seedlings, but inoculation of S. indica enhanced it to 30.52% as compared to metal-stressed seedlings (Fig. 3C).

Effect of S. indica on carbohydrate content under different Cd concentrations. A TSS (total soluble sugars), B RS (reducing sugars), and C NRS (non reducing sugars)
Effect of S. indica + Cd on the Vitamin A and C Content of B. juncea Seedlings
The content of vitamin A was found to increase under the condition of Cd stress, which was 20.12%, 23.17%, and 6.7% increases in 10 µM Cd, 30 µM Cd, and 50 µM Cd treatments, respectively, as compared to CN seedlings (Fig. 4A). The least increase in vitamin A was at 50 µM Cd, which suggests toxic effects of Cd that weaken the machinery of plants after prolonged exposures. Serendipita indica inoculation helped to maintain levels of vitamin A and seedlings inoculated with S. indica exhibited a 19.51% increase in content as compared to CN seedlings. In the case of vitamin C content, similar trends of increase under Cd-stressed seedlings were observed and S. indica inoculated seedlings demonstrated a notable increase of 6.33% as compared to CN seedlings (Fig. 4B).

Effect of S. indica on vitamin A (A) and vitamin C (B) under different levels of cadmium (Cd) stress
Cd Stress Increases TPC and TFC
TPC is found to increase in conditions of heavy metal stress, which is a response of plants to act against any stressful condition. These biomolecules act as excellent chelating agents and have tendency to act as ROS scavengers. An increase in TPC was found under all the treatments with Cd, and again, the maximum increase was evident in the 50 µM Cd treatment. Notably, the results revealed that S. indica inoculation increased the levels of phenols as compared to CN seedlings but reduced the level in the case of metal-stressed seedlings as compared to their respective metal concentrations. Serendipita indica inoculation enhanced the levels of TPC by 26.66% as compared to CN seedlings, which can provide evidence to explore S. indica for biofortification of B. juncea seedlings (Fig. 5A). Furthermore, the maximum increase in TPC in the combination of Cd and S. indica was found in S. indica + 30 µM treatment, which was 55.29% more as compared to the CN seedlings of B. juncea. Similarly, in the case of TFC, an increase in content under Cd was evident from the biochemical results, and the S. indica inoculation demonstrated an increase of 9% as compared to CN seedlings of B. juncea (Fig. 5B). Also, in the case of Cd-stressed seedlings inoculated with S. indica, it was observed that S. indica + 30 µM treatment showed an enhancement of 28.13% as compared to the respective metal concentration of 30 µM Cd.

Effect of S. indica on phenol and flavanol content under different levels of cadmium (Cd) stress. A TPC (total phenol content) and B TFC (total flavonoid content)
Overall, the results of this study revealed that S. indica is a potential endophyte for improving growth parameters in terms of FW, DW, RL, and SL. Also, S. indica inoculation improved the contents of sugar, which suggests the role of this endophyte in improving the energy source of seedlings even under Cd stress. Furthermore, managing the levels of TPC and TFC proves the efficacy of S. indica in mitigating the deleterious impacts of Cd stress as phenols are known to play roles of chelating agents and ROS scavengers along with other growth stimulating functions.
