
Abstract
Pesticides are commonly found in the environment and pose a risk to target and non-target species; therefore, employing a set of bioassays to rapidly assess the toxicity of these chemicals to diverse species is crucial. The toxicity of nine individual pesticides from organophosphate, organochlorine, phenylurea, dinitroaniline, carbamate, and viologen chemical classes and a mixture of all the compounds were tested in three bioassays (Hydra vulgaris, Lemna minor, and Caenorhabditis elegans) that represent plant, aquatic, and soil-dwelling species, respectively. Multiple endpoints related to growth and survival were measured for each model, and EC10 and EC50 values were derived for each endpoint to identify sensitivity patterns according to chemical classes and target organisms. L. minor had the lowest EC10 and EC50 values for seven and five of the individual pesticides, respectively. L. minor was also one to two orders of magnitude more sensitive to the mixture compared to H. vulgaris and C. elegans, where EC50 values were calculated to be 0.00042, 0.0014, and 0.038 mM, respectively. H. vulgaris was the most sensitive species to the remaining individual pesticides, and C. elegans consistently ranked the least sensitive to all tested compounds. When comparing the EC50 values across all pesticides, the endpoints of L. minor were correlated with each other while the endpoints measured in H. vulgaris and C. elegans were clustered together. While there was no apparent relationship between the chemical class of pesticide and toxicity, the compounds were more closely clustered based on target organisms (herbicide vs insecticide). The results of this study demonstrate that the combination of these plant, soil, and aquatic specie can serve as representative indicators of pesticide pollution in environmental samples.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pesticides are diverse classes of chemicals that are intentionally released into the environment to control unwanted plants, insects, fungi, and other organisms (Fujita et al. 2010; Hernández et al. 2013; Warne and Reichelt-Brushett 2023). However, only 1% of the applied chemicals have been estimated to effectively control the target organisms, and the remaining residues enter the environment where they can interact with non-target organisms (Grégoire et al. 2022; Hernández et al. 2013; Tudi et al. 2021; Warne and Reichelt-Brushett 2023). Considering the leaching, runoff from agricultural lands, and spray drifting that can occur after applications, humans, plants, and other organisms living in the surrounding environment are at risk of exposure and possible toxicity (Cunha et al. 2012; Leu et al. 2004; Sharma et al. 2019). For example, aquatic species, bees, and earthworms living near citrus (Cunha et al. 2012) and plum orchards (Reinecke and Reinecke 2007) have been shown to have elevated risks of toxicity due to spray drifting after pesticide applications. Membrane devices deployed into streams that drain from two golf courses also showed significant toxicity to a fish species, which correlated with the application schedule of chlorinated pesticides on the courses (Metcalfe et al. 2008). Previous studies have also demonstrated that exposure to just 1% of the recommended field dose of metsulfuron methyl and chlorsulfuron was linked to severe growth inhibition of aquatic plant species, food crops (Boutin et al. 2000), and fruit trees (Bhatti et al. 1995; Cedergreen and Streibig 2005). These studies demonstrate that even low levels of pesticides in the environment pose a significant risk to the ecosystem.
The global application of all pesticides was approximately 4.2 million tons in 2019 (Warne and Reichelt-Brushett 2023), and over 1000 individual chemicals are currently registered for use worldwide (Johansen 2003; Schwingl et al. 2021; WHO 2022). Pesticides account for roughly 20% of the compounds listed on the Agency for Toxic Substances and Disease Registry Substance Priority List (SPL) (Simonsen et al. 2008), where nearly 75% and 17% of the listed pesticides belong to the legacy organochlorine and organophosphate chemical classes, respectively (ATSDR 2022). Because of the complexity of these existing applications, presence at priority sites, and the development of new agrochemicals over time, there is a critical need for bioassay methods that can quickly assess the toxicity of individual compounds and complex mixtures to target and non-target species (Fochtman et al. 2000; Ullah and Zorriehzahra 2015). To assess the toxicity of individual pesticides and mixtures, the use of several bioassays from similar ecological niches (e.g., a set of aquatic organisms) or different groups (e.g., microorganisms, aquatic species, and other invertebrates) have been used (Hernando et al. 2003; Nowell et al. 2014). Utilizing a battery of bioassays in hazard assessment is advantageous as a single model organism cannot accurately nor reliably depict the toxicity of a chemical or represent the whole environment (Mariani et al. 2006; Repetto 2013).
Hydra vulgaris is a freshwater cnidarian that is used to assess the toxicity of contaminants to aquatic species because (1) hydra species play an important role in their ecosystem, (2) it has well-described morphological endpoints, (3) and is sensitive to environmental contaminants (Galliot 2012; Karntanut and Pascoe 2000). Lemna minor is a floating macrophyte that is used to represent aquatic plants in toxicity testing because (1) macrophytes play an important ecological role as primary producers (de Alkimin et al. 2020), (2) its ease of cultivation, (3) has a high reproduction rate that yields a genetically homogenous population, and (4) is sensitive to pesticides (Aliferis et al. 2009; Wang 1990). Caenorhabditis elegans is a soil nematode that is useful for understanding the toxicity of pesticides to soil species because (1) nematodes account for the largest number of soil-dwelling species and are essential for maintaining soil quality (Sochová et al. 2006), (2) its ease of cultivation, and (3) has diverse histological, biochemical, and behavioral endpoints that are available to characterize chemical toxicity (Hunt 2017). These bioassays were utilized to represent important ecological niches in aquatic and soil environments that contain aquatic, plant, and soil invertebrate species that can be directly or indirectly affected by pesticides. These bioassays are advantageous for toxicity testing of environmental chemicals like pesticides because they combine the whole-organism approach of in vivo methods and the scalability and throughput of smaller, cell-based, in vitro systems (Hunt 2017; Ueda and Nagai 2021).
In the current study, nine pesticides from important chemical classes, including organochlorines, organophosphates, urea-type, dinitroanilines, carbamates, and viologens (Table 1), were chosen to represent diverse compounds with varying ranges of toxicities and target organisms. As the SPL is not a list of the most toxic compounds, chemicals that are not currently listed were also included in the study to further represent compounds that have environmental and human relevance. We also included a pesticide mixture because organisms are exposed to a complex mixture of chemicals in the environment, and the toxicities of these mixtures are typically different than the individual compounds. For example, interactions between chemicals in the mixtures may act synergistically or antagonistically to alter the sample toxicity (Bart et al. 2022; Hernández et al. 2013; Hernando et al. 2003; Rider and Simmons 2018; Wang et al. 2021).
The goals of the current study were to assess the toxicity of diverse chemical classes of insecticides and herbicides to a battery of bioassays and use this information to rapidly (1) compare the sublethal responses of the bioassays and (2) determine if specific classes of pesticides are more toxic to the current set of bioassays than others. These results will be used to identify sensitive endpoints for each bioassay that can be used to rapidly assess and predict the toxicity of new pesticides, complex mixtures, and environmental samples to monitor soil and water quality and verify remediation efforts.
Materials and methods
Reagents and materials
Nine pesticides that represent different chemical classes, target organisms, and physiochemical properties were included in the present study (Table 1) (PubChem, NIH). These pesticides also have varying LogP values, molecular weights (MW), and water solubilities, which are known to be important factors for chemical toxicity and bioaccumulation factors (Fujita et al. 2010; Silva et al. 2022). Crystalline 2,4,6-trichlorophenol, pentachlorophenol, lindane, diazinon, glyphosate, linuron, aldicarb, and paraquat dichloride were purchased from Sigma-Aldrich (St. Louis, MO). Trifluralin was purchased from ChemServ (Minneapolis, MN). Stock concentrations of individual pesticides were created by dissolving each chemical in water (glyphosate, paraquat, aldicarb) or 100% acetonitrile, which was chosen based on the chemical’s water solubility (Table 1) (PubChem, NIH) to prevent chemical precipitation during the experiments. The pesticide mixture was created by dissolving 2 mg/mL of each chemical into 100% acetonitrile. Cell culture grade dimethyl sulfoxide (DMSO) and HPLC grade acetonitrile were purchased from Sigma-Aldrich.
