
Abstract
Medicinal plants are the backbone of modern medicine. In recent times, there is a great urge to discover nootropic medicinal plants to reverse cognitive dysfunction owing to their less adverse effects. Alzheimer’s disease (AD) is an age-related neurodegenerative disorder characterized by the inevitable loss of cognitive function, memory and language impairment, and behavioral disturbances, which turn into gradually more severe. Alzheimer’s has no current cure, but symptomatic treatments are available and research continues. The number of patients suffering from AD continues to rise and today, there is a worldwide effort under study to find better ways to alleviate Alzheimer’s pathogenesis. In this review, the nootropic and anti-Alzheimer’s potentials of 6 medicinal plants (i.e., Centella asiatica, Clitoria ternatea, Crocus sativus, Terminalia chebula, Withania somnifera, and Asparagus racemosus) were explored through literature review. This appraisal focused on available information about neuroprotective and anti-Alzheimer’s use of these plants and their respective bioactive compounds/metabolites and associated effects in animal models and consequences of its use in human as well as proposed molecular mechanisms. This review progresses our existing knowledge to reveal the promising linkage of traditional medicine to halt AD pathogenesis. This analysis also avowed a new insight to search the promising anti-Alzheimer’s drugs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Alzheimer’s disease (AD) is a progressive, irreversible neurodegenerative disorder that principally influences the older people and is believed to be accountable for most of the dementia cases in people aged 65 or older [1, 2]. The neuropathological hallmarks of AD include deposits of extracellular amyloid β (Aβ) in the form of senile plaques (SPs) and formation of intracellular abnormal tau protein as neurofibrillary tangles (NFTs) [3, 4]. In the USA, AD is presently graded as the sixth dominant reason of death; however, current statistics specify that the disorder may position third, immediately after heart disease and cancer, as a leading reason of dying for older humans [5]. Considering the recent evidence, in the USA, nearly 5.5 million people are living with AD dementia, among them 5.3 million people aged 65 and older and about 200,000 people are below age 65 and feature younger-onset Alzheimer’s [6].
The morphological changes of brain-related with typical aging differ from those that happen with AD [7]. Amid the most remarkable is the aggregation of Aβ and tau hyperphosphorylation in Alzheimer’s brain [8, 9]. Studies suggest that each of these is responsible for the disease process; however, the actual series of events is still obscure [10, 11]. As the disease advances, nerve cells in the brain become damaged to a greater extent [12] and this damage ends in the signs and symptoms of AD. One of the most familiar early symptoms is trouble in remembering the latest events (i.e., short-term memory loss) [13]. Subsequently, as the disease progresses symptoms can incorporate difficulty with language, disorientation, alterations in mood and personality, poor judgment and decision making, loss of motivation, and behavioral issues [14, 15]. As the condition aggravates, people with AD regularly draw away from the circle of relatives and society [16] and progressively, bodily capabilities are misplaced that lead to death [17].
Until now, researchers do not absolutely recognize what causes AD in the majority of the population [18]. Approximately 70% of the risk is thought to be genetic with a lot of genes typically concerned [19, 20]. Diverse risk factors comprise a history of head injuries, high blood pressure, depression or environmental, and way of lifestyle [16]. The disease progression is linked with plaques and tangles inside the brain [21, 22]. AD may vary from individual to individual on account of the significance of any one of these factors in growing or declining the risk of developing [23].
In AD, increased oxidative stress (OS) is proven to be a distinguished and early characteristic of susceptible neurons [24]. Increased reactive oxygen species (ROS) levels are connected with augmented accumulation of Aβ and development of SPs [25]. Furthermore, it is obvious that OS takes places before the formation of NFTs and SPs, where each of which can be neuropathological markers of AD. Nevertheless, the actual mechanisms accountable for this exaggerated effect remain ambiguous [26]. Generally, abnormal function of the central cholinergic system leads to cognitive impairment in AD patients. The change of this acetylcholinesterase (AChE) activity is the key hallmark of AD [27]. No treatments can stop, cure, slow or reverse its progression, although a few may transiently alleviate symptoms [28].
Nature has been a great origin of therapeutic agents from the beginning of human civilization and has continued to offer humankind with novel therapies. In the AD treatment, the use of natural products is a useful way to raise acetylcholine (ACh) levels essential for memory function [29]. Research on flora with AChE inhibitors activity is presently ongoing, with the intention of inventing novel potent compounds which are more affordable and show less adverse effects than synthetic drugs. Furthermore, chemical components with antioxidant activity can be observed in plants at high concentrations and can be used in the prevention of diverse degenerative diseases [30]. Phytochemical research has proven the existence of numerous valuable compounds, including alkaloids, triterpenes, polyphenols, tannins, sterols, flavonoids, and lignans that demonstrate diverse pharmacological activities, such as anti-cholinesterase, anti-Alzheimer, anti-amyloidogenic, hypolipidemic, antioxidant, and anti-inflammatory effects [31]. Therefore, the purpose of the present review is to provide an ample precise analysis of various studies on medicinal plants (Centella asiatica, Clitoria ternatea, Crocus sativus, Terminalia chebula, Withania somnifera, and Asparagus racemosus) that revealed auspicious nootropic effects and promising against AD pathogenesis.
Materials and Methods
Search Strategy
The available information about traditional nootropic and anti-Alzheimer’s actions of the medicinal plants was collected from various electronic sources like PubMed, EBSCO, and other databases. Furthermore, available books were also used.
Plants Selection
Numerous medicinal plants are effective for nootropic and anti-Alzheimer’s activities. Amid copious plants, 6 promising anti-Alzheimer’s medicinal plants were considered (Table 1). In this appraisal, well-documented anti-Alzheimer’s medicinal plants like Ginkgo biloba and Bacopa monnieri were exempted.
Results and Discussion
Phytopharmakons are attaining the growing importance both as the traditional system of medicine and as modern medicine due to their therapeutic effects. Novel antioxidants can be considered as a safe and effective way of strengthening the body’s defense system against free radicals; ultimately, these can also facilitate protection against degenerative nerve diseases like AD [39]. Here, we represent the potential effects of various medicinal plants as therapeutic in Alzheimer’s pathogenesis.
Centella asiatica
Cognition-Enhancing, Antioxidant, and Anti-Alzheimer’s Effects
Centella asiatica (C. asiatica) is quite well-known to have the capacity to increase attention span and concentration, re-vitalize nervous system and the brain and also to combat aging [40]. A study found that C. asiatica exhibited antioxidant and cognitive-enhancing properties in normal rats. In rats, for 21 days, the effect of an aqueous extract of C. asiatica (i.e., 100, 200, and 300 mg/kg) was estimated in OS induced by streptozotocin (STZ) and cognitive impairment [41]. In an elevated plus maze (EPM) test and passive avoidance test (PAT), it was found that the rats, which were treated with C. asiatica, exhibited an upsurge in cognitive behavior in a dose-dependent manner. In case of rats which were treated with C. asiatica at doses of 200 mg/kg and 300 mg/kg found to have a noteworthy reduction in malondialdehyde (MDA) and an upsurge in catalase (CAT) and glutathione (GSH) levels. Impairment in endogenous antioxidant mechanism or OS is regarded as a crucial factor in AD. In addition, in humans, cognitive deficits are often found in the older people and the STZ in rats has been associated to sporadic form of AD. In the above study, generation of free radical in the model was found to be connected with cognitive impairments. Furthermore, the findings of the aforesaid study proposed the notable efficacy of C. asiatica in the prevention of OS and cognitive deficits [41].
In a study by Xu et al., [42] stated that the cyclic adenosine monophosphate (cAMP) response element binding protein phosphorylation was increased in both neuroblastoma cell line that expresses Aβ (i.e., 1–42) and in rat embryonic cortical primary cell culture. Moreover, the enhanced cAMP phosphorylation in response element binding and the involvement of 2 major single components were tested. On the other hand, when inhibitors were used in this study, it was found that extracellular signal-regulated kinase/ribosomal S6 kinase signaling pathway might facilitate this effect of C. asiatica extract [42]. Conversely, in a different study by Jayashree et al., [43] reported a significant increase in case of superoxide dismutase (SOD), glutathione peroxidase (GPx) and CAT in lymphoma-bearing mice, when 50 mg/kg/day of a crude methanolic extract of C. asiatica was given orally for 2 weeks. In contrast, it was also noticed that antioxidants like ascorbic acid and GSH were reduced in the animals [43].
In a study, asiatic acid’s derivatives of C. asiatica were found to show substantial neuroprotective effects on cultured cortical cells through their modulation of the mechanism involved in cellular oxidative defense. Henceforth, these agents are efficacious enough to protect neurons from the oxidative damage triggered upon excess glutamate exposure [44]. In another study by Mook-Jung et al., [45] when tested on hippocampal slices and B103 cell cultures, it was confirmed that the asiaticoside derivatives mediated protective effects against Aβ neurotoxicity. Out of 28 of the asiaticoside derivatives, strong inhibition in the cell death induced by free radical and Aβ was exerted by three components including asiatic acid. These agents or derivatives perhaps are good candidates as a treatment of AD that particularly defends neurons from Aβ toxicity [45].
Neuroprotective and Anti-Alzheimer’s Effects
Kumar et al. [46] reported the neuroprotective potential of C. asiatica against memory impairment mediated via colchicine and OS in rats. When the extract of C. asiatica was used for chronic oral treatment (i.e., 150 and 300 mg/kg) for 25 days, beginning 4 days prior to administration of colchicine, this considerably (P < 0.05) attenuated memory impairment induced by colchicine and oxidative damage. Moreover, a marked reversal of colchicines administered rise in AChE activity was also achieved by C. asiatica [46]. Another study in mice examined the neuroprotective potential of C. asiatica aqueous extract in cognitive impairment triggered by scopolamine in mice. Improvement in cognitive impairment mediated by C. asiatica was also compared against a standard drug (i.e., donepezil 50 μg/kg). Conversely, studies conducted in EPM models revealed that C. asiatica considerably (P < 0.001) reduces the retention transfer latency when used at a dose of 300 mg per kg [47].
Soumyanath et al. [48] observed in a study that effects of a water extract of C. asiatica (i.e., prepared by refluxing C. asiatica [120 g] with water [1.5 L] for 2 h, filtering to eliminate plant debris and freeze-drying to produce a residue [11.5 g]) in the Tg2576 mouse, which was a murine model of AD with greater Aβ burden. It was found that water extract of C. asiatica attenuated behavioral abnormalities associated with Aβ in these mice when administered orally. Water extract also protected MC65 human neuroblastoma cells and SH-SY5Y cells from toxicity triggered by endogenously produced and exogenously introduced Aβ, respectively. In MC65 cells, water extract of C. asiatica reduced the intracellular formation of Aβ aggregates. This water extract did not protect neurons from OS and glutamate toxicity and oxidative damage or exhibit anticholinesterase activity, mechanisms of current treatments of Alzheimer’s [48]. Table 2 represents the neuroprotective anticholinergic, cognitive enhancer and antioxidant effects of bioactive constituents of C. asiatica.
