
Abstract
Aims
We aimed to identify the optimal combination of selenium (from Na2SeO4) and secondary elements required for obtaining the best nutritive characteristics and quality of 'Qingcui' plums.
Methods
Five treatments (Na2SeO4 with MgSO4 and CaSO4) and a control treatment without Se, Mg, Ca, or S were applied to 2-year-old 'Qingcui' plum trees in a greenhouse. Plum leaves and fruit were analyzed for color, nutrient, and quality characteristics during the growing and deciduous seasons as well as the following year.
Results
Leaf color parameters, pigment content, and specific weight significantly improved by CaSO4 addition during the growing and deciduous seasons. Ca and Mg contents during the growth, defoliation, and young leaf stages of the following year exhibited consistent trends under the different treatments, whereas Se and S contents differed according to the season and treatment. Thus, Mg and S showed high mobility and reutilization rates, whereas Ca and S showed poor mobility and reutilization rates. Leaf and fruit nutrient contents decreased in the order of Ca > Mg > S and S > Ca > Mg, respectively. The combination of 77 mg Na2SeO4, 10 g MgSO4, and 19 g CaSO4 yielded optimal fruit quality in terms of fruit weight, hardness, and soluble solids and fruit Se content. A large amount of Ca was present in the cell wall, whereas S and Mg were mainly observed as soluble components.
Conclusions
Our study revealed the best combination of Se and secondary elements leading to optimal leaf nutrition and fruit Se content and quality in 'Qingcui' plums.
Similar content being viewed by others

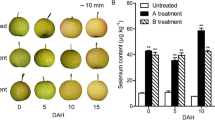
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Selenium (Se) is an essential trace element for humans; Se deficiency in the soil is the main factor leading to Se deficiency in the human body. Globally, the area of Se-deficient soils is much larger than that of Se-rich soils, and in China, approximately 51% of cultivated land is characterized as Se-deficient (Dinh et al. 2018; EI-Ramady et al. 2022). Therefore, the production of safe, high-quality, and Se-rich agricultural products has become a popular research topic in contemporary agricultural sciences in China. Generally, fruits have a weak Se enrichment ability, which can be enhanced by root application or foliar spraying of Se fertilizer. Furthermore, Se benefits crop plants by improving crop growth, quality, and resistance to stress (Golubkina et al. 2019; Pezzarossa et al. 2012; Xu et al. 2022). The soil application of sodium selenate (Na2SeO4) promotes the growth and leaf Se content of 'Qingcui' plum trees more effectively than does the root application of selenomethionine (Sun et al. 2020). However, Na2SeO4 easily migrates and leaches into the deeper soil layers, posing a high environmental risk. Conversely, the addition of organic matter can stabilize Se and improve its utilization rate by plants (Moreno et al. 2013; Wang et al. 2017). Weed compost fertilizer is rich in organic matter, is convenient to obtain, and can replace some chemical fertilizers to improve economic outputs (Sun et al. 2022; Tanveer et al. 2021). Therefore, the combination of Se and weed compost will likely improve the stability of soil Se and the Se utilization rate of crops.
China boasts the largest planting area and highest output of Prunus salicina Lindl in Asia. The planting area of the 'Qingcui' plum in Chongqing, Sichuan, Guizhou, Yunnan, and other provinces is increasing annually, forming a characteristic and favorable 'Qingcui' plum-planting region in Southwest China. The 'Qingcui' plum belongs to the crisp type of plums, with crispness being one of the key determinants of fruit quality that is closely related to its nutritional characteristics, especially the contents of secondary elements. The content of Ca in soft plums is lower than that of Mg (Zhou et al. 2023), while the Ca content in crisp plum leaves is second only to that of nitrogen (N) and significantly higher than that of Mg (Ma et al. 2023; Sun et al. 2023). Ca strengthens the cell membranes and walls; thus, the proper application of Ca fertilizer can reduce the incidence of plum fruit cracking and maintain fruit hardness (Hosein-Beigi et al. 2019; Ma et al. 2023; Yan et al. 2021). Progressive fruit cracking significantly reduces the Ca content and, to a lesser extent, Mg content in crisp plum fruits (Ma et al. 2023).
Selenium and sulfur (S) belong to the same group of elements and exhibit similar metabolic profiles in plants; thus, competition and synergy exist between these two elements (Galeas et al. 2007). However, plants exhibit significant differences in Se, Ca, and Mg metabolism. Hawrylak-Nowak (2008) reported that the application of Se increased Ca content in plants but had no significant impact on Mg and S contents, whereas Xia et al. (2020) showed that root application of Se increased Ca and Mg contents. Conversely, Xu et al. (2022) showed that the root application of Se increased the content of Ca but had no significant effect on Mg, possibly because of the synergistic or antagonistic interactions between these two ions. When plants experience Se stress, Ca is the main element associated with Se (Lanza et al. 2021). Therefore, although treatment with different doses of Se can increase the Se content of wild plants, Ca, Mg, and S contents differ according to the dose and plant species (Drahoňovský et al. 2016).
