
Abstract
Ovarian reserve is a term used to estimate the total number of immature follicles present in the ovaries. Between birth and menopause, there is a progressive decrease in the number of ovarian follicles. Ovarian aging is a continuous physiological phenomenon, with menopause being the clinical mark of the end of ovarian function. Genetics, measured as family history for age at the onset of menopause, is the main determinant. However, physical activity, diet, and lifestyle are important factors that can influence the age of menopause. The low estrogen levels after natural or premature menopause increased the risk for several diseases, resulting in increased mortality risk. Besides that, the decreasing ovarian reserve is associated to reduced fertility. In women with infertility undergoing in vitro fertilization, reduced markers of ovarian reserve, including antral follicular count and anti-Mullerian hormone, are the main indicators of reduced chances of becoming pregnant. Therefore, it becomes clear that the ovarian reserve has a central role in women’s life, affecting fertility early in life and overall health later in life. Based on this, the ideal strategy for delaying ovarian aging should have the following characteristics: (1) be initiated in the presence of good ovarian reserve; (2) maintained for a long period; (3) have an action on the dynamics of primordial follicles, controlling the rate of activation and atresia; and (4) safe use in pre-conception, pregnancy, and lactation. In this review, we therefore discuss some of these strategies and its feasibility for preventing a decline in the ovarian reserve.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ovaries are glands of the female reproductive system responsible for the development of the gamete (oocyte) and production of female sex hormones, which regulate reproductive and non-reproductive functions [1]. Anatomically, the ovary can be divided into two compartments, medulla (inner layer, composed of loose vascularized connective tissue) and cortex (outer layer containing the ovarian follicles) [1]. The functional unit of the ovary is the follicle, which is constituted by an oocyte surrounded, initially, only by granulosa cells and, later, also by theca cells. The ovarian follicle can be classified into different types (primordial, primary, secondary, antral, and pre-ovulatory) according to the degree of oocyte maturity and its histological structure [1, 2].
Ovarian reserve is a term used to estimate the total number of immature follicles present in the ovaries. In humans, the maximum number of ovarian follicles is reached during a woman’s intrauterine life, containing about 7 million follicles around the 20th week of gestation. Between birth, menarche, and menopause, there is a progressive decrease in the number of ovarian follicles, estimated for approximately 1 to 2 million, 300–400,000, and 1000 follicles, respectively [3]. The primordial follicles remain in a state of quiescence and may enter directly into atresia or start the maturation process (folliculogenesis), culminating in ovulation or follicular atresia during any stage. During a woman’s reproductive life, approximately 400 ovulations occur [1, 2, 4].
Folliculogenesis is a continuous process, divided into two stages, beginning with the activation of primordial follicles and ending with follicular rupture. Inhibitory and stimulatory intraovarian factors trigger the first stage (initial recruitment) of folliculogenesis, starting about 150 days before ovulation, independent of follicle-stimulating hormone (FSH) [2]. Activation of primordial follicles induces modifications of granulosa cells around the oocyte, which assume a cuboidal shape in a single layer (primary follicle) and later in multiple layers (secondary follicle). Then, granulosa cells continue proliferating and theca cells appear, forming the preantral and antral follicles, at which point the second stage begins (cyclic recruitment) of folliculogenesis, dependent on FSH [2].
Theca and granulosa cells are responsible for the synthesis and secretion of estrogen, the main ovarian hormone. The presence of estrogen receptors in other non-reproductive organs (brain, bone, liver, intestine, skin, and salivary glands) demonstrates the relevance of the ovary in the regulation of different body functions [1, 2, 5]. The duration of menacme, the period in which the ovary plays its physiological role in full, depends not only on the ovarian reserve, but also on the rate of activation of primordial follicles and follicular atresia. Numerous local or systemic mechanisms are responsible for ovarian aging, with reproductive and non-reproductive repercussions [6, 7]. Basically, ovarian aging is characterized by a decline in follicular quantity and quality, and changes in the synthesis of ovarian hormones, resulting in a drop in estrogen levels [2].
Progressive ovarian aging promotes the depletion of female gonadal function, initiating a period of transition from the reproductive to the non-reproductive phase in a woman’s life, known as the climacteric [2, 8, 9]. Therefore, a good understanding of folliculogenesis and understanding of the ovarian aging process are fundamental for anti-aging interventions to be proposed.
Lessons from menopause studies
Ovarian aging is a continuous physiological phenomenon, with menopause being the clinical mark of the end of ovarian function. Usually, menopause occurs between the ages of 45 and 55 years [10,11,12]. Numerous epidemiological studies evaluate conditions that can influence the age of menopause. Genetics, measured as family history for age at the onset of menopause, is the main determinant. Ethnicity, physical activity, diet, and habits are other significant factors that influence the age of menopause [11].
Current smoking, low socioeconomic status, low education, unemployment, early menarche, nulliparity, unilateral oophorectomy, vigorous physical activity, severe weight loss, vegetarian or high-carbohydrate diet, and high consumption of polyunsaturated fats are conditions that accelerate the onset of menopause [11, 13]. On the other hand, multiparity, first pregnancy at an advanced age, use of oral contraceptive pills, Japanese ethnicity, higher body mass index, moderate physical activity, and moderate alcohol consumption are associated with late menopause [11, 13].
Menopause-induced hypoestrogenism has short- and long-term consequences for women’s health. Hot flashes, night sweats, palpitations, headache, vaginal dryness, burning and genital irritation, dyspareunia, urinary urgency, dysuria, and recurrent urinary tract infection are short-term repercussions, while osteoporosis and cardiovascular and neurological diseases are long-term consequences [14]. The North American Menopause Society recommends hormone therapy as a supportive intervention to improve vasomotor symptoms and relieve genitourinary menopausal syndrome, as well as prevent bone loss and fractures, especially in women under 60 years of age and less than 10 years after menopause onset [8].
Several studies have evaluated ovarian aging markers with the potential to predict the onset of menopause, such as antral follicle count (AFC) and blood levels of inhibins A and B, follicle-stimulating hormone (FSH), estradiol, and anti-Mullerian hormone (AMH) [15,16,17,18,19,20]. AMH is produced by the granulosa cells of growing follicles, from primary follicle to small antral follicles. AMH is currently the most promising marker for predicting age at natural menopause [15, 18].
Pathological ovarian aging can anticipate the occurrence of menopause. The term “early menopause” is used to refer to the onset of menopause before age 45, while premature ovarian insufficiency (POI) is when the last menstrual period occurs before age 40 [21]. POI is a gynecological condition that affects approximately 1% of women. In about 90% of cases, the cause of POI is unknown (idiopathic POI), and specific diagnoses are genetic abnormalities (such as Turner syndrome, Fragile X syndrome, and Autosomal gene mutations), exposure to gonadotoxic agents (chemotherapy and radiotherapy), autoimmune disorders (such as lymphocytic oophoritis, thyroid disease, Addison’s disease, and celiac disease), viral infections, oophorectomy, and endometriosis [21,22,23].
The low estrogen levels detected in the POI have consequences for women’s health, potentially more serious than those observed in physiological menopause [24]. Malek et al. observed a 15% increase in the risk of all-cause mortality among women with early age at menopause [25]. Mortality was also higher among women with POI (34.7% vs. 19.3%, p < 0.001) in a long-term cohort of Chilean women followed for three decades, especially deaths from cardiovascular disease (12.0% vs. 5.1%; OR 2.55, 95% CI 1.21–5.39) [26]. Other studies have confirmed that women with POI have an increased risk of death from all causes, as well as a higher prevalence of cardiovascular disease, autoimmune conditions, osteoporosis, cognitive dysfunction, mood, and sexual disorders [24, 27,28,29,30,31].