Anti-Alzheimer’s Effects in Human Studies
A limited number of randomized, blinded and/or placebo-controlled clinical studies have proved the effect of C. asiatica in enhancing memory and improving cognitive functions. In a double-blind, placebo-controlled, randomized study, Wattanathorn et al. [69] examined the C. asiatica effect on cognitive function of the healthy elderly volunteer. Moreover, 28 healthy elderly subjects received the extract of the plant once daily for two months at various doses ranging from 750, 500, and 250 mg. By using the event-related potential and the computerized test battery, cognitive performance was evaluated. In contrast, the mood was evaluated by employing Bond-Lader Visual Analogue Scales prior to the trial and after single, one and two months following treatment. The findings revealed that working memory and enhanced N100 component amplitude of event-related potential were increased due to the high dose of the plant extract. Moreover, following the C. asiatica treatment, enhancements of self-rated mood were also noticed [69].
Another study was conducted by Tiwari et al. [70] involving 60 elderly subjects of age group sixty-five and above. In that study, Mini-Mental State Examination (MMSE) was used as the diagnostic tool. Furthermore, to monitor the subjects, parameters like Instrumental Activities of Daily Living, Activities of Daily Living, and Yesavage Geriatric Depression Scale were used. On the other hand, in a dose of 500 mg twice daily for 6 months, aqueous extract of C. asiatica was prescribed. Remarkably, a promising improvement was noticed in mild cognitive impairment (MCI) together with other problems such as constipation, loss of appetite, insomnia, and hypertension [70]. Figure 1 depicts the possible mechanism of action for anti-Alzheimer’s potential of C. asiatica.

Possible mechanism of action for anti-Alzheimer’s action of Centella asiatica extracts. Binding of CAE to cell surface receptor generates a signal that causes the production of a second messenger, which in turn activates a protein kinase that translocates to the nucleus and activates CREB. The activated CREB then binds to a CRE region and regulates gene expression to enhance the arborization of neurons and improve cognitive performances [42]. CAE, Centella asiatica extracts; CREB, cyclic AMP response element binding protein; CRE, cyclic AMP response element
Clitoria ternatea
Learning, Memory-Enhancing, and Anti-Alzheimer’s Effects
In a study by Rai et al. [71] assessed the effects of Clitoria ternatea (C. ternatea) aqueous root extract on memory and learning in neonatal rat pups (7 days old) by using PAT, EPM, and T-maze test or spatial learning tests. Interestingly, it was found in this study that memory in rats increased considerably when an extract of C. ternatea roots was used at different doses as an oral treatment. Conversely, neonatal rat pups were intubated at the dose of 50 mg/kg and 100 mg/kg of aqueous root extract of C. ternatea for 30 days during their growth spurt period. Memory improving properties was observed with C. ternatea root extract; nonetheless, it did not disturb motor activity. In addition, in the case of both time points of behavioral tests, it showed enhanced retention and spatial learning performance. Henceforth it was evident that permanent change in the brain mediated by C. ternatea extracts resulted in enhanced learning and memory. This memory increasing activity was significantly higher in neonatal rats (i.e., especially during their growth spurt period) treated for 30 days with C. ternatea extract (100 mg/kg body weight). So, it seems that treatment with C. ternatea extract can cause permanent changes in the rat’s brain and can enhance memory and learning [71].
In rats, in case of the alcoholic extracts of aerial parts and roots of C. ternatea (i.e., 300 and 500 mg/kg) lessened amnesia induced by electroshock through conditional avoidance response paradigm [72]. C. ternatea extract, at the dose of 300 mg/kg, triggered substantial retention of memory and the root parts found to be significantly effective; however, the dose of 300 mg/kg appeared very high. The researchers in this study also assessed the most likely mechanism by which C. ternatea exerts the anti-amnesic effects on central cholinergic activity. For this assessment, they evaluated the whole brain’s acetylcholine (ACh) content and AChE activity at different parts of the rat brain, specifically midbrain, cerebral cortex, cerebellum, and medulla oblongata. In this study, in comparison with aerial parts, C. ternatea extract of roots was found to be more effective in lessening memory deficits. Alternatively, the pathway via which C. ternatea generated retention of memory seems to be analogous to pyritinol (i.e., standard cerebro protective drug). In rat hippocampus, the effects of aqueous C. ternatea root extract on the ACh content were discovered [73, 74]. In contrast, when C. ternatea aqueous root extracts were used at a dose of 100 mg/kg for 30 days in neonatal rat pups (i.e., 7 days old) and young adult rats (i.e., 60 days old), substantial increase in the content of ACh was noticed in their hippocampi in comparison with that of age-matched controls. In 90-day old control rats, hippocampal ACh content was found to be considerably less as compared to 37-day old control rats. In the central nervous system, ACh is considered as one of the key neurotransmitters. ACh helps to enhance attention and also ease the process of learning. As a result, in rat hippocampus, increased ACh content possibly the neurochemical basis for enhanced learning and memory. Treatment with aqueous C. ternatea root extract might be effective in strengthening disrupted cholinergic transmission in specific age-related memory disorders. Furthermore, in normal individuals, learning and enhanced memory may also be facilitated by aqueous root extract of C. ternatea. However, it is yet to be discovered whether this extract exerts similar action in senile animals or not [75].
Rai et al. [76], in another study, showed that retention in the PAT memory can also be ominously improved by using C. ternatea. When young adult rats were orally intubated for 30 days at a dose of 100 mg/kg per day, increased retention was noticed 48 h after the learning process in the same group of C. ternatea treated young adult rats in comparison with age-matched saline controls. This further specifies an upsurge in the capacity to maintain avoidance memories, suggesting possible effects exerted by C. ternatea on amygdaloid neurons. These findings collectively specify that treatment by aqueous root extract of C. ternatea has a specific contribution in the increment of amygdaloid neuronal dendritic arborization and also hippocampal CA3 neurons [76]. In a different study Rai et al., [77] further assessed the effect of C. ternatea aqueous root extract on the dendritic cytoarchitecture of amygdala neurons [77]. Along with age-matched saline controls, when Wistar rats (i.e., either sex) were orally intubated with C. ternatea aqueous root extract (i.e., 50 and 100 mg/kg) for 30 days and exposed to PAT, a significant increase was found in retention of passive avoidance and learning. These rats were sacrificed by decapitation after the PAT. The amygdala was treated for Golgi staining and following staining neurons were traced by using a camera lucida and examined. The findings of the study revealed a substantial increase in branching points, dendritic processes, and dendritic intersections that arise from the soma of amygdaloid neurons in aqueous root extract treated rats when compared to age-matched saline controls. Besides that, in the aqueous C. ternatea root extract, treated rats enhanced dendritic arborization of amygdaloid neurons links with the improved passive avoidance learning and memory. The results of the study showed that aqueous root extract of C. ternatea improves memory by triggering the functional growth of amygdala neurons. Alternatively, an aqueous root extract of C. ternatea also influences the structures of the brain which governs learning and memory specifically amygdala and hippocampus. These changes related to behavior are considered comparatively permanent and denotes that C. ternatea can affect neurotransmitter biosynthesis namely ACh, which plays a significant role in the learning process and also in memory. In addition, these changes can also cause permanent changes in the brain. Moreover, C. ternatea perhaps also facilitates long-term potentiation [71]. The aqueous root extract of C. ternatea was not only found to support neurogenesis in the amygdala; nevertheless, it also triggers the release of neuromodulators or hormones that control the neurotransmitters and neuromodulators activity specifically which play vital roles in memory and learning, thus playing roles in improving memory and learning. Following comprehensive clinical trials in adults, it was detected that treatment by aqueous root extract of C. ternatea possibly be foreseen as an enhancer of memory and also might be beneficial in the treatment of amygdala-related neurodegenerative disorders [77].
Neuroprotective, Cognition-Enhancing, and Anti-Alzheimer’s Effects
Talpate et al. [78] analyzed the nootropic and neuroprotective activity of ethanol extract of C. ternatea extract on diabetes-associated cognitive impairment in a rat model. The ethanol extract of C. ternatea (200 and 400 mg/kg) caused a substantial upsurge in spatial reference memory (P < 0.001), spatial working memory (P < 0.05) and spatial working-reference (P < 0.001) in retention trials on Morris water maze, Y-maze and radial arm maze accordingly. Instead, marked decrease in lipid peroxide (P < 0.001), AChE activity (P < 0.05), total nitric oxide (P < 0.001) and significant increase in CAT, SOD, and GSH levels were detected in animals treated with ethanol extract of C. ternatea (200 and 400 mg/kg) in comparison with diabetic control group [78].
Recently, in a study, Rajashree et al., [79] in the streptozocin-induced young diabetic rats, examined the effects of the alcoholic roots extracts of the C. ternatea on the behavioral and cognitive changes. A substantial improvement was observed only in the memory and learning among the diabetic group and C. ternatea groups of rats [79]. In another study by Ravishankar et al., [80] examined the neuroprotective effect of ethanolic C. ternatea root extracts against amnesia triggered by stress. In this study, conditioned avoidance response by employing Cook’s pole climbing apparatus and transfer latency using EPM were used in stress-induced and normal rats to evaluate cognitive-improving activities. When C. ternatea was orally administered daily at doses of 150 and 300 mg/kg, it improves cognition in a dose-dependent way in normal rats. In extract treated stress-induced rats, fast retrieval was observed as compared to the stress control group [80]. The neuroprotective anticholinergic, cognitive, and memory-enhancing, as well as antioxidant, effects of the C. asiatica extracts and its bioactive metabolites are specified in Table 3 and Table 4.
Neuropharmacological Effects
In rats and mice, C. ternatea root’s ethanol extract was assessed for various neuropharmacological actions including exploratory behavior, general behavior, sleeping time induced by phenobarbitone and muscle relaxant activity. At the doses of 100 and 150 mg/kg, the extract caused a decrease in spontaneous activity, reduction in the exploratory behavioral pattern by Y-maze and the head dip test, decrease in the muscle relaxant activity by rotarod, traction tests, and 300 inclined screens [94]. Furthermore, C. ternatea extract considerably increased the phenobarbitone-mediated sleeping time [94]. In rats, Kulkarni et al. [81] observed the effect exerted by an alcoholic extract of aerial part of C. ternatea on special discrimination. When alcoholic extract was given orally at a dose of 460 mg/kg, a significant increase in the time taken to traverse the maze was noticed, which was alike to that generated by chlorpromazine; nonetheless, at a lower dose (i.e., 230 mg/kg), chlorpromazine was not effective.
Nootropic and Anxiolytic Effects
EPM and object recognition tests were used to evaluate the nootropic activity of C. ternatea aerial parts extract [82]. On the 9th day, a marked increase in inflection ratio was noticed in the case of the animals, which were treated with C. ternatea (100 mg/kg). It was noticed in the object recognition test that mice which were treated with C. ternatea took considerably less amount of time to find out the familiar object as compared to the new object and markedly decreased the discrimination index. So, the increased discrimination index and inflection ratio clearly indicate the nootropic activity of the species. In addition, C. ternatea also met a key condition for nootropic activity, which is specifically the memory improvement in cognitive deficit’s absence. In association with the nootropic drug’s assumption, a decrease in transfer latency was observed by C. ternatea in the EPM test. However, improved inflection ratio on the 9th day denoted C. ternatea’s less significant effect on long-term memory. Conversely, Jain et al. [82] conducted a study by employing the light or dark exploration and EPM test to evaluate the anxiolytic potential of a methanolic extract of aerial part of C. ternatea. It was observed that when C. ternatea (100–400 mg/kg) was administered orally, it augmented the time spent in the light-box and the time spent in the open arm. In contrast, in the same study, it was noticed that there is a reduction in time spent in the dark box. Failure to exert any substantial effect in both animal models of anxiety was noticed with the oral administration of C. ternatea at a dose of 30 mg/kg; however, a weaker effect was detected in case of both the animal models. Malik et al. [95] stated that C. ternatea aqueous methanol extract showed supreme memory-improving and anxiolytic effect at the doses of 100 and 200 mg/kg.