Hydra vulgaris assay
A population of H. vulgaris polyps was obtained from Environment Canada (Montreal, Qc) and was maintained in hydra medium according to established methods (Wang et al. 2021). For toxicity analysis, non-budding polyps were exposed to increasing concentrations of each pesticide or the pesticide mixture (Supplemental Fig. S1) prepared in 4 mL of hydra medium. Each pesticide was tested in at least four concentrations. Over 92 h, the morphology of the polyps was scored according to the Wilby scale, where a score of 0 represented a dead, disintegrated polyp and ten represented a normal, healthy polyp (Wang et al. 2021). To prevent the precipitation of the chemicals during the experiment, up to 1% DMSO was included in the test solution to overcome the low water solubility of several of these compounds. Preliminary studies indicated that exposure to 1% DMSO or acetonitrile did not significantly affect the morphology of the polyps. For diazinon and linuron, a metabolic activation package (MAP) (2.4 µg/mL mice hepatic microsomal cytochrome P450, 225 µM NADPH, and 25 µM MgCl2) was included in the test solution. MAP was added to the test solutions because (1) H. vulgaris is metabolically inactive (Galliot 2012) and (2) these compounds require metabolic activation to exert toxic effects (Ellison et al. 2012; Uren Webster et al. 2015). For each experiment, triplicate analyses of each chemical exposure and vehicle and blank solution controls were included. Relative morphology scores were derived by expressing raw values as percent control, which was set to 1.
Lemna minor assay
A small community of plants was obtained from AquaHabit (Chatham, England) and was maintained in accordance with established methods and standard guidelines (Drost et al. 2007; OECD 2006) in Steinburg Medium under white, fluorescent lights with a 400 ft-c intensity set to a 16:8-h light to dark cycle at 25 °C (Rivenbark et al. 2022). Dosimetry experiments were conducted in sterile, transparent, 24-well plates with fitted lids (VWR, Radnor, PA). In each well, two plants (seven to eight fronds total) were exposed to increasing concentrations of each pesticide or the mixture prepared (Supplemental Fig. S1) in 2 mL Steinburg Medium for 7 days under the growth light. Each day, plants were observed for changes in frond number and surface area, which was measured using ImageJ (NIH, Bethesda, MD). At the end of the exposure period, all types of chlorophyll were extracted from the surviving plants by homogenizing (Homogenizer 150, Fisher Scientific) the plants in 1.5 mL of 80% acetonitrile. After a 48 h incubation in the dark at 4 °C, the total chlorophyll content was quantified by measuring the absorbance of the solution using UV/visible spectroscopy (Shimadzu UV-1800, Kyoto, Japan) set to 663 nm (Drost et al. 2007). Each pesticide was tested in at least four concentrations. For each experiment, triplicate analyses of each chemical exposure and vehicle and blank solution controls were included. Preliminary studies demonstrated that the inclusion of 1% acetonitrile in the exposure medium did not adversely impact the growth of plants. Relative surface area, frond number, and chlorophyll content values were derived by expressing raw values as percent control, which was set to 1.
Caenorhabditis elegans assay
Wildtype nematodes (Bristol N2) and Escherichia coli (E. coli) (strains OP50-1, NA22) were purchased from the Caenorhabditis Genetics Center (University of Minnesota) and maintained according to established protocols on 8P agar seeded with E. coli NA22 (Rivenbark et al. 2022). Large populations of larval stage 1 (L1) nematodes were obtained by a bleaching, washing, and incubation process. For dosimetry studies, groups of 2000 nematodes were transferred to a microcentrifuge tube containing 15 µL E. coli OP50-1, increasing concentrations of the test chemical (Supplemental Fig. S1), and sufficient K-media complete needed to achieve a final solution volume of 1 mL (Boyd et al. 2012). The tubes were incubated for up to 48 h on a rocking platform. After 24 and 48 h of exposure, nematodes were assessed for survival and body length. Survival was quantified by counting the number of alive nematodes in 10 µL of supernatant from each tube using a microscope (Olympus SZ61 zoom stereomicroscope, Olympus, Waltham, MA). Then, the nematodes in the tubes were washed three times with M9 solution, transferred to nematode growth media plates containing a lawn of E. coli OP50-1, and incubated for 48 h at 20 °C. After 48 h, the nose touch response was measured following the established procedures (Chatzigeorgiou and Schafer 2011) to assess the impact of pesticide exposure on neuronal health. Then the nematodes were paralyzed with 25 mM sodium azide, and body length was measured using the CellSens Entry (version 3) software that was connected to the microscope. Each pesticide was tested in at least four concentrations. All experiments were conducted in triplicate, and relevant vehicle and blank solution controls were included; the inclusion of 1% acetonitrile in the exposure medium did not adversely impact the endpoints of the nematodes. Relative body lengths, survival, and nose-touch responses were calculated by expressing raw values as percent control, which was set to 1.
Statistical analysis
An ANOVA with post-hoc Tukey HSD test (GraphPad Software) was used to determine statistical significance in the experimental results, with significance achieved at p ≤ 0.05. Each experiment was conducted in triplicate, and the resulting values were employed to compute averages and standard deviations and produce graphical representations. The dose–response modeling of the experimental results was conducted in R (4.3.0) using the four-parameter log-logistic functional form in the drm function within the drc library (Ritz et al. 2015). The 10% effective concentration (EC10), 50% effective concentration (EC50), and their 95% confidence intervals were estimated using the ED function within the drc library. The delta method was used for confidence interval calculations. A lack-of-fit test was also performed to compare the four-parameter log-logistic model to the one-way ANOVA model where p ≤ 0.05 was considered statistically significant. The fitted dose–response curves were visualized using the ggplot2 and ggprism libraries in R. In addition, the correlation and the grouping among the three model organism endpoints were explored with hierarchical clustering using Spearman correlation as the similarity/dissimilarity index and Ward’s minimum variance method with squared distances as the linkage method (Aghayev et al. 2023; Onel et al. 2019). Prior to the clustering analysis, the dataset was checked for missing values, and any missing values were imputed using bagged regression trees via the preprocess function of the caret library. Finally, the clustering results were visualized using the pheatmap library in R.
Results
Toxicity endpoints and dosimetry curves for 2,4,6-trichlorophenol
After exposure to 0–0.051 mM of 2,4,6-trichlorophenol for 92 h, H. vulgaris polyps had a dose- and time-dependent decline in morphological scores (Fig. 1A). Exposure to ≥ 0.032 mM 2,4,6-trichlorophenol caused complete disintegration all polyps after 42 h (score = 0), while lower doses (0.013 and 0.025 mM) had moderate toxicity to morphology and resulted in scores of 6–8. Based on the 92-h toxicity data, the EC10 and EC50 for 2,4,6-trichlorpohneol were calculated to be 0.0090 and 0.022 mM, respectively (Fig. 1B, Table 2). Results of lack of fit testing (p = 1.2E-9) indicated the dose–response curve displayed a better fit to the ANOVA model compared to the four-parameter log-logistic function (Table 2), which may be due to the lack of model fitting to the curve in regions x ≥ 0.032 mM. Dose–response curves for H. vulgaris morphological scores corresponding to pentachlorophenol, lindane, diazinon, glyphosate, linuron, trifluralin, aldicarb, paraquat, and the mixture are depicted in the supplemental material (Supplemental Fig. S2). All pesticides showed a similar dose- and time-dependent effect on H. vulgaris morphology over the exposure period, and 92-h EC10 and EC50 values were derived from these experiments (Table 2).