Currently, there is no reliable intervention to reverse ovarian aging in women diagnosed with POI. Estrogen hormone therapy, with or without progesterone, is recommended to improve quality of life and reduce comorbidities. However, regular practice of moderate physical activity, healthy diet, maintenance of healthy habits, and early diagnosis and control of acute or chronic diseases are good ovarian antiaging strategies.
Lessons from reproductive medicine
Infertility in women is defined as the inability to establish pregnancy spontaneously after a period of 12 months with regular sexual intercourse; the period is reduced to 6 months when the woman is over 35 years old [21]. Disorders such as polycystic ovary syndrome, low ovarian reserve, and POI are responsible for up to 25% of female infertility cases [19, 21]. Interestingly, epidemiological studies suggest that there is an association between female reproductive aspects and life expectancy and risk of chronic diseases [32, 33].
The live birth rates of women undergoing infertility treatment by in vitro fertilization (IVF) are directly related to the ovarian response to controlled stimulation. The Bologna and POSEIDON (Patient-Oriented Strategies Encompassing IndividualizeD Oocyte Number) criteria are the two most used ways to define poor ovarian response (POR). Both are based on the woman’s age, ovarian reserve biomarkers, and number of oocytes recovered from previous ovarian stimulations [34,35,36]. According to the Bologna criteria, a woman with a POR must have at least two of the following conditions: advanced maternal age (> 40 years), a previous poor response to controlled stimulation (defined as ≤ 3 oocytes retrieved), or an abnormal ovarian reserve test (antral follicle count [AFC] < 5–7 follicles and/or AMH < 1.1 ng/mL) [34,35,36]. POSEIDON criteria divide patients into four groups based on a combination of factors: group 1—patients < 35 years old, presenting with adequate values of AFC (≥ 5 follicles) and/or AMH ≥ 1.2 ng/mL (subgroup 1a: < 4 oocytes retrieved and subgroup 1b: 4–9 oocytes retrieved, after standard ovarian stimulation); group 2—patients ≥ 35 years old, presenting with adequate values of AFC (≥ 5 follicles) and/or AMH ≥ 1.2 ng/mL (subgroup 2a: < 4 oocytes retrieved and subgroup 2b: 4–9 oocytes retrieved, after standard ovarian stimulation); group 3—patients < 35 years old, presenting with poor values of AFC (< 5 follicles) and/or AMH (< 1.2 ng/mL); and group 4—POR patients ≥ 35 years old, presenting with poor values of AFC (< 5 follicles) and/or AMH (< 1.2 ng/mL) [34,35,36].
POR accounts for up to 20% of women undergoing to IVF [37, 38]. Some factors responsible for POR are already well recognized and associated with ovarian aging, such as endometriosis, prior ovarian surgery, chemotherapy, radiation therapy, smoking, infections, and autoimmune disorders [39, 40]. However, little is known about the mechanisms responsible for POR in women with normal or abnormal ovarian reserve biomarkers [39].
In order to increase the number and quality of aspirated oocytes and, consequently, increase live birth rate, several studies have evaluated different ovarian stimulation protocols associated or not with adjuvant therapies in cases of POR. Pretreatment using coenzyme Q10 (CoQ10), testosterone, dehydroepiandrosterone (DHEA), and myoinositol, as well as, the use of gonadotropins associated with luteinizing hormone (LH), growth hormone (GH), clomiphene citrate, and letrozole, are some of the strategies suggested for the management of patients with POR [41,42,43]. However, to date, there are no adjuvant therapy that shown convincing results for management of POR patients [41]. A recent meta-analysis has shown that dehydroepiandrosterone (DHEA) and CoQ10 are promising adjuvant therapies for raising the clinical pregnancy rate (OR 2.46, 95% CI 1.16 to 5.23 and OR 2.22, 95% CI 1.08–4.58, respectively), while GH raised the number of oocytes retrieved (weighted mean difference 1.72, 95% CI 0.98 to 2.46) [41]. However, the authors concluded that high-level randomized controlled trial (RCT) studies using uniform standards for POR still need to be performed [41].
It is not uncommon for patients to undergo several IVF cycles with multiple rounds of ovarian hyperstimulation. Thus, since the establishment of IVF as a treatment technique for infertile couples, the medium- and long-term impact of repeated ovarian hyperstimulation on ovarian aging and estrogen-dependent diseases has been investigated [44, 45]. The use of gonadotropins at supraphysiological doses promotes maximum recruitment and maturation of small follicles, but the effects on primordial follicles (unresponsive to physiological levels of FSH) are poorly understood [45, 46].
Most studies have not observed an association between the use of gonadotropins for fertility treatment with accelerated ovarian aging or anticipation of menopausal symptoms [47]. However, women who had low number of aspirated oocytes in IVF cycles are at risk of anticipation of menopause symptoms and early menopause [44, 48,49,50]. Women who have multiple IVF attempts also have repeated transvaginal ultrasound-guided needle ovarian aspirations, with potential complications (bleeding, infection, and fibrosis) in the short and long term, which can contribute to accelerated ovarian aging.
Currently, female fertility preservation is possible through the vitrification of oocytes, embryos, or ovarian tissue [51, 52]. Among these three methods, vitrification of ovarian tissue, initially proposed for oncologic patients prior to chemotherapy or radiotherapy, and subsequent transplantation can restore not only the female reproductive potential, but also the ovarian hormone function [51]. Ovarian tissue cryopreservation and transplantation (OTCT) was performed for the first time in 1999 in a 29-year-old patient, with surgical menopause at age 28, who did not obtain improvement of menopause symptoms induced using conventional hormone therapy [53].
Recently, the American Society for Reproductive Medicine (ASRM) recognized that OTCT may be indicated for fertility preservation in situations where ovarian hyperstimulation for oocyte retrieval is contraindicated [54]. A systematic review of 309 cases of OTCT revealed that the intention of the vast majority of patients was the restoration of reproductive function, but on nine patients, the procedure was used with non-reproductive purpose, to restore ovarian endocrine function [55]. This same systematic review highlighted that OTCT was able to restore endocrine function in up to 85% of cases [55].
Despite the great progress in ovarian tissue cryopreservation studies, OTCT still has limitations. It is necessary to reduce the percentage of loss of primordial follicles during the freezing–thawing process, especially during revascularization after auto transplantation, increasing graft survival [56]. In addition to the success variables related to technical conditions, the woman’s age at the time of freezing and the amount of cryopreserved ovarian tissue are other determining factors for maintaining the endocrine and reproductive function of the thawed and transplanted ovarian tissue [57]. Oktay et al. found that with about a third of an ovary cryopreserved at a mean age of 29.3 (9–44) years, the mean longevity of transplanted ovarian tissue was 26.9 (4–144) months [55]. Surgical removal of ovarian tissue can also interfere with the age of onset of menopause. Studies are not clear about the real impact of a unilateral oophorectomy on the age of menopause, which may accelerate the onset of menopause from 1 to 7 years before control groups, not submitted to oophorectomy [56]. Thus, in light of early studies in reproductive medicine, OTCT is a promising intervention to delay the onset of menopause. Studies are also needed to identify the best protocols for freezing, thawing, and transplanting ovarian tissue, the ideal age for removing the ovarian cortex, and the number and interval of transplants [57,58,59].