Anti-Alzheimer’s Effects in Human Studies
In a randomized, crossover study, Chusak et al. [96], in 15 healthy men, estimated the effects of C. ternatea flower extract on antioxidant status and postprandial plasma glycemia response. Surprisingly, after 30 min of ingestion, the insulin and postprandial plasma glucose levels were inhibited when consuming sucrose and 2 g and 1 g C. ternatea flower extract. Furthermore, insulin concentration in the fasting state and glucose level in plasma was not altered due to this extract consumption. Instead, in the individuals who received 2 g and 1 g C. ternatea flower extract, a decrease in the level of MDA and a marked rise in plasma antioxidant capacity (i.e., oxygen radical absorbance capacity, trolox equivalent antioxidant capacity, ferric reducing ability of plasma, and protein thiol) were detected. In addition, sucrose-stimulated reduction in trolox equivalent antioxidant capacity and oxygen radical absorbance capacity and also the increase in plasma MDA were protected due to the C. ternatea flower extract consumption. These results suggested that, without hypoglycemia in the fasting state, plasma antioxidant capacity is increased due to an acute ingestion of C. ternatea flower extract. It has been summarized, in adults, following detailed clinical trials, that treatment with aqueous root extract of C. ternatea may be beneficial in the treatment of neurodegenerative diseases relating the amygdala and might also be foreseen as an enhancer of memory [77]. The possible mechanism of action for anti-Alzheimer’s potential of C. ternatea is illustrated in Fig. 2.

Possible mechanism of action for anti-Alzheimer’s action of Clitoria ternatea extracts. Binding of LPS to TLR causes the generation of ROS from NADPH oxidase and mitochondria. This generated ROS activate NF-kB via phosphorylation by the IkB kinase, which then translocates to the nucleus and binds target DNA to produce COX-2, thus causing inflammation. The extract of Clitoria ternatea by blocking few steps of the aforementioned pathway exerts anti-inflammatory effects as well as improves learning, memory, and cognition [97]. CTE, Clitoria ternatea extracts; NADPH, reduced nicotinamide adenine dinucleotide phosphate; LPS, lipopolysaccharide; TIR, Toll-interleukin receptor; ROS, reactive oxygen species; pNF-KB, phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells; COX-2, cyclooxygenase-2
Crocus sativus
Anti-Alzheimer’s Effects in Animal Models
Promising antioxidant properties were found with the aqueous-ethanolic extract of Crocus sativus (C. sativus) stigmas, which were greater as compared to those of tomato and carrot in a time and concentration-dependent manner, in consort with Aβ fibrillogenesis inhibition. Papandreou et al. [98] stated that the ester of crocetin, trans-crocin-4, inhibited Aβ fibrillogenesis. For the sporadic AD, intracerebroventricular streptozotocin injection to rodents has been found to be commonly used animal model [41, 99, 100]. Khalili et al., [101] exposed that treatment with an extract of C. sativus (i.e., 30 mg/kg) for 21 days might considerably improve cognition impairments facilitated by intracerebroventricular streptozotocin injection in rats. In fact, crocin (30 mg/kg) has been revealed to have an antagonizing role on the streptozotocin-induced cognitive deficits in rats and exerted higher correct choices and lower errors in Y-maze than vehicle-treated streptozotocin-injected rats [102]. In a different study, Geromichalos et al. [103] exhibited moderate (i.e., up to 30%) inhibitory activity exerted by saffron extract on AChE and inhibited breakdown of ACh which is the key therapeutic approach for Alzheimer’s.
The active constituents of C. sativus, at 1–10 μM was found to defend HT22 cells against neuronal cell death mediated by Aβ1-42 and reduced production of OS increased by Aβ1-42. Moreover, these findings revealed that crocetin has powerful neuroprotective effect against cytotoxicity triggered by Aβ1-42 in hippocampal cells by weakening OS, which ultimately recommends that crocetin might offer a beneficial therapeutic measure against disorders which are linked to Aβ [102]. Table 5 and Table 6 show the neuroprotective, anti-Alzheimer’s, cognitive-enhancing, and antioxidant effects of the C. sativus extracts and its bioactive constituents.
Anti-Alzheimer’s Effects in Human Studies
Akhondzadeh et al. [118] reported that when saffron at a dose of 30 mg/day (i.e., 15 mg twice daily) was administered in subjects of 55 years and older, it was found that saffron’s effectiveness was equivalent to donepezil to treat mild-to-moderate Alzheimer’s (Table 7). Furthermore, in the case of side effects, the extent of an extract of saffron was identical to those of donepezil apart from vomiting, which is more commonly observed in the donepezil-treated group [118]. Another study was conducted by giving saffron for 16 weeks to 46 patients who had mild to the moderate AD and it was seen that in comparison with placebo, cognitive functions were noticeably better in the saffron treated group [118]. Recently, Farokhnia et al. [119] in a double-blind randomized clinical trial reported that 1-year administration of saffron extract in the form of capsules exhibited to be comparable with memantine (i.e., a standard drug that block NMDA receptors) in decreasing cognitive deficits in patients with moderate to the severe AD. In Fig. 3, the possible mechanism of action for anti-Alzheimer’s potential of C. sativus is displayed.

Possible mechanism of action for anti-Alzheimer’s action of Crocus sativus extracts. CSE enhances Aβ clearance across the BBB and improves the tightness of the BBB limiting entry of unwanted large molecules, as well as exerts an anti-inflammatory effect whereby it reduces astrocytes activation and brain interleukin-1 beta [120]. CSE, Crocus sativus extracts; Aβ, amyloid beta; BBB, blood-brain barrier
Terminalia chebula
Anticholinesterase, Cognition-Enhancing, and Anti-Alzheimer’s Effects
In many studies, it has been reported that Terminalia chebula (T. chebula) has anticholinesterase properties [121,122,123,124,125,126,127]. Sancheti et al. [122] revealed that methanolic crude extract of the T. chebula fruits with a concentration of 5 mg/mL inhibited AChE and butyrylcholinesterase (BChE) about 89% and 95%, respectively. The researchers extracted 1,2,3,4,6-penta-O-galloyl-β-D-glucose with a gallotannin structure from T. chebula by chromatographic methods, and they showed it to be the most potent AChE and BChE inhibitor. Then again, concentration-mediated inhibitory activity on AChE and BChE with IC50 values of 29.9 ± 0.3 μM and 27.6 ± 0.2 μM successively was revealed when a study with bioassay of 1,2,3,4,6-penta-O-galloyl-β-D-glucose was conducted. Instead, for deducing the inhibitory potential of 1,2,3,4,6-penta-O-galloyl-β-D-glucose, a thin-layer chromatography assay was carried out with tacrine as a positive control, and the AChE inhibitory activity was visualized clearly in the assay [122]. In addition, the inhibitory potential of methanolic fruit extracts of T. chebula on AChE, the ethyl acetate fraction showed relatively remarkable AChE inhibitory activity. Sulaiman et al. [128] revealed that fraction of ethyl acetate with doses of 1, 5, 15, and 25 mg/mL inhibited AChE by 29.36%, 32.44%, 45.82%, and 62.32%, respectively. This fraction was more effective than the fraction of chloroform and methanol in AChE inhibition [128]. In a similar study, Vinutha et al. [121] used the methanolic and aqueous extracts of the T. chebula fruit and the outcome showed that IC50 values of the T. chebula aqueous extract (i.e., minimum inhibition 12.45%) are more potent than the methanolic extract (i.e., minimum inhibition 1.21%).
The effect of gallotannins on AChE inhibition, tannic acid as an active component of T. chebula, has a potential inhibition effect on AChE. Then again, the competitive inhibition of AChE by tannic acid was showed by Upadhyay and Singh [124]. This study focused on the effects of tannic acid from the T. chebula fruit. In the nervous tissue of Lymnaea acuminate, in vivo treatment with 80% of LC50 of tannic acid during 96 h caused a remarkable reduction in AChE activity. Furthermore, when the tissue was exposed to 80% of LC50 of tannic acid for 96 h, optimum inhibition of AChE activity was detected, which suggests that inhibition of AChE was time- and dose-dependent in the 96 h process. In vitro treatment showed that tannic acid triggered dose-dependent AChE inhibition significantly; such that 0.04 mM tannic acid inhibited the AChE activity to 37% of the control in the nervous tissue of Lymnaea acuminate [124]. In addition, another study on AChE activity showed that tannic acid completely neutralized the AChE activity in Naja kaouthia venom [129]. Instead, when administered orally, Parle and Vasudevan [125] in a study showed that the mixture of medicinal plants containing T. chebula with doses of 50, 100, and 200 mg/kg for 15 days can reduce the AChE activity in the brain of young and aged mice. PAT and maze tests were done on day 16 and the results exhibited a noteworthy dose-dependent inhibition in transfer latency and a marked increase in step-down latency tests, indicating significant memory improvement [125]. In a different study, Dhivya et al. [127] revealed that AChE inhibitory activity of various plant species containing T. chebula was reported to have AChE inhibition of 41.06 ± 5.6% (0.1 mg/mL).
In a similar study, AChE inhibition of the methanolic extract of T. chebula showed a moderate inhibition with an extract with IC50 value of 180 ± 14.6 μg/mL and the percentage of AChE inhibition was 41.06 ± 5.6 at 0.1 mg/mL [130]. In addition, the percentage of AChE inhibition in the extract of T. chebula was 13 [131]. Furthermore, in another research, T. chebula could inhibit 89% of AChE activity (5 mg/mL) and 95% of BChE activity (5 mg/mL) [122].
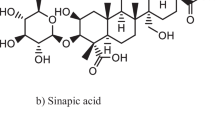
Interestingly, ellagic acid and gallic acid, as phenolic compounds in the T. chebula fruits, could also inhibit AChE. Nag and De [126] in their study stated that the inhibition in the effect of the methanolic extract of T. chebula (i.e., gallic acid: 0.25 μg, ellagic acid: 0.08 μg) has a linear relationship with increasing dosage (i.e., IC50: 10.96). In mice, Rao et al. [132] examined the learning- and memory-enhancing properties of different doses of T. chebula ethanolic extract (100, 200, and 400 mg/kg). The results suggested, when administered acutely, T. chebula ethanolic extracts also improve the process of learning and memory retrieval capacity in an inverse dose-dependent manner in mice. In Table 8, the neuroprotective, cognitive-enhancing, and antioxidant effects of the bioactive compounds of T. chebula are given.