Toxicity of 2,4,6-trichlorophenol as shown by the morphological scores of H. vulgaris (A) and the corresponding dose–response curve at 92 h of exposure (B). Data represent the average value from triplicate analysis ± the standard deviation. * indicates a significant difference (p ≤ 0.05) from the vehicle control group after 92 h of exposure
For L. minor, significant toxicity was observed to surface area and frond number content after exposure to ≥ 0.0025 mM 2,4,6-trichlorophenol (Fig. 2 A and B) (p ≤ 0.05). Among the measured endpoints, chlorophyll content showed a slightly higher sensitivity to 2,4,6-trichlorophenol exposure as a 15 ± 3% decrease in chlorophyll content was observed after exposure to 0.0013 mM for 7 days (Fig. 2C) (p ≤ 0.05), but this concentration only inhibited the surface area and frond number by 5.8 ± 0.9% and − 3 ± 7%, respectively (p ≥ 0.05). Lack of fit testing also indicated the ANOVA model fit the dose–response curves the best (p ≤ 0.05, Table 3). Based on the dosimetry curve (Fig. 2D), the 168-h EC50 values for surface area, frond number, and chlorophyll content were calculated to be 0.0016, 0.0019, and 0.0011 mM, respectively (Table 3). Dose–response curves for pentachlorophenol, lindane, diazinon, glyphosate, linuron, trifluralin, aldicarb, paraquat, and the mixture with L. minor are depicted in the supplemental material (Supplemental Fig. S3). Toxicity studies with all of the pesticides demonstrated that the toxicity was dose- and time-dependent and yielded similar 168-h EC10 and EC50 values for surface area and frond number. However, chlorophyll content had a slightly lower EC10 value, compared to the frond number and surface area (p = 0.07 and 0.23, respectively).

Toxicity of 2,4,6-trichlorophenol on the surface area (A), frond number (B), and chlorophyll content (C) during a 7-day exposure. Dose–response curves of the 7 day toxicity data for all three measured endpoints (D). Data represent the average value from triplicate analysis ± the standard deviation. * indicates a significant difference (p ≤ 0.05) on day 7 of exposure compared to the vehicle control group
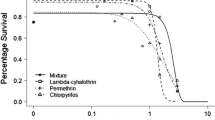
After 48 h of exposure to 0.10 mM 2,4,6-trichlorophenol, nematodes had a 5.9 ± 4% and 16 ± 3% impairment to body length and behavior, respectively (Fig. 3 A and B) (p ≤ 0.05). However, this concentration only caused a 2.6 ± 4% decrease in nematode survival (Fig. 3C) (p ≥ 0.05), and significant lethality was only observed in concentrations ≥ 0.25 mM. Similar to the other bioassays, toxicity was significantly higher after longer exposure durations. The 48 h EC50 values for body length, behavior, and survival were calculated to be 0.18, 0.25, and 0.23 mM, respectively for 2,4,6-trichlorophenol (Table 4). Dose–response curves for body length and survival displayed a good fit to the four-parameter log-logistic and ANOVA models (p ≥ 0.05) while the nose touch data displayed a slightly better fit to the ANOVA model (p = 0.019) (Table 4). Dose–response curves for pentachlorophenol, lindane, diazinon, glyphosate, linuron, trifluralin, aldicarb, paraquat, and the mixture with C. elegans are depicted in the supplemental material (Supplemental Fig. S4).

Toxicity of 2,4,6-trichlorophenol on the body length (A), nose touch response (B), and survival rate (C). The 48 h toxicity data was used to create the dose–response graphs for the three endpoints (D). Data represent the average value from triplicate analysis ± the standard deviation. * indicates a significant difference (p ≤ 0.05) after 48 h of exposure compared to the vehicle control group
Based on the calculated EC10 values, L. minor was the most sensitive bioassay to 2,4,6-trichlorophenol exposure, followed by H. vulgaris then C. elegans. When comparing the most sensitive endpoints for each bioassay, the chlorophyll content of L. minor had the lowest EC10 value compared to the morphology and body length of H. vulgaris and C. elegans, respectively (p ≤ 0.05). For EC50, H. vulgaris had the lowest values compared to the surface area and body length of L. minor and C. elegans, respectively (p ≤ 0.05).
EC10 and EC50 values for the remaining pesticides
For pentachlorophenol, the EC10 of the chlorophyll content of L. minor (0.00073 mM) was one to two-2 orders of magnitude lower than the values for frond number and surface area (0.0053–0.0086 mM) and the endpoints in H. vulgaris and C. elegans (0.0018–0.21 mM) (p ≤ 0.05). The morphology of H. vulgaris had the lowest EC50 value (0.0019 mM), which was similar to the values obtained by the chlorophyll content of L. minor (0.0023 mM) (p = 0.67) but was two orders of magnitude lower than the values for the survival of C. elegans (0.11 mM) (p ≤ 0.05). For lindane, the chlorophyll content of L. minor had the lowest EC10 and EC50 values (0.0039 and 0.0061 mM, respectively), which were one to two orders of magnitude lower than the values calculated for H. vulgaris and C. elegans. The morphology of H. vulgaris and the survival of C. elegans had similar EC10 and EC50 values p = 0.82 and 0.73, respectively).
The chlorophyll content of L. minor had the lowest EC10 value for diazinon (0.0016 mM), but it was not significantly lower than the values for the surface area or frond number of L. minor or the morphology of H. vulgaris (p ≥ 0.05). However, the morphology of H. vulgaris had the lowest EC50 value compared to all other measured endpoints (0.055 mM). For glyphosate, H. vulgaris was the most sensitive bioassay with an EC10 value (0.00036 mM) two to three orders of magnitude smaller than values derived from L. minor and C. elegans experiments. However, L. minor had lower EC50 values for glyphosate compared to the other bioassays (0.034–0.054 mM vs 0.15–0.22 mM).
For linuron, the chlorophyll content of L. minor had the lowest EC10 and EC50 values (0.00010 and 0.00015 mM, respectively), which was two to three orders of magnitude lower than the values derived from H. vulgaris and C. elegans dosimetry curves. C. elegans was the least sensitive to linuron and had EC50 values for its endpoints between 0.15 and 0.65 mM. Similarly, L. minor had the lowest EC10 and EC50 values that were several orders of magnitude smaller than the other bioassays after exposure to trifluralin. Specifically, the chlorophyll content of L. minor had an EC10 of 0.000042 mM, compared to 0.0057 mM and 0.19 mM for the morphology of H. vulgaris and nose touch response of C. elegans, respectively (p ≤ 0.05).
The morphology of H. vulgaris had the lowest EC10 and EC50 after aldicarb exposure (0.0043 and 0.0046 mM, respectively) compared to the other bioassays. Interestingly, the surface area of L. minor and nose touch of C. elegans are four orders of magnitude less sensitive to aldicarb exposure with EC50 values of 11 and 9.6 mM, respectively. For the mixture, the chlorophyll content of L. minor had the lowest EC10 value (0.00013 mM), compared to 0.0014 and 0.037 mM for the morphology of H. vulgaris and nose touch of C. elegans, respectively.
Correlations between bioassays
The distribution of EC10 and EC50 values of individual pesticides and the mixture for L. minor (Fig. 4 A and C) and C. elegans (Fig. 4 B and D) is depicted in Fig. 4. L. minor was more sensitive to the tested compounds and mixture, indicated by the lower EC10 and EC50 values. L. minor had a greater range of EC10 (10−4–100 mM) and EC50 (10−4–100 mM) values overall. Effective concentration values for each pesticide are distributed throughout the concentration range and do not overlap significantly. However, C. elegans displayed high variability in EC10 and EC50 values for each individual pesticide, and the range of EC10 and EC50 values was between 10−2–100 mM and 10−1–101 mM, respectively.

Distribution of EC10 (A–B) and EC50 (C–D) values (mM) for each pesticide between the measured endpoints for L. minor and C. elegans
When comparing EC10 values, L. minor was the most sensitive species to seven individual pesticides and the mixture, and H. vulgaris was the most sensitive species to the remaining two pesticides (aldicarb and glyphosate). For EC50, L. minor had the lowest values for five individual pesticides and the mixture, and H. vulgaris had the lowest for the remaining four compounds (2,4,6-trichlorophenol, pentachlorophenol, diazinon, and aldicarb). C. elegans consistently had the highest EC10 and EC50 values for all the tested compounds.