Lessons from animal studies
Ovarian aging, evaluated from a comparative perspective between rodents and humans, also results from follicle loss starting in the prenatal period and an exponential depletion of primordial follicles associated with loss of fertility in middle age [60]. Furthermore, both species share progressively increasing irregularities in ovulatory cycles and increased pregnancy loss as oocyte depletion becomes imminent [60]. In C57BL/6 mice, the ovarian reserve is reduced in half around 10 months of age compared to 2-month-old mice [60], and at 18 months is reduced approximately 10 times [61]. Our group also observed a 75% reduction in primordial follicle density in C57BL/6 mice from 3 to 12 months of age [62] and severely compromised fertility around 14 months of age. Around this age, mice start to present irregularities in the estrous cycle and may enter a persistent anovulatory state, known as estropause [63]. This reinforces the idea that the ovarian reserve in mice, as in humans, is depleted with age and this process is completed in first half of the lifespan [60, 61].
The loss of ovarian function and the resulting decline in circulating estrogen levels are associated to various metabolic changes during induced menopause in rodents, similar to observed in women during natural menopause, such as increased adiposity, changes in lipid metabolism, hypertension, and insulin resistance [64, 65]. Although mice do not naturally enter in menopause, it can be induced chemically or by ovariectomy. Chemical menopause induction can be achieved by treatment with 4-vinylcyclohexene diepoxide (VCD). Treatment with VCD causes atresia of primordial and primary follicles leading to exhaustion of the ovarian reserve but retention of ovarian tissue [66]. One study showed that as young as 6-month-old rats induced to menopause by ovariectomy have a slight increase in systolic blood pressure (SBP), body weight, insulin resistance, and plasma cholesterol compared to intact females of the same age. This reinforces the notion that metabolic changes are driven by ovarian function independent of age [64]. These metabolic changes observed in ovariectomized rodents are a consequence of the hypoestrogenic state, similar to observed in women [65].
As estradiol reduction is one of the main effects of menopause, several studies have evaluated the benefits of exogenous estradiol replacement. Studies in murine models point to some beneficial effects of estradiol replacement therapy. These effects include improvement of glucose homeostasis and insulin sensitivity [67], improvement of the innate immune response [68], and reduced arterial pressure [69]. Interestingly, the transplantation of young ovaries into older females can increase lifespan [70], suggesting a protective effect of the ovarian tissue itself. Transplantation of young ovarian tissue to post-reproductive mice was able to significantly restore the cardioprotective benefits, similarly to what is seen at reproductive age [71]. These protective effects of ovarian tissue transplantation were also observed even when the young ovaries were follicle depleted with VCD prior to transplantation [72], suggesting that other factors beyond estrogen levels may play a role. Therefore, novel therapies to replace estrogen could be developed, improving quality of life without side effects of hormone replacement therapies.
Faced with the hormonal and physiological changes induced by menopause, some alternatives also aim at delaying the onset of menopause in preclinical models, such as ovarian transplantation of young mice into old mice [70, 72], dietary strategies [61, 73], and use of drugs, such as rapamycin [74, 75] and metformin [76, 77] have been developed. Dietary strategies have been proved widely effective in preserving the decline of follicle reserve in mice with aging. Caloric restriction (CR), ranging from 10 to 30% reduction of calorie intake [78], is an intervention that has been proven efficient in promoting overall longevity [79] as well as extended fertility in females, due to its impact on the control of oxidative stress, insulin resistance, and reduction of the inflammatory state [80, 81]. CR can extend fertility by preserving the ovarian reserve [78, 82], reducing mTOR and FOXO3 activation [77, 83, 84], the main regulators of primordial follicle activation [83]. Mice under 30% CR can still reproduce successfully around 15–16 months of age, when ad libitum-fed mice could not [61]. In addition, CR also regulated DNA repair mechanisms, preventing the accumulation of DNA damage [85, 86], which could impact oocyte quality. However, some studies have associated the positive effects of CR to the restriction of protein [87]. Protein restriction also results in lower activity of the mTOR pathway in the ovary, decreasing the activation of primordial follicles and preserving the ovarian reserve [88]. The reduction in protein supply also results in lower adiposity and greater insulin sensitivity in humans and rodents, similar to that observed with 30% CR [87, 89]. On the other hand, hypercaloric diets (high-fat and/or high-carbohydrate diets) cause harmful metabolic adaptions, decreasing the ovarian reserve. Mice exposed to a high-fat diet have decreased ovarian reserve and increased insulin resistance and inflammation in the ovarian tissues [90, 91]. Interestingly, this unhealthy diet decreased ovarian reserve even in the absence of body weight gain [90].
Interestingly, the nematode C. elegans and the fly D. melanogaster can also serve as model for reproductive aging. C. elegans also have decreased reproductive potential as they age in both self-fertile and mated reproduction [92]. As reviewed elsewhere, CR, metformin, and rapamycin treatment can also extend reproductive lifespan in C. elegans [92], resembling the effects observed in mice. Similarly to the observed in humans and mice, an hyperactive germline in C. elegans is linked to reduced lifespan [93], suggesting a central role for reproductive fitness to regulate overall lifespan. The female fly D. melanogaster also has decreased reproductive potential with age [94]. In the fly, lifetime egg production was maximized in a diet containing a low protein to carbohydrate content [95]. The overall dietary restriction and rapamycin treatment both reduce daily egg laying, however increasing overall lifespan [96], further confirm a link between reproduction and lifespan in this species.
Repurposing of FDA-approved drugs to target aging
Based on ovarian physiology, animal studies, lessons from menopause and reproductive medicine studies, and the ideal strategy for delaying ovarian aging should have the following characteristics: (1) be initiated in the presence of good ovarian reserve; (2) maintained for a long period; (3) have an action on the dynamics of primordial follicles, controlling the rate of activation and atresia; and (4) safe use in pre-conception, pregnancy, and lactation.
Recently, Kulkarni et al. suggested the off-label use of FDA-approved drugs to target aging [97]. Based on animal and human studies that evaluated the effect of drugs on hallmarks of aging, healthspan, and lifespan, the authors scored (from 0 to 12) nine drugs according to potential anti-aging effects. Sodium-glucose linked transporter 2 (SGLT-2) inhibitors was the drug with the highest antiaging potential (score 12), followed by metformin (score 11), acarbose (score 9), rapamycin (score 9), methylene blue (score 9), angiotensin-converting enzyme inhibitors and angiotensin receptor blockers (ACEi/ARB) (score 8), dasatinib (+ quercetin) (score 6), aspirin (score 6), and N-acetyl cyeteine (NAC) (score 5) [97].
These nine potential anti-aging drugs belong to different classes. Three of these nine drugs are oral hypoglycemic agents, the SGLT-2 inhibitors, metformin, and acarbose [97]. The impact of these gerotherapeutic agents on ovarian physiology is poorly understood. The benefit of using these hypoglycemic agents for female fertility is essentially due to adequate glycemic control and weight loss, with improvement in the ovulatory function [98]. However, the medium- and long-term effects on ovarian aging are not well known.
The effects of metformin on female reproductive outcomes are extensively studied, especially in patients with polycystic ovary syndrome. In addition to improving spontaneous pregnancy rates, increase implantation rates in IVF cycles, and decrease miscarriage rates among PCOS patients, metformin regulates ovarian mechanisms that may contribute to a delay in ovarian aging [98]. Studies in animal models have revealed that metformin reduces follicular atresia, oxidative stress, and autophagy in granulosa cells via the PI3K/AKT/mTOR pathway [99, 100]. Qin et al. suggested that metformin may delay the ovarian aging process, probably by inducing SIRT1 expression and reducing oxidative damage [76]. Recently, Landry et al. observed that metformin was able to prevent age-associated ovarian fibrosis by modulating the proportion of fibroblasts, myofibroblasts, and immune cells [101]. At a dosage of 100 mg/kg, metformin was able to prevent the decline in the ovarian reserve with age in older mice [76]. In addition, metformin has chemoprotective effects in the ovaries of young mice exposed to cyclophosphamide [102]. Therefore, metformin due improvement of insulin sensitivity has similar effects to a 30% CR diet, and can be an alternative to dietary interventions.