Senescence is the last process of leaf development. The reactivation of nutrients and the degradation of macromolecules are critical physiological processes in senescence. From the appearance of leaves during senescence, leaf color changes from green to yellow or red, which is a typical leaf senescence phenomenon (Peerzada and Iqbal 2021). This is mainly due to the decrease in various photosynthetic pigments and anthocyanin synthesis, which can be reflected by color parameters and pigment content (Wang et al. 2022). From the perspective of nutrition, leaf senescence is characterized by a transition from assimilation to reuse of nutrients, which is reflected in the decline in leaf weight; plant characteristics and nutrient status affect the reuse of elements (Guo et al. 2021; Jacques et al. 2021; Maillard et al. 2015). In general, the mobility and degree of reuse of Mg in plants is high and the mobility and degree of reuse of S in plants is low, while the mobility of Ca in plants is challenging and the degree of reutilization is very low (Lin et al. 2004; Inail et al. 2022; Oliveira and Santana 2020). However, there are also differences among species. For example, Ca has a certain mobility in Koompassia malaccensis (Gambia), and its reutilization rate is higher than that of Mg (Ong et al. 2015).
The Se application with different combinations of secondary elements on ‘Qingcui’ plum leaf nutrition and fruit quality has not been studied in this plum species before. In this study, we explored the effects of the combined application of Se and secondary elements on the nutrient characteristics of 'Qingcui' plums. Specifically, we mixed the same dose of Se with various essential elements in different proportions and semi-decomposed weeds in the field. We then analyzed the color parameters, pigment content, essential element contents, and leaf nutrient reutilization rate during the growth period (August) and defoliation period (December) as well as the nutrient content in young leaves of the following year (April), fruit quality during the mature period (July), and differences in the subcellular components of essential elements. The aim of this study was to determine the optimal application rate of Se and secondary elements for the efficient production of high-quality 'Qingcui' plums.
Materials and methods
Experimental design and treatments
The experiments were conducted in a multi-span plastic greenhouse in Lidu Town, Fuling, Chongqing, China (29° 45'N, 107° 15'E). The soil is purple clay, with an organic matter content of 15.8 g/kg, a pH of 6.1, and total N, P, K, Ca, Mg, S, Se, and Na contents of approximately 1.45, 2.32, 58.01, 10.73, 15.47, 4.13, 0.00018, and 0.85 g/kg, respectively. The soil Se content is low, the Ca content is at a moderate level, and the Mg, S, and Na contents are at a high level.
Two-year-old 'Qingcui' plum trees (with a rootstock of wild peach (Prunus persica L.)) with uniform growth were planted on ridges on December 12, 2019. The ridge height and width were approximately 40 cm and 150 cm, respectively, and the distances between trees and between ridges were 5 m and 1.5 m, respectively. Trees were maintained under conventional management conditions for more than 1 year. Fertilization was performed on February 6, 2021.
The application of 26.6 mg/plant Se (from Na2SeO4) to 'Qingcui' plum trees can enhance the Se content in the leaves (Sun et al. 2020) and fruit of the following year to those of the standard of Se-enriched fruit. Therefore, in this study, we used 32.18 mg of Se (from 77 mg of Na2SeO4) per plant as the source of Se for plant treatment. CaSO4 is often used as a base fertilizer; it is insoluble in water and was used as a source of Ca and S. MgSO4, which is not easily soluble in water, was used as the source of Mg and S. After mixing Na2SeO4 with CaSO4 and MgSO4 in different amounts, we added semi-decomposed weeds collected from the field and placed the contents into a non-woven bag (20 × 30 cm). Finally, the bag was sealed and applied to the roots of the trees in one hole. The planting hole was 30 cm deep and located approximately 60 cm from the tree trunk. We also performed an experiment without Se, Ca, Mg, or S as a control. The five experimental treatments were as follows: T1: 32.18 mg Se; T2: T1 + 2.02 g Mg + 2.66 g S; T3: T2 + 4.48 g S + 5.59 g Ca; T4: T3 + 4.48 g S + 5.59 g Ca; T5: T4 + 4.48 g S + 5.59 g Ca. Three replicates were included per treatment, and a single tree was used as a biological replicate.
The experimental parameters were measured during the growing season in August 2021, the defoliation season in January 2022, the end of the flowering period in April 2022, and the fruit-ripening season in July 2022 (too little fruit was produced for analysis in 2021). In the experimental stage, i.e., the dormancy period of the 'Qingcui' plum (February 2022), the bagged fertilizer was dug out, the non-woven bags were replaced, and Se, Ca, Mg, S, and semi-decomposed weeds were replenished.
Leaf color
A precision colorimeter (WR-10QC, Weifu, Shenzhen, China) was used to measure the lightness (L*), redness (a*), and yellowness (b*) values. During the analysis, we avoided measurements at the leaf vein, repeated the measurement for three trees per treatment, and assessed 20 leaves randomly on each tree. We then calculated the color saturation (chroma, C) and color shading (hue, H) using a* and b*\(\left(\text{C}=\sqrt{\left(\text{a}\right)^2+\left(\text{b}\right)^2}\right)\) and \(h=\text{tan}^{-1}(\text{b}/\text{a})\), respectively (Adhikary et al. 2021), to quantify pigment concentration and leaf color.