Importantly, the toxicity results indicated that some endpoints within a single bioassay were more sensitive to chemical exposure than the others; for example, the chlorophyll content of L. minor had the lowest EC10 and EC50 values for eight and five chemicals, respectively. For C. elegans, the survival rate had the lowest EC10 and EC50 values for four and five pesticides, respectively. The distribution of EC10 and EC50 values for all pesticides across the most sensitive endpoint for L. minor and C. elegans is shown in Fig. 5 A and B. These results demonstrated that L. minor has a greater range of EC10 and EC50 values for the tested pesticides and mixture compared to H. vulgaris and C. elegans. When comparing the most sensitive endpoints, L. minor generally had the lowest EC10 and EC50 values for the tested chemicals, followed by H. vulgaris and C. elegans.

Distribution of EC10 (A) and EC50 (B) (mM) values across all tested pesticides for the most sensitive endpoint in each bioassay
The relationship between the EC10 and EC50 values for each bioassay’s endpoints after chemical exposures was explored using clustering analysis coupled with dissimilarity matrices (Fig. 6 A and B) based on the Spearman correlation coefficient. For EC10 values, the endpoints were closely clustered into three distinct groups that correspond to each bioassay. The decreased distance between pairs of features (< 1), such as frond number-chlorophyll content (0.15) reflects the positive correlation between the features. Increased distance between features (> 1), such as body lengthmorphology (1.73), indicates the strong negative association between the endpoints. The endpoints measured in L. minor were the most tightly clustered, with distances of 0–0.15. While the three endpoints measured in C. elegans were also clustered together, they had a higher pair-wise distance (0.47–0.74). Values derived from H. vulgaris are negatively correlated with the endpoints from the other bioassays (distance > 1), except for the weak positive correlation with the survival of C. elegans (0.81). For EC50 values, the endpoints of each bioassay were clustered into two main clades, where H. vulgaris and C. elegans were clustered in the same clade and L. minor was clustered alone (Fig. 6B). Additionally, the overall similarity of endpoints in the sample bioassay decreased. For example, the distances between the endpoints of L. minor and C. elegans increased to 0.01–0.19 and 0.59–0.90, respectively. Interestingly, the distance between the morphology of H. vulgaris and the other measured endpoints decreased slightly and was negatively correlated with the surface area of L. minor.

Clustering heatmap with distance matrix and dendrograms depicting the relationships between the measured endpoints for all model organisms based on their EC10 (A) and EC50 (B) values for all chemical exposures based on the Spearman correlation coefficient
Correlation between pesticides
Figure 7 A and B depict the relationship between the EC10 and EC50 values for all the pesticides according to the Spearman correlation method. For EC10 values, two main clades were created from the associated dendrogram (Fig. 6A). In the first clade, glyphosate, paraquat, pentachlorophenol, trifluralin, and the mixture were clustered together. In clade 2, diazinon and aldicarb had the highest dissimilarity from the other pesticides (distance 0.5–1.04 and 0.43–0.86, respectively). When categorized by chemical class, there was no direct relationship with the overall cluster structure. A slight relationship between the target organism and cluster structure was observed as three herbicides and one insecticide were clustered in clade 1, and three insecticides and two herbicides were clustered in clade 2. The mixture had high similarity between most of the compounds (distance = 0.04–0.32), except for diazinon and aldicarb, where the distance was 0.82 and 0.64, respectively

Clustering heatmap with distance matrix and dendrograms depicting the comparison of EC10 (A) and EC50 (B) values of all the tested pesticides and the mixture according to the Spearman correlation coefficient
For EC50 values, the clustering structure also yielded two primary clades, where diazinon and aldicarb were strongly clustered away from the other pesticides (Fig. 6B). Diazinon had the highest dissimilarity from the other pesticides (distances = 0.96–1.68). Similar to the EC10 results, no clear relationship between the clustering and chemical class was observed. However, several of the chemicals with the same target organism were tightly clustered together and had low distances. For example, insecticides diazinon and aldicarb were consistently clustered closely and had a distance of 0.32. Pairs of herbicides, such as 2,4,6-trichlorohenol-glyphosate and trifluralin-paraquat were closely clustered together and had pair-wise distances of 0.04. The mixture also displayed high similarity to most of the compounds, except for diazinon and aldicarb (distance 1.68 and 1.07, respectively).
Discussion
Toxicity of individual pesticides
The main mode of action for 2,4,6-trichlorophenol toxicity is through acetylcholinesterase inhibition (Matsumura et al. 1997), which causes a variety of effects in plants and animals. In plants, acetylcholinesterase plays a vital role in water retention and photosynthesis (Wessler et al. 2001); in animals, it is essential for neuronal communication (Devi et al. 2023; Matsumura et al. 1997). This mode of action explains the increased sensitivity of L. minor to 2,4,6-trichlorophenol exposure for all measured endpoints (Fig. 2A-D) as well as the rapid deterioration of the nose touch response observed in C. elegans at high concentrations (Fig. 3B). The EC10 and EC50 values derived for H. vulgaris and L. minor in the current study are in accordance with values for other aquatic organisms, like Scenedesmus obliquus (a green microalgae) and Daphnia magna (a crustacean) (Xing et al. 2012). Pentachlorophenol inhibits ATP-ases and can produce reactive oxygen species (Maheshwari et al. 2023). Plants have previously been shown to be more sensitive to pentachlorophenol than other organisms due to the inhibition of photosynthesis caused by exposure (Huber et al. 1982; Repetto et al. 2001). The EC50 values for the endpoints in L. minor and C. elegans calculated in the current study are in accordance with published values for these bioassays (Huber et al. 1982; Kammenga et al. 1994; Repetto et al. 2001) and other hydra species (Silva et al. 2001). Because the number of chlorinated substitutions contributes to the toxicity of the compound, EC10 and EC50 estimates for all the bioassays were lower for pentachlorophenol than 2,4,6-trichlorophenol (Freitag et al. 1994).
Lindane is a legacy organochlorine insecticide that was banned under the Stockholm Convention due to concerns over its toxicity and environmental persistence (Nolan et al. 2012).
While there is no main mode of action for insects, the neurotoxicity of lindane is among the most concerning effects after exposures (Nolan et al. 2012), which explains the lower values of EC10 and EC50 for the nose touch response of C. elegans compared to the body length (Yu et al. 2022). L. minor was two orders of magnitude more sensitive to lindane compared to the other models, likely because of the impact of lindane on plant development and photosynthesis (Pereira et al. 2010). Diazinon is an organophosphate insecticide that has been previously shown to cause toxicity to aquatic species after the contamination of waters (Bailey et al. 2000). The main mode of action is the inhibition of acetylcholinesterase (Velki et al. 2017). Previous studies have also demonstrated that significant toxicity to aquatic species was observed at levels at and below the recommended application doses (Natal-da-Luz et al. 2012), highlighting the importance of investigating the toxicity of low doses to more species. Interestingly, L. minor, H. vulgaris, and C. elegans were similarly susceptible to diazinon toxicity when comparing EC10 estimates.
Glyphosate is one of the most commonly used broad-spectrum organophosphate herbicides worldwide that inhibits the biosynthesis of essential amino acids. Specifically, it targets 5-enolpyruvylshikimate-3-phosphate synthase and causes a decrease in plant growth and survival (Gill et al. 2018; Singh et al. 2020). H. vulgaris and L. minor were sensitive to glyphosate exposure, likely because aromatic amino acids also play a key role in the health of these species. While C. elegans can obtain amino acids from their bacterial food source (Zečić et al. 2019), L. minor and H. vulgaris mainly rely on the biosynthesis of these molecules to survive, increasing their sensitivity to glyphosate exposure (Trovato et al. 2021). The increased sensitivity of aquatic species and non-target plant species to this compound highlights the importance of using diverse species to understand the environmental impact of chemicals classified as herbicides. Linuron is a phenylurea herbicide that directly decreases the photosynthetic ability of plants by inhibiting the function of the electron transport chain (Maharaj et al. 2020; Santos et al. 2014), which explains why L. minor was several orders of magnitude more sensitive to exposure compared to the other bioassays. The EC50 values for L. minor (Gatidou et al. 2015) and C. elegans (Zöngür and Sari 2023) are in accordance with previously published values. Importantly, the EC10 values derived for L. minor are an order of magnitude lower than levels applied to fields (Durand and Barcelo 1992), demonstrating the possibility of non-target plant toxicity after applications.
