
Abstract
Aging and poor nutrition are independent risk factors for the development of chronic disease. When young animals are given diets high in fat or sugar, they exhibit hallmarks of aging like mitochondrial dysfunction and inflammation, and also develop a greater risk for age-related disease. The same mitochondrial dysfunction and inflammation that progress with aging may also further predispose older individuals to dietary insults by fat and sugar. The purpose of this work is to review the most recent studies that address the impact of fat and sugar consumption on hallmarks of aging (mitochondrial dysfunction and inflammation). Findings from these studies show that obesogenic, high-fat diets can exacerbate age-related disease and hallmarks of aging in young animals, but high-fat diets that are non-obesogenic may play a beneficial role in old age. In contrast, high-sugar diets do not require an obesogenic effect to induce mitochondrial dysfunction or inflammation in young rodents. Currently, there is a lack of experimental studies addressing the impact of sugar in the context of aging, even though empirical evidence points to the detrimental effect of sugar in aging by contributing to a variety of age-related diseases.

Mitochondrial dysfunction and altered cellular communication (e.g. inflammation) progress with advancing age and increase the risk for age-related disease (ARD). Given the physiological changes that occur with age, the impact of high-fat (HFD) and high-sugar diets (HSD) may differ in later and earlier stages of life. HFD can promote the development of hallmarks of aging in young animals and can also exacerbate the risk for ARD when consumed at an old age. However, non-obesogenic high-fat diets may also reduce the risk for ARD in old age by acting on these hallmarks of aging. On the other hand, HSD promotes mitochondrial dysfunction and inflammation without necessarily inducing weight gain in young animals. Empirical evidence points to sugar as a major contributor to age-related disease and more experimental studies are needed to clarify whether aged individuals are more susceptible to its effects
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
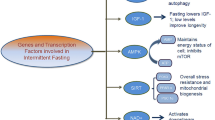
Aging is a major risk factor for developing the same chronic diseases that have been previously linked with fat and sugar consumption, such as Alzheimer’s disease (AD), cardiovascular disease (CVD), and type 2 diabetes (T2DM) [1,2,3]. Americans have been recommended to restrict their saturated fat consumption for its association with CVD since 1977 [4]. This has led to the widespread marketing strategy of labeling processed food as “low-fat” in an effort to combat metabolic disease that continues to impact people’s food choices today. There has also been growing popularity of high-fat containing-diets like ketogenic diets that stimulate debates among experts arguing against the restriction of fat [5, 6]. More recently, Americans have also been called to restrict their consumption of added sugars [7], especially as the contribution of processed foods to the American diet continues to grow in conjunction with the prevalence of metabolic disease [8]. (Fig. 1)
While there is a reasonable assumption that fat and sugar could only promote disease in a vulnerable population, the high fat content of ketogenic diets has proven beneficial in older people with cognitive decline [9, 10], high-fat feeding improves healthspan in mouse models of accelerated aging [11,12,13], and being overweight or obese has been unexpectedly linked to reduced mortality in older adults [14]. Therefore, it is important to clarify how fat and sugar may differentially affect health in advanced age, as fat and sugar could either exacerbate or perhaps reduce the risk for age-related disease and mortality. Currently, there are no differences in the recommended intakes of sugar or fat between younger and older adults. According to the Dietary Guidelines for Americans, < 30 and < 10% of energy should be coming from fats and added sugars, respectively. The main difference in dietary recommendation for people of advanced age is to reduce caloric intake as a result of decreased metabolic rate and reduced physical activity. Based on the estimated caloric needs for people over the age of 71 and sugar and saturated fat consumption values from the National Data in Usual Intakes, individuals in 75–95th percentile of intakes consumed 1.5–2.4 times more sugar and 2.0–2.8 times more saturated fat than recommended [7, 15].
This review attempts to highlight some of the recent literature examining the interactions between aging and fat or sugar consumption. There has been significant progress in nutritional interventions, such as caloric and protein restriction, to delay the process of aging, [16, 17]. Our goal is to address a different question, and that is if fat and sugar are more or less detrimental in old age. Mitochondrial dysfunction and altered cellular communication (e.g. inflammation) are two hallmarks of aging [18] that not only progress with advancing age, but are also associated with diets high in fat and sugar. Here, we will briefly provide evidence that fat and sugar consumption in youth can lead to hallmarks of aging by promoting mitochondrial dysfunction and inflammation. We will then review recent studies that address the effects of sugar and fat consumption in an aged environment to discuss the interaction between old age and high-fat or high-sugar consumption in the development of age-related diseases. We also discuss evidence showing that high-fat consumption can be both detrimental and beneficial to health based on whether it is obesogenic; whereas, the obesogenic effect is not necessary for sugar to disrupt health. By appreciating age-related differences in the effects of common components of our diet like fat and sugar, we have a better chance of improving health in advanced age.