As stated earlier T. chebula contains copious biochemicals and has a wide range of medicinal properties. The high content of phenolic constituents displays strong antioxidant and neuroprotective effects. The possible mechanism of action for anti-Alzheimer’s potential of T. chebula is specified in Fig. 4.

Possible mechanism of action for anti-Alzheimer’s action of Terminalia chebula extracts. Binding of TNF-α to TNFR causes activation of MyD88 and TRAF, thus activating MAPK. This activated MAPK finally activates NF-kB and translocate to the nucleus and binds target DNA to produce inflammation. TCE by blocking few steps of aforementioned pathway exerts anti-inflammatory effects as well as increases antioxidant enzymes, and blocks AChE and mTOR pathways [148, 149]. TCE, Terminalia chebula extracts; TNF-α, tumor necrosis factor alpha; TNFR, tumor necrosis factor receptor; MyD88, myeloid differentiation primary response protein 88; TRAF, tumor necrosis factor associated factors; MAPK, mitogen-activated protein kinases; NF-kB, nuclear factor kappa-light-chain-enhancer of activated B cells
Withania somnifera
Anticholinesterase, Memory-Enhancing, and Anti-Alzheimer’s Effects
In rats, the influence of withaferin and sitoindosides VII-X isolated from the aqueous methanol extract of roots of cultivated varieties of Withania somnifera (W. somnifera) were evaluated on glutamatergic, cholinergic, and GABAergic receptors of the brain. The compounds mediated slight enhancement of AChE activity was observed in the globus pallidus and lateral septum, and reduced AChE activity was observed in the vertical diagonal band [150]. Indeed, the observed changes were found associated with the improved binding with the M1-muscarinic receptor in lateral and medial septum as well as in frontal cortices. Then again, the increase in binding sites of M2- muscarinic receptor was noticed in many cortical regions, such as frontal, parietal, cingulate and retrosplenial cortex. These findings recommend that the compounds favorably affect the events that take place in the cortical and basal forebrain cholinergic-signal transduction cascade. Increased cortical muscarinic ACh receptor capacity induced by the drug could somewhat explain the memory improving and cognition-enhancing potential of W. somnifera extracts in animals and also in humans [150].
In an investigation by Zhao et al. [151] reported that withanoside IV initiated outgrowth of neurite in the cultured cortical neurons of rat. Oral intake of withanoside IV fundamentally enhanced memory shortages in Aβ-injected mice and forestalled the loss of dendrites, axons, as well as neural connections. On the other hand, sominone was recognized as the fundamental metabolite following oral administration of withanoside IV. Then, sominone initiated dendritic and axonal recovery and synaptic recreation altogether cultured cortical neurons of rat impaired by Aβ. Alternatively, withanoside IV may also enhance neuronal abnormality present in AD and sominone is the active principle after metabolism. In a different study employing the EPM test revealed that animals, which were treated with reserpine, additionally demonstrated poor retaining of memory. Ceaseless W. somnifera intake markedly turned reserpine-instigated retaining deficiencies [152]. In various examinations with W. somnifera root extricate enhanced retention in the PAT. W. somnifera likewise switched the scopolamine-instigated interruption of acquisition and retention and constricted the amnesia created by intense treatment with electroconvulsive shock, promptly after training. Perpetual therapy with electroconvulsive shock, for six progressive days at 24-h interims, disturbed memory consolidation on the 7th day. Day by day intake of W. somnifera for the period of 6 days altogether enhanced memory consolidation in mice which received a chronic electroconvulsive shock. W. somnifera, intake on day 7, likewise constricted the interruption of memory consolidation caused by chronic electroconvulsive shock. In EPM test, W. somnifera reversed the scopolamine-induced delay in transfer latency on the 1st day. These discoveries recommended that W. somnifera shows a nootropic-like impact in amnesic and naïve mice [153]. Table 9 and Table 10 denote the anti-Alzheimer’s, neuroprotective, antioxidant, and memory-enhancing activities of the extracts of W. somnifera and its bioactive constituents.
Anti-Alzheimer’s Effects in Human Studies
In a double-blind clinical trial, W. somnifera was studied for its anti-aging properties. At a dosage of 3 g daily for 1 year, a group of 101 healthy males, 50–59 years old was given the herb. The subjects experienced a noteworthy improvement in seated stature, hair melanin, red blood cell count, and hemoglobin. Moreover, nail calcium was preserved and there was a decrease in the serum cholesterol level. Instead, in case of sexual performance, 70% of the study subjects reported enhancement [163]. In adults with MCI, the safety and efficacy of W. somnifera in improving cognitive functioning and memory were evaluated by Choudhary et al., [164] in a double-blind, randomized, placebo-controlled, prospective study. Moreover, for 8 weeks, subjects were treated with either placebo or root extract of W. somnifera (300 mg 2 times a day). Interestingly, following 8 weeks of experiment, in case of both general and immediate memory, substantial enhancements were confirmed in the group which was treated with W. somnifera, as compared to the placebo group. This was demonstrated by Wechsler Memory Scale III subtest scores for family pictures II, verbal paired associates II, logical memory II, faces II, family pictures I, faces I, verbal paired associates I, and logical memory I. Considerably improvement in information-processing speed, sustained attention, and executive function was demonstrated by the treatment group, as specified by scores on the Mackworth Clock test, Trail-Making test part A, Wisconsin Card Sort test, and the Eriksen Flanker task [164]. Henceforth, W. somnifera could be effective in improving both general and immediate memory in people with MCI along with improvement in information processing speed, attention, and executive functions. A framework of the anti-Alzheimer’s, neuroprotective, memory-enhancing, and antioxidant effects of W. somnifera is mentioned in Fig. 5.

Asparagus racemosus
Neuroprotective and Anti-Alzheimer’s Effects
In Ayurvedic medicine, Asparagus racemosus (A. racemosus) is a quite well-known nervine tonic. Copiuos studies suggested neuroprotective effects of A. racemosus. Furthermore, in mice, Parihar and Hemnani [168] examined the effect of the methanolic extricate of A. racemosus roots against kainic acid-mediated hippocampal as well as striatal neuronal injury. Administration of kainic acid caused excitotoxic brain lesions in the anesthetized mice. The researchers reported injury of striatal and hippocampal regions of the brain as well as augmented lipid peroxidation, better protein carbonyl content, abated GPx and reduced GSH activity. The study suggested that reduced GSH is an imperative antioxidant that plays a pivotal role as a nucleophilic forager of deadly compounds and also acts as a substrate in the GSH peroxidase-facilitated damage of hydroperoxides that would then accrue to lethal levels in brain tissues [168]. In mice, A. racemosus extract was found to improve GPx and reduced activity of GSH and decline in lipid peroxidation as well as protein carbonyl contents. The researchers avowed that this extract of the plant shows the contribution of an antioxidant by reducing oxidative impairment induced by free radical [168].
Recently, in ovariectomized rats, Lalert et al. [169] studied the mechanisms and the effects of root extract of ethanol A. racemosus on the change of estrogen receptor (ER) subtypes and brain-derived neurotrophic factor. Moreover, a marked decrease in BDNF and a down-regulation of estrogen receptor-α, and -β in the hippocampus and frontal cortex were exhibited by the ovariectomized rats. In ovariectomized rats, it was also confirmed that root extract of A. racemosus and 17α-ethynylestradiol enhanced estrogen receptor-α, and -β and brain-derived neurotrophic factor in the hippocampus and frontal cortex. These findings collectively suggested that, in the ovariectomized rat, the increase of brain-derived neurotrophic factor and estrogen receptors up-regulation might be associated in the neuroprotective effects of root extract of ethanol A. racemosus. The neuroprotective, anticholinergic, and cognitive-enhancing, and antioxidant potential of the extracts of A. racemosus and its bioactive constituents are displayed in Table 11 and Table 12.
Anti-Alzheimer’s Effects in Human Studies
A. racemosus possesses multiple neuropharmacological activities to improve cognition and enhance memory (Fig. 6). The neuroprotective effects of root extract of A. racemosus were analyzed by Saxena et al. [170] in 16 clinical outpatients of a hospital for known stress-related disorders i.e., patients receiving treatment for cognitive changes, memory deficits. In addition, these were subjected to recall and memory retention test. As compared to stress group individuals, a substantial increase in test score was observed in the individuals whom were treated with extract. Interestingly, females were found to exhibit less effect of stress on short-term memory as compared to males. Therefore it can be hypothesized that, in cognition, estrogen has a significant contribution [183]. Furthermore, estrogen is well-known as a neuroprotective agent, a finding that driven many investigators to find a sidetrack or breakthrough for the protection from neurodegenerative diseases and cerebrovascular diseases [184, 185].

Asparagus racemosus extracts reduce oxidative stress by blocking membranal lipid peroxidation and protein carbonyl as well as improve cognitive functions by raising GPx, GSH BDNF, and ERs [168, 169]. GPx, glutathione peroxidase; GSH, reduced glutathione; BDNF, brain-derived neurotrophic factor; ERα, estrogen receptor alpha
Conclusion
The explored medicinal plants in this appraisal play a propitious role in the controlling AD. The utmost benefit of natural medicine has little adverse effects compared to modern medicine. Heretofore, Alzheimer’s is incurable albeit many compounds are under research. Merely a few of them show substantial denouements in clinical trials. The traditional medicine comprises appreciated info on medicines. The represented auspicious candidates could undertake an inquiry for the extensive further studies and clinical trials to detect lead compounds to abate AD pathogenesis.