Trifluralin is a dinitroaniline herbicide that inhibits microtubule formation during mitosis in plants (Coleman et al. 2020), but is known to impact aquatic species through the same mode of action (Fernandes et al. 2013). L. minor was the most sensitive species to exposure, followed by H. vulgaris and C. elegans. The EC50 value for H. vulgaris was similar to those derived for an aquatic larval frog species, Lithobates clamitans (Weir et al. 2012). Perhaps, the inclusion of endpoints that are related to mitotic processes in nematodes, like the quantification of offspring (Sant'anna et al. 2016), would have demonstrated a higher sensitivity than the measured endpoints in C. elegans. However, the EC10 values for all the bioassays were higher than levels reported in soils in cotton fields (3.2–6.0 ppb or 8.9–18 nM) (Li et al. 2021).
Aldicarb is a broad-acting carbamate insecticide that inhibits acetylcholinesterase (Blacker et al. 2010), like 2,4,6-trichlorophenol. However, it has been suggested that these effects are not observed at human or environmentally relevant exposure levels and are rapidly reversible (Blacker et al. 2010). The low toxicity observed in L minor and C. elegans in general may be due to the high water solubility and low LogP of aldicarb that decreases the baseline toxicity (Mayer and Reichenberg 2006). Additionally, aldicarb is rapidly metabolized in the environment and in animals to less toxic intermediates (Blacker et al. 2010); since H. vulgaris is metabolically inactive, this detoxification process could be inhibited, leading to the increased toxicity that was observed.
Paraquat is a viologen and bipyridinium herbicide that inhibits photosynthesis and promotes the formation of reactive oxygen species in plants and animals (Blanco-Ayala et al. 2014). This mode of action explains the increased sensitivity of L. minor to exposure compared to the other bioassays. Previous studies have also demonstrated that among other tested pesticides, paraquat was among the most toxic compounds to L. minor (Tagun and Boxall 2018). H. vulgaris was more sensitive to exposure than the average C. elegans response, likely because quaternary ammonium compounds are known to have high toxicity to aquatic species compared to soil organisms (Blanco-Ayala et al. 2014; Zhang et al. 2015). Importantly, levels of paraquat detected in surface waters (Thi Hue et al. 2018) were above estimated EC10 and EC50 values derived for H. vulgaris and L. minor.
Chemical mixtures have previously been shown to have toxicities different from the constituent parts due to additive, synergistic, or antagonistic activity between the individual chemicals (Bart et al. 2022; Hernández et al. 2013; Hernando et al. 2003; Rider and Simmons 2018; Wang et al. 2021). For H. vulgaris and L. minor, the pesticide mixture had one of the lowest EC10 and EC50 values across all the tested chemicals. However, it was not the most toxic substance evaluated according to EC50 values, either because of dilution effects or antagonism between the individual pesticides (Hernando et al. 2003; Wang et al. 2021; Zhang et al. 2015). However, the toxicity testing of chemical mixtures is important because the mixture toxicity is difficult to predict (Deneer 2000).
Correlation between bioassays
Using a set of bioassays that covers diverse ecological niches, levels, and endpoints is advantageous for understanding the toxicity of individual compounds and mixtures (Hernando et al. 2003; Repetto 2013). The current set of bioassays combined organisms that have standardized toxicity testing guidelines (EPA 2009; Pollino and Holdway 1999; Queirós et al. 2019) and have been individually employed previously for the assessment of other pesticides, wastewaters, sediments, polycyclic aromatic hydrocarbons, and complex chemical mixtures (Gatidou et al. 2015; Graves et al. 2005; Höss et al. 2009; Taraldsen and Norberg-King 1990).
Fairchild et. al utilized Lemna minor and Selenastrum capricornutum (a green algae) to characterize the toxicity of several herbicides and found that both species were equally sensitive to exposure (Fairchild et al. 1997). The inclusion of the addition of Myriophyllum aquaticum (a rooted macrophyte) in the bioassay battery was suggested to expand the toxicity assessment to more sensitive aquatic plant species. When exposed to atrazine in aqueous medium, L. minor was shown to be more sensitive to the aquatic species D. magnia (Klementová et al. 2019) but had comparable results to M. aquaticum (Teodorović et al. 2012). M. aquaticum was more sensitive to atrazine than L. minor when both species were exposed to contaminated sediments, likely because of the roots present in M. aquaticum that promote direct contact with the sediment. Because the herbicides in the present study were exposed to the bioassays by spiked aqueous mediums, L. minor was likely more sensitive than M. aquaticum may have been (Park et al. 2021).
H. vulgaris has been shown to be a more sensitive species to the pesticide 4-chlorophenol than Hydra viridissima but was less sensitive than the fish species Pimephales promelas and Leepomis macrochirus (Pollino and Holdway 1999). Consistent with previous studies (Lui and Wrischer 2002), H. vulgaris was the most sensitive species to insecticides, and rapid deterioration of the tentacles and morphological scores were observed after exposure, which is consistent with the sensitivity of this species to aldicarb, diazinon, and pentachlorophenol observed in the current study. The inclusion of H. vulgaris in toxicity testing is important as these species play an important role in their ecosystem and may be used as an early indicator of pollution in the environment (Beach and Pascoe 1998; Pollino and Holdway 1999).
Pesticides are among the most frequently used chemicals for toxicity testing with C. elegans (Queirós et al. 2019) and have been shown to produce LD50 values that are correlated with oral LD50 in rats, mice, and rabbits (Boyd et al. 2012; Hunt 2017). C. elegans also displayed a high correlation with zebrafish in 59–79% of tested chemicals in previous studies (Boyd et al. 2012). While less commonly reported to be used in a biological battery, C. elegans play an important role in the current study to describe ecological and mammalian toxicity caused by pesticides (Cole et al. 2004; Hunt 2017).
Interestingly, each bioassay had an endpoint that was more sensitive to pesticide exposure. For L. minor, chlorophyll content generally was more sensitive to pesticide exposure compared to surface area and frond number. This may be due to the role of chlorophyll degradation in the plant senescence cycle where chlorophylls are broken down to release nutrients and promote plant viability (Lisiewska et al. 2006). Therefore, the loss of chlorophyll may be an earlier and more sensitive sign of plant toxicity compared to the overall surface area. This finding is in accordance with previous studies that demonstrate chlorophyll content is a more sensitive endpoint than frond number for pesticide exposure (Fekete-Kertész et al. 2015). The close clustering of the L. minor endpoints was likely because these endpoints are more related to each other than the group of endpoints measured in C. elegans. For example, plants with high surface area are more likely to have a higher abundance of fronds and chlorophylls as these endpoints contribute to the overall growth of the plants.
For C. elegans, the survival rate had the lowest EC50 values for most of the pesticides compared to body length and nose touch response. The variation in the dose–response curves for C. elegans is larger than those observed in L. minor and H. vulgaris, possibly due to the endpoints of C. elegans corresponding to different mechanisms of toxicity, i.e., survival, growth, and neurological system (Anderson et al. 2001). Nematodes that show sensitivity to nose touch assay may not have growth stunting as neurological endpoints are not shown to be related to growth outcomes directly (Anderson et al. 2001). Therefore, varying endpoints reflecting different mechanisms of action are needed as broad-range indicators for chemical mixtures and environmental samples. Interestingly, lethality endpoints have been shown to be correlative with behavioral and locomotive endpoints (Anderson et al. 2001). This also reflects the wide array of endpoints available in the C. elegans model and implemented in the current study to detect toxicity to a variety of different endpoints and biological systems. The varying degrees of clustering and correlation between the endpoints of all organisms demonstrate the need to include different types of model organisms in the toxicity analysis of chemicals and design mixtures to better describe the hazards.