Discussion
Cd toxicity detrimentally impacts overall health of the plants [31]. It significantly declines the productivity of plants as it leads to oxidative burst due to Cd entry into plants, disrupts cellular machinery, hinders metabolism and normal physiology of plants like mineral nutrition, thereby impacting optimum governing of the metabolic processes [11, 15, 31, 32]. Cd holds no biological role in plants therefore; any minute concentration of it will lead to toxic effects. The same has been evident in the results of present study where Cd impacted all the metabolic processes beginning from seed germination. The seed germination was found to decrease in the results of the study thereby suggesting the phytotoxic impacts of Cd on the reserve food materials of the seed which accelerates the reduction in seed germination %age [33,34,35]. Also, reduction in seed germination can be attributed to the detrimental effects of Cd on seeds’ reserve food and functioning of enzymes which trigger mobilization of reserve food materials [33]. Similar reduction in seed germination has been observed in studies of Vigna radiata, Oryza sativa and Triticum aestivum [33,34,35]. Furthermore, Cd impacts the seedling growth which was evident from the reduction of SL and RL in B. juncea seedlings and the similar results have been obtained in O. sativa and mungbean seedlings thereby demonstrating the impacts of Cd on early seedling growth traits [33, 34]. Reduction in SL, RL, FW and DW was evident in C. cajan seedlings and T. aestivum seedlings exposed to toxic levels of Cd [18, 36, 37]. The reduction in the biomass characteristics of early plant growth when exposed to Cd stress in present study suggests that even minimal concentration of Cd leads to deteriorating outcomes pertaining to seedling growth which directly impact the future plant health (Fig. 2). The reduction in growth parameters can directly be attributed to interference of Cd with cell division which leads to perturbed growth [38]. However, in order to build a strong retaliating capacity of B. juncea seedlings, inoculation with S. indica proved to be beneficial in ameliorating the growth parameters like RL, SL, FW and DW. The improved growth traits with S. indica inoculation have been reported previously in different plant species under variety of stress condition [39,40,41]. Additionally, S. indica can be inoculated in broad spectrum of plants [42] and this study adds to the previous research of successful colonization of S. indica in B. juncea [43].
One of the major targets during stress is carbohydrate content of a plant and sugar contents show fluctuation during conditions of stress [44, 45]. Results of the present study revealed increase in TSS and RS content during mild toxic concentrations of Cd (10 µM Cd) and this further declined during higher Cd concentration (50 µM Cd) (Fig. 3). Increase in content of sugars has been reported under heavy metal stress condition as a response of plant to act against stress [44, 46]. Sugars act as vital energy sources for the plants and help them to combat stress condition along with acting as ROS scavengers and osmolytes during conditions of stress thereby adding to the stress management efficacy of the plants [45,46,47,48,49,50]. Reducing sugars have tendency to neutralize ROS being produced during stress, therefore, carbohydrates have dual role of scavenging ROS besides acting as energy sources [50]. Serendipita indica inoculation has been known to enhance the levels of carbohydrates in terms of TSS, NRS and RS under both stressed and unstressed conditions [51, 52]. Nevertheless, this study adds to the existing reservoir of knowledge that S. indica inoculation helps in enhancing sugars in B. juncea.
Vitamin C is known to play several physiological functions in living organisms along with acting as a potential antioxidant [53]. It has direct role to play in scavenging ROS and chelation of Cd ions [11]. The results of the present study revealed that S. indica inoculation enhanced the levels of Vitamin C which helps in strengthening the non-enzymatic antioxidant defense system of B. juncea seedlings under Cd stress along with promoting the nutritional value [54]. Contents of vitamin A were found to enhance under treatment of both Cd stress and S. indica inoculation. Endophytes have been reported to enhance the levels of vitamins in various crops which pinpoints their role in shielding the plants as well as biofortification [55, 56]. Furthermore, this is pioneer study to report S. indica inoculation effect on vitamin content in B. juncea seedlings under Cd stress which can be further explored and use S. indica as potent candidate of biofortification.
A substantial increase in TPC and TFC was detected in B. juncea seedlings with or without S. indica inoculation exposed to Cd stress conditions (Fig. 5). TPC and TFC act as essential metabolites in plants which are known to play variety of roles under heavy metal stress [57, 58]. They are essential chelating agents which help in sequestering Cd ions thereby preventing them to cause metabolic damage to plants [59]. Also, phenols and flavonoids have tendency to act as antioxidants and neutralize ROS which are produced during conditions of Cd stress and avoid any inevitable damage associated with them [57, 59, 60]. Increase in content in TPC and TFC is a strategy of plant to avoid any deleterious impacts of stress and similar results have been obtained by Ref. [18, 59]. Serendipita indica inoculation is known to increase tolerance ability of plants under heavy metal stress by enhancing secondary metabolites like TFC and TPC [23]. Therefore, this study presents a significant evidence of enhancement of TPC and TFC with S. indica inoculation which is beneficial in improving chelating capacity of plants along with consolidating the antioxidant defense system of plants.