In youth: fat and sugar contribute to hallmarks of aging and accelerate aging
The consumption of high fat diets (HFD) can recapitulate mitochondrial dysfunction and inflammation in young rodents (2–6 months old) [19,20,21,22,23], suggesting that these diets can promote hallmarks of aging in healthy, younger environment. A key consequence of HFD is the induction of obesity, which is a major risk factor for age-related disease and an important link between the effects of HFD and hallmarks of aging. Obesity is a complex disease that is influenced by factors including family history of obesity, lifestyle, that increases the risk for T2DM, CVD, and a variety of chronic diseases. Despite its complexity, obesity can be easily induced experimentally in rodents by increasing fat content in the diet, where 45–60% of energy is derived from fat (HFD-45 or HFD-60, respectively) compared to standard chow that usually contains < 5% [24]. Application of this diet strategy to induce obesity in young animals resulted in reduced mitochondrial respiration in the aorta [19], altered mitochondrial morphology in neuronal synapses [20], reduced mitochondrial density in the myocardium [21], and reduced activity and expression of mitochondrial enzymes in liver [22], myocardium [21], and skeletal muscle [25]. However, a study that differentiated the effects of caloric excess and fat content found that the excess calories from both HFD-60 and high-calorie-low-fat diet led to cardiac hypertrophy, increased malondialdehyde (oxidative stress), and reduced activity of mitochondrial complexes in the myocardium; whereas feeding fat without caloric excess did not result in any of the mentioned outcomes [26]. The composition of fat also plays a significant role in the obesogenic effect of HFD. The most used HFD diets contain lard fat that is made up of saturated long-chain triglycerides. A diet modified to have equal fat composition as HFD-45 but with most of the fat replaced by medium-chain triglycerides (HF-MCT) did not increase body weight or intrahepatic lipid compared to HFD-45. The HF-MCT increased the liver’s and adipose tissue’s ability to oxidize fat by increasing thermogenic expression for genes encoding carnitine palmitoyl acyl transferase-1 (CPT1) and peroxisome proliferator-activated receptor alpha coactivator 1-alpha (PGC-1α) in 13-week-old mice [22]. These studies support the hypothesis that obesity is a critical link for HFD to impair mitochondrial function.
High sugar diets (HSD) are less commonly used as a tool to induce mitochondrial dysfunction and oxidative stress, but sugar’s effect on tissue function should not be overlooked. Unlike HFD, the disruptions of tissue function by sugar do not always require a significant expansion of fat mass. For instance, the acute hyperglycemia that occurs after consuming 600 ml of a sweetened beverage resulted in detectable decreases in microvascular and macrovascular endothelial function in healthy 30-year-old men. To explore mechanisms, young rats were injected with glucose to induce acute hyperglycemia, which led to increased aortic reactive oxygen species (ROS) production, decreased bioavailability of nitric oxide, and a reduction in microvascular blood flow. Endothelial function was preserved in rats that were also treated with antioxidant n-acetylcysteine, suggesting the role of sugar-induced redox stress in promoting endothelial dysfunction [27]. Sugar intake, in the form of fructose, can also induce mitochondrial changes in the brain without inducing significant body weight gain or adiposity. As early as 2 weeks into consuming a diet where 30% of calories came from fructose, a reduction in citrate synthase and cytochrome oxidase was seen in the brains of young adult rats [28]. After 12 weeks of drinking water with 10%g/vol fructose, there was a sharp decrease in antioxidant enzyme activities of superoxide-dismutase 1 (SOD1) and catalase in the cortex and striatum that was associated with increased behavioral hyperactivity in 9-month-old mice [29].
Like HFD, HSD can also be obesogenic and induce mitochondrial dysfunction and oxidative stress. Consuming 30%g/vol fructose in drinking water for 5.5 months led to a 20% increase in body mass as a result of intra-abdominal fat expansion compared to chow-fed young rats [30]. Consuming 30%g/vol sucrose in drinking water also increased hydrogen peroxide generation and lipid peroxidation in the kidney [31]. Another study comparing the effects HFD and HSD (both 30%g/g or g/vol) found that despite both diets inducing similar weight gain (12%) and similar levels of hepatic fat content after 8–12 months, only HSD-induced hyperleptinemia and hyperinsulinemia in addition to increased lipogenic gene expression like sterol regulatory element-binding transcription factor 1 (SREBP1) and peroxisome proliferator-activated receptors gamma (PPARλ) increased nitrosylation of mitochondrial proteins, and increased expression proteins involved in mitochondrial fission like dynamin-related protein 1 relative to fusion protein mitofusin [32]. Another study comparing multiple diets given for 10 weeks to 6-week-old mice, found that fructose (30% g/vol) and, to a lesser extent glucose, decreased mitochondrial density and affected mitochondrial fusion and fission in the liver, when HFD-60 alone did not. Mice fed glucose and fructose were 30% heavier than controls, while the mice fed the HFD were 40–60% heavier [33]. Thus, there is a differential effect of HFD and HSD in inducing mitochondrial dysfunction in young rodents.
Inflammation is prominent in aging and leads to “altered cellular communication” that plays a role in enhanced activation of proinflammatory pathways, atherosclerosis, and decline in immune system response with aging. Both HFD and HSD can induce inflammation in a variety of tissues of young rodents, and similar to effects on mitochondria, the effects of HFD are linked to the development of obesity whereas the effects of HSD are not always associated with changes in body weight. Sixteen-week-old male mice were given either a HFD (45%kcal from fat), HSD (32%kcal from sucrose), or HFHSD for 15 weeks [34]. All diets increased body weight and heart mass to the same extent. All diets also increased gene expression of fibrosis and inflammation in heart tissue (IL1- β and TNF α). The effect of diet was absent in mice lacking the gene for NLRP3-inflammasome, suggesting that NLRP3-inflammasome inhibition may be protective against metabolic perturbations induced by both HFD and HSD. Besides the heart, HFD can induce inflammation in the lungs of very young and adult mice [35], and HFD can induce changes in the gut microbiome that are associated with changes in body weight and inflammation of colon and liver [36]. Because HFD is intended to induce obesity in most studies, it is difficult to differentiate the effects of fat from the effects of obesity on inflammation. However, HFD has been shown to reduce inflammation in brains of mice with accelerated aging and in livers of mice with complex III deficiency without inducing weight gain [13, 37].