Specific leaf weight
We collected leaf blades and used a perforator to obtain six disks with a diameter of 4.9 mm, which were placed in a drying oven at 80 °C, dried, and weighed to calculate the specific leaf weight (leaf dry weight/leaf area).
Pigment content in leaves
Six round leaf disks from the same area were obtained from the leaves using a perforator. These samples were extracted with 95% ethanol in the dark at 4 °C until the leaves completely lost their green color (Sun et al. 2020). Next, the chlorophyll a, chlorophyll b, chlorophyll a + b, and carotenoid contents were measured at 556, 649, and 470 nm using a UV 16A spectrophotometer (Shimadzu Corporation, Kyoto, Japan) (Lichtenthaler and Wellburn 1983).
Total Se, Ca, Mg, and S contents and plant stem diameter
Twenty leaves from the short branches of the long branches at the periphery of the tree were collected, dried at 60 °C, and ground. Se, Ca, Mg, and S contents were determined according to the method described by Wang et al. (2021). Briefly, after the sample was soaked in 5 mL nitric acid solution overnight, it was heated at 160℃ for decomposition for 4 h. After cooling to room temperature about 25 °C, the sample was diluted to 25 mL with 1% nitric acid solution. Inductively coupled plasma mass spectrometry (Flash 2000, Thermo Fisher Scientific, Massachusetts, USA) was used to determine the elemental contents. The nutrient element reutilization rate was calculated as (leaf element content in growth period—leaf element content in following period)/leaf element content in growth period × 100% (Lin et al. 2004). In January 2022, a vernier caliper was used to measure the diameter of the plant stem 15 cm above the ground.
Determination of fruit quality
Twelve fruit weights were measured using an electronic scale, and the transverse and longitudinal diameters of the sampled fruit were measured using vernier calipers. The hardness of the fruit was measured using a sclerometer (Lvbo, GY-1, Hebei, China), and the soluble solids were measured using a digital refractometer (Atogo, ATC-1, Tokyo, Japan). The fruit titrate acid content (%) was determined using 0.1 mol L−1 of NaOH with phenolphthalein as the pH indicator.
Isolation and determination of Ca, Mg, and S in fruit subcellular fractions
First, a 0.5 g leaf sample was weighed, added to 50 mmol L−1 Tris HCl buffer (pH = 7.5), 250 mmol L−1 sucrose, 1 mmol L−1 dithioerythritol (C4H10O2S2), 5 mmol L−1 ascorbic acid, and 10 mL mixed extractant, and then ground into a homogenate. The homogenate was centrifuged at 4000 rpm for 15 min at room temperature, with the residue representing the cell wall component. The supernatant was centrifuged at 16,000 rpm for 45 min; thereafter, the precipitate comprised the organelles and the supernatant represented the soluble cellular components. After drying, 5 mL nitric acid was added to dissolve all components, which were then transferred to a polytetrafluoroethylene digestion tank for overnight immersion. We then covered the inner lid and kept the sample at 150℃–170℃ for 4 h–6 h before dilution to 25 mL with 1% nitric acid (Wang et al. 2023). Subsequently, the Ca, Mg, and S contents were determined using an inductively coupled plasma optical emission spectrometer (ICAP7200HS Duo, Thermo Scientific, USA).
Data analysis
The experiment was conducted using a completely randomized block design with six treatments and three replications per treatment. The data were analyzed using the Fisher least significant difference test and t-test at p < 0.05 using Origin 8.5 software. Significant differences between means were assessed using the analysis of variance (one-way ANOVA) procedure.
Results
Leaf color
During the growing season, leaf L* values were significantly higher in the treatment groups T4 and T5 than in treatment groups T2 (Na2SeO4 with MgSO4) and T3 (T2+4.48g S + 5.59g Ca) (Fig. 1). However, the L* values were not significantly different from those of the control. Simultaneously, the a* values in the T4 and T5 treatments were significantly lower than those in the T2, T3, and control treatments. Both b* and C values were the highest in the T5 treatment (p < 0.05). Trees under the T5 treatment showed the lowest h value (significantly lower than that in all other treatments except for that of the control). The application of Se and secondary elements affected the color of plum leaves to varying degrees. The L* and h values of leaves were the highest in the T5 group, followed by those in the T4 group.