Rapamycin is a substance originally produced by the bacterium Streptomyces hygroscopicus. In the past, rapamycin was used as an antifungal drug, later as an immunosuppressive agent [103]. Rapamycin acts in tissues by inhibiting to mammalian target of rapamycin (mTOR), a member of the phosphatidylinositol 3-kinase family of protein kinases. In this sense, mTOR is a serine/threonine protein kinase that functions as a master regulator of cell growth and metabolism in response to nutritional and hormonal stimuli [104], and is suggested as a CR mimetic. The mTOR signaling is involved in follicular dynamics, and its activation in granulosa cells promotes follicular development [105], and its inhibition can prolong ovarian lifespan [103]. Animal studies have shown that treatment of female mice with rapamycin prevented activation of primordial follicles when compared to untreated controls [82, 106, 107]. Our previous study suggested that treatment with rapamycin (4 mg/kg) prevents the decline in the ovarian reserve of mice similarly to 30% CR [82]. Additionally, rapamycin administered to young animals (5 mg/kg) exposed to chemotherapy also decreased the activation of the mTOR pathway, preserving the ovarian reserve [108].
Among many adaptations with aging, there is an accumulation of senescent cells with age in several species [6, 109]. Senescent cells have decreased capacity for proliferation [110,111,112], however remain metabolically active, secreting pro-inflammatory factors known as senescence-associated secretory phenotype (SASP) [113]. Our group demonstrated an accumulation of senescent cells with aging also in the ovary of mice [62]. Furthermore, we demonstrated that obese mice accumulate more senescent cells in the ovary than normal littermates [114], suggesting that obesity accelerates the ovarian aging phenotype. Interestingly, senolytic drugs, such as dasatinib and quercetin (D + Q), were able to reduce cellular senescence in the ovary of obese mice, without affecting the ovarian reserve. [114]. In young ovaries exposed to chemotherapy, an increase in cellular senescence was also observed, which was prevented by treatment with D + Q [115]. Furthermore, metformin along with D + Q was able to reduce ovarian senescence even more, maintain regular estrous cycle, and prevent excessive primordial follicle activation [116].
NAC is a stable form of the essential amino acid L-cysteine. NAC is converted to L-cysteine after ingestion and subsequently converted to glutathione [117]. NAC has important antioxidant and anti-inflammatory properties [117]. Animal studies have observed that NAC is able to reduce ovarian damage by upregulating local antioxidant capacity and reducing ovarian secretion of pro-inflammatory cytokines [118, 119]. In humans, the antioxidant action of NAC contributed to reduce cell damage in ovarian tissue cryopreservation protocols [120, 121]. NAC also has an antifibrotic action, already described in tissues such as muscle, aorta, and kidney [122,123,124]. Therefore, although there are no studies evaluating the long-term effect of NAC on ovarian physiology, these properties may contribute to delaying ovarian aging. There are no animal or human studies evaluating a possible impact of SGLT-2 inhibitors, acarbose, methylene blue, ACEi/ARB, and aspirin on ovarian aging.
We found no clear evidence in the literature that these intervention strategies can have a negative impact on overall fertility in animal models and humans. However, it should be noted that interventions that prevent ovarian aging result in retention of follicles in the primordial stage, which will have a negative effect on fertility during the period of the intervention. For example, in mice under 30% CR, follicles progress slower to secondary and tertiary stages, and, therefore, we observe a preserved pool of primordial follicles [82], which is desirable in the long term. However, as these follicles are not progressing to the secondary and tertiary stages, we observe a reduction in ovulation rate, reflecting in decreased fertility during the course of CR [78]. However, once mice are put back in ad libitum, feeding pregnancy rate increases [78] and is maintained above control group in aged mice due to preserved ovarian reserve [61]. Therefore, in a proposed dietary/pharmacological intervention protocol to preserve the ovarian reserve, one should consider removing the treatment once pregnancy is desired. However, more studies are necessary to understand how long these interventions should be applied and how long before pregnancy is desired they should be interrupted, in order to maximize preservation of the ovarian reserve without compromising fertility.
New possibilities for ovarian anti-aging interventions
Recently, numerous studies, including those by Johnson et al. and White et al., have described the presence of ovarian germline stem cells (GSCs) in ovaries of non-mammals, mammals, and humans [125,126,127,128]. These findings began to question the dogma, established by Zuckerman in 1951, of the inability of new ovarian germ cells to arise [129].
Currently, studies seek to define protocols for ovarian rejuvenation or ovarian regeneration, through interventions that stimulate the emergence of new follicular units from GSCs. Injection of platelet-rich plasma (PRP) and stem cells into the ovaries are the two most studied treatments at present, especially in patients suffering from ovarian insufficiency and low ovarian reserve or undergoing IVF treatments with a poor prognosis [130,131,132].
In 2018, Sills et al. were the first to describe that intraovarian injection of autologous PRP significantly improved the response of four patients with diminished ovarian reserve as determined by at least one prior IVF cycle canceled for poor follicular recruitment response. Decreased FSH and increased AMH levels were also observed after the intervention. All four patients had at least 1 day five blastocyst available for cryopreservation [130]. Currently, numerous studies are being conducted to validate the use of PRP as an alternative treatment for female infertility associated with low ovarian reserve [133, 134].
Intraovarian injection of mesenchymal stem cells (MSCs) is another promising alternative to stimulate ovarian rejuvenation. MSCs can originate from several sources, such as amniotic fluid, endometrium, germ cells, skin, umbilical cord, bone marrow, and adipose tissue [131]. Stimulation of angiogenesis and cell proliferation, a decrease of apoptosis, modulation of immune cells function, and regulation of gene expression in the ovary are possible mechanisms involved in the process of ovarian rejuvenation that are observed after MSC transplantation [131]. However, clinical studies are needed to evaluate the impact of treating women with low ovarian reserve undergoing IVF cycles.
Conclusion
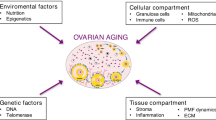
Based on the evidence presented in this review, it becomes clear that the ovarian reserve has a central role in the female life. The progressive decline in the ovarian reserve with age is natural; however, it can be accelerated by several factors, like diet and lifestyle. Once the ovarian reserve starts to become severely compromised, females experience a decline in natural fertility, as well as in the response to assisted reproductive technologies. Later in life, once this ovarian reserve is depleted, females experience changes in the body physiology, resulting in increased chronic disease and mortality risk. Therefore, the use of strategies to prevent the decline in the ovarian reserve should be investigated to improve fertility, healthspan, and lifespan. There are several drugs that can be repurposed for this aim; however, we need to better understand the best moment that these therapies must be initiated to promote the desired effects (Fig. 1).

Ovarian aging results from activation of primordial follicles. This decline in the ovarian reserve leads to reduce fertility and menopause later in the female life. Despite some lifestyle factors are a risk for decreased ovarian reserve, some strategies can prevent this decline
References
Oktem O, Oktay K. The ovary: anatomy and function throughout human life. Ann N Y Acad Sci. 2008;1127:1–9.
Baerwald AR, Adams GP, Pierson RA. Ovarian antral folliculogenesis during the human menstrual cycle: a review. Hum Reprod Update. 2012;18:73–91.
Baker TG. A quantitative and cytological study of germ cells in human ovaries. Proc R Soc Lond B Biol Sci. 1963;158:417–33.
Crawford NM, Steiner AZ. Age-related infertility. Obstet Gynecol Clin North Am. 2015;42:15–25.