Role of endophyte S. indica can be greatly acknowledged in inculcating Cd stress tolerance in B. juncea seedlings as per the results of present study. Serendipita indica is reported to shield rice against Cd stress conditions as reported by Adil et al. [61], in soybean [62], wheat [63], fenugreek [64]. Serendipita indica helps in strengthening the healthy establishment of seeding stage of the crop which is necessary It is imperative to elucidate the molecular mechanisms underlying this potential of endophyte before its implications in fields under variable environmental cues.
Conclusion
The study tested the efficacy of S. indica inoculation against toxicity of Cd in B. juncea seedlings. The major advantage of utilizing S. indica in the study is that it is capable of producing axenic cultures and inoculates with broad spectrum of plants and provides them immense benefits as obtained in B. juncea. Serendipita indica has been proved to be beneficial in the present study as it improved overall growth parameters like RL, SL, FW and DW along with enhancing the carbohydrate levels which serve as source of energy for plants along with acting as non-enzymatic defense molecules to scavenge ROS. It also managed levels of vitamins, TPC and TFC which also have role in scavenging ROS which may be generated upon entry of Cd into B. juncea seedlings. The enhanced tolerance level of B. juncea against Cd stress helped in inculcating tolerance without compromising the growth which suggests S. indica can be utilized as microbial partner in phytoremediation as well as bioremediation process. However, this study requires field experimental validations which are in progress. Overall, S. indica has potential to enhance capacity of B. juncea seedlings to overcome Cd stress without compromising its growth.
References
Jacoby R, Peukert M, Succurro A, Koprivova A, Kopriva S (2017) The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front Plant Sci 8:292271
Baron NC, Rigobelo EC (2022) Endophytic fungi: a tool for plant growth promotion and sustainable agriculture. Mycology 13:39–55
Grabka R, d’Entremont TW, Adams SJ, Walker AK, Tanney JB, Abbasi PA, Ali S (2022) Fungal endophytes and their role in agricultural plant protection against pests and pathogens. Plants 11:384
Abdelaziz ME, Abdelsattar M, Abdeldaym EA, Atia MA, Mahmoud AWM, Saad MM, Hirt H (2019) Piriformospora indica alters Na+/K+ homeostasis, antioxidant enzymes and LeNHX1 expression of greenhouse tomato grown under salt stress. Sci Hortic 256:108532
Saddique MAB, Ali Z, Khan AS, Rana IA, Shamsi IH (2018) Inoculation with the endophyte Piriformospora indica significantly affects mechanisms involved in osmotic stress in rice. Rice 11:1–12
Verma A, Shameem N, Jatav HS, Sathyanarayana E, Parray JA, Poczai P, Sayyed RZ (2022) Fungal endophytes to combat biotic and abiotic stresses for climate-smart and sustainable agriculture. Front Plant Sci 13:953836
Liu YANG, Jin-Li CAO, Zou YN, Qiang-Sheng WU, Kamil KUČA (2020) Piriformospora indica: a root endophytic fungus and its roles in plants. Not Bot Horti Agrobo 48:1–13
Gill SS, Gill R, Anjum NA, Sharma KK, Johri AK, Tuteja N (2016) Piriformospora indica: potential and significance in plant stress tolerance. Front Microbiol 7:184779
Verma S, Varma A, Rexer KH, Hassel A, Kost G, Sarbhoy A, Bisen P, Bütehorn B, Franken P (1998) Piriformospora indica, gen. et sp. nov., a new root-colonizing fungus. Mycologia 90:896–903
Mensah RA, Li D, Liu F, Tian N, Sun X, Hao X, Lai Z, Cheng C (2020) Versatile Piriformospora indica and its potential applications in horticultural crops. Hort Plant J 6:111–121
Haider FU, Liqun C, Coulter JA, Cheema SA, Wu J, Zhang R, Wenjun M, Farooq M (2021) Cadmium toxicity in plants: impacts and remediation strategies. Ecotoxicol Environ Saf 211:111887