Effect of Se in combination with secondary elements on plum leaf color during the growth phase (A) and deciduous season (B). Different lowercase letters indicate significant differences between treatments (P < 0.05). T1: 32.18 mg Se; T2: T1 + 2.02 g Mg + 2.66 g S; T3: T2 + 4.48 g S + 5.59 g Ca; T4: T3 + 4.48 g S + 5.59 g Ca; T5: T4 + 4.48 g S + 5.59 g Ca
In the deciduous season, the L* value in the T5 group was significantly higher than that in the other treatments. The L* value of T3 was significantly lower than that of the other treatments (except for that of T2). The a* value of T5 was larger than 0.00, indicating that the leaves began to turn red, whereas the a* value of other treatments was still negative, indicating green leaves. The lowest a* values, indicating the greenest leaves, were observed in the control and T1 groups. The b* value gradually increased with an increase in Ca application amount, reaching a maximum in T5 and indicating that the color was the yellowest under this treatment. The C and b* values were consistent with those of a*; the values were significantly higher in the T5 group than in the other groups, whereas the difference between the other treatments was not significant. This indicates that the T5 treatment, containing 57 g of CaSO4, promoted the change in leaf color to yellow and red.
Specific leaf weight and leaf pigments
During the growing season, the specific leaf weight increased with increasing fertilizer dosage and reached a maximum value under the T5 treatment, which was significantly higher (35.10%) than that in the control treatment (Table 1). The chlorophyll and carotenoid contents also varied under different treatments. The content of chlorophyll a was significantly higher under the T2 and T5 treatments than under the control treatment, by 71.79% and 64.10%, respectively. The content of chlorophyll b was also significantly higher under the T2, T3, and T5 treatments than under the control treatment by 25.00%, 31.25%, and 43.75%, respectively. Chlorophyll a + b content was significantly higher in T2 and T5 than in the control (60.00% and 58.18% of the control, respectively). Finally, chlorophyll a/b ratios were significantly higher in the T3, T4, and T5 groups than in the control, by 11.67%, 11.25%, and 15.83%, respectively. Different treatments significantly increased the content of carotenoids, with the carotenoid contents in T5 and T2 being higher than those in the control (70.83% and 65.28%, respectively). Specific leaf weight and leaf pigment content were significantly increased by the combined application of Se and secondary elements.
During the deciduous season, the specific leaf weight and chlorophyll content were significantly lower than those during the growing season. Moreover, we observed no significant differences in specific leaf weight between the different treatments and the control. The contents of chlorophyll a, b, and a + b were the highest under the T3 treatment, significantly higher than those in the control group by 36.36%, 85.71%, and 59.26%, respectively, and significantly higher than those seen under the other treatments. The T5 treatment showed the lowest contents of chlorophyll a, b, and a + b, which were lower than the control by 54.55% (p < 0.05), 28.57%, and 44.44% (p < 0.05), respectively. The chlorophyll a/b value was the highest in the control group; however, this difference was not significant. Similarly, the carotenoid value was also the highest in the control treatment (p < 0.05) and significantly lower under the T4 and T5 treatments by 50.88% and 45.61%, respectively. The degradation of leaf pigment was the slowest under T3 and fastest under T5.
Leaf Se, Ca, Mg, and S contents
During the growing season, the leaf Se content was the lowest in the control group (Fig. 2A). After adding Na2SeO4 fertilizer (T1), the Se content in leaves increased by 102.78%. Compared with T1, the combined application of Se with MgSO4 (T2), as well as MgSO4 plus CaSO4 (19 g, T3; 38 g, T4), had no significant effect on the Se content of the leaves. However, in the T5 treatment with 57 g CaSO4, the Se content of the leaves was significantly increased by 118.57% from that in T1. The Ca and Mg contents were the highest in the control group. Na2SeO4 application (T1) reduced the Ca and Mg contents in the leaves, but not by a significant amount. The T2 treatment significantly reduced the leaf Ca and Mg contents by 24.05% and 17.91%, respectively. We observed no significant difference in Ca or Mg between the control and T3 group; however, the contents of Ca and Mg decreased with an increase in CaSO4 dose. After treatment with 38 g (T4) and 57 g (T5) of CaSO4, the Ca content decreased by 20.17% and 28.59%, respectively, from that in the control, whereas Mg decreased by 18.82% and 30.30%, respectively, from that in the control. Changes in S contents in the leaves were not significant during the growing season.

Effect of Se in combination with secondary elements on the Se, Ca, S, and Mg contents of plum leaves during the growth phase (A) and deciduous season (B) and the new leaves (C) in the following year. Different lowercase letters indicate significant differences between treatments (P < 0.05). T1: 32.18 mg Se; T2: T1 + 2.02 g Mg + 2.66 g S; T3: T2 + 4.48 g S + 5.59 g Ca; T4: T3 + 4.48 g S + 5.59 g Ca; T5: T4 + 4.48 g S + 5.59 g Ca
During the deciduous season, the Se content in the falling leaves was the lowest in the control group and the highest in T1 (273.38% higher than that in the control, p < 0.05; Fig. 2B), followed by T3, T4, and T5, which also had significantly higher Se content than did the control. Conversely, the Se content in T2 was significantly lower than that in T1, but not significantly different from the other treatments. The Ca and Mg contents were the lowest in T5 and significantly lower than those in the control (24.40% and 20.61%, respectively); however, we observed no significant difference between the other treatments and the control. The S content was the highest in T4, which was 13.08% higher than that in the control (p < 0.05); we observed no significant differences between the other treatments and the control. Thus, compared with the changes seen in the contents of other elements, different treatments had the greatest impact on leaf Se contents.