Abbreviations
- AD:
-
Alzheimer disease
- Aβ:
-
amyloid β
- SPs:
-
senile plaques
- NFTs:
-
neurofibrillary tangles
- OS:
-
oxidative stress
- ACh:
-
acetylcholine
- AChE:
-
acetylcholinesterase
- BChE:
-
butyrylcholinesterase
References
Uddin MS, Haque A, Mamun AA et al (2016) Searching the linkage between high fat diet and Alzheimer′s disease: a debatable proof stand for ketogenic diet to alleviate symptoms of Alzheimer′s patient with APOE ε4 allele. J Neurol Neurophysiol 07:1–9. https://doi.org/10.4172/2155-9562.1000397
Mendiola-Precoma J, Berumen LC, Padilla K, Garcia-Alcocer G (2016) Therapies for prevention and treatment of Alzheimer’s disease. Biomed Res Int 2016:1–17. https://doi.org/10.1155/2016/2589276
Uddin MS, Mamun AA, Hossain MS et al (2016) Exploring the effect of Phyllanthus emblica L. on cognitive performance, brain antioxidant markers and acetylcholinesterase activity in rats: promising natural gift for the mitigation of Alzheimer’s disease. Ann Neurosci 23:218–229. https://doi.org/10.1159/000449482
Uddin MS, Mamun AA, Labu ZK, Hidalgo-Lanussa O, Barreto GE, Ashraf G (2018) Autophagic dysfunction in Alzheimer’s disease: cellular and molecular mechanistic approaches to halt Alzheimer’s pathogenesis. J Cell Physiol. https://doi.org/10.1002/jcp.27588
Khajehnejad M, Saatlou F, Mohammadzade H (2017) Alzheimer’s disease early diagnosis using manifold-based semi-supervised learning. Brain Sci 7:109. https://doi.org/10.3390/brainsci7080109
Alzheimer’s disease statistics—Alzheimer’s news today. https://alzheimersnewstoday.com/alzheimers-disease-statistics/. Accessed 12 Oct 2018
Ohnishi T, Matsuda H, Tabira T et al (2001) Changes in brain morphology in Alzheimer disease and normal aging: is Alzheimer disease an exaggerated aging process? AJNR Am J Neuroradiol 22:1680–1685
Uddin MS, Mamun AA, Takeda S et al (2018) Analyzing the chance of developing dementia among geriatric people: a cross-sectional pilot study in Bangladesh. Psychogeriatrics. https://doi.org/10.1111/psyg.12368
Uddin MS, Amran MS (2018) Handbook of research on critical examinations of neurodegenerative disorders. IGI Global, Pennsylvania
Murphy MP, LeVine H (2010) Alzheimer’s disease and the amyloid-β peptide. J Alzheimers Dis 19:311–323. https://doi.org/10.3233/JAD-2010-1221
O’Brien RJ, Wong PC (2011) Amyloid precursor protein processing and Alzheimer’s disease. Annu Rev Neurosci 34:185–204. https://doi.org/10.1146/annurev-neuro-061010-113613
Singh S, Kushwah AS, Singh R, Farswan M, Kaur R (2012) Current therapeutic strategy in Alzheimer’s disease. Eur Rev Med Pharmacol Sci 16:1651–1664
Parra MA, Abrahams S, Fabi K et al (2008) Short-term memory binding deficits in Alzheimer’s disease. Brain 132:1057–1066. https://doi.org/10.1093/brain/awp036
Mamum AA, Uddin MS, Wahid F et al (2016) Neurodefensive effect of Olea europaea L. in Alloxan-induced cognitive dysfunction and brain tissue oxidative stress in mice: incredible natural nootropic. J Neurol Neurosci 7:1–9. https://doi.org/10.21767/2171-6625.1000126
Tarawneh R, Holtzman DM (2012) The clinical problem of symptomatic Alzheimer disease and mild cognitive impairment. Cold Spring Harb Perspect Med 2:a006148–a006148. https://doi.org/10.1101/cshperspect.a006148
Burns A, Iliffe S (2009) Alzheimer’s disease. BMJ 338:b158–b158. https://doi.org/10.1136/bmj.b158
National Institute on Aging What are the signs of Alzheimer’s disease? https://www.nia.nih.gov/health/what-are-signs-alzheimers-disease. Accessed 13 Oct 2018
Uddin MS, Stachowiak A, Mamun AA et al (2018) Autophagy and Alzheimer’s disease: from molecular mechanisms to therapeutic implications. Front Aging Neurosci 10:1–18. https://doi.org/10.3389/fnagi.2018.00004
Ballard C, Gauthier S, Corbett A et al (2011) Alzheimer’s disease. Lancet 377:1019–1031. https://doi.org/10.1016/S0140-6736(10)61349-9
Uddin MS, Kabir MT, Mamun AA et al (2018) APOE and Alzheimer’s disease: evidence mounts that targeting APOE4 may combat Alzheimer’s pathogenesis. Mol Neurobiol. https://doi.org/10.1007/s12035-018-1237-z
Armstrong RA (2011) The pathogenesis of Alzheimer’s disease: a reevaluation of the “amyloid cascade hypothesis”. Int J Alzheimers Dis 2011:630865. https://doi.org/10.4061/2011/630865
Smith AD (2002) Imaging the progression of Alzheimer pathology through the brain. Proc Natl Acad Sci 99:4135–4137. https://doi.org/10.1073/pnas.082107399
Buchman AS, Bennett DA (2011) Loss of motor function in preclinical Alzheimer’s disease. Expert Rev Neurother 11:665–676. https://doi.org/10.1586/ern.11.57
Aliev G, Obrenovich ME, Reddy VP et al (2008) Antioxidant therapy in Alzheimer’s disease: theory and practice. Mini Rev Med Chem 8:1395–1406
Moneim AEA (2015) Oxidant/antioxidant imbalance and the risk of Alzheimer’s disease. Curr Alzheimer Res 12:335–349
Zhao Y, Zhao B (2013) Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxidative Med Cell Longev 2013:1–10. https://doi.org/10.1155/2013/316523
García-Ayllón M-S, Small DH, Avila J, Sáez-Valero J (2011) Revisiting the role of acetylcholinesterase in Alzheimer’s disease: cross-talk with P-tau and β-amyloid. Front Mol Neurosci 4:22. https://doi.org/10.3389/fnmol.2011.00022
DeFina PA, Moser RS, Glenn M et al (2013) Alzheimer’s disease clinical and research update for health care practitioners. J Aging Res 2013:1–9. https://doi.org/10.1155/2013/207178
Barbosa Filho JM, Medeiros KCP, Diniz M de FFM et al (2006) Natural products inhibitors of the enzyme acetylcholinesterase. Rev Bras Farmacogn 16:258–285. https://doi.org/10.1590/S0102-695X2006000200021
Rahman K (2007) Studies on free radicals, antioxidants, and co-factors. Clin Interv Aging 2:219–236
Bredie SJH, Jong MC (2012) No significant effect of ginkgo biloba special extract EGb 761 in the treatment of primary Raynaud phenomenon. J Cardiovasc Pharmacol 59:215–221. https://doi.org/10.1097/FJC.0b013e31823c0bed
ITIS Integrated Taxonomic Information System. https://www.itis.gov. Accessed 13 Oct 2018
Zahara K, Bibi Y, Tabassum S (2014) Clinical and therapeutic benefits of Centella asiatica. Pure Appl Biol 3:152–159
Al-Snafi AE (2016) Pharmacological importance of Clitoria ternatea—a review. IOSR J Pharm www.iosrphr.org 6:68–83
Saxena RB (2010) Botany, taxonomy and cytology of Crocus sativus series. Ayu 31:374–381. https://doi.org/10.4103/0974-8520.77153
Upadhyay A, Agrahari P, Singh DK (2014) A review on the pharmacological aspects of Terminalia chebula. Int J Pharmacol 10:289–298. https://doi.org/10.3923/ijp.2014.289.298
Narinderpal K, Junaid N, Raman B (2013) A review on pharmacological profile of Withania somnifera (Ashwagandha). Res Rev J Bot Sci 2:6–14
Sachan AK, Das DR, Dohare SL, Shuaib M (2012) Asparagus racemosus (Shatavari): an overview. Int J Pharm Chem Sci ISSN 2277 5005(1):588–592
Uddin MS, Nasrullah M, Hossain MS et al (2016) Evaluation of nootropic activity of Persicaria flaccida on cognitive performance, brain antioxidant markers and acetylcholinesterase activity in rats: implication for the management of Alzheimer’s disease. Am J Psychiatry Neurosci 4:26–37. https://doi.org/10.11648/j.ajpn.20160402.12
Brinkhaus B, Lindner M, Schuppan D, Hahn EG (2000) Chemical, pharmacological and clinical profile of the East Asian medical plant Centella asiatica. Phytomedicine 7:427–448
Veerendra Kumar MH, Gupta YK (2003) Effect of Centella asiatica on cognition and oxidative stress in an intracerebroventricular streptozotocin model of Alzheimer’s disease in rats. Clin Exp Pharmacol Physiol 30:336–342. https://doi.org/10.1046/j.1440-1681.2003.03842.x
Xu Y, Cao Z, Khan I, Luo Y (2008) Gotu Kola (Centella asiatica) extract enhances phosphorylation of cyclic AMP response element binding protein in neuroblastoma cells expressing amyloid beta peptide. J Alzheimers Dis 13:341–349
Jayashree G, Kurup Muraleedhara G, Sudarslal S, Jacob VB (2003) Anti-oxidant activity of Centella asiatica on lymphoma-bearing mice. Fitoterapia 74:431–434
Lee MK, Kim SR, Sung SH et al (2000) Asiatic acid derivatives protect cultured cortical neurons from glutamate-induced excitotoxicity. Res Commun Mol Pathol Pharmacol 108:75–86
Mook-Jung I, Shin JE, Yun SH et al (1999) Protective effects of asiaticoside derivatives against beta-amyloid neurotoxicity. J Neurosci Res 58:417–425
Kumar A, Dogra S, Prakash A (2009) Neuroprotective effects of Centella asiatica against Intracerebroventricular colchicine-induced cognitive impairment and oxidative stress. Int J Alzheimers Dis 2009:1–8. https://doi.org/10.4061/2009/972178
Bhumireddy Papireddy Venkata S, Kanniappan L, Bondha Ramakishnaiyah R et al (2014) Evaluation of the neuroprotective effects of Centella asiatica against scopolamine induced cognitive impairment in mice. Indian J Pharm Educ Res 48:31–34. https://doi.org/10.5530/ijper.48.4.5
Soumyanath A, Zhong Y-P, Henson E et al (2012) Centella asiatica extract improves behavioral deficits in a mouse model of Alzheimer’s disease: investigation of a possible mechanism of action. Int J Alzheimers Dis 2012:1–9. https://doi.org/10.1155/2012/381974
Nasir MN, Habsah M, Zamzuri I et al (2011) Effects of asiatic acid on passive and active avoidance task in male Spraque–Dawley rats. J Ethnopharmacol 134:203–209. https://doi.org/10.1016/j.jep.2010.12.010
Nasir MN, Abdullah J, Habsah M et al (2012) Inhibitory effect of asiatic acid on acetylcholinesterase, excitatory post synapticpotential and locomotor activity. Phytomedicine 19:311–316. https://doi.org/10.1016/j.phymed.2011.10.004
Zhang X, Wu J, Dou Y et al (2012) Asiatic acid protects primary neurons against C2-ceramide-induced apoptosis. Eur J Pharmacol 679:51–59. https://doi.org/10.1016/j.ejphar.2012.01.006
Xu C-L, Qu R, Zhang J, Li LF, Ma SP (2013) Neuroprotective effects of madecassoside in early stage of Parkinson’s disease induced by MPTP in rats. Fitoterapia 90:112–118. https://doi.org/10.1016/j.fitote.2013.07.009
Ling Y, Gong Q, Xiong X et al (2017) Protective effect of madecassoside on H2O2-induced oxidative stress and autophagy activation in human melanocytes. Oncotarget 8:51066–51075. https://doi.org/10.18632/oncotarget.17654