Zulfiqar U, Farooq M, Hussain S, Maqsood M, Hussain M, Ishfaq M, Ahmad M, Anjum MZ (2019) Lead toxicity in plants: impacts and remediation. J Environ Manage 250:109557
Labudda M, Dziurka K, Fidler J, Gietler M, Rybarczyk-Płońska A, Nykiel PB, Morkunas I, Muszyńska E (2022) The alleviation of metal stress nuisance for plants—a review of promising solutions in the face of environmental challenges. Plants 11:2544
Palansooriya KN, Shaheen SM, Chen SS, Tsang DC, Hashimoto Y, Hou D, Bolan NS, Rinklebe J, Ok YS (2020) Soil amendments for immobilization of potentially toxic elements in contaminated soils: a critical review. Environ Int 134:105046
Kubier A, Wilkin RT, Pichler T (2019) Cadmium in soils and groundwater: a review. Appl Geochem 108:104388
Tomza-Marciniak A, Pilarczyk B, Marciniak A, Udała J, Bąkowska M, Pilarczyk R (2019) Cadmium, Cd. mammals and birds as bioindicators of trace element contaminations in terrestrial environments: an ecotoxicological assessment of the northern hemisphere, pp 483–532
Shi T, Zhang Y, Gong Y, Ma J, Wei H, Wu X, Zhao L, Hou H (2019) Status of cadmium accumulation in agricultural soils across China (1975–2016): from temporal and spatial variations to risk assessment. Chemosphere 230:136–143
Kaushik S, Ranjan A, Singh AK, Sirhindi G (2024) Methyl jasmonate reduces cadmium toxicity by enhancing phenol and flavonoid metabolism and activating the antioxidant defense system in Cajanus cajan. Chemosphere 346:140681
Kaur G, Kaushik S, Madaan I, Sidhu A, Kaur H, Sirhindi G (2023) Exogenous application of biostimulants and Cd stress tolerance. In: Biostimulants in alleviation of metal toxicity in plants. Academic Press, pp 207–227
Goswami S, Das S (2015) A study on cadmium phytoremediation potential of Brassica juncea. Int J Phytoremediation 17:583–588
Izadi F, Ghabooli M, Rostami M, Movahedi Z (2021) Evaluation of fungus Piriformospora indica effects on some morphophysiological traits of radish under heavy metal stress. J Agric Sci Sustain Prod 31:117–129
Nanda R, Agrawal V (2018) Piriformospora indica, an excellent system for heavy metal sequestration and amelioration of oxidative stress and DNA damage in Cassia angustifolia Vahl under copper stress. Ecotoxicol Environ Saf 156:409–419
Khalid M, Kayani SI, Tang K (2020) The ameliorative effects of exogenous inoculation of Piriformospora indica on molecular, biochemical and physiological parameters of Artemisia annua L. under arsenic stress condition. Ecotoxicol Environ Saf 206:111202
Su Z, Zeng Y, Li X, Perumal AB, Zhu J, Lu X, Lin F (2021) The endophytic fungus Piriformospora indica-assisted alleviation of cadmium in tobacco. J Fungi 7:675
Hui F, Liu J, Gao Q, Lou B (2015) Piriformospora indica confers cadmium tolerance in Nicotiana tabacum. J Environ Sci 37:184–191
Hill TW, Kafer E (2001) Improved protocols for Aspergillus minimal medium: trace element and minimal medium salt stock solutions. Fungal Genet Rep 48:20–21
Loewus FA (1952) Improvement in anthrone method for determination of carbohydrates. Anal Chem 24:219
Somogyi M (1952) Determination of reducing sugars by Nelson-Somogyi method. J Biol Chem 200:245
Chinoy JJ, Gurumurti K (1976) Colorimetric determination of ascorbic acid turnover in plants. Springer, New York
Bayfield RF, Cole ER (1980) Colorimetric estimation of vitamin A with trichloroacetic acid. In: Methods in enzymology, vol 67. Academic Press, pp 189–195
Zulfiqar U, Ayub A, Hussain S, Waraich EA, El-Esawi MA, Ishfaq M, Ahmad M, Ali N, Maqsood MF (2022) Cadmium toxicity in plants: recent progress on morpho-physiological effects and remediation strategies. J Soil Sci Plant Nutr 22:212–269