HSD from sugar sweetened beverages (SSB) can also induce systemic and adipose inflammation. Mice that drank Coca-Cola for 13 weeks were 10% heavier than controls, and also had 37% more circulating MCP-1, and 30% greater HOMA-1R [38]. When SSB were combined with HFD, SSB exacerbated inflammatory markers suggesting an interaction between fat and sugar. In the brain, sucrose and fructose water can induce neuroinflammation and promote increases in genes associated with AD like amyloid beta even in the absence of major changes in adiposity or body weight [28, 39]. Thus, HFD and HSD can induce inflammation in young animals, and while HFD tends to be obesogenic, the HSD can promote inflammation in the absence of major weight gain.
Provided that fat and sugar promote mitochondrial dysfunction and inflammation, we looked for recent studies where HFD or HSD were given to rodents for longer than 12 months to determine if the rate of aging and development of age-related disease accelerated. Unfortunately, we were unable to find recent studies using chronic interventions of HSD. On the other hand, HFD was fed to rodents long term (16–21 months) to induce long-term obesity and better mimic age-related disease such as non-alcoholic steatohepatitis (NASH) [40], chronic kidney disease [23], and sarcopenia [23, 41]. Altogether, chronic consumption of HFD can accelerate the development of age-related tissue pathology when it is used to promote obesity, and similar studies should test the effects of chronic HSD consumption.
We described studies showing that HFD and HSD induce mitochondrial dysfunction and inflammation in young rodents. However, it is important to clarify the limitations in translating these findings to humans, as metabolic disease is also influenced by a variety of factors including physical inactivity, alcohol consumption, lack of sleep, socioeconomic status, and food selection in humans [42]. These factors can be controlled in rodent studies, but the diets used (i.e. HFD-60) are meant to rapidly induce obesity and metabolic stress by exceeding the fat content normally consumed by laboratory rodents (5%) and thus, do not fully translate to what is consumed by humans (~ 30–45%) [24]. Human diets are typically more mixed, and thus there may be an interactive effect between fats and sugars, or even a tertiary dietary component that may not appear in these controlled rodent studies dedicated to either sugar or fat. Studies using HF in conjunction with HS show more metabolic defects than HF or HS alone [34]. Rodent studies on sugar consumption are less consistent in the strategy used to add sugar, with some replacing complex carbohydrates for sugar in the diet, others replacing drinking water with sugar water, and some using different sweeteners (fructose vs. glucose). However, the use of sucrose, high fructose corn syrup, or commercially available SSBs increase the translation potential of these studies, as these are the same added sugars that make up a variety of processed foods in the human diet.
In the context of aging: effects of fat
The likelihood of getting a disease like CVD, T2DM, or AD increases with the number of risk factors present in addition to aging, such as genetics, the environment, and diet. In the laboratory where risks-factors can be better controlled, HFD has been used as a “third hit” by inducing obesity in aged animals when a stressor is insufficient to recapitulate the pathophysiology seen clinically. To find a model for heart failure with preserved ejection fraction (HFpEF), 18–22-month-old female mice were given a HFD-60 for 12 weeks to induce obesity and treated with angiotensin II (ANGII). Without ANGII treatment, HFD caused enlargement of the left ventricle and increased the aortic pressure gradient. However, it did not lead to fibrosis, a reduction in capillary density, nor gene expression of hypertrophy and fibrotic genes until animals were also treated with ANGII [43]. Another study using 16-month-old rats, found that providing HFD-45 for 6 months led to increased body weight and mild cardiac hypertrophy without changes in diastolic or systolic function in normotensive rats. However, the effect of HFD-45 exacerbated cardiac pathology in hypertensive rats, with detectable increase in left ventricular mass, decrease in ejection fraction, as well as a sharp increase in atrial expression of inflammatory and hypertrophic genes [44]. In addition to cardiovascular pathology, HFD can also promote retinal pathology to mimic age-related macular degeneration (AMD), the leading cause of blindness in the elderly. Young (7-month-old) and old (20-month-old) transgenic mice expressing either the AMD-associated variant or the full CFH variant (normal) were provided with a HFD + cholesterol diet, and only old mice with the AMD-associated variant exhibited decreased visual function and retinal pigment epithelium stress [45]. These studies show that HFD-induced obesity can act as a metabolic stressor in aged animals but may require additional stressors to lead to clinically relevant disease models.