Xu C-L, Wang Q-Z, Sun L-M et al (2012) Asiaticoside: attenuation of neurotoxicity induced by MPTP in a rat model of parkinsonism via maintaining redox balance and up-regulating the ratio of Bcl-2/Bax. Pharmacol Biochem Behav 100:413–418. https://doi.org/10.1016/j.pbb.2011.09.014
Liu S, An J, Qi F, Yang L, Tian Z, Zhao MG (2014) Neuroprotective effects of Asiaticoside. Neural Regen Res 9:1275–1282. https://doi.org/10.4103/1673-5374.137574
Gupta R, Shukla RK, Chandravanshi LP et al (2017) Protective role of quercetin in cadmium-induced cholinergic dysfunctions in rat brain by modulating mitochondrial integrity and MAP kinase signaling. Mol Neurobiol 54:4560–4583. https://doi.org/10.1007/s12035-016-9950-y
Lei X, Chao H, Zhang Z et al (2015) Neuroprotective effects of quercetin in a mouse model of brain ischemic/reperfusion injury via anti-apoptotic mechanisms based on the Akt pathway. Mol Med Rep 12:3688–3696. https://doi.org/10.3892/mmr.2015.3857
Wang D-M, Li S-Q, Wu W-L et al (2014) Effects of long-term treatment with quercetin on cognition and mitochondrial function in a mouse model of Alzheimer’s disease. Neurochem Res 39:1533–1543. https://doi.org/10.1007/s11064-014-1343-x
Hong J-T, Yen J-H, Wang L, Lo YH, Chen ZT, Wu MJ (2009) Regulation of heme oxygenase-1 expression and MAPK pathways in response to kaempferol and rhamnocitrin in PC12 cells. Toxicol Appl Pharmacol 237:59–68. https://doi.org/10.1016/j.taap.2009.02.014
Jang S, Kelley KW, Johnson RW (2008) Luteolin reduces IL-6 production in microglia by inhibiting JNK phosphorylation and activation of AP-1. Proc Natl Acad Sci 105:7534–7539. https://doi.org/10.1073/pnas.0802865105
Lin TY, Lu CW, Wang SJ (2016) Luteolin protects the hippocampus against neuron impairments induced by kainic acid in rats. Neurotoxicology 55:48–57. https://doi.org/10.1016/j.neuro.2016.05.008
Prakash A, Shur B, Kumar A (2013) Naringin protects memory impairment and mitochondrial oxidative damage against aluminum-induced neurotoxicity in rats. Int J Neurosci 123:636–645. https://doi.org/10.3109/00207454.2013.785542
Huang Y-N, Wang J-Y, Lee C-T et al (2012) l-Ascorbate attenuates methamphetamine neurotoxicity through enhancing the induction of endogenous heme oxygenase-1. Toxicol Appl Pharmacol 265:241–252. https://doi.org/10.1016/j.taap.2012.08.036
Cherdyntseva N, Ivanova A, Ivanov V et al (2013) Ascorbic acid glucoside reduces neurotoxicity and glutathione depletion in mouse brain induced by nitrotriazole radiosensitazer. J Cancer Res Ther 9:364–369. https://doi.org/10.4103/0973-1482.119303
Li Y, Shi W, Li Y et al (2008) Neuroprotective effects of chlorogenic acid against apoptosis of PC12 cells induced by methylmercury. Environ Toxicol Pharmacol 26:13–21. https://doi.org/10.1016/j.etap.2007.12.008
Lee C-W, Won T-J, Kim H-R et al (2011) Protective effect of chlorogenic acid against Aβ-induced neurotoxicity. Biomol Ther 19:181–186. https://doi.org/10.4062/biomolther.2011.19.2.181
Zhao L, Wang J, Wang Y, Fa X (2013) Apigenin attenuates copper-mediated β-amyloid neurotoxicity through antioxidation, mitochondrion protection and MAPK signal inactivation in an AD cell model. Brain Res 1492:33–45. https://doi.org/10.1016/j.brainres.2012.11.019
Balez R, Steiner N, Engel M et al (2016) Neuroprotective effects of apigenin against inflammation, neuronal excitability and apoptosis in an induced pluripotent stem cell model of Alzheimer’s disease. Sci Rep 6:31450. https://doi.org/10.1038/srep31450
Wattanathorn J, Mator L, Muchimapura S et al (2008) Positive modulation of cognition and mood in the healthy elderly volunteer following the administration of Centella asiatica. J Ethnopharmacol 116:325–332. https://doi.org/10.1016/j.jep.2007.11.038
Tiwari S, Singh S, Patwardhan K et al (2008) Effect of Centella asiatica on mild cognitive impairment (mci) and other common age-related clinical problems. Dig J Nanomater Biostructures 3:215–220
Rai KS, Murthy KD, Karanth KS, Rao MS (2001) Clitoria ternatea (Linn) root extract treatment during growth spurt period enhances learning and memory in rats. Indian J Physiol Pharmacol 45:305–313
Taranalli AD, Cheeramkuzhy TC (2000) Influence of Clitoria ternatea extracts on memory and central cholinergic activity in rats. Pharm Biol 38:51–56. https://doi.org/10.1076/1388-0209(200001)3811-BFT051
Rai KS, Murthy KD, Karanth KS et al (2002) Clitoria ternatea root extract enhances acetylcholine content in rat hippocampus. Fitoterapia 73:685–689
Mukherjee PK, Kumar V, Houghton PJ (2007) Screening of Indian medicinal plants for acetylcholinesterase inhibitory activity. Phyther Res 21:1142–1145. https://doi.org/10.1002/ptr.2224
Ng TB, Wang HX, Liu F, Xia LX (2004) Plants beneficial to the aging brain. Neuroembryology Aging 3:136–141. https://doi.org/10.1159/000094576
Rai KS, Dilip Murthy K, Karanth KS, Rao MS (2001) Clitoria ternatea (linn) root extract treatment during growth spurt period enhances learning and memory in rats. Indian J Physiol Pharmacol 45(3):305–313
Rai KS, Murthy KD, Rao MS, Karanth KS (2005) Altered dendritic arborization of amygdala neurons in young adult rats orally intubated withClitorea ternatea aqueous root extract. Phyther Res 19:592–598. https://doi.org/10.1002/ptr.1657
Talpate KA, Bhosale UA, Zambare MR, Somani RS (2014) Neuroprotective and nootropic activity of Clitorea ternatea Linn. (Fabaceae) leaves on diabetes induced cognitive decline in experimental animals. J Pharm Bioallied Sci 6:48–55. https://doi.org/10.4103/0975-7406.124317
Rajashree R, Patil R, Khlokute SD, Goudar SS (2017) Effect of Salacia reticulata W. and Clitoria ternatea L. on the cognitive and behavioral changes in the streptozotocin-induced young diabetic rats. J Basic Clin Physiol Pharmacol 28:107–114. https://doi.org/10.1515/jbcpp-2016-0024
Ravishankar K, Parvathi M (2012) Study of protective effect of ethanolic root extract of Clitoria ternatea against stress induced amnesia. J Pharm Res 5:2763–2766
Kulkarni C, Pattanshetty JR, Amruthraj G (1988) Effect of alcoholic extract of Clitoria ternatea Linn. on central nervous system in rodents. Indian J Exp Biol 26:957–960
Jain NN, Ohal CC, Shroff SK et al (2003) Clitoria ternatea and the CNS. Pharmacol Biochem Behav 75:529–536
Yao X-Y, Bai Q (2017) Taraxerol protects the human hepatic L02 cells from hydrogen peroxide-induced apoptosis. Bangladesh J Pharmacol 12:20. https://doi.org/10.3329/bjp.v12i2.30985
Berté TE, Dalmagro AP, Zimath PL et al (2018) Taraxerol as a possible therapeutic agent on memory impairments and Alzheimer’s disease: effects against scopolamine and streptozotocin-induced cognitive dysfunctions. Steroids 132:5–11. https://doi.org/10.1016/J.STEROIDS.2018.01.002
Kiliç I, Yeşiloğlu Y (2013) Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim Acta A Mol Biomol Spectrosc 115:719–724. https://doi.org/10.1016/j.saa.2013.06.110
Guven M, Aras AB, Akman T, Sen HM, Ozkan A, Salis O, Sehitoglu I, Kalkan Y et al (2015) Neuroprotective effect of p-coumaric acid in rat model of embolic cerebral ischemia. Iran J Basic Med Sci 18:356–363
Rao N, Mittal S, Menghani E (2013) Antioxidant potential and validation of bioactive Β-Sitosterol in Eulophia campestris wall. Adv Biores 4:136–142
Ayaz M, Junaid M, Ullah F et al (2017) Anti-Alzheimer’s studies on β-Sitosterol isolated from Polygonum hydropiper L. Front Pharmacol 8:697. https://doi.org/10.3389/fphar.2017.00697
Tzeng C-W, Yen F-L, Wu T-H et al (2011) Enhancement of dissolution and antioxidant activity of Kaempferol using a nanoparticle engineering process. J Agric Food Chem 59:5073–5080. https://doi.org/10.1021/jf200354y
Kähkönen MP, Heinonen M (2003) Antioxidant activity of anthocyanins and their aglycons. J Agric Food Chem 51:628–633. https://doi.org/10.1021/JF025551I
Pojer E, Mattivi F, Johnson D, Stockley CS (2013) The case for anthocyanin consumption to promote human health: a review. Compr Rev Food Sci Food Saf 12:483–508. https://doi.org/10.1111/1541-4337.12024
Mo EK, Han BH, Kim SM et al (2012) Identification of d-friedoolean-13-en-3-one (Taraxerone) as an antioxidant compound from sedom (Sedum sarmentosum). Food Sci Biotechnol 21:485–489. https://doi.org/10.1007/s10068-012-0061-x
Estévez L, Mosquera RA (2008) Molecular structure and antioxidant properties of delphinidin. J Phys Chem A 112:10614–10623. https://doi.org/10.1021/jp8043237
Boominathan R, Parimaladevi B, Mandal SC (2003) Studies on Neuropharmacological effects of Clitoria ternatea Linn. Root extract in rats and mice. Nat Prod Sci 9:260–263
Malik J, Karan M, Vasisht K (2011) Nootropic, anxiolytic and CNS-depressant studies on different plant sources of shankhpushpi. Pharm Biol 49:1234–1242. https://doi.org/10.3109/13880209.2011.584539
Chusak C, Thilavech T, Henry CJ, Adisakwattana S (2018) Acute effect of Clitoria ternatea flower beverage on glycemic response and antioxidant capacity in healthy subjects: a randomized crossover trial. BMC Complement Altern Med 18:6. https://doi.org/10.1186/s12906-017-2075-7
Nair V, Bang WY, Schreckinger E et al (2015) Protective role of ternatin anthocyanins and quercetin glycosides from butterfly pea (Clitoria ternatea Leguminosae) blue flower petals against lipopolysaccharide (LPS)-induced inflammation in macrophage cells. J Agric Food Chem 63:6355–6365. https://doi.org/10.1021/acs.jafc.5b00928
Papandreou MA, Kanakis CD, Polissiou MG et al (2006) Inhibitory activity on amyloid-β aggregation and antioxidant properties of Crocus sativus stigmas extract and its crocin constituents. J Agric Food Chem 54:8762–8768. https://doi.org/10.1021/jf061932a
Lannert H, Hoyer S (1998) Intracerebroventricular administration of streptozotocin causes long-term diminutions in learning and memory abilities and in cerebral energy metabolism in adult rats. Behav Neurosci 112:1199–1208
Labak M, Foniok T, Kirk D, Rushforth D, Tomanek B, Jasiński A, Grieb P (2010) Metabolic changes in rat brain following intracerebroventricular injections of streptozotocin: a model of sporadic Alzheimer’s disease. Acta Neurochir Suppl 106:177–181