Bashri G, Singh S, Prasad SM, Ansari MJ, Usmani S, Alfarraj S, Alharbi SA, Brestic M (2021) Kinetin mitigates Cd-induced damages to growth, photosynthesis and PS II photochemistry of Trigonella seedlings by up-regulating ascorbate-glutathione cycle. PLoS ONE 16:e0249230
Anwar S, Shafiq F, Nisa ZU, Usman U, Ashraf MY, Ali N (2021) Effect of cadmium stress on seed germination, plant growth and hydrolyzing enzymes activities in mungbean seedlings. J Seed Sci 43:e202143042
He J, Ren Y, Chen X, Chen H (2014) Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol Environ Saf 108:114–119
Ahmad I, Akhtar MJ, Zahir ZA, Jamil A (2012) Effect of cadmium on seed germination and seedling growth of four wheat (Triticum aestivum L.) cultivars. Pak J Bot 44:1569–1574
Shi Z, Yang S, Han D, Zhou Z, Li X, Liu Y, Zhang B (2018) Silicon alleviates cadmium toxicity in wheat seedlings (Triticum aestivum L.) by reducing cadmium ion uptake and enhancing antioxidative capacity. Environ Sci Pollut Res 25:7638–7646
Zhou Z, Wei C, Liu H, Jiao Q, Li G, Zhang J, Zhang B, Jin W, Lin D, Chen G, Yang S (2022) Exogenous ascorbic acid application alleviates cadmium toxicity in seedlings of two wheat (Triticum aestivum L.) varieties by reducing cadmium uptake and enhancing antioxidative capacity. Environ Sci Pollut Res 29:21739–21750
Shiyu QIN, Hongen LIU, Zhaojun NIE, Rengel Z, Wei GAO, Chang LI, Peng ZHAO (2020) Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: a review. Pedosphere 30:168–180
Yun P, Xu L, Wang SS, Shabala L, Shabala S, Zhang WY (2018) Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoot. Plant Growth Regul 86:323–331
Swetha S, Padmavathi T (2020) Mitigation of drought stress by Piriformospora indica in Solanum melongena L. cultivars. Proc Natl Acad Sci India Sect B Biol Sci 90:585–593
Su ZZ, Wang T, Shrivastava N, Chen YY, Liu X, Sun C, Yin Y, Gao QK, Lou BG (2017) Piriformospora indica promotes growth, seed yield and quality of Brassica napus L. Microbiol Res 199:29–39
Qiang X, Weiss M, Kogel KH, Schäfer P (2012) Piriformospora indica—a mutualistic basidiomycete with an exceptionally large plant host range. Mol Plant Pathol 13:508–518
Sharaya R, Tuteja N, Gill R, Gill SS (2023) Reactive oxygen species (ROS) scavenging and methylglyoxal (MG) detoxification confers salinity tolerance in mustard cultivars inoculated with Piriformospora (Serendipita) indica. S Afr J Bot 163:630–645
Handa N, Kohli SK, Kaur R, Sharma A, Kumar V, Thukral AK, Arora S, Bhardwaj R (2018) Role of compatible solutes in enhancing antioxidative defense in plants exposed to metal toxicity. In: Plants under metal and metalloid stress: responses, tolerance and remediation. pp 207–228
Keunen ELS, Peshev D, Vangronsveld J, Van Den Ende WIM, Cuypers ANN (2013) Plant sugars are crucial players in the oxidative challenge during abiotic stress: extending the traditional concept. Plant Cell Environ 36:1242–1255
Lalelou FS, Fateh M (2014) Effects of different concentrations of zinc on chlorophyll, starch, soluble sugars and proline content in Cucurbita pepo. Int J Biosci 10:6–12
Nägele T, Gibon Y, Le Hir R (2022) Plant sugar metabolism, transport and signalling in challenging environments. Physiol Plant 174:1–3
Siddiqui MH, Alamri S, Al-Khaishany MY, Khan MN, Al-Amri A, Ali HM, Alsahli AA (2019) Exogenous melatonin counteracts NaCl-induced damage by regulating the antioxidant system, proline and carbohydrates metabolism in tomato seedlings. Int J Mol Sci 20(2):353
Ende WVD, Peshev D (2013) Sugars as antioxidants in plants. In: Crop improvement under adverse conditions. Springer, New York, pp 285–307
Ahmad F, Singh A, Kamal A (2020) Osmoprotective role of sugar in mitigating abiotic stress in plants. In: Protective chemical agents in the amelioration of plant abiotic stress: biochemical and molecular perspectives. Wiley, New York, pp 53–70