The sensitivity to insults in old age by obesogenic-HFD can be attributed to low grade chronic inflammation that progresses with aging. For instance, fibrosis of the kidney was exacerbated in old rats after only 15 days of consuming HFD-60 [46]. Young rats did not show any evidence of fibrosis or any changes in the proteins involved in mitochondrial respiration, inflammation, or lipid metabolism with HFD, whereas old rats, which started with elevated inflammation to begin with, had the highest protein levels of p-p65, COX-2, and iNOS, and the lowest content of lipid oxidation proteins like CPT1α. This finding highlights a vulnerability to metabolic insults by HFD that is specific to the aged kidney. Another study using a short-term diet intervention found that 3 days of HFD-60 led to 4% increase in body weight in 24-month-old rats and 2% increase in body weight in young 3-month-old. It also led to cognitive deficits in old but not young rats, specifically contextual memory and cued-fear memory. HFD only increased interleukin 1 beta (IL1ß) expression in the hippocampus and amygdala of old but not younger rats [47], which suggests that the old brain is particularly susceptible to inflammation by HFD that is associated with impaired memory. Similar findings were found by a different group, where only old rats (23 months) exhibited increased inflammatory markers like CD68 in the hippocampus and hypothalamus after 12 weeks of HFD-60 compared to a middle-aged (13 months) and young group (3 months) and were also the only group with a significant gain in body weight with HFD [48]. Behaviorally, the HFD increased the number of errors in a Barnes maze probe trial in the old and middle-aged rats but not young, indicating an HFD-dependent decline in spatial learning and memory that was unique to aged groups. Similar susceptibilities of old rodents to HFD have been shown in the vasculature. To determine if hyperlipidemia would increase the susceptibility of aged mice to atherosclerosis compared to young mice, 2–3-month-old and 18–19-month-old C57/BL6 mice were given AAV-Pcsk9 injection to degrade lipoprotein lipase receptor and induce hypercholesterolemia in addition to a HFD-45 for 10 weeks [19]. Both young and old mice developed similar degrees of weight gain, fat mass, hypercholesterolemia, and circulating monocytes. However, only aged mice showed an amplified atherosclerotic response with larger and more necrotic arterial lesions and impaired aortic mitochondrial function. The authors showed that the elevation of IL-6 seen with aging, primed the vasculature to atherosclerosis when presented with a hypercholesterolemic insult. Thus, the progression of inflammation with age predisposes old animals to metabolic insults by an obesogenic HFD.
Despite old age amplifying the stress induced by HFD-induced obesity, fat consumption may promote some beneficial effects in context of aging in both animals and humans. These studies often modify the timing, fat source, or composition of diet that reduces their obesogenic effect. One study used self-foraging of sunflower seeds to increase fat intake in 16 month-old mice for 9 weeks, which led to a smaller increase in body weight (24%) compared to commonly used HFD-60 (40–60%) [49]. Old-HFD mice showed improvements in locomotory muscle function, specifically soleus and extensor digitorium longus, evidenced by increased muscle mass, cross-sectional area, and power output. Another study tested the effects of ad libitum HFD-75 versus consuming it every other week starting at 12 months old until death. Feeding a cyclic-HFD-75 prevented obesity and tended to reduce mid-life mortality compared to CHOW. The opposite was true in mice fed HF-75 ad libitum who lived shorter lives [50]. The timing of HFD to prevent obesity has also been tested in old humans. In a cross-over design, older adults (55–75 years old) were given fat-based breakfasts (45%kcal from fat, 35% from carbohydrate) or carbohydrate-based breakfast (20%kcal from fat, and 60%kcal from carbohydrate) for 4 weeks. This led to an increase in total fat consumption of only 10% in the fat-based breakfast group compared to carbohydrate-based breakfast group and did not increase body weight nor plasma cholesterol. However, eating a fat-based breakfast was enough to lower 24-h respiratory quotient (RQ) and reduce c-reactive protein that is indicative of increased whole body fat oxidation and lower inflammation, respectively [51]. In obesity and in conditions of insulin resistance, the rates of far oxidation by skeletal muscle decrease [52], therefore, increasing fat oxidation throughout the day may be beneficial for overweight and obese older adults to prevent further weight gain. Recommending fat consumption to older, overweight or obese individuals sounds counterintuitive, but it has been shown in a 5-year longitudinal randomized control trial that included over 4000 women and 3000 men who were obese and overweight that Mediterranean diets rich in healthy fats (olive oil and nuts) (~ 40% kcal from fat) do not increase body weight throughout a 5-year period and in fact promote better weight maintenance than the group consuming a control diet (< 30% kcal from fat) and could thus be beneficial in reducing risk for cardiovascular events in people over 65 [53].
Ketogenic diets (KD) sit in a different realm than the commonly used high fat diets; nevertheless, it is relevant to discuss them as these diets have the highest fat composition tested, with > 60% of calories deriving from fat and almost no calories from carbohydrate to induce ketosis (a rise in blood ketones). Unlike HFD, KD is not given with the purpose to induce obesity. To prevent obesity, KDs can be given in a cyclic manner (every other week) [50], or by controlling the calories consumed in order to see benefits from fat intake. It was demonstrated by Newman et al. that mice fed ad libitum KD-85 had increased rates of mortality, but mice fed a cyclic-KD had reduced mid-life mortality and did not develop obesity seen in ad libitum feeders, suggesting that fat intake can be beneficial when it does not promote obesity. Studies testing the effects of KD in older people tend to report benefits, particularly in the aging brain and in neurological disease [9, 54, 55]. For instance, Parkinson’s patients aged 60 ± 7 years old demonstrated greater improvements in non-motor symptoms (MDS-UPDRS scores) after consuming a KD for 8 weeks compared to those consuming a low-fat-high-carbohydrate diet, despite both groups having similar reductions in body weight, plasma triglycerides, and HbA1C [56]. A recent study also showed that a ketogenic formula made up of medium chain triglycerides for 12 weeks had positive effects on working memory, short-term memory, and processing in 20 older Japanese adults (73.4 ± 6.0 years old) with mild to moderate AD, although results should be interpreted with caution as there was no placebo group [57]. A pilot trial found that KD intervention of 3 months in participants with AD (age 73.1 ± 9.0) improved cognition scores without increasing body weight, blood lipid profile, or impairing metabolic status (HOMAIR, blood glucose/insulin). After a 1-month wash-out period, cognition scores returned to baseline suggesting that the improvements may have been due to the KD diet itself, but a larger study is needed to confirm their findings [10]. Findings are consistent with animal studies using isocaloric KD or cyclic-KD that show improvements in cognition without weight gain [50, 58]. KD is also contraindicated in a variety of conditions that are common with aging, such as cardiac arrhythmias, frailty, heart failure, recent stroke or myocardial infarction, liver failure, and kidney disease. In fact, 16 weeks of KD consumption promoted cardiac pathology in young rats by impairing mitochondria and promoting cardiac fibrosis as a direct result from the elevation of ketone beta-hydroxybutyrate [59]. The evidence supporting these contraindications has been recently reviewed by Watanabe et al., and suggests that there is a no definitive conclusion as to the harm of KD in the presence of these conditions, but providing KD to fragile population should be approached with caution [60], and further research is needed to determine KD’s benefits in vulnerable populations like seniors.