Overland MR, Li Y, Derpinghaus A, Aksel S, Cao M, Ladwig N, Cunha GR, Himelreich-Perić M, Baskin LS. Development of the human ovary: fetal through pubertal ovarian morphology, folliculogenesis and expression of cellular differentiation markers. Differentiation. 2023;129:37–59.
Secomandi L, Borghesan M, Velarde M, Demaria M. The role of cellular senescence in female reproductive aging and the potential for senotherapeutic interventions. Hum Reprod Update. 2022;28:172–89.
Park SU, Walsh L, Berkowitz KM. Mechanisms of ovarian aging. Reproduction. 2021;162:R19–33.
The 2022 Hormone Therapy Position Statement of The North American Menopause Society Advisory Panel. The 2022 hormone therapy position statement of The North American Menopause Society. Menopause. 2022;(29):767–94.
Katainen R, Engblom JR, Polo-Kantola P. Climacteric-related symptoms in menopause transition and beyond: a prospective 19-year follow-up study on previously hysterectomized women. Menopause. 2018;25:890–6.
McKinlay SM, Brambilla DJ, Posner JG. The normal menopause transition. Maturitas. 2008;61:4–16.
Gold EB. The timing of the age at which natural menopause occurs. Obstet Gynecol Clin North Am. 2011;38:425–40.
El Khoudary SR, Greendale G, Crawford SL, Avis NE, Brooks MM, Thurston RC, Karvonen-Gutierrez C, Waetjen LE, Matthews K. The menopause transition and women’s health at midlife: a progress report from the Study of Women’s Health Across the Nation (SWAN). Menopause. 2019;26:1213–27.
Ceylan B, Özerdoğan N. Factors affecting age of onset of menopause and determination of quality of life in menopause. Turk J Obstet Gynecol. 2015;12:43–9.
Newson L. Menopause and cardiovascular disease. Post Reprod Health. 2018;24:44–9.
de Kat AC, Broekmans FJM, Lambalk CB. Role of AMH in prediction of menopause. Front Endocrinol (Lausanne). 2021;12:733731.
Depmann M, Broer SL, van der Schouw YT, Tehrani FR, Eijkemans MJ, Mol BW, Broekmans FJ. Can we predict age at natural menopause using ovarian reserve tests or mother’s age at menopause? Syst Lit Rev Menopause. 2016;23:224–32.
Robertson DM. Inhibins and activins in blood: predictors of female reproductive health? Mol Cell Endocrinol. 2012;359:78–84.
Moolhuijsen LME, Visser JA. Anti-Müllerian hormone and ovarian reserve: update on assessing ovarian function. J Clin Endocrinol Metab. 2020;105:3361–73.
Unuane D, Tournaye H, Velkeniers B, Poppe K. Endocrine disorders & female infertility. Best Pract Res Clin Endocrinol Metab. 2011;25:861–73.
Lambalk CB, van Disseldorp J, de Koning CH, Broekmans FJ. Testing ovarian reserve to predict age at menopause. Maturitas. 2009;63:280–91.
Vander Borght M, Wyns C. Fertility and infertility: definition and epidemiology. Clin Biochem. 2018;62:2–10.
Coulam CB, Adamson SC, Annegers JF. Incidence of premature ovarian failure. Obstet Gynecol. 1986;67:604–6.
Chon SJ, Umair Z, Yoon MS. Premature ovarian insufficiency: past, present, and future. Front Cell Dev Biol. 2021;9:672890.
Podfigurna-Stopa A, Czyzyk A, Grymowicz M, Smolarczyk R, Katulski K, Czajkowski K, Meczekalski B. Premature ovarian insufficiency: the context of long-term effects. J Endocrinol Invest. 2016;39:983–90.
Malek AM, Vladutiu CJ, Meyer ML, Cushman M, Newman R, Lisabeth LD, Kleindorfer D, Lakkur S, Howard VJ. The association of age at menopause and all-cause and cause-specific mortality by race, postmenopausal hormone use, and smoking status. Prev Med Rep. 2019;15:100955.
Blümel JE, Mezones-Holguín E, Chedraui P, Soto-Becerra P, Arteaga E, Vallejo MS. Is premature ovarian insufficiency associated with mortality? A three-decade follow-up cohort. Maturitas. 2022;163:82–7.
Jacobsen BK, Knutsen SF, Fraser GE. Age at natural menopause and total mortality and mortality from ischemic heart disease: the Adventist Health Study. J Clin Epidemiol. 1999;52:303–7.
Popat VB, Calis KA, Vanderhoof VH, Cizza G, Reynolds JC, Sebring N, Troendle JF, Nelson LM. Bone mineral density in estrogen-deficient young women. J Clin Endocrinol Metab. 2009;94:2277–83.
Bakalov VK, Anasti JN, Calis KA, Vanderhoof VH, Premkumar A, Chen S, Furmaniak J, Smith BR, Merino MJ, Nelson LM. Autoimmune oophoritis as a mechanism of follicular dysfunction in women with 46, XX spontaneous premature ovarian failure. Fertil Steril. 2005;84:958–65.
Bove R, Secor E, Chibnik LB, Barnes LL, Schneider JA, Bennett DA, De Jager PL. Age at surgical menopause influences cognitive decline and Alzheimer pathology in older women. Neurology. 2014;82:222–9.
Schmidt PJ, Luff JA, Haq NA, Vanderhoof VH, Koziol DE, Calis KA, Rubinow DR, Nelson LM. Depression in women with spontaneous 46, XX primary ovarian insufficiency. J Clin Endocrinol Metab. 2011;96:E278–87.
Bellver J, Rodríguez-Tabernero L, Robles A, Muñoz E, Martínez F, Landeras J, García-Velasco J, Fontes J, Álvarez M, Álvarez C, Acevedo B, Group of interest in Reproductive Endocrinology (GIER) of the Spanish Fertility Society (SEF). Polycystic ovary syndrome throughout a woman’s life. J Assist Reprod Genet. 2018;35:25–39.
Ehrlich S. Effect of fertility and infertility on longevity. Fertil Steril. 2015;103:1129–35.
Conforti A, Esteves SC, Picarelli S, Iorio G, Rania E, Zullo F, De Placido G, Alviggi C. Novel approaches for diagnosis and management of low prognosis patients in assisted reproductive technology: the POSEIDON concept. Panminerva Med. 2019;61:24–9.
Drakopoulos P, Bardhi E, Boudry L, Vaiarelli A, Makrigiannakis A, Esteves SC, Tournaye H, Blockeel C. Update on the management of poor ovarian response in IVF: the shift from Bologna criteria to the Poseidon concept. Ther Adv Reprod Health. 2020;14:2633494120941480.
Abu-Musa A, Haahr T, Humaidan P. Novel physiology and definition of poor ovarian response; clinical recommendations. Int J Mol Sci. 2020;21:2110.
Zhuang J, Li H, Li X, Tian D, Yang D, Zhu M. The incidence of unexpected poor ovarian response in Chinese young women. Medicine (Baltimore). 2019;98:e14379.
Venetis CA, Kolibianakis EM, Tarlatzi TB, Tarlatzis BC. Evidence-based management of poor ovarian response. Ann N Y Acad Sci. 2010;1205:199–206.
Badawy A, Wageah A, El Gharib M, Osman EE. Prediction and diagnosis of poor ovarian response: the dilemma. J Reprod Infertil. 2011;12:241–8.
Nikolaou D, Templeton A. Early ovarian ageing: a hypothesis. Detection and clinical relevance. Hum Reprod. 2003;18:1137–9.