C. elegans consistently ranked the least sensitive to all chemicals among the three bioassays. The addition of nematicides in future studies may reveal an increased sensitivity of the nematode model compared to the plant and aquatic models in a set of pesticides. Additionally, it has been previously shown that nematode species may be more sensitive to chemical exposures conducted in solid-phase mediums (soils and sediments) compared to the aqueous phase (Kim et al. 2020). However, conducting the testing in an aqueous medium is advantageous as the chemical is dissolved and freely available, whereas it may be sequestered by organic matter in soil matrices (Sarkar et al. 2021).
Conclusions
Because pesticides can ubiquitously occur in soil and aquatic environments and pose a risk to target and non-target organisms in the environment, using a set of bioassays that incorporates environmentally relevant species to characterize toxicity is important. In the current study, we used a novel set of living organisms that are representative of important environmental niches containing plant, aquatic, and soil species to compare the toxicities of nine individual pesticides and a mixture. Each bioassay displayed varying sensitivity to the individual pesticides and the mixture L. minor had the lowest EC10 and EC50 values for most of the individual pesticides tested, but these values may not be representative of non-target or more resistant species’ responses to these compounds. More importantly, the most sensitive endpoint of each bioassay displayed distinct but slightly overlapping EC10 and EC50 ranges for all of the chemicals tested. These representative bioassays can be used in combination to characterize the toxicity of individual pesticide exposure to living organisms and predict the toxicity of real-life environmental samples and the prioritization of contaminated sites for further chemical analysis and remediation.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Aghayev Z, Szafran AT, Tran A, Ganesh HS, Stossi F, Zhou L, Mancini MA, Pistikopoulos EN, Beykal B (2023) Machine learning methods for endocrine disrupting potential identification based on single-cell data. Chem Eng Sci 281:119086
Aliferis KA, Materzok S, Paziotou GN, Chrysayi-Tokousbalides M (2009) Lemna minor L. as a model organism for ecotoxicological studies performing 1H NMR fingerprinting. Chemosphere 76(7):967–973
Anderson GL, Boyd WA, Williams PL (2001) Assessment of sublethal endpoints for toxicity testing with the nematode Caenorhabditis elegans. Environ Toxicol Chem: Int J 20(4):833–838
ATSDR (2022) Substance priority list
Bailey HC, Deanovic L, Reyes E, Kimball T, Larson K, Cortright K, Connor V, Hinton DE (2000) Diazinon and chlorpyrifos in urban waterways in northern California, USA. Environ Toxicol Chem: Int J 19(1):82–87
Bart S, Short S, Jager T, Eagles EJ, Robinson A, Badder C, Lahive E, Spurgeon DJ, Ashauer R (2022) How to analyse and account for interactions in mixture toxicity with toxicokinetic-toxicodynamic models. Sci Total Environ 843:157048
Beach MJ, Pascoe D (1998) The role of Hydra vulgaris (Pallas) in assessing the toxicity of freshwater pollutants. Water Res 32(1):101–106
Bhatti MA, Felsot AS, Al-Khatib K, Kadir S, Parker R (1995) Effects of simulated chlorsulfuron drift on fruit yield and quality of sweet cherries (Prunus avium L.). Environ Toxicol Chem: Int J 14(3):537–544
Blacker AM, Kelly ID, Lantz JL, Mihlan GJ, Jones RL, Young BM (2010) Aldicarb: toxicity, exposure and risks to humans. Hayes' Handb Pestic Toxic 1619–1632
Blanco-Ayala T, Andérica-Romero A, Pedraza-Chaverri J (2014) New insights into antioxidant strategies against paraquat toxicity. Free Radical Res 48(6):623–640
Boutin C, Lee HB, Peart ET, Batchelor PS, Maguire RJ (2000) Effects of the sulfonylurea herbicide metsulfuron methyl on growth and reproduction of five wetland and terrestrial plant species. Environ Toxicol Chem: Int J 19(10):2532–2541
Boyd WA, Smith MV, Freedman JH (2012) Caenorhabditis elegans as a model in developmental toxicology. Methods Mol Biol 15–24
Cedergreen N, Streibig JC (2005) The toxicity of herbicides to non-target aquatic plants and algae: assessment of predictive factors and hazard. Pest Manag Sci: Formerly Pestic Sci 61(12):1152–1160
Chatzigeorgiou M, Schafer WR (2011) Lateral facilitation between primary mechanosensory neurons controls nose touch perception in C. elegans. Neuron 70(2):299–309
Cole RD, Anderson GL, Williams PL (2004) The nematode Caenorhabditis elegans as a model of organophosphate-induced mammalian neurotoxicity. Toxicol Appl Pharmacol 194(3):248–256
Coleman NV, Rich DJ, Tang FH, Vervoort RW, Maggi F (2020) Biodegradation and abiotic degradation of trifluralin: a commonly used herbicide with a poorly understood environmental fate. Environ Sci Technol 54(17):10399–10410
Cunha JP, Chueca P, Garcerá C, Moltó E (2012) Risk assessment of pesticide spray drift from citrus applications with air-blast sprayers in Spain. Crop Prot 42:116–123
de Alkimin GD, Santos J, Soares AM, Nunes B (2020) Ecotoxicological effects of the azole antifungal agent clotrimazole on the macrophyte species Lemna minor and Lemna gibba. Comp Biochem Physiol c: Toxicol Pharmacol 237:108835
Deneer JW (2000) Toxicity of mixtures of pesticides in aquatic systems. Pest Manag Sci: Formerly Pestic Sci 56(6):516–520
Devi RK, Ganesan M, Chen T-W, Chen S-M, Rasheed RA, Al-Onazi WA, Elshikh MS, Liu X, Yu J (2023) Hexagonal nanosheets of pyrrochlore-type lanthanum stannate for sensitive detection of chlorinated pesticide in food and environmental samples. Food Chem 404:134516
Drost W, Matzke M, Backhaus T (2007) Heavy metal toxicity to Lemna minor: studies on the time dependence of growth inhibition and the recovery after exposure. Chemosphere 67(1):36–43
Durand G, Barcelo D (1992) Environmental degradation of atrazine, linuron and fenitrothion in soil samples. Toxicol Environ Chem 36(3–4):225–234
Ellison CA, Tian Y, Knaak JB, Kostyniak PJ, Olson JR (2012) Human hepatic cytochrome P450-specific metabolism of the organophosphorus pesticides methyl parathion and diazinon. Drug Metab Dispos 40(1):1–5
EPA (2009) OECD Guidelines for the testing of chemicals 221: Lemna specific growth inhibition test
Fairchild J, Ruessler D, Haverland P, Carlson A (1997) Comparative sensitivity of Selenastrum capricornutum and Lemna minor to sixteen herbicides. Arch Environ Contam Toxicol 32:353–357
Fekete-Kertész I, Kunglné-Nagy Z, Gruiz K, Magyar Á, Farkas É, Molnár M (2015) Assessing toxicity of organic aquatic micropollutants based on the total chlorophyll content of Lemna minor as a sensitive endpoint. Periodica Polytech, Chem Eng 59(4):262–271
Fernandes TC, Pizano MA, Marin-Morales MA (2013) Characterization, modes of action and effects of trifluralin: a review, IntechOpen
Fochtman P, Raszka A, Nierzedska E (2000) The use of conventional bioassays, microbiotests, and some “rapid” methods in the selection of an optimal test battery for the assessment of pesticides toxicity. Environ Toxicol: Int J 15(5):376–384