In the context of aging: effects of sugar
The benefits of fat cannot be purely attributed to increasing fat intake as there is usually a concomitant decrease in carbohydrate intake. For instance, a carbohydrate-based breakfast led to increased RQ that was positively correlated with higher fasting glucose and insulin, and reduced insulin sensitivity [51]. The authors suggested that a carbohydrate-based breakfast may lead to greater metabolic perturbations than a fat-based breakfast in older adults. On the other hand, consuming a low-fat-high-carbohydrate diet still improved MDS-UPDRS scores in older individuals with Parkinson’s Disease, though less so than KD [56]. To clarify, carbohydrates are the primary source of energy for the human body, and starches and whole grains are considered to be part of a healthy diet. However, there is growing evidence that added sugars (sucrose and high-fructose-corn-syrup) contribute to disease and may exacerbate age-related disease.
Compared to the published work on HFD, there are very few experimental studies that have tested the effects of sugar in the context of aging, although empirical evidence suggests that sugar intake is implicated in increased risk of age-related diseases. A report from Centers for Disease Control and Prevention provided strong evidence that added sugar consumption increased the risk of CVD-mortality in the US population, and that 71% of the population exceeded the recommended intake [2]. The hazard ratio was 1.83 and 1.67 for people over and under 60, respectively. A recent study also found that SSB consumption increased the risk for congestive heart failure in African Americans independent of risk factors like age or BMI [61]. An opposite association was found between sugar intake and CVD risk in older adults living in Hong Kong, where there was a lower risk for CVD with increased sugar consumption; though the average sugar intake for this population was 3%, and only 6% for the highest quintile with the lowest risk of CVD, suggesting the caution that a diet too low in sugar (less than 3% calories from sugar) may also be unfavorable in old age [62]. Empirical evidence also shows that blood pressure is associated with sugar consumption in women between 65 and 80 years old after adjusting for BMI. It was concluded that reducing sugar intake by 2.3 tsp would result in a drop in systolic and diastolic blood pressure of 8.4 mmHg and 3.7 mmHg, respectively [63]. Also in women > 60 years old who participated in the Nurse’s Health Study an increase of 1 serving/day of SSB were 1.3 times more likely to have frailty (fatigue, poor strength, reduced aerobic capacity, having several chronic illnesses, and significant weight loss during the previous year) after adjusting for BMI [64]. Finally, it has been hypothesized that fructose metabolism plays a causative role in the development of AD by decreasing mitochondrial function and cerebral energy levels [3]. A recent study also found that simple sugars did not increase inflammatory markers in plasma or adipose tissue of 65-year-old adults after 4.5 weeks compared to refined and unrefined carbohydrates, suggesting that the effects of sugar on health may not be solely dependent on inflammation [65].
Based on what we know about fat and sugar consumption during youth and fat consumption during old age, we can make some hypotheses as to sugar’s effect in the context of aging. Sugar and fat induced mitochondrial dysfunction and inflammation in young animals, but the effects of fat were associated with obesity whereas the effects of sugar were not. Obesogenic fat diets can exacerbate inflammation and increase the risk for disease in advanced age, but non-obesogenic fat can also decrease risk for age-related disease when it does not cause weight gain. Non-obesogenic fat consumption can promote the expression of genes required to oxidize fat resulting in mitochondrial biogenesis, thermogenesis, and increased fatty acid oxidation enzymes like CPT1. So, having the ability to oxidize fatty acids while preventing excess weight gain would reduce stress on other organs like liver, adipose tissue, and skeletal muscle and thus prevent the inflammation commonly seen in obesity. Unlike fat, sugar does not promote fatty acid oxidation and instead enhances de novo lipogenesis by activating SREBP, carbohydrate response element binding protein (ChREBP) in the liver, and acetate production by microbiota in the gut [66]. In addition, sugar promotes inflammation by damaging the intestinal barrier [67] and contributes to increases in post-translational modifications of proteins such as O-GlcNAcylation that increase cytokine production [68] and advanced-glycated-end products (AGEs) that contribute to inflammation, suppress sirtuins, and promote oxidative stress [69]. Since aged individuals already have higher levels of inflammation, AGEs, and mitochondrial dysfunction, and sugar does not appear to improve but only worsen these factors, it is reasonable to hypothesize that excess sugar could be particularly detrimental in the context of aging independent of obesity.