Zhang Y, Zhang C, Shu J, Guo J, Chang HM, Leung PCK, Sheng JZ, Huang H. Adjuvant treatment strategies in ovarian stimulation for poor responders undergoing IVF: a systematic review and network meta-analysis. Hum Reprod Update. 2020;26:247–63.
Wang Z, Yang A, Bao H, Wang A, Deng X, Xue D, Tan H, Zhou Y, Wu C, Chen ZJ, Shi Y. Effect of dehydroepiandrosterone administration before in vitro fertilization on the live birth rate in poor ovarian responders according to the Bologna criteria: a randomised controlled trial. BJOG. 2022;129:1030–8.
Hart RJ. Stimulation for low responder patients: adjuvants during stimulation. Fertil Steril. 2022;117:669–74.
Muharam R, Sumapraja K, Pratama G, Azyati M, Prabowo KA. Impact of IVF on the timing and symptoms of menopause. Int J Womens Health. 2021;13:889–93.
Bai L, Pan H, Zhao Y, Chen Q, Xiang Y, Yang X, Zhu Y. The exploration of poor ovarian response-related risk factors: a potential role of growth differentiation factor 8 in predicting ovarian response in IVF-ET patient. Front Endocrinol (Lausanne). 2021;12:708089.
Whang J, Ahn C, Kim S, Seok E, Yang Y, Han G, Jo H, Yang H. Effects of repeated ovarian stimulation on ovarian function and aging in mice. Dev Reprod. 2021;25:213–23.
Elder K, Mathews T, Kutner E, Kim E, Espenberg D, Faddy M, Gosden R. Impact of gonadotrophin stimulation for assisted reproductive technology on ovarian ageing and menopause. Reprod Biomed Online. 2008;16:611–6.
de Boer EJ, den Tonkelaar I, te Velde ER, Burger CW, van Leeuwen FE, OMEGA-project group. Increased risk of early menopausal transition and natural menopause after poor response at first IVF treatment. Hum Reprod. 2003;18:1544–52.
de Boer EJ, den Tonkelaar I, te Velde ER, Burger CW, Klip H, van Leeuwen FE, OMEGA-project group. A low number of retrieved oocytes at in vitro fertilization treatment is predictive of early menopause. Fertil Steril. 2002;77:978–85.
Szmidt NA, Bhattacharya S, Maheshwari A. Does poor ovarian response to gonadotrophins predict early menopause? A retrospective cohort study with minimum of 10-year follow-up. Hum Fertil (Camb). 2016;19:212–9.
Kasaven LS, Saso S, Getreu N, O’Neill H, Bracewell-Milnes T, Shakir F, Yazbek J, Thum MY, Nicopoullos J, Ben Nagi J, Hardiman P, Diaz-Garcia C, Jones BP. Age-related fertility decline: is there a role for elective ovarian tissue cryopreservation? Hum Reprod. 2022;37:1970–9.
Fisch B, Abir R. Female fertility preservation: past, present and future. Reproduction. 2018;156:F11–27.
Oktay K, Karlikaya G. Ovarian function after transplantation of frozen, banked autologous ovarian tissue. N Engl J Med. 2000;342:1919.
Practice Committee of the American Society for Reproductive Medicine. Fertility preservation in patients undergoing gonadotoxic therapy or gonadectomy: a committee opinion. Fertil Steril. 2019;112:1022–33.
Pacheco F, Oktay K. Current success and efficiency of autologous ovarian transplantation: a meta-analysis. Reprod Sci. 2017;24:1111–20.
Oktay KH, Marin L, Petrikovsky B, Terrani M, Babayev SN. Delaying reproductive aging by ovarian tissue cryopreservation and transplantation: is it prime time? Trends Mol Med. 2021;27:753–61.
Chen J, Han Y, Shi W, Yan X, Shi Y, Yang Y, Gao H, Li Y. Ovarian tissue bank for fertility preservation and anti-menopause hormone replacement. Front Endocrinol (Lausanne). 2022;13:950297.
von Wolff M, Stute P. Cryopreservation and transplantation of ovarian tissue exclusively to postpone menopause: technically possible but endocrinologically doubtful. Reprod Biomed Online. 2015;31:718–21.
Kolibianaki EE, Goulis DG, Kolibianakis EM. Ovarian tissue cryopreservation and transplantation to delay menopause: facts and fiction. Maturitas. 2020;142:64–7.
Finch CE. The menopause and aging, a comparative perspective. J Steroid Biochem Mol Biol. 2014;142:132–41.
Selesniemi K, Lee HJ, Tilly JL. Moderate caloric restriction initiated in rodents during adulthood sustains function of the female reproductive axis into advanced chronological age. Aging Cell. 2008;7:622–9.
Ansere VA, Ali-Mondal S, Sathiaseelan R, Garcia DN, Isola JVV, Henseb JD, Saccon TD, Ocañas SR, Tooley KB, Stout MB, Schneider A, Freeman WM. Cellular hallmarks of aging emerge in the ovary prior to primordial follicle depletion. Mech Ageing Dev. 2021;194:111425.
Koebele SV, Bimonte-Nelson HA. Modeling menopause: the utility of rodents in translational behavioral endocrinology research. Maturitas. 2016;87:5–17.
Bitto A, Altavilla D, Bonaiuto A, Polito F, Minutoli L, Di Stefano V, Giuliani D, Guarini S, Arcoraci V, Squadrito F. Effects of aglycone genistein in a rat experimental model of postmenopausal metabolic syndrome. J Endocrinol. 2009;200:367–76.
Medina-Contreras J, Villalobos-Molina R, Zarain-Herzberg A, Balderas-Villalobos J. Ovariectomized rodents as a menopausal metabolic syndrome model. A minireview. Mol Cell Biochem. 2020;475:261–76.
Brooks HL, Pollow DP, Hoyer PB. The VCD mouse model of menopause and perimenopause for the study of sex differences in cardiovascular disease and the metabolic syndrome. Physiology (Bethesda). 2016;31:250–7.
Camporez JP, Jornayvaz FR, Lee HY, Kanda S, Guigni BA, Kahn M, Samuel VT, Carvalho CR, Petersen KF, Jurczak MJ, Shulman GI. Cellular mechanism by which estradiol protects female ovariectomized mice from high-fat diet-induced hepatic and muscle insulin resistance. Endocrinology. 2013;154:1021–8.
Schneider AH, Kanashiro A, Dutra SGV, Souza RDN, Veras FP, Cunha FQ, Ulloa L, Mecawi AS, Reis LC, Malvar DDC. Estradiol replacement therapy regulates innate immune response in ovariectomized arthritic mice. Int Immunopharmacol. 2019;72:504–10.
Pollow DP Jr, Romero-Aleshire MJ, Sanchez JN, Konhilas JP, Brooks HL. ANG II-induced hypertension in the VCD mouse model of menopause is prevented by estrogen replacement during perimenopause. Am J Physiol Regul Integr Comp Physiol. 2015;309:R1546–52.
Mason JB, Parkinson KC, Habermehl TL. Orthotopic ovarian transplantation procedures to investigate the life- and health-span influence of ovarian senescence in female mice. J Vis Exp. 2018;132:56638.
Mason JB, Cargill SL, Griffey SM, Reader JR, Anderson GB, Carey JR. Transplantation of young ovaries restored cardioprotective influence in postreproductive-aged mice. Aging Cell. 2011;10:448–56.
Habermehl TL, Underwood KB, Welch KD, Gawrys SP, Parkinson KC, Schneider A, Masternak MM, Mason JB. Aging-associated changes in motor function are ovarian somatic tissue-dependent, but germ cell and estradiol independent in post-reproductive female mice exposed to young ovarian tissue. Geroscience. 2022;44:2157–69.