Khalili M, Kiasalari Z, Rahmati B, Narenjkar J (2010) Behavioral and histological analysis of Crocus sativus effect in intracerebroventricular streptozotocin model of Alzheimer disease in rats. Iran J Pathol 5:27–33
Khalili M, Hamzeh F (2010) Effects of active constituents of Crocus sativus L., crocin on streptozocin-induced model of sporadic Alzheimer’s disease in male rats. Iran Biomed J 14:59–65
Geromichalos GD, Lamari FN, Papandreou MA et al (2012) Saffron as a source of novel acetylcholinesterase inhibitors: molecular docking and in vitro enzymatic studies. J Agric Food Chem 60:6131–6138. https://doi.org/10.1021/jf300589c
Zhang Y, Shoyama Y, Sugiura M, Saito H (1994) Effects of Crocus sativus L. on the ethanol-induced impairment of passive avoidance performances in mice. Biol Pharm Bull 17:217–221
Pitsikas N, Sakellaridis N (2006) Crocus sativus L. extracts antagonize memory impairments in different behavioural tasks in the rat. Behav Brain Res 173:112–115. https://doi.org/10.1016/j.bbr.2006.06.005
Papandreou MA, Tsachaki M, Efthimiopoulos S et al (2011) Memory enhancing effects of saffron in aged mice are correlated with antioxidant protection. Behav Brain Res 219:197–204. https://doi.org/10.1016/j.bbr.2011.01.007
Ghadrdoost B, Vafaei AA, Rashidy-Pour A et al (2011) Protective effects of saffron extract and its active constituent crocin against oxidative stress and spatial learning and memory deficits induced by chronic stress in rats. Eur J Pharmacol 667:222–229. https://doi.org/10.1016/j.ejphar.2011.05.012
Linardaki ZI, Orkoula MG, Kokkosis AG et al (2013) Investigation of the neuroprotective action of saffron (Crocus sativus L.) in aluminum-exposed adult mice through behavioral and neurobiochemical assessment. Food Chem Toxicol 52:163–170. https://doi.org/10.1016/j.fct.2012.11.016
Hosseinzadeh H, Sadeghnia HR, Ghaeni FA et al (2011) Effects of saffron (Crocus sativus L.) and its active constituent, crocin, on recognition and spatial memory after chronic cerebral hypoperfusion in rats. Phyther Res 26:n/a-n/a. https://doi.org/10.1002/ptr.3566
Escribano J, Alonso GL, Coca-Prados M, Fernandez JA (1996) Crocin, safranal and picrocrocin from saffron (Crocus sativus L.) inhibit the growth of human cancer cells in vitro. Cancer Lett 100:23–30
Farkhondeh T, Samarghandian S, Shaterzadeh Yazdi H, Samini F (2018) The protective effects of crocin in the management of neurodegenerative diseases: a review. Am J Neurodegener Dis 7:1–10
Heidari S, Mehri S, Hosseinzadeh H (2017) Memory enhancement and protective effects of crocin against D-galactose aging model in the hippocampus of Wistar rats. Mashhad Univ Med Sci 20:1250–1259. https://doi.org/10.22038/IJBMS.2017.9541
Mohamadpour AH, Ayati Z, Parizadeh M-R et al (2013) Safety evaluation of crocin (a constituent of saffron) tablets in healthy volunteers. Iran J Basic Med Sci 16:39–46
Yoshino Y, Ishisaka M, Umigai N et al (2014) Crocetin prevents amyloid β1-42-induced cell death in murine hippocampal cells. Pharmacol Pharm 05:37–42. https://doi.org/10.4236/pp.2014.51007
Power R, Coen RF, Beatty S et al (2018) Supplemental retinal carotenoids enhance memory in healthy individuals with low levels of macular pigment in a randomized, double-blind, placebo-controlled clinical trial. J Alzheimers Dis 61:947–961. https://doi.org/10.3233/JAD-170713
Xue C, Rosen R, Jordan A, Hu D-N (2015) Management of ocular diseases using lutein and zeaxanthin: what have we learned from experimental animal studies? J Ophthalmol 2015:1–11. https://doi.org/10.1155/2015/523027
Wang Z, Fan J, Wang J et al (2016) Protective effect of lycopene on high-fat diet-induced cognitive impairment in rats. Neurosci Lett 627:185–191. https://doi.org/10.1016/j.neulet.2016.05.014
Akhondzadeh S, Shafiee Sabet M, Harirchian MH et al (2010) A 22-week, multicenter, randomized, double-blind controlled trial of Crocus sativus in the treatment of mild-to-moderate Alzheimer’s disease. Psychopharmacology 207:637–643. https://doi.org/10.1007/s00213-009-1706-1
Farokhnia M, Shafiee Sabet M, Iranpour N et al (2014) Comparing the efficacy and safety of Crocus sativus L. with memantine in patients with moderate to severe Alzheimer’s disease: a double-blind randomized clinical trial. Hum Psychopharmacol Clin Exp 29:351–359. https://doi.org/10.1002/hup.2412
Batarseh YS, Bharate SS, Kumar V et al (2017) Crocus sativus extract tightens the blood-brain barrier, reduces amyloid β load and related toxicity in 5XFAD mice. ACS Chem Neurosci 8:1756–1766. https://doi.org/10.1021/acschemneuro.7b00101
Vinutha B, Prashanth D, Salma K et al (2007) Screening of selected Indian medicinal plants for acetylcholinesterase inhibitory activity. J Ethnopharmacol 109:359–363. https://doi.org/10.1016/j.jep.2006.06.014
Sancheti S, Sancheti S, Um B-H, Seo S-Y (2010) 1,2,3,4,6-penta-O-galloyl-β-d-glucose: a cholinesterase inhibitor from Terminalia chebula. South African J Bot 76:285–288. https://doi.org/10.1016/J.SAJB.2009.11.006
Murray AP, Faraoni MB, Castro MJ et al (2013) Natural AChE inhibitors from plants and their contribution to Alzheimer’s disease therapy. Curr Neuropharmacol 11:388–413. https://doi.org/10.2174/1570159X11311040004
Upadhyay A, Singh DK (2011) Inhibition kinetics of certain enzymes in the nervous tissue of vector snail Lymnaea acuminata by active molluscicidal components of Sapindus mukorossi and Terminalia chebula. Chemosphere 85:1095–1100. https://doi.org/10.1016/j.chemosphere.2011.07.041
Parle M, Vasudevan M (2007) Memory enhancing activity of Abana : an Indian ayurvedic poly-herbal formulation
Nag G, De B (2011) Acetylcholinesterase inhibitory activity of Terminalia chebula, terminalia bellerica and Emblica officinalis and some phenolic compounds. Int J Pharm Pharm Sci 3:121–124
Dhivya P, Sobiya M, Selvamani P, Latha S (2014) An approach to Alzheimer’s disease treatment with cholinesterase inhibitory activity from various plant species. Int J PharmTech Res 6:1450–1467
Sulaiman CT, Sadashiva CT, George S, Balachandran I (2012) Journal of tropical medicinal plants. Tropical Botanics Sdn Bhd
Surveswaran S, Cai Y-Z, Xing J et al (2010) Antioxidant properties and principal phenolic phytochemicals of Indian medicinal plants from Asclepiadoideae and Periplocoideae. Nat Prod Res 24:206–221. https://doi.org/10.1080/14786410802228827
Mathew M, Subramanian S (2014) In vitro screening for anti-cholinesterase and antioxidant activity of methanolic extracts of ayurvedic medicinal plants used for cognitive disorders. PLoS One 9:e86804. https://doi.org/10.1371/journal.pone.0086804
Ali SK, Hamed AR, Soltan MM, Hegazy UM, Elgorashi EE, El-Garf IA, Hussein AA (2013) In-vitro evaluation of selected Egyptian traditional herbal medicines for treatment of alzheimer disease. BMC Complement Altern Med 13:121. https://doi.org/10.1186/1472-6882-13-121
Rao N, Palaksha MN, Satish S, Ravishankar (2013) The effects of Ethanolic extract in dried fruits of Terminalia chebula on learning and memory in mice. Asian J Biomed Pharm Sci 3(20):59–62
Surveswaran S, Cai Y-Z, Corke H, Sun M (2007) Systematic evaluation of natural phenolic antioxidants from 133 Indian medicinal plants. Food Chem 102:938–953. https://doi.org/10.1016/J.FOODCHEM.2006.06.033
Kim HJ, Kim J, Kang KS et al (2014) Neuroprotective effect of Chebulagic acid via autophagy induction in SH-SY5Y cells. Biomol Ther (Seoul) 22:275–281. https://doi.org/10.4062/biomolther.2014.068
Bhadoriya U, Sharma P, Solanki SS (2012) In vitro free radical scavenging activity of gallic acid isolated from Caesalpinia Decapetala wood. Asian Pacific J Trop Dis 2:S833–S836. https://doi.org/10.1016/S2222-1808(12)60274-6
Ibrahim AE, El Kareem RA, Sheir MA (2015) Elucidation of acrylamide genotoxicity and neurotoxicity and the protective role of gallic acid and green tea. J Forensic Toxicol Pharmacol 04. https://doi.org/10.4172/2325-9841.1000135
Kilic I, Yeşiloğlu Y, Bayrak Y (2014) Spectroscopic studies on the antioxidant activity of ellagic acid. Spectrochim Acta A Mol Biomol Spectrosc 130:447–452. https://doi.org/10.1016/j.saa.2014.04.052
Farbood Y, Sarkaki A, Dianat M et al (2015) Ellagic acid prevents cognitive and hippocampal long-term potentiation deficits and brain inflammation in rat with traumatic brain injury. Life Sci 124:120–127. https://doi.org/10.1016/j.lfs.2015.01.013
Zhao X, Sun H, Hou A et al (2005) Antioxidant properties of two gallotannins isolated from the leaves of Pistacia weinmannifolia. Biochim Biophys Acta Gen Subj 1725:103–110. https://doi.org/10.1016/j.bbagen.2005.04.015
Kujawski R, Kujawska M, Ożarowski M, Baraniak J, Laskowska H, Nowocień T, Borowska M, Szulc M et al (2016) Perspectives for gallotannins neuroprotective potential-current experimental evidences Comparison of extracts from root of Rhodiola rosea inhibitory action on EtOH tolerance development in rats View project Małgorzata Kujawska. J Med Sci 85:313–318. https://doi.org/10.20883/jms.2016.172
Kähkönen M, Kylli P, Ollilainen V, Salminen JP, Heinonen M (2012) Antioxidant activity of isolated Ellagitannins from red raspberries and cloudberries. J Agric Food Chem 60:1167–1174. https://doi.org/10.1021/jf203431g
Sowndhararajan K, Kim S (2017) Neuroprotective and cognitive enhancement potentials of Angelica gigas Nakai root: a review. Sci Pharm 85:21. https://doi.org/10.3390/scipharm85020021
Hussein RH, Khalifa FK (2014) The protective role of ellagitannins flavonoids pretreatment against N-nitrosodiethylamine induced-hepatocellular carcinoma. Saudi J Biol Sci 21:589–596. https://doi.org/10.1016/j.sjbs.2014.03.004
Lee H-S, Jung S-H, Yun B-S, Lee K-W (2007) Isolation of chebulic acid from Terminalia chebula Retz. and its antioxidant effect in isolated rat hepatocytes. Arch Toxicol 81:211–218. https://doi.org/10.1007/s00204-006-0139-4