Ghabooli M (2014) Effect of Piriformospora indica inoculation on some physiological traits of barley (Hordeum vulgare) under salt stress. Chem Nat Compd 50:1082–1087
Kaboosi E, Ghabooli M, Karimi R (2022) Piriformospora indica inoculants enhance flowering, yield, and physiological characteristics of tomato (Lycopersicon esculentum) in different growth phases. Iran J Plant Physiol 12:4183–4194
Fenech M, Amaya I, Valpuesta V, Botella MA (2019) Vitamin C content in fruits: biosynthesis and regulation. Front Plant Sci 9:413553
Arora M, Saxena P, Abdin MZ, Varma A (2020) Interaction between Piriformospora indica and Azotobacter chroococcum diminish the effect of salt stress in Artemisia annua L. by enhancing enzymatic and non-enzymatic antioxidants. Symbiosis 80:61–73
Molina-Montenegro MA, Escobedo VM, Atala C (2023) Inoculation with extreme endophytes improves performance and nutritional quality in crop species grown under exoplanetary conditions. Front Plant Sci 14:1139704
Waqas M, Kim YH, Khan AL, Shahzad R, Asaf S, Hamayun M, Kang SM, Khan MA, Lee IJ (2017) Additive effects due to biochar and endophyte application enable soybean to enhance nutrient uptake and modulate nutritional parameters. J Zhejiang Univ Sci B 18:109
Chen S, Wang Q, Lu H, Li J, Yang D, Liu J, Yan C (2019) Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia obovata under Cd and Zn stress. Ecotoxicol Environ Saf 169:134–143
Martinez V, Mestre TC, Rubio F, Girones-Vilaplana A, Moreno DA, Mittler R, Rivero RM (2016) Accumulation of flavonols over hydroxycinnamic acids favors oxidative damage protection under abiotic stress. Front Plant Sci 7:838
Yadav P, Kaur R, Kohli SK, Sirhindi G, Bhardwaj R (2016) Castasterone assisted accumulation of polyphenols and antioxidant to increase tolerance of B. juncea plants towards copper toxicity. Cogent Food Agric 2:1276821
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Adil MF, Sehar S, Ma Z, Tahira K, Askri SMH, El-Sheikh MA, Ahmad A, Zhou F, Zhao P, Shamsi IH (2024) Insights into the alleviation of cadmium toxicity in rice by nano-zinc and Serendipita indica: modulation of stress-responsive gene expression and antioxidant defense system activation. Environ Pollut 350:123952
Wang X, Fan X, Wang W, Song F (2022) Use of Serendipita indica to improve soybean growth, physiological properties, and soil enzymatic activities under different Cd concentrations. Chem Biol Technol Agric 9:66
Aliloo AA, Shahabıvand S (2016) Piriformospora indica promotes growth and antioxidant activities of wheat plant under cadmium stress. Yuzuncu Yıl Univ J Agric Sci 26:333–340
Sakhai F, Movahedi Z, Ghabooli M, Mohseni Fard E (2021) Effect of Piriformospora indica inoculation on some morphophysiological traits of fenugreek under cadmium stress. Iran J Seed Sci Technol 10:123–140
Acknowledgements
The Department of Botany, Punjabi University, Patiala (coordinate, DSA-I of UGC and FIST of DST, New Delhi) is acknowledged for providing laboratory and infrastructural facilities.
Author information
Authors and Affiliations
Contributions
GK: Performed experiments and prepared original draft; PS: Data analysis; SK: Helped in experiments; IM: Aided in culturing; AV: Editing; SSG: Conceptualization of idea and provided with S. indica cultures for the study; GS: Conceptualized idea, supervision, reviewing & editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kaur, G., Singh, P., Kaushik, S. et al. Endophyte-Mediated Reinforcement of Morpho-Physiology of Brassica juncea Under Cadmium Toxicity. Indian J Microbiol (2024). https://doi.org/10.1007/s12088-024-01375-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12088-024-01375-7