Conclusion
The rise in fat and sugar consumption has undoubtedly contributed to the rise in non-communicable disease and mortality in recent years. However, dietary guideline recommendations for adults as young as 18 years old and older individuals are almost identical [7]. Only 8% of published literature on dietary policies and programs in the United States has focused on older adults (> 65 years) [70]. While healthy behaviors such as physical activity and eating a balanced nutritious diet are recognized to benefit all age groups, the focus of fat and sugar continue to influence food guidelines, marketing, and food choices today [71]. A recent review of dietary guidelines worldwide found that most graphics from 78 countries conveyed the restriction of sugar and fats/oils [72]. Thus, we focused this review on summarizing some of the recent evidence on the influence of fat and sugar in hallmarks of aging.
Experimental studies have been able to show that old rodents can be more susceptible to mitochondrial dysfunction and inflammation with HFD-induced obesity, partly due to low grade chronic inflammation in advanced age. However, most evidence comes from studies on obesity, which is recognized to be a metabolic condition also influenced by a variety of factors (i.e. lower physical activity) and may not be a direct result of solely fat consumption. In fact, HFD can also be controlled or modified, such as in a cyclic diet or high intake of healthy fats so it does not promote obesity and is potentially beneficial in the context of aging, especially to the aging brain, although more research is needed to determine their safety in older populations. Nevertheless, the trends on fat consumption have gone to extremes (low to very high fat), and more research is necessary to (1) assess whether reasonable, sustainable levels of fat consumption may be particularly beneficial to the older population given the benefits seen with HFD, and (2) how the call for fat restriction could be especially harmful in older age.
On the other hand, added sugar does not appear to be beneficial in youth or in old age as it induces mitochondrial dysfunction and inflammation, even in the absence of weight gain. Sugar consumption does not reliably induce weight gain in humans or mice, which may be a reason why it is less often used as a metabolic stressor like HFD in experimental studies. There is a significant gap in the literature regarding the effects of sugar in old age, despite epidemiological evidence of sugar accelerating the development of conditions associated with aging (e.g. frailty, CVD). Future research should focus on the effects of high sugar intake in older humans and rodents compared to younger counterparts, and determine whether the aged population is more susceptible to mortality and morbidity by this ubiquitous food additive.
Data availability
Review does not include any unpublished data. All work presented is available to the public.
Code availability
N/A
References
Micha R, et al. Association between dietary factors and mortality from heart disease, stroke, and type 2 diabetes in the United States. JAMA. 2017;317(9):912–24.
Yang Q, et al. Added sugar intake and cardiovascular diseases mortality among US adults. JAMA Intern Med. 2014;174(4):516–24.
Johnson RJ, et al. Cerebral fructose metabolism as a potential mechanism driving Alzheimer’s disease. Front Aging Neurosci. 2020;12:560865.
Harcombe Z, et al. Evidence from randomised controlled trials did not support the introduction of dietary fat guidelines in 1977 and 1983: a systematic review and meta-analysis. Open Heart. 2015;2(1):e000196.
Ludwig DS, et al. Dietary fat: from foe to friend? Science. 2018;362(6416):764–70.
Astrup A, et al. Saturated fats and health: a reassessment and proposal for food-based recommendations: JACC State-of-the-Art Review. J Am Coll Cardiol. 2020;76(7):844–57.
Agriculture, U.S.D.o.H.a.H.S.a.U.S.D.o., 2010–2015 Dietary Guidelines for Americans. December 2015.
Martínez Steele E, et al. Dietary share of ultra-processed foods and metabolic syndrome in the US adult population. Prev Med. 2019;125:40–8.
Morrill SJ, Gibas KJ. Ketogenic diet rescues cognition in ApoE4+ patient with mild Alzheimer’s disease: a case study. Diabetes Metab Syndr. 2019;13(2):1187–91.
Taylor MK, et al. Feasibility and efficacy data from a ketogenic diet intervention in Alzheimer’s disease. Alzheimers Dement (N Y). 2018;4:28–36.
Oike H, Ogawa Y, Azami K. Long-term feeding of a high-fat diet ameliorated age-related phenotypes in SAMP8 mice. Nutrients. 2020;12(5).
Li Z, et al. A high-fat diet reverses metabolic disorders and premature aging by modulating insulin and IGF1 signaling in SIRT6 knockout mice. Aging Cell. 2020;19(3):e13104.
Purhonen J, et al. Ketogenic diet attenuates hepatopathy in mouse model of respiratory chain complex III deficiency caused by a Bcs1l mutation. Sci Rep. 2017;7(1):957.
Javed AA, et al. Body mass index and all-cause mortality in older adults: a scoping review of observational studies. Obes Rev. 2020;21(8):e13035.
Institute, N.C., Usual Dietary Intakes: Food Intakes, U.S. Population, 2007 – 10. 2019.
Balasubramanian P, Mattison JA, Anderson RM. Nutrition, metabolism, and targeting aging in nonhuman primates. Ageing Res Rev. 2017;39:29–35.
Hill CM, Kaeberlein M. Anti-ageing effects of protein restriction unpacked. Nature. 2021;589(7842):357–8.
López-Otín C, et al. The hallmarks of aging. Cell. 2013;153(6):1194–217.
Tyrrell DJ, et al. Age-associated mitochondrial dysfunction accelerates atherogenesis. Circ Res. 2020;126(3):298–314.