In the following year, the nutrient levels of young leaves mainly originated from their own stored nutrition. The combined application of Se and secondary elements significantly increased the Se content in leaves, especially in T3 and T5, which had 391.49% and 491.42% higher contents than those in the control, respectively (p < 0.05); however, the difference between T1 and the control was not significant (Fig. 2C). The Ca and Mg contents did not differ significantly among the treatments. The S content of young leaves was the highest in the control and 22.23% and 17.41% higher than that in T3 and T4, respectively (p < 0.05).
Stem diameter and Se, Ca, Mg, and S element reutilization
Compared with that seen in the control, the different treatments showed little effect on stem diameter. However, stem diameters were significantly higher in T2 and T4 than in T3, by 12.54% and 12.19%, respectively (Table 2). Se exhibited the lowest mobility of the elements, resulting in a negative Se reutilization rate in the leaves. However, after treatment with 57 g of CaSO4 (T5), Se mobility improved and the reutilization rate was greater than 0.00%. The mobility of Ca in the leaves was also poor, but we observed some reutilization in the control. Se application significantly reduced the reutilization rate of Ca, whereas all other treatments resulted in an insignificant reduction in the reutilization rate of Ca. The combined application of Se and secondary elements reduced the reutilization rate of Ca. Treatments T2 and T3 increased the reutilization rate of elemental S. In general, the reutilization rate of Mg was the highest, followed by that of S, whereas the reutilization rates of Ca and Se were the lowest.
Fruit quality and nutrition
The fruit weight was the highest under the T2 and T3 treatments and the lowest under the T5 treatment (Table 3). The different treatments had little effect on fruit transverse and longitudinal diameters, but the longitudinal diameter was 34.96% higher in T3 than in the control. Fruit hardness was the lowest in T4 (p < 0.05), yet hardness in the other treatments was not significantly different from that in the control. The highest soluble solids content was observed in the T5 treatment and was significantly higher than that under T1 and T4. There was no significant effect on titratable acid among the total treatments. The Se content was significantly lower in the fruit than in the leaves and was not even detected in the fruit in the control. Combining the application of Se and secondary elements significantly increased the Se content in the fruit (except in T5). The Se content of the fruit was the highest in T3, which was significantly higher than that in T2. In summary, fruit quality was comprehensively better under T3 than under other treatments.
The S content in fruit was the highest, followed by those of Ca and Mg. The contents of Ca, Mg, and S in the soluble components of cells were the highest, followed by those in the cell wall and organelles (Fig. 3). The combined application of Se and secondary elements had no significant effect on the Ca contents of organelles, soluble components, or cell walls. Differences between the different treatments were not significant, except that the Ca content of the soluble components of cells was significantly higher (by 101.62%) in T2 than in T4. Moreover, the highest Mg content in the cell wall was found in T2, which was significantly higher than that in T1 and T3 (by 46.60% and 37.78%, respectively). The Mg contents in the organelles under the T1 and T3 treatments were 97.15% and 76.83% higher than those under T2, respectively. The distribution of Se in the soluble components and cell walls was not significant under the different treatments; however, the S content in the organelles was the highest under T1 and significantly higher than that in the control (by approximately 90.87%). The lowest S content was observed in T2; however, this was not significantly different from that in the control.

Effect of Se in combination with secondary elements on the Se, Ca, S, and Mg contents of the organelle fraction, soluble fraction, and cell wall of fruit. Different lowercase letters indicate significant differences between treatments (P < 0.05). T1: 32.18 mg Se; T2: T1 + 2.02 g Mg + 2.66 g S; T3: T2 + 4.48 g S + 5.59 g Ca; T4: T3 + 4.48 g S + 5.59 g Ca; T5: T4 + 4.48 g S + 5.59 g Ca
Discussion
The quantitative analyses of leaf color changes from green to yellow and red using a color difference meter, leaf color parameters, and chlorophyll content are helpful for understanding the senescence process of plants under different treatments (Tang et al. 2016; Wang et al. 2020). Leaf color patterns can also be used to analyze leaf nutrient concentrations (Keskin et al. 2018; Oliveira and Santana 2020). Color comprises three elements, namely lightness, hue, and chroma, which are reflected in the values of L*, a*, C, and total anthocyanin content (Adhikary et al. 2021; Zhou et al. 2022). In the current study, during the growing season, only the treatment containing a high CaSO4 dose (T5) significantly affected the color parameters, with other treatments showing no significant difference from the control. The L* value, which reflects the brightness of the color, increased as brightness increased during leaf aging. The a* and b* values represent the hues and did not change significantly during the senescence of 'Qingcui' plum leaves, indicating that the leaves typically remained green and yellow. The C and H values reflect the chroma; during 'Qingcui' plum senescence, C did not change significantly whereas H increased significantly. Thus, although the pigment concentration was not obvious, the leaves lost their green color and anthocyanins were synthesized. The T5 treatment showed significant color changes. Our results show that the application of nutrients changes leaf color and pigment parameters during leaf senescence. Moreover, the decomposition rate of chlorophyll was higher than that of carotenoids, and the late anthocyanin synthesis leaves were red (T5). Indeed, color parameters are highly correlated with N and Ca content (Keskin et al. 2018). Increases in the application dose of CaSO4 reduced the Ca and Mg contents in the leaves and led to consistent changes in leaf color parameters. Thus, these results also indicate a correlation between leaf color parameters and Ca and Mg contents.