Masoro EJ. Overview of caloric restriction and ageing. Mech Ageing Dev. 2005;126:913–22.
Richardson MC, Guo M, Fauser BC, Macklon NS. Environmental and developmental origins of ovarian reserve. Hum Reprod Update. 2014;20:353–69.
Luo LL, Xu JJ, Fu YC. Rapamycin prolongs female reproductive lifespan. Cell Cycle. 2013;12:3353–4.
Qin X, Du D, Chen Q, Wu M, Wu T, Wen J, Jin Y, Zhang J, Wang S. Metformin prevents murine ovarian aging. Aging (Albany NY). 2019;11:3785–94.
Schneider A, Saccon TD, Garcia DN, Zanini BM, Isola JVV, Hense JD, Alvarado-Rincón JA, Cavalcante MB, Mason JB, Stout MB, Bartke A, Masternak MM. The interconnections between somatic and ovarian aging in murine models. J Gerontol A Biol Sci Med Sci. 2021;76:1579–86.
Isola JVV, Zanini BM, Hense JD, Alvarado-Rincón JA, Garcia DN, Pereira GC, Vieira AD, Oliveira TL, Collares T, Gasperin BG, Stout MB, Schneider A. Mild calorie restriction, but not 17α-estradiol, extends ovarian reserve and fertility in female mice. Exp Gerontol. 2022;159:111669.
Green CL, Lamming DW, Fontana L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat Rev Mol Cell Biol. 2022;23:56–73.
Heilbronn LK, Ravussin E. Calorie restriction and aging: review of the literature and implications for studies in humans. Am J Clin Nutr. 2003;78:361–9.
Flanagan EW, Most J, Mey JT, Redman LM. Calorie restriction and aging in humans. Annu Rev Nutr. 2020;40:105–33.
Garcia DN, Saccon TD, Pradiee J, Rincón JAA, Andrade KRS, Rovani MT, Mondadori RG, Cruz LAX, Barros CC, Masternak MM, Bartke A, Mason JB, Schneider A. Effect of caloric restriction and rapamycin on ovarian aging in mice. Geroscience. 2019;41:395–408.
Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science. 2003;301:215–8.
Li L, Fu YC, Xu JJ, Lin XH, Chen XC, Zhang XM, Luo LL. Caloric restriction promotes the reserve of follicle pool in adult female rats by inhibiting the activation of mammalian target of rapamycin signaling. Reprod Sci. 2015;22:60–7.
Heydari AR, Unnikrishnan A, Lucente LV, Richardson A. Caloric restriction and genomic stability. Nucleic Acids Res. 2007;35:7485–96.
Vermeij WP, Dollé ME, Reiling E, Jaarsma D, Payan-Gomez C, Bombardieri CR, Wu H, Roks AJ, Botter SM, van der Eerden BC, Youssef SA, Kuiper RV, Nagarajah B, van Oostrom CT, Brandt RM, Barnhoorn S, Imholz S, Pennings JL, de Bruin A, Gyenis Á, Pothof J, Vijg J, van Steeg H, Hoeijmakers JH. Restricted diet delays accelerated ageing and genomic stress in DNA-repair-deficient mice. Nature. 2016;537:427–31.
Solon-Biet SM, McMahon AC, Ballard JW, Ruohonen K, Wu LE, Cogger VC, Warren A, Huang X, Pichaud N, Melvin RG, Gokarn R, Khalil M, Turner N, Cooney GJ, Sinclair DA, Raubenheimer D, Le Couteur DG, Simpson SJ. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metab. 2014;19:418–30.
Zhuo Y, Hua L, Feng B, Jiang X, Li J, Jiang D, Huang X, Zhu Y, Li Z, Yan L, Jin C, Che L, Fang Z, Lin Y, Xu S, Li J, Wu D. Fibroblast growth factor 21 coordinates adiponectin to mediate the beneficial effects of low-protein diet on primordial follicle reserve. EBioMedicine. 2019;41:623–35.
Henagan TM, Laeger T, Navard AM, Albarado D, Noland RC, Stadler K, Elks CM, Burk D, Morrison CD. Hepatic autophagy contributes to the metabolic response to dietary protein restriction. Metabolism. 2016;65:805–15.
Skaznik-Wikiel ME, Swindle DC, Allshouse AA, Polotsky AJ, McManaman JL. High-fat diet causes subfertility and compromised ovarian function independent of obesity in mice. Biol Reprod. 2016;94:108.
Nteeba J, Ross JW, Perfield JW 2nd, Keating AF. High fat diet induced obesity alters ovarian phosphatidylinositol-3 kinase signaling gene expression. Reprod Toxicol. 2013;42:68–77.
Scharf A, Pohl F, Egan BM, Kocsisova Z, Kornfeld K. Reproductive aging in Caenorhabditis elegans: from molecules to ecology. Front Cell Dev Biol. 2021;9:718522.
Shi C, Murphy CT. piRNAs regulate a Hedgehog germline-to-soma pro-aging signal. Nat Aging. 2023;3:47–63.
Miller PB, Obrik-Uloho OT, Phan MH, Medrano CL, Renier JS, Thayer JL, Wiessner G, Bloch Qazi MC. The song of the old mother: reproductive senescence in female drosophila. Fly (Austin). 2014;8:127–39.
Lee KP, Simpson SJ, Clissold FJ, Brooks R, Ballard JW, Taylor PW, Soran N, Raubenheimer D. Lifespan and reproduction in Drosophila: new insights from nutritional geometry. Proc Natl Acad Sci U S A. 2008;105:2498–503.
Steenwinkel TE, Hamre KK, Werner T. The use of non-model Drosophila species to study natural variation in TOR pathway signaling. PLoS ONE. 2022;17:e0270436.
Kulkarni AS, Aleksic S, Berger DM, Sierra F, Kuchel GA, Barzilai N. Geroscience-guided repurposing of FDA-approved drugs to target aging: a proposed process and prioritization. Aging Cell. 2022;21(4):e13596.
Joham AE, Norman RJ, Stener-Victorin E, Legro RS, Franks S, Moran LJ, Boyle J, Teede HJ. Polycystic ovary syndrome. Lancet Diabetes Endocrinol. 2022;10:668–80.
Xu B, Dai W, Liu L, Han H, Zhang J, Du X, Pei X, Fu X. Metformin ameliorates polycystic ovary syndrome in a rat model by decreasing excessive autophagy in ovarian granulosa cells via the PI3K/AKT/mTOR pathway. Endocr J. 2022;69:863–75.
Yao J, Ma Y, Zhou S, Bao T, Mi Y, Zeng W, Li J, Zhang C. Metformin prevents follicular atresia in aging laying chickens through activation of PI3K/AKT and calcium signaling pathways. Oxid Med Cell Longev. 2020;2020:3648040.
Landry DA, Yakubovich E, Cook DP, Fasih S, Upham J, Vanderhyden BC. Metformin prevents age-associated ovarian fibrosis by modulating the immune landscape in female mice. Sci Adv. 2022;8:eabq1475.
Huang CC, Chou CH, Yang YS, Ho HN, Shun CT, Wen WF, Chen SU, Chen MJ. Metformin: a novel promising option for fertility preservation during cyclophosphamide-based chemotherapy. Mol Hum Reprod. 2021;27:gaaa084.
Guo Z, Yu Q. Role of mTOR signaling in female reproduction. Front Endocrinol (Lausanne). 2019;10:692.
Corton JM, Gillespie JG, Hardie DG. Role of the AMP-activated protein kinase in the cellular stress response. Curr Biol. 1994;4:315–24.