Freitag D, Ballhorn L, Behechti A, Fischer K, Thumm W (1994) Structural configuration and toxicity of chlorinated alkanes. Chemosphere 28(2):253–259
Fujita T, Nishimura K, Takayama C, Yoshida M, Uchida M (2010) Hydrophobicity as a key physicochemical parameter of environmental toxicology of pesticides. Hayes' Handb Pestic Toxicol 1229–1252
Galliot B (2012) Hydra, a fruitful model system for 270 years. Int J Dev Biol 56(6-7–8):411–423
Gatidou G, Stasinakis AS, Iatrou EI (2015) Assessing single and joint toxicity of three phenylurea herbicides using Lemna minor and Vibrio fischeri bioassays. Chemosphere 119:S69–S74
Gill JPK, Sethi N, Mohan A, Datta S, Girdhar M (2018) Glyphosate toxicity for animals. Environ Chem Lett 16:401–426
Graves AL, Boyd WA, Williams PL (2005) Using transgenic Caenorhabditis elegans in soil toxicity testing. Arch Environ Contam Toxicol 48(4):490–494
Grégoire G, Fortin J, Ebtehaj I, Bonakdari H (2022) Novel hybrid statistical learning framework coupled with random forest and grasshopper optimization algorithm to forecast pesticide use on golf courses. Agriculture 12(7):933
Hernández AF, Parrón T, Tsatsakis AM, Requena M, Alarcón R, López-Guarnido O (2013) Toxic effects of pesticide mixtures at a molecular level: their relevance to human health. Toxicology 307:136–145
Hernando M, Ejerhoon M, Fernandez-Alba A, Chisti Y (2003) Combined toxicity effects of MTBE and pesticides measured with Vibrio fischeri and Daphnia magna bioassays. Water Res 37(17):4091–4098
Höss S, Jänsch S, Moser T, Junker T, Römbke J (2009) Assessing the toxicity of contaminated soils using the nematode Caenorhabditis elegans as test organism. Ecotoxicol Environ Saf 72(7):1811–1818
Huber W, Schubert V, Sautter C (1982) Effects of pentachlorophenol on the metabolism of the aquatic macrophyte Lem na minor L. Environ Pollut Series a, Ecol Biol 29(3):215–223
Hunt PR (2017) The C. elegans model in toxicity testing. J Appl Toxicol 37(1):50–59
Johansen BE (2003) The dirty dozen: toxic chemicals and the earth’s future, Bloomsbury Publishing USA
Kammenga J, Van Gestel C, Bakker J (1994) Patterns of sensitivity to cadmium and pentachlorophenol among nematode species from different taxonomic and ecological groups. Arch Environ Contam Toxicol 27:88–94
Karntanut W, Pascoe D (2000) A comparison of methods for measuring acute toxicity to Hydra vulgaris. Chemosphere 41(10):1543–1548
Kim SW, Kim D, Jeong S-W, An Y-J (2020) Size-dependent effects of polystyrene plastic particles on the nematode Caenorhabditis elegans as related to soil physicochemical properties. Environ Pollut 258:113740
Klementová Š, Hornychová L, Šorf M, Zemanová J, Kahoun D (2019) Toxicity of atrazine and the products of its homogeneous photocatalytic degradation on the aquatic organisms Lemna minor and Daphnia magna. Environ Sci Pollut Res 26:27259–27267
Leu C, Singer H, Stamm C, Müller SR, Schwarzenbach RP (2004) Variability of herbicide losses from 13 fields to surface water within a small catchment after a controlled herbicide application. Environ Sci Technol 38(14):3835–3841
Li Y, Li C, Li B, Ma Z (2021) Trifluralin residues in soils from main cotton fields of China and associated ecological risk. Chemosphere 284:131300
Lisiewska Z, Kmiecik W, Korus A (2006) Content of vitamin C, carotenoids, chlorophylls and polyphenols in green parts of dill (Anethum graveolens L.) depending on plant height. J Food Compos Anal 19(23):134–140
Lui A, Wrischer M (2002) Effect of insecticides (Dimiline WP 25, Torak EC 24 and Gamacide 20) on hydra (Hydra vulgaris Pallas). Int J Dev Biol 35(3):335–340
Maharaj S, El Ahmadie N, Rheingold S, El Chehouri J, Yang L, Souders CL II, Martyniuk CJ (2020) Sub-lethal toxicity assessment of the phenylurea herbicide linuron in developing zebrafish (Danio rerio) embryo/larvae. Neurotoxicol Teratol 81:106917
Maheshwari N, Khan AA, Mahmood R, Salam S (2023) Pentachlorophenol-induced hemotoxicity diminishes antioxidant potential and oxidizes proteins, thiols, and lipids in rat blood: an in vivo study. Heliyon 9(5)
Mariani L, De Pascale D, Faraponova O, Tornambè A, Sarni A, Giuliani S, Ruggiero G, Onorati F, Magaletti E (2006) The use of a test battery in marine ecotoxicology: the acute toxicity of sodium dodecyl sulfate. Environ Toxicol: Int J 21(4):373–379
Matsumura H, Matsuoka M, Igisu H, Ikeda M (1997) Cooperative inhibition of acetylcholinesterase activities by hexachlorophene in human erythrocytes. Arch Toxicol 71:151–156
Mayer P, Reichenberg F (2006) Can highly hydrophobic organic substances cause aquatic baseline toxicity and can they contribute to mixture toxicity? Environ Toxicol Chem: Int J 25(10):2639–2644
Metcalfe TL, Dillon PJ, Metcalfe CD (2008) Detecting the transport of toxic pesticides from golf courses into watersheds in the Precambrian Shield region of Ontario, Canada. Environ Toxicol Chem: Int J 27(4):811–818
Natal-da-Luz T, Moreira-Santos M, Ruepert C, Castillo LE, Ribeiro R, Sousa JP (2012) Ecotoxicological characterization of a tropical soil after diazinon spraying. Ecotoxicology 21:2163–2176
Nolan K, Kamrath J, Levitt J (2012) Lindane toxicity: a comprehensive review of the medical literature. Pediatr Dermatol 29(2):141–146
Nowell LH, Norman JE, Moran PW, Martin JD, Stone WW (2014) Pesticide toxicity index—a tool for assessing potential toxicity of pesticide mixtures to freshwater aquatic organisms. Sci Total Environ 476:144–157
OECD (2006) Test No. 221: Lemna sp. Growth Inhibition Test
Onel M, Beykal B, Ferguson K, Chiu WA, McDonald TJ, Zhou L, House JS, Wright FA, Sheen DA, Rusyn I (2019) Grouping of complex substances using analytical chemistry data: a framework for quantitative evaluation and visualization. PLoS One 14(10):e0223517
Park J, Yoo E-J, Shin K, Depuydt S, Li W, Appenroth K-J, Lillicrap AD, Xie L, Lee H, Kim G (2021) Interlaboratory validation of toxicity testing using the duckweed Lemna minor root-regrowth test. Biology 11(1):37
Pereira RC, Monterroso C, Macías F (2010) Phytotoxicity of hexachlorocyclohexane: effect on germination and early growth of different plant species. Chemosphere 79(3):326–333
Pollino CA, Holdway DA (1999) Potential of two hydra species as standard toxicity test animals. Ecotoxicol Environ Saf 43(3):309–316
Queirós L, Pereira J, Gonçalves F, Pacheco M, Aschner M, Pereira P (2019) Caenorhabditis elegans as a tool for environmental risk assessment: emerging and promising applications for a “nobelized worm.” Crit Rev Toxicol 49(5):411–429
Reinecke S, Reinecke A (2007) The impact of organophosphate pesticides in orchards on earthworms in the Western Cape, South Africa. Ecotoxicol Environ Saf 66(2):244–251
Repetto G, Jos A, Hazen M, Molero M, Del Peso A, Salguero M, Del Castillo P, Rodrıguez-Vicente M, Repetto M (2001) A test battery for the ecotoxicological evaluation of pentachlorophenol. Toxicol in Vitro 15(4–5):503–509
Repetto G (2013) Encyclopedia of aquatic ecotoxicology. Férard J, Blaise C (ed), Springer, Dordrecht
Rider CV, Simmons JE (2018) Chemical mixtures and combined chemical and nonchemical stressors: exposure, toxicity, analysis, and risk. Springer
Ritz C, Baty F, Streibig JC, Gerhard D (2015) Dose-response analysis using R. Plos One 10(12):e0146021