Leaf weight and leaf pigmentation were significantly lower during leaf senescence than during the growing season, which is consistent with the regular leaf senescence law. Under low-Ca or high-Ca stress, the chlorophyll content of plants is low (Weng et al. 2022); however, the foliar application of calcium chloride can delay the loss of chlorophyll from biomass during storage (Rastegar et al. 2022). In this study, treatment with high levels of CaSO4 (T5) had an obvious effect on the leaves during the growing season; colors were more green and yellow, and the specific leaf weight and leaf pigment content were the highest. However, in the defoliation stage, the leaves showed obvious signs of senescence; that is, with an increase in CaSO4 dose, the leaves became increasingly red and yellow. Future experiments over a longer period of time are required to better determine the physiological effects of sodium selenate and essential element ratios on 'Qingcui' plum leaves. The treatment with 19 g of CaSO4 (T3) showed the best effect on leaf color, retaining the yellow and green colors most effectively. In addition, the treatment with sodium selenate and Mg (T2), which is the key metal ion in chlorophyll, significantly increased the leaf pigment content during the growing season.
In this study, leaf and fruit Se contents were notably higher under T3, showing that the application of sodium selenate with Ca, Mg, and S can significantly improve the reutilization rate of Se. One of the nutritional characteristics of 'Qingcui' plums is that Ca is the second most abundant essential element in this fruit. However, in our study, all five treatments reduced the leaf Ca content during the growing season, which may be related to the characteristics of the plant species and local soil (Xu et al. 2022). We also observed a significant negative correlation between Ca content in the soil (CaSO4 dose) and Mg content in the leaves. Thus, attention should be paid to the balance of cations in the soil (Neilsen and Edwards 1982). The Ca content was also notably higher than the Mg content in the leaves and fruit of 'Qingcui' plums, consistent with the results of Ma et al. (2023) and Sun et al. (2023). Furthermore, Se and secondary element contents were significantly higher in the leaves than in the fruit and decreased in the order of Ca > Mg > S > Se and S > Ca > Mg > Se, respectively. Thus, we speculated that S had the strongest transfer ability from the source (leaves) to the sink (fruit). Mg and S showed the highest nutrient reutilization rate in 'Qingcui' plum leaves, followed by Se and Ca, similar to the results obtained for phyllostachys pubescens (Lin et al. 2004), Eucalyptus (Inail et al. 2022; Oliveira and Santana 2020) and so on. Se showed the lowest mobility in the leaves of 'Qingcui' plums and was lost during leaf senescence.
The key determinants of 'Qingcui' plum quality include fruit crispness and sweetness. Secondary elements affect both plum fruit quality and post-harvest storage quality (Cuquel et al. 2011; Dar et al. 2016). The results of this study show that the combined application of Se and secondary elements has a significant effect on the appearance of fruit (single fruit weight, horizontal and longitudinal diameter) but not on the intrinsic quality of the fruit (hardness, soluble solids content, titratable acid content). Fruit Se content was more than 10 g/kg dry weight (except for that under T5 and the control). Thus, dried fruit can be used as a Se supplement; however, the Se content in fresh 'Qingcui' fruit was too low to reach the Se enrichment standard (10–500 μg/kg fresh weight). Therefore, future attempts should increase the dose of sodium selenate to allow the fruit to reach the Se enrichment standard. Fruit firmness is closely related to the composition and structure of the cell walls, where Ca is typically distributed (Wang et al. 2023). In our study, fruit Ca content was mainly distributed in the solid fraction and cell wall and rarely in the organelle fraction. Moreover, the fruit showed the lowest firmness, soluble solids, and Ca, Mg, and S contents under the T4 treatment. Ca was the most abundant element in the fruit cell wall, which may explain the crisp characteristics of 'Qingcui' plums. The appropriate application of Ca fertilizer has been shown to reduce the rate of plum fruit cracking and maintain fruit hardness (Yan et al. 2021). The fruit cracking rate is also significantly correlated with the Ca content in the leaves and soil (Ma et al. 2023). In this study, no obvious fruit cracking was observed in the fruit, and an increase in the CaSO4 dose mainly affected the Ca content in the soluble fraction of cells, with no significant effect on the Ca content in the cell walls and organelle fraction.