Adhikari D, Zheng W, Shen Y, Gorre N, Hämäläinen T, Cooney AJ, Huhtaniemi I, Lan ZJ, Liu K. Tsc/mTORC1 signaling in oocytes governs the quiescence and activation of primordial follicles. Hum Mol Genet. 2010;19:397–410.
Sato Yorino, Kawamura K. Rapamycin treatment maintains developmental potential of oocytes in mice and follicle reserve in human cortical fragments grafted into immune-deficient mice. Mol Cell Endocrinol. 2020;504:110694.
Dou X, Sun Y, Li J, Zhang J, Hao D, Liu W, Wu R, Kong F, Peng X, Li J. Short-term rapamycin treatment increases ovarian lifespan in young and middle-aged female mice. Aging Cell. 2017;16:825–36.
Zhou L, Xie Y, Li S, Liang Y, Qiu Q, Lin H, Zhang Q. Rapamycin prevents cyclophosphamide-induced over-activation of primordial follicle pool through PI3K/Akt/mTOR signaling pathway in vivo. J Ovarian Res. 2017;10:56.
Schafer MJ, Miller JD, LeBrasseur NK. Cellular senescence: implications for metabolic disease. Mol Cell Endocrinol. 2017;455:93–102.
Childs BG, Durik M, Baker DJ, van Deursen JM. Cellular senescence in aging and age-related disease: from mechanisms to therapy. Nat Med. 2015;21:1424–35.
Broekmans FJ, Soules MR, Fauser BC. Ovarian aging: mechanisms and clinical consequences. Endocr Rev. 2009;30:465–93.
Bernadotte A, Mikhelson VM, Spivak IM. Markers of cellular senescence. Telomere shortening as a marker of cellular senescence. Aging (Albany NY). 2016;8:3–11.
Kirkland JL, Tchkonia T. Cellular senescence: a translational perspective. EBioMedicine. 2017;21:21–8.
Hense JD, Garcia DN, Isola JV, Alvarado-Rincón JA, Zanini BM, Prosczek JB, Stout MB, Mason JB, Walsh PT, Brieño-Enríquez MA, Schadock I, Barros CC, Masternak MM, Schneider A. Senolytic treatment reverses obesity-mediated senescent cell accumulation in the ovary. Geroscience. 2022;44:1747–59.
Gao Y, Wu T, Tang X, Wen J, Zhang Y, Zhang J, Wang S. Increased cellular senescence in doxorubicin-induced murine ovarian injury: effect of senolytics. Geroscience. 2023. https://doi.org/10.1007/s11357-023-00728-2
Du D, Tang X, Li Y, Gao Y, Chen R, Chen Q, Wen J, Wu T, Zhang Y, Lu H, Zhang J, Wang S. Senotherapy protects against cisplatin-induced ovarian injury by removing senescent cells and alleviating DNA damage. Oxid Med Cell Longev. 2022;2022:9144644.
Raghu G, Berk M, Campochiaro PA, Jaeschke H, Marenzi G, Richeldi L, Wen FQ, Nicoletti F, Calverley PMA. The multifaceted therapeutic role of N-acetylcysteine (NAC) in disorders characterized by oxidative stress. Curr Neuropharmacol. 2021;19:1202–24.
Cao X, Guo L, Zhou C, Huang C, Li G, Zhuang Y, Yang F, Liu P, Hu G, Gao X, Guo X. Effects of N-acetyl-l-cysteine on chronic heat stress-induced oxidative stress and inflammation in the ovaries of growing pullets. Poult Sci. 2023;102:102274.
Fan L, Guan F, Ma Y, Zhang Y, Li L, Sun Y, Cao C, Du H, He M. N-Acetylcysteine improves oocyte quality through modulating the Nrf2 signaling pathway to ameliorate oxidative stress caused by repeated controlled ovarian hyperstimulation. Reprod Fertil Dev. 2022;34:736–50.
Fabbri R, Sapone A, Paolini M, Vivarelli F, Franchi P, Lucarini M, Pasquinelli G, Vicenti R, Macciocca M, Venturoli S, Canistro D. Effects of N-acetylcysteine on human ovarian tissue preservation undergoing cryopreservation procedure. Histol Histopathol. 2015;30:725–35.
Barrozo LG, Paulino LRFM, Silva BR, Barbalho EC, Nascimento DR, Neto MFL, Silva JRV. N-acetyl-cysteine and the control of oxidative stress during in vitro ovarian follicle growth, oocyte maturation, embryo development and cryopreservation. Anim Reprod Sci. 2021;231:106801.
Yosef B, Zhou Y, Mouschouris K, Poteracki J, Soker S, Criswell T. N-Acetyl-L-cysteine reduces fibrosis and improves muscle function after acute compartment syndrome injury. Mil Med. 2020;185(Suppl 1):25–34.
Zhu QY, Tai S, Tang L, Xiao YC, Tang JJ, Chen YQ, Shen L, He J, Ouyang MQ, Zhou SH. N-acetyl cysteine ameliorates aortic fibrosis by promoting M2 macrophage polarization in aging mice. Redox Rep. 2021;26:170–5.
Honma S, Tani I, Sakai M, Soma I, Toriyabe K, Yoshida M. Effect of N-acetyl cysteine on renal interstitial fibrosis in mice. Biol Pharm Bull. 2020;43:1940–4.
Ozakpinar OB, Maurer AM, Ozsavci D. Ovarian stem cells: from basic to clinical applications. World J Stem Cells. 2015;7:757–68.
Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature. 2004;428:145–50.
Kirilly D, Xie T. The Drosophila ovary: an active stem cell community. Cell Res. 2007;17:15–25.
White YA, Woods DC, Takai Y, Ishihara O, Seki H, Tilly JL. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat Med. 2012;18:413–21.
Green SH, Zuckerman S. Further observations on oocyte numbers in mature rhesus monkeys (Macaca mulatta). J Endocrinol. 1954;10:284–90.
Sills ES, Rickers NS, Li X, Palermo GD. First data on in vitro fertilization and blastocyst formation after intraovarian injection of calcium gluconate-activated autologous platelet rich plasma. Gynecol Endocrinol. 2018;34:756–60.
Ahmadian S, Mahdipour M, Pazhang M, Sheshpari S, Mobarak H, Bedate AM, Rahbarghazi R, Nouri M. Effectiveness of stem cell therapy in the treatment of ovarian disorders and female infertility: a systematic review. Curr Stem Cell Res Ther. 2020;15:173–86.
Sheikhansari G, Aghebati-Maleki L, Nouri M, Jadidi-Niaragh F, Yousefi M. Current approaches for the treatment of premature ovarian failure with stem cell therapy. Biomed Pharmacother. 2018;102:254–62.
Cakiroglu Y, Saltik A, Yuceturk A, Karaosmanoglu O, Kopuk SY, Scott RT, Tiras B, Seli E. Effects of intraovarian injection of autologous platelet rich plasma on ovarian reserve and IVF outcome parameters in women with primary ovarian insufficiency. Aging (Albany NY). 2020;12:10211–22.
Hosseinisadat R, Farsi Nejad A, Mohammadi F. Intra-ovarian infusion of autologous platelet-rich plasma in women with poor ovarian reserve: a before and after study. Eur J Obstet Gynecol Reprod Biol. 2023;280:60–3.
Funding
FUNCAP PS1-0186–00240.01.00/21; UNIFOR 60/2022; CAPES; CNPq; FAPERGS; NIH/NIA R56AG074499.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Cavalcante, M.B., Sampaio, O.G.M., Câmara, F.E.A. et al. Ovarian aging in humans: potential strategies for extending reproductive lifespan. GeroScience 45, 2121–2133 (2023). https://doi.org/10.1007/s11357-023-00768-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-023-00768-8