Martins IV, et al. Mitochondrial abnormalities and synaptic loss underlie memory deficits seen in mouse models of obesity and Alzheimer’s disease. J Alzheimers Dis. 2017;55(3):915–32.
Chen D, et al. A high-fat diet impairs mitochondrial biogenesis, mitochondrial dynamics, and the respiratory chain complex in rat myocardial tissues. J Cell Biochem. 2018;119(11):9602.
Rial SA, et al. A high-fat diet enriched in medium chain triglycerides triggers hepatic thermogenesis and improves metabolic health in lean and obese mice. Biochim Biophys Acta Mol Cell Biol Lipids. 2020;1865(3):158582.
Laurentius T, et al. High-fat diet-induced obesity causes an inflammatory microenvironment in the kidneys of aging Long-Evans rats. J Inflamm (Lond). 2019;16:14.
Speakman JR. Use of high-fat diets to study rodent obesity as a model of human obesity. Int J Obes (Lond). 2019;43(8):1491–2.
Choi JW, et al. Carnitine induces autophagy and restores high-fat diet-induced mitochondrial dysfunction. Metabolism. 2018;78:43–51.
Emelyanova L, et al. High calories but not fat content of lard-based diet contribute to impaired mitochondrial oxidative phosphorylation in C57BL/6J mice heart. PLoS ONE. 2019;14(7):e0217045.
Loader J, et al. Effects of sugar-sweetened beverage consumption on microvascular and macrovascular function in a healthy population. Arterioscler Thromb Vasc Biol. 2017;37(6):1250–60.
Cigliano L, et al. Short-term fructose feeding induces inflammation and oxidative stress in the hippocampus of young and adult rats. Mol Neurobiol. 2018;55(4):2869–83.
Dos Santos B, et al. Fructose intake impairs cortical antioxidant defenses allied to hyperlocomotion in middle-aged C57BL/6 female mice. Neurochem Res. 2020;45(12):2868–83.
Żebrowska E, et al. High-sugar diet disrupts hypothalamic but not cerebral cortex redox homeostasis. Nutrients. 2020;12(10).
Ruiz-Ramírez A, et al. Kidney dysfunction induced by a sucrose-rich diet in rat involves mitochondria ROS generation, cardiolipin changes, and the decline of autophagy protein markers. Am J Physiol Renal Physiol. 2020;318(1):F53–66.
Cruz Hernández, J.H., et al., High sugar but not high fat diet consumption induces hepatic metabolic disruption and up-regulation of mitochondrial fission-associated protein Drp1 in a model of moderate obesity. Arch Physiol Biochem, 2020: p. 1–8.
Softic S, et al. Dietary sugars alter hepatic fatty acid oxidation via transcriptional and post-translational modifications of mitochondrial proteins. Cell Metab. 2019;30(4):735-753.e4.
Pavillard LE, et al. NLRP3-inflammasome inhibition prevents high fat and high sugar diets-induced heart damage through autophagy induction. Oncotarget. 2017;8(59):99740–56.
Hegab AE, et al. High fat diet activates adult mouse lung stem cells and accelerates several aging-induced effects. Stem Cell Res. 2018;33:25–35.
Kim SJ, et al. Dietary fat intake and age modulate the composition of the gut microbiota and colonic inflammation in C57BL/6J mice. BMC Microbiol. 2019;19(1):193.
Lam, V., et al., Chronic high fat feeding paradoxically attenuates cerebral capillary dysfunction and neurovascular inflammation in senescence-accelerated-murine-prone strain 8 mice. Nutr Neurosci, 2019: p. 1–9.
Lee G, et al. Three-month daily consumption of sugar-sweetened beverages affects the liver, adipose tissue, and glucose metabolism. J Obes Metab Syndr. 2020;29(1):26–38.
Yeh SH, et al. A high-sucrose diet aggravates Alzheimer’s disease pathology, attenuates hypothalamic leptin signaling, and impairs food-anticipatory activity in APPswe/PS1dE9 mice. Neurobiol Aging. 2020;90:60–74.
Velázquez KT, et al. Prolonged high-fat-diet feeding promotes non-alcoholic fatty liver disease and alters gut microbiota in mice. World J Hepatol. 2019;11(8):619–37.
Laurentius T, et al. Long-chain fatty acids and inflammatory markers coaccumulate in the skeletal muscle of sarcopenic old rats. Dis Markers. 2019;2019:9140789.
Remus Popa A, et al. Risk factors for adiposity in the urban population and influence on the prevalence of overweight and obesity. Exp Ther Med. 2020;20(1):129–33.
Withaar, C., et al., The effects of liraglutide and dapagliflozin on cardiac function and structure in a multi-hit mouse model of heart failure with preserved ejection fraction. Cardiovasc Res, 2020.
Shiou YL, et al. High fat diet aggravates atrial and ventricular remodeling of hypertensive heart disease in aging rats. J Formos Med Assoc. 2018;117(7):621–31.
Landowski M, et al. Human complement factor H Y402H polymorphism causes an age-related macular degeneration phenotype and lipoprotein dysregulation in mice. Proc Natl Acad Sci U S A. 2019;116(9):3703–11.
Ha S, et al. Short-term intake of high fat diet aggravates renal fibrosis in aged Sprague-Dawley rats. Exp Gerontol. 2020;142:111108.
Spencer SJ, et al. High-fat diet and aging interact to produce neuroinflammation and impair hippocampal- and amygdalar-dependent memory. Neurobiol Aging. 2017;58:88–101.