Rivenbark KJ, Wang M, Lilly K, Tamamis P, Phillips TD (2022) Development and characterization of chlorophyll-amended montmorillonite clays for the adsorption and detoxification of benzene. Water Res 221:118788
Sant’anna V, de Souza W, Vommaro RC (2016) Anthelmintic effect of herbicidal dinitroanilines on the nematode model Caenorhabditis elegans. Exp Parasitol 167:43–49
Santos SM, Videira RA, Fernandes MA, Santos MS, Moreno AJ, Vicente JA, Jurado AS (2014) Toxicity of the herbicide linuron as assessed by bacterial and mitochondrial model systems. Toxicol in Vitro 28(5):932–939
Sarkar B, Mukhopadhyay R, Ramanayaka S, Bolan N, Ok YS (2021) The role of soils in the disposition, sequestration and decontamination of environmental contaminants. Philos Trans R Soc B 376(1834):20200177
Schwingl PJ, Lunn RM, Mehta SS (2021) A tiered approach to prioritizing registered pesticides for potential cancer hazard evaluations: implications for decision making. Environ Health 20:1–14
Sharma A, Kumar V, Shahzad B, Tanveer M, Sidhu GPS, Handa N, Kohli SK, Yadav P, Bali AS, Parihar RD (2019) Worldwide pesticide usage and its impacts on ecosystem. SN Appl Sci 1:1–16
Silva J, Iannacone J, Cifuentes A, Troncoso L, Bay-Schmith E, Larrain A (2001) Assessment of sensitivity to pentachlorophenol (PCP) in 18 aquatic species, using acute and chronic ecotoxicity bioassays. Ecotoxicol Environ Restor 4(1):10–17
Silva AM, Martins-Gomes C, Silva TL, Coutinho TE, Souto EB, Andreani T (2022) In vitro assessment of pesticides toxicity and data correlation with pesticides physicochemical properties for prediction of toxicity in gastrointestinal and skin contact exposure. Toxics 10(7):378
Simonsen L, Fomsgaard IS, Svensmark B, Spliid NH (2008) Fate and availability of glyphosate and AMPA in agricultural soil. J Environ Sci Health B 43(5):365–375
Singh S, Kumar V, Gill JPK, Datta S, Singh S, Dhaka V, Kapoor D, Wani AB, Dhanjal DS, Kumar M (2020) Herbicide glyphosate: toxicity and microbial degradation. Int J Environ Res Public Health 17(20):7519
Sochová I, Hofman J, Holoubek I (2006) Using nematodes in soil ecotoxicology. Environ Int 32(3):374–383
Tagun R, Boxall AB (2018) The response of Lemna minor to mixtures of pesticides that are commonly used in Thailand. Bull Environ Contam Toxicol 100(4):516–523
Taraldsen JE, Norberg-King TJ (1990) New method for determining effluent toxicity using duckweed (Lemna minor). Environ Toxicol Chem: Int J 9(6):761–767
Teodorović I, Knežević V, Tunić T, Čučak M, Lečić JN, Leovac A, Tumbas II (2012) Myriophyllum aquaticum versus Lemna minor: sensitivity and recovery potential after exposure to atrazine. Environ Toxicol Chem 31(2):417–426
Thi Hue N, Nguyen TPM, Nam H, Hoang Tung N (2018) Paraquat in surface water of some streams in Mai Chau Province, the Northern Vietnam: concentrations, profiles, and human risk assessments. J Chem 2018:1–8
Trovato M, Funck D, Forlani G, Okumoto S, Amir R (2021) Amino acids in plants: regulation and functions in development and stress defense, p. 772810. Front Media SA
Tudi M, Daniel Ruan H, Wang L, Lyu J, Sadler R, Connell D, Chu C, Phung DT (2021) Agriculture development, pesticide application and its impact on the environment. Int J Environ Res Public Health 18(3):1112
Ueda K, Nagai T (2021) Relative sensitivity of duckweed Lemna minor and six algae to seven herbicides. J Pestic Sci 46(3):267–273
Ullah S, Zorriehzahra MJ (2015) Ecotoxicology: a review of pesticides induced toxicity in fish. Adv Anim Vet Sci 3(1):40–57
Uren Webster TM, Perry MH, Santos EM (2015) The herbicide linuron inhibits cholesterol biosynthesis and induces cellular stress responses in brown trout. Environ Sci Technol 49(5):3110–3118
Velki M, Di Paolo C, Nelles J, Seiler T-B, Hollert H (2017) Diuron and diazinon alter the behavior of zebrafish embryos and larvae in the absence of acute toxicity. Chemosphere 180:65–76
Wang W (1990) Literature review on duckweed toxicity testing. Environ Res 52(1):7–22
Wang M, Chen Z, Rusyn I, Phillips TD (2021) Testing the efficacy of broad-acting sorbents for environmental mixtures using isothermal analysis, mammalian cells, and H vulgaris. J Hazard Mater 408:124425
Warne MSJ, Reichelt-Brushett A (2023) Marine pollution–monitoring, management and mitigation, pp. 155–184, Springer
Weir SM, Yu S, Salice CJ (2012) Acute toxicity of herbicide formulations and chronic toxicity of technical-grade trifluralin to larval green frogs (Lithobates clamitans). Environ Toxicol Chem 31(9):2029–2034
Wessler I, Kilbinger H, Bittinger F, Kirkpatrick CJ (2001) The non-neuronal cholinergic system the biological role of non-neuronal acetylcholine in plants and humans. Japan J Pharmacol 85(1):2–10
WHO (2022) Peticie residues in food
Xing L, Liu H, Giesy JP, Yu H (2012) pH-dependent aquatic criteria for 2, 4-dichlorophenol, 2, 4, 6-trichlorophenol and pentachlorophenol. Sci Total Environ 441:125–131
Yu Y, Hua X, Chen H, Wang Z, Han Y, Chen X, Yang Y, Xiang M (2022) Glutamatergic transmission associated with locomotion-related neurotoxicity to lindane over generations in Caenorhabditis elegans. Chemosphere 290:133360
Zečić A, Dhondt I, Braeckman BP (2019) The nutritional requirements of Caenorhabditis elegans. Genes Nutr 14(1):1–13
Zhang C, Cui F, Zeng G-M, Jiang M, Yang Z-Z, Yu Z-G, Zhu M-Y, Shen L-Q (2015) Quaternary ammonium compounds (QACs): a review on occurrence, fate and toxicity in the environment. Sci Total Environ 518:352–362
Zöngür A, Sari M (2023) Herbicides widely used in the world: an investigation of toxic effects on Caenorhabditis elegans. Biologia Futura 1–12
Funding
This work was supported by the Superfund Hazardous Substance Research and Training Program (National Institute of Environmental Health Sciences) (P42 ES027704) and the United States Department of Agriculture (Hatch 6215). The manuscript contents are solely the responsibility of the grantee and do not necessarily represent the official views of the NIH.
Author information
Authors and Affiliations
Contributions
Kelly Rivenbark: study design, execution of in vitro experiments, preparation of original draft. Hasan Nikkhah: software, visualization, writing—review and editing. Meichen Wang: study design, execution of in vitro experiments, writing—review and editing. Burcu Beykal: software, visualization, funding acquisition, writing—review and editing. Timothy Phillips: funding acquisition, writing—review and editing.
Corresponding author
Ethics declarations
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
All authors have read and approved the manuscript for submission and publication.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Bruno Nunes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Rivenbark, K.J., Nikkhah, H., Wang, M. et al. Toxicity of representative organophosphate, organochlorine, phenylurea, dinitroaniline, carbamate, and viologen pesticides to the growth and survival of H. vulgaris, L. minor, and C. elegans. Environ Sci Pollut Res 31, 21781–21796 (2024). https://doi.org/10.1007/s11356-024-32444-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-024-32444-5