Conclusions
The application of Se fertilizer to the roots of trees can improve Se contents both during the growing season and in young leaves. In this study, we observed that the combined application of Se and other secondary elements resulted in a greater increase in Se contents in 'Qingcui' plum trees than did only Se treatment. During the growing season, the addition of CaSO4 increased leaf color parameters, pigment content, and specific leaf weight in a dose-dependent manner. However, over the long term, a high dosage of CaSO4 reduced fruit weight and Se content. Mg showed the highest nutrient reutilization rate, followed by S, whereas the mobility of Ca and Se was poor and was mainly lost in the leaves. Treatment with 77 mg Na2SeO4, 10 g MgSO4, and 19 g CaSO4 showed optimal improvement in leaf nutrition, Se content, and fruit quality.
Data availability
All data are available in agreement with the open data policy.
Change history
07 May 2024
A Correction to this paper has been published: https://doi.org/10.1007/s11104-024-06703-w
References
Adhikary T, Gill PPS, Jawandha SK, Bhardwaj RD, Anurag RK (2021) Efficacy of postharvest sodium nitroprusside application to extend storability by regulating physico-chemical quality of pear fruit. Food Chem 346:128934
Cuquel FL, Motta ACV, Tutida I, Mio LLMD (2011) Nitrogen and potassium fertilization affecting the plum postharvest quality. Rev Bras Frutic 33:328–336
Dar G, Misger F, Kumar A, Rather J (2016) Growth, fruit set and yield of Santa Rosa plum as affected by nitrogen and boron under rainfed conditions of Kashmir Valley. J Applied Nat Sci 8(4):2081–2086
Dinh QT, Cui Z, Huang J, Tran TAT, Wang D, Yang W, Zhou F, Wang M, Yu D, Liang D (2018) Selenium distribution in the Chinese environment and its relationship with human health: a review. Environ Int 112:294–309
Drahoňovský J, Száková J, Mestek O, Tremlová J, Kaňa A, Najmanová J, Tlustoš P (2016) Selenium uptake, transformation and inter-element interactions by selected wildlife plant species after foliar selenate application. Environ Exp Bot 125:12–19
EI-Ramady H, El-Sakhawy T, Omara AED, Prokisch J, Brevik EC (2022) Selenium and nano-selenium for plant nutrition and crop quality. In: Hossain MA, Ahammed GJ, Kolbert Z, El-Ramady H, Islam T, Schiavon M (eds) Selenium and nano-selenium in environmental stress management and crop quality improvement. Sustainable Plant Nutrition in a Changing World. Springer, Cham
Galeas ML, Zhang LH, Freeman JL, Wegner M, Pilon-Smits EA (2007) Seasonal fluctuations of selenium and sulfur accumulation in selenium hyperaccumulators and related nonaccumulators. New Phytol 173(3):517–525
Golubkina N, Zamana S, Seredin T, Poluboyarinov P, Sokolov S, Baranova H, Krivenkov L, Pietrantonio L, Caruso G (2019) Effect of selenium biofortification and beneficial microorganism inoculation on yield, quality and antioxidant properties of shallot bulbs. Plants (Basel) 8(4):102
Guo Y, Ren G, Zhang K, Li Z, Miao Y, Guo H (2021) Leaf senescence: progression, regulation, and application. Mol Hortic 1(1):1–25
Hawrylak-Nowak B (2008) Effect of selenium on selected macronutrients in maize plants. J Elementol 13(4):513–519
Hosein-Beigi M, Zarei A, Rostaminia M, Erfani-Moghadam J (2019) Positive effects of foliar application of Ca, B and GA3 on the qualitative and quantitative traits of pomegranate (Punica granatum L.) cv. ‘Malase-Torshe-Saveh.’ Sci Hortic 254:40–47
Inail MA, Hardiyanto EB, Thaher E (2022) The effect of Acacia mangium harvest residue management on productivity and soil characteristics of a subsequent Eucalyptus pellita plantation in South Sumatra, Indonesia. Forest Ecol Manag 519:120322
Jacques C, Forest M, Durey V, Salon C, Ourry A, Prudent M (2021) Transient nutrient deficiencies in Pea: consequences on nutrient uptake, remobilization, and seed quality. Front Plant Sci 12:785221–785221
Keskin M, Sekerli YE, Gunduz K (2018) Influence of leaf water content on the prediction of nutrient stress in strawberry leaves using chromameter. Int J Agric Biol 20:2103–2109
Lanza MGDB, Silva VM, Montanha GS, Lavres J, de Carvalho HWP, Dos Reis AR (2021) Assessment of selenium spatial distribution using μ-XFR in cowpea (Vigna unguiculata L. Walp.) plants: Integration of physiological and biochemical responses. Ecotoxicol Environ Safe 207:111216
Lichtenthaler H, Wellburn A (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc T 11(5):591–592
Lin YM, Peng ZQ, Lin P (2004) Dynamics of leaf mass, leaf area and element retranslocation efficiency during leaf senescence in Phyllostachys pubescens. Acta Bot Sin 46(11):1316–1323