Moser VA, et al. Effects of aging, high-fat diet, and testosterone treatment on neural and metabolic outcomes in male brown Norway rats. Neurobiol Aging. 2019;73:145–60.
Hill C, et al., Does dietary-induced obesity in old age impair the contractile performance of isolated mouse soleus, extensor digitorum longus and diaphragm skeletal muscles? Nutrients. 2019;11(3).
Newman JC, et al. Ketogenic diet reduces midlife mortality and improves memory in aging mice. Cell Metab. 2017;26(3):547-557.e8.
Bush NC, et al. A high-fat compared with a high-carbohydrate breakfast enhances 24-hour fat oxidation in older adults. J Nutr. 2018;148(2):220–6.
Kelley DE, Simoneau JA. Impaired free fatty acid utilization by skeletal muscle in non-insulin-dependent diabetes mellitus. J Clin Invest. 1994;94(6):2349–56.
Estruch R, et al. Effect of a high-fat Mediterranean diet on bodyweight and waist circumference: a prespecified secondary outcomes analysis of the PREDIMED randomised controlled trial. Lancet Diabetes Endocrinol. 2019;7(5):e6–17.
Augustin K, et al. Mechanisms of action for the medium-chain triglyceride ketogenic diet in neurological and metabolic disorders. Lancet Neurol. 2018;17(1):84–93.
Vinciguerra, F., et al., Influence of the Mediterranean and ketogenic diets on cognitive status and decline: a narrative review. Nutrients. 2020;12(4).
Phillips MCL, et al. Low-fat versus ketogenic diet in Parkinson’s disease: a pilot randomized controlled trial. Mov Disord. 2018;33(8):1306–14.
Ota M, et al. Effects of a medium-chain triglyceride-based ketogenic formula on cognitive function in patients with mild-to-moderate Alzheimer’s disease. Neurosci Lett. 2019;690:232–6.
Roberts MN, et al. A ketogenic diet extends longevity and healthspan in adult mice. Cell Metab. 2017;26(3):539-546.e5.
Xu S, et al. Ketogenic diets inhibit mitochondrial biogenesis and induce cardiac fibrosis. Signal Transduct Target Ther. 2021;6(1):54.
Watanabe M, et al. Scientific evidence underlying contraindications to the ketogenic diet: an update. Obes Rev. 2020;21(10):e13053.
DeChristopher LR, Auerbach BJ, Tucker KL. High fructose corn syrup, excess-free-fructose, and risk of coronary heart disease among African Americans- the Jackson Heart Study. BMC Nutr. 2020;6(1):70.
Liu ZM, et al. Dietary sugar intake was associated with increased body fatness but decreased cardiovascular mortality in Chinese elderly: an 11-year prospective study of Mr and Ms OS of Hong Kong. Int J Obes (Lond). 2018;42(4):808–16.
Mansoori, S., et al., Added sugar intake is associated with blood pressure in older females. Nutrients. 2019;11(9).
Struijk EA, et al. Sweetened beverages and risk of frailty among older women in the Nurses’ health study: a cohort study. PLoS Med. 2020;17(12):e1003453.
Meng H, et al. Effect of dietary carbohydrate type on serum cardiometabolic risk indicators and adipose tissue inflammatory markers. J Clin Endocrinol Metab. 2018;103(9):3430–8.
Zhao S, et al. Dietary fructose feeds hepatic lipogenesis via microbiota-derived acetate. Nature. 2020;579(7800):586–91.
Todoric J, et al. Fructose stimulated de novo lipogenesis is promoted by inflammation. Nat Metab. 2020;2(10):1034–45.
Banerjee PS, Lagerlöf O, Hart GW. Roles of O-GlcNAc in chronic diseases of aging. Mol Aspects Med. 2016;51:1–15.
Nowotny K, et al. Dietary advanced glycation end products and their relevance for human health. Ageing Res Rev. 2018;47:55–66.
Russo R, et al. Dietary policies and programs in the United States: a narrative review. Prev Med Rep. 2020;19:101135.
Kuster, I. and N. Vila, Healthy lifestyle and eating perceptions: correlations with weight and low-fat and low-sugar food consumption in adolescence. 2017, Taylor & Francis: Frontiers in Life Science. p. 48–62.
Herforth A, et al. A global review of food-based dietary guidelines. Adv Nutr. 2019;10(4):590–605.
Acknowledgements
We thank Jeffrey Leung for his help in the literature review.
Funding
The work was supported by P.S.R. and D.J.M. (P01-AG001751-36), A.P.V. (T32-HL007028-43).
Author information
Authors and Affiliations
Contributions
A.P.V conceived the original idea and wrote the manuscript with input from all authors. A.P.V, N.N. and D.O. conducted literature search and discussed relevant articles. D.J.M. helped plan and revise the manuscript and D.J.M. and P.S.R. secured funding.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
A.P.V., D.J.M., N.N., D.H.O. declare that this was all their original work. No experiments were conducted for this project.
Consent for publication
Authors A.P.V., P.S.R., D.J.M., N.N., D.H.O. give consent of publication.
Conflict of interest
Authors A.P.V., P.S.R., D.J.M., N.N., and D.H.O. declare no conflicts of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Valencia, A.P., Nagaraj, N., Osman, D.H. et al. Are fat and sugar just as detrimental in old age?. GeroScience 43, 1615–1625 (2021). https://doi.org/10.1007/s11357-021-00390-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11357-021-00390-6