Klika KD, Saleem A, Sinkkonen J et al (2004) The structural and conformational analyses and antioxidant activities of chebulinic acid and its thrice-hydrolyzed derivative, 2,4-chebuloyl-β-D-glucopyranoside, isolated from the fruit of Terminalia chebula. ARKIVOC 7:83–105
Romanová D, Vachálková A, Cipák L et al (2001) Study of antioxidant effect of apigenin, luteolin and quercetin by DNA protective method. Neoplasma 48:104–107
Aloqbi A, Omar U, Yousr M, Grace M, Lila MA, Howell N (2016) Antioxidant activity of pomegranate juice and Punicalagin. Antioxid Act Pomegranate Juice Punicalagin Nat Sci 8:235–246. https://doi.org/10.4236/ns.2016.86028
Shanmuganathan S, Angayarkanni N (2018) Chebulagic acid Chebulinic acid and Gallic acid, the active principles of Triphala, inhibit TNFα induced pro-angiogenic and pro-inflammatory activities in retinal capillary endothelial cells by inhibiting p38, ERK and NFkB phosphorylation. Vasc Pharmacol 108:23–35. https://doi.org/10.1016/J.VPH.2018.04.005
Lee Y, Byun HS, Seok JH et al (2016) Terminalia Chebula provides protection against dual modes of necroptotic and apoptotic cell death upon death receptor ligation. Sci Rep 6:25094. https://doi.org/10.1038/srep25094
Schliebs R, Liebmann A, Bhattacharya SK et al (1997) Systemic administration of defined extracts from Withania somnifera (Indian Ginseng) and Shilajit differentially affects cholinergic but not glutamatergic and GABAergic markers in rat brain. Neurochem Int 30:181–190
Zhao J, Nakamura N, Hattori M et al (2002) Withanolide derivatives from the roots of Withania somnifera and their neurite outgrowth activities. Chem Pharm Bull (Tokyo) 50:760–765
Naidu PS, Singh A, Kulkarni SK (2006) Effect of Withania somnifera root extract on reserpine-induced orofacial dyskinesia and cognitive dysfunction. Phyther Res 20:140–146. https://doi.org/10.1002/ptr.1823
Dhuley JN (2001) Nootropic-like effect of ashwagandha (Withania somnifera L.) in mice. Phytother Res 15:524–528
Kumar V, Dey A, Hadimani MB et al (2015) Chemistry and pharmacology of withania somnifera: An update. TANG [HUMANITAS Med] 5:1.1–1.13. https://doi.org/10.5667/tang.2014.0030
Tohda C, Joyashiki E (2009) Sominone enhances neurite outgrowth and spatial memory mediated by the neurotrophic factor receptor, RET. Br J Pharmacol 157:1427–1440. https://doi.org/10.1111/j.1476-5381.2009.00313.x
Kuboyama T, Tohda C, Komatsu K (2006) Withanoside IV and its active metabolite, sominone, attenuate Aβ(25-35)-induced neurodegeneration. Eur J Neurosci 23:1417–1426. https://doi.org/10.1111/j.1460-9568.2006.04664.x
Ostrovskaya RU, Gruden MA, Bobkova NA et al (2007) The nootropic and neuroprotective proline-containing dipeptide noopept restores spatial memory and increases immunoreactivity to amyloid in an Alzheimer’s disease model. J Psychopharmacol 21:611–619. https://doi.org/10.1177/0269881106071335
Li T, Wang L, Hu Q et al (2017) Neuroprotective roles of l-cysteine in attenuating early brain injury and improving synaptic density via the CBS/H2S pathway following subarachnoid hemorrhage in rats. Front Neurol 8:176. https://doi.org/10.3389/fneur.2017.00176
Batumalaie K, Amin MA, Murugan DD et al (2016) Withaferin A protects against palmitic acid-induced endothelial insulin resistance and dysfunction through suppression of oxidative stress and inflammation. Sci Rep 6:27236. https://doi.org/10.1038/srep27236
Murugan S, Ameesh M, Devaraja R et al (2015) A study on the efficacy of Withaferin-A in reducing oxidative stress against DEN induced hepatocellular carcinoma. IOSR J Dent Med Sci e-ISSN 14:23–31. https://doi.org/10.9790/0853-14922331
Kuboyama T, Tohda C, Komatsu K (2005) Neuritic regeneration and synaptic reconstruction induced by withanolide A. Br J Pharmacol 144:961–971. https://doi.org/10.1038/sj.bjp.0706122
Jayaprakasam B, Padmanabhan K, Nair MG (2010) Withanamides in Withania somnifera fruit protect PC-12 cells from Î2-amyloid responsible for Alzheimer’s disease. Phyther Res 24:859–863. https://doi.org/10.1002/ptr.3033
Bone K (1996) Clinical applications of Ayurvedic and Chinese herbs : monographs for the Western herbal practitioner. Phytotherapy Press
Choudhary D, Bhattacharyya S, Bose S (2017) Efficacy and safety of Ashwagandha (Withania somnifera (L.) Dunal ) root extract in improving memory and cognitive functions. J Diet Suppl 14:599–612. https://doi.org/10.1080/19390211.2017.1284970
Kurapati KRV, Atluri VSR, Samikkannu T, Nair MPN (2013) Ashwagandha (Withania somnifera) reverses β-Amyloid1-42 induced toxicity in human neuronal cells: implications in HIV-associated neurocognitive disorders (HAND). PLoS One 8:e77624. https://doi.org/10.1371/journal.pone.0077624
Bhattacharya A, Ghosal S, Bhattacharya SK (2001) Anti-oxidant effect of Withania somnifera glycowithanolides in chronic footshock stress-induced perturbations of oxidative free radical scavenging enzymes and lipid peroxidation in rat frontal cortex and striatum. J Ethnopharmacol 74:1–6
Afshari AR, Sadeghnia HR, Mollazadeh H (2016) A review on potential mechanisms of Terminalia chebula in Alzheimer’s disease. Adv Pharmacol Sci 2016:1–14. https://doi.org/10.1155/2016/8964849
Parihar MS, Hemnani T (2004) Experimental excitotoxicity provokes oxidative damage in mice brain and attenuation by extract of Asparagus racemosus. J Neural Transm 111:1–12. https://doi.org/10.1007/s00702-003-0069-8
Lalert L, Kruevaisayawan H, Amatyakul P et al (2018) Neuroprotective effect of Asparagus racemosus root extract via the enhancement of brain-derived neurotrophic factor and estrogen receptor in ovariectomized rats. J Ethnopharmacol 225:336–341. https://doi.org/10.1016/J.JEP.2018.07.014
Saxena G, Singh M, Meena P, Barber S, Sharma D, Shukla S, Bhatnagar M (2007) Neuroprotective effects of Asparagus racemosus Linn root extract: an experimental and clinical evidence. Ann Neurosci 14:57–63. https://doi.org/10.5214/156
Ahmad MP, Hussain A, Siddiqui HH et al (2015) Effect of methanolic extract of Asparagus racemosus Willd. on lipopolysaccharide induced-oxidative stress in rats. Pak J Pharm Sci 28:509–513
Sarbishegi M, Heidari Z, Mahmoudzadeh-Sagheb H et al (2016) Neuroprotective effects of Withania coagulans root extract on CA1 hippocampus following cerebral ischemia in rats. Avicenna J phytomedicine 6:399–409
Maheep B, Prahlad M, Satyendra B, Chetan J (2013) Neuroprotective response of the hippocampus region of the brain to Withania somnifera and Asparagus racemosus root extract: an in vitro study. J Med Plants Res 7:2259–2264. https://doi.org/10.5897/JMPR12.497
Sharma K, Bhatnagar M, Kulkarni SK (2010) Effect of Convolvulus pluricaulis Choisy and Asparagus racemosus Willd on learning and memory in young and old mice: a comparative evaluation. Indian J Exp Biol 48:479–485
Almutairi MM, Alanazi WA, Alshammari MA, Alotaibi MR, Alhoshani AR, al-Rejaie SS, Hafez MM, al-Shabanah OA (2017) Neuro-protective effect of rutin against cisplatin-induced neurotoxic rat model. BMC Complement Altern Med 17:472. https://doi.org/10.1186/s12906-017-1976-9
Sharma S, Ali A, Ali J et al (2013) Rutin: therapeutic potential and recent advances in drug delivery. Expert Opin Investig Drugs 22:1063–1079. https://doi.org/10.1517/13543784.2013.805744
Pany S, Pal A, Sahu PK (2014) Neuroprotective effect of quercetin in neurotoxicity induced rats: role of neuroinflammation in neurodegeneration. Asian J Pharm Clin Res 7:152–156
Sarkar S, Mukherjee A, Swarnakar S, Das N (2016) Nanocapsulated ascorbic acid in combating cerebral ischemia reperfusion- induced oxidative injury in rat brain. Curr Alzheimer Res 13:1363–1373
Dolu N, Khan A, Dokutan Ş (2015) Effect of vitamin E administration on learning of the young male rats. J Exp Neurosci 9:81–85. https://doi.org/10.4137/JEN.S29843
Zandi PP, Anthony JC, Khachaturian AS et al (2004) Reduced risk of Alzheimer disease in users of antioxidant vitamin supplements. Arch Neurol 61:82–88. https://doi.org/10.1001/archneur.61.1.82
Hu Y, Xia Z, Sun Q et al (2005) A new approach to the pharmacological regulation of memory: sarsasapogenin improves memory by elevating the low muscarinic acetylcholine receptor density in brains of memory-deficit rat models. Brain Res 1060:26–39. https://doi.org/10.1016/j.brainres.2005.08.019
Sarkaki A, Amani R, Badavi M et al (2008) Pre-treatment effect of different doses of soy isoflavones on spatial learning and memory in an ovariectomized animal model of Alzheimer’s disease. Pak J Biol Sci 11:1114–1119
Carlson LE, Sherwin BB (1999) Relationships among cortisol (CRT), dehydroepiandrosterone-sulfate (DHEAS), and memory in a longitudinal study of healthy elderly men and women. Neurobiol Aging 20:315–324
Dhandapani KM, Brann DW (2002) Protective effects of estrogen and selective estrogen receptor modulators in the brain. Biol Reprod 67:1379–1385
Alkayed NJ, Murphy SJ, Traystman RJ, Hurn PD, Miller VM (2000) Neuroprotective effects of female gonadal steroids in reproductively senescent female rats. Stroke 31:161–168
Acknowledgments
The authors are grateful to the Department of Pharmacy, Southeast University, Dhaka, Bangladesh.
Funding
The authors received no financial support for the research, authorship, and publication of this manuscript.
Author information
Authors and Affiliations
Contributions
This work was carried out in collaboration between all authors. MSU and GMA designed the study, wrote the protocol, and managed the analyses of the study. MSU, AAM, and MTK prepared the draft of the manuscript. MSU prepared the figures for the manuscript. MJ and BM participated in literature searches. GEB reviewed the scientific contents of the manuscript. All authors read and approved the final submitted version of the manuscript.
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Uddin, M.S., Al Mamun, A., Kabir, M.T. et al. Nootropic and Anti-Alzheimer’s Actions of Medicinal Plants: Molecular Insight into Therapeutic Potential to Alleviate Alzheimer’s Neuropathology. Mol Neurobiol 56, 4925–4944 (2019). https://doi.org/10.1007/s12035-018-1420-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12035-018-1420-2