Ma X, Xiang P, Yuan X, Liu X (2023) Logistic regression analysis of relationship between severity of fruit splitting and mineral element content of Prunus salicina Lindl. J Plant Nutr 46(10):2488–2495
Maillard A, Diquélou S, Billard V, Laîné P, Garnica M, Prudent M, Garcia-Mina JM, Yvin JC, Ourry A (2015) Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front Plant Sci 6:317
Moreno RG, Burdock R, Álvarez MCD, Crawford JW (2013) Managing the selenium content in soils in semiarid environments through the recycling of organic matter. Appl Environ Soil Sci 283468:1–10
Neilsen G, Edwards T (1982) Relationships between Ca, Mg, and K in soil, leaf, and fruits of Okanagan apple orchards. Can J Soil Sci 62(2):365–374
Oliveira L, Santana RC (2020) Exploratory analysis of nutrient concentrations in Eucalyptus leaf color patterns. Adv for Sci 7(2):973–979
Ong CS, Juan JC, Yule CM (2015) Litter fall production and chemistry of Koompassia malaccensis and Shorea uliginosa in a tropical peat swamp forest: plant nutrient regulation and climate relationships. Trees 29(2):527–537
Peerzada YY, Iqbal M (2021) Leaf senescence and ethylene signaling. In: Aftab T, Hakeem KR (eds) Plant growth regulators. Springer, Cham, pp 53–171
Pezzarossa B, Remorini D, Gentile ML, Massai R (2012) Effects of foliar and fruit addition of sodium selenate on selenium accumulation and fruit quality. J Sci Food Agr 92(4):781–786
Sun X, Han G, Ye S, Luo Y, Zhou X (2020) Effects of selenium on serotonin synthesis and the glutathione redox cycle in plum leaves. J Soil Sci Plant Nutr 20(4):2212–2221
Sun X, Luo Y, Han G, Zhang W (2023) Effects of semi-decomposed weeds as substrate on plant growth, antioxidant capacity, and leaf nutrition of plum trees. J Plant Nutr 46(6):823–834
Tang GL, Li XY, Lin LS, Zeng FJ (2016) Impact of girdling and leaf removal on Alhagi sparsifolia leaf senescence. Plant Growth Regul 78(2):205–216
Tanveer A, Asghar MS, Sarwar M, Saleem MF, Nadeem M, Munir MK, Zafar M, Rizwan M, Sarwar G (2021) Improving the productivity of okra (Abelmoschus esculentus L.) by strengthening the impact of applied nutrients through alligator weed compost. Pak J Agr Sci 58(4):1131–1139
Wang D, Zhou F, Yang W, Peng Q, Man N, Liang D (2017) Selenate redistribution during aging in different Chinese soils and the dominant influential factors. Chemosphere 182:284–292
Wang HL, Zhang Y, Xia X, Li Z (2020) Research advances in leaf senescence of woody plants. Sci Sinica Chim 50(2):196–206
Wang P, Fu C, Wang L, Yan T (2022) Delayed autumnal leaf senescence following nutrient fertilization results in altered nitrogen resorption. Tree Physiol 42:1549–1559
Wang C, Tao Y, Han Y, Zhang R, Li L, Gao Y (2023) Influences of subcellular Ca redistribution induced by γ irradiation on the fruit firmness of refrigerated blueberries. Postharvest Biol Tec 195:112146
Weng X, Li H, Ren C, Zhou Y, Zhu W, Zhang S, Liu L (2022) Calcium regulates growing and nutrient absorption in poplar seedlings. Front Plant Sci 13:887098–887098
Xia Q, Yang Z, Shui Y, Liu X, Chen J, Khan S, Wang J, Gao Z (2020) Methods of selenium application deferentially modulate plant growth, selenium accumulation and speciation, protein, anthocyanins and concentrations of mineral elements in purple-grained wheat. Front Plant Sci 11:1114
Xu X, Wang J, Wu H, Yuan Q, Wang J, Cui J, Lin A (2022) Effects of selenium fertilizer application and tomato varieties on tomato fruit quality: a meta-analysis. Sci Hortic 304:111242
Yan G, Yang Y, Li L, Zeng X, Bie Y, Mao H, Wang L, Xiong B (2021) Study on the effect difference of two kinds of control techniques on cracking fruit of Qingcui plum. IOP Conf Ser Earth Environ Sci 792(1):012042
Zhou D, Lin Y, Fang Z, Ye X, Chen W (2022) Correlation and path analysis of fruit quality and mineral element content of ‘Furongli’ from 10 kinds of different origins in Fujian. J Food Saf Qual 13(4):1176–1184
Zhou Y, Ye Y, Zhu G, Xu Y, Tan J, Liu J (2023) Diversity, classification, and EST-SSR-based association analysis of caladium ornamental traits. Physiol Plantarum 175:e13841
Acknowledgements
This research was financially supported by the National Natural Science Foundation of China (Grant No. 31901972).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Martin J. Hodson.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sun, X., Gao, S., Ling, M. et al. Effects of selenium application with different combinations of secondary elements on ‘Qingcui’ plum leaf nutrition and fruit quality. Plant Soil 501, 425–436 (2024). https://doi.org/10.1007/s11104-024-06566-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-024-06566-1