
Abstract
Key message
This study shows that two emergent tree species growing in a nutrient-limited tropical peat swamp forest use different mechanisms for nutrient regulation.
Abstract
The main aim of this study is to understand the contribution of litterfall to nutrient supply and regulation of two emergent tree species in a tropical peat swamp forest (TPSF) in Peninsular Malaysia. The questions posed by this study include: (1) Do coexisting tree species in TPSF differ in terms of litter production, physico-chemical traits and litterfall patterns? (2) How do these relate to possible nutrient regulation mechanisms of the tree species and do the strategies involved differ between a nitrogen (N) fixing legume (Koompassia malaccensis, Family Fabaceae) and a dipterocarp (Shorea uliginosa, Family Dipterocarpaceae)? (3) Is litterfall timing of the selected tree species driven by climatic variables? Litterfall was collected from litter traps placed under the selected trees in the Sungai Karang TPSF. Green leaves of the selected tree species were also collected. Both leaf litter and green leaves were assessed for their physico-chemical properties (toughness, total phenols, total tannins, proximate fibre–cellulose–lignin and a range of nutrients) and compared. Results obtained indicate that K. malaccensis relies on a combination of high litter quality (higher N coupled with lower amount of tannins, fibre and cellulose) and low annual litterfall mass (388.17 g m−2) for nutrient regulation while the opposite strategy is used by S. uliginosa, which was found to produce an annual litterfall mass of 918.14 g m−2. Due to the aseasonal environment, no significant relationships were observed between climatic variables and litterfall. Therefore, litterfall characteristics in aseasonal tropical regions may depend more on species-specific physico-chemical properties than on climatic variables.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As the first and main process that returns nutrients from above-ground living biomass to the forest floor, plant litter forms the forest organic layer, which enriches and maintains the nutrient supply in the soil (Liu et al. 2004). The timing and amount of litterfall produced exert a major control over the forest nutrient cycling process. Therefore, studies of litterfall are useful in providing information on limiting nutrients and nutrient use efficiency (NUE) in forest ecosystems. Litterfall collection is a standard non-destructive method that has been used by global researchers since the 1950s to determine forest net primary production (productivity), phenology and most importantly, turnover rates of organic and inorganic matter (nutrient cycling) (Lowman 1988; Williams-Linera and Tolome 1996; Hansen et al. 2009; Chave et al. 2010). The components of litterfall include leaves, twigs, bark and reproductive organs such as flowers and seeds, but of these, leaf litter contributes to the highest amount of transferrable nutrients via decomposition (Chave et al. 2010).
Over the years, the effects of litterfall on nutrient cycling have been studied extensively especially in terms of nutrient return via leaf litter decomposition. However, to further understand the underlying plant nutrient use strategies, it is also helpful to compare nutrient and physico-chemical properties between green leaves and senesced leaves (Hättenschwiler et al. 2008; Biswas and Khan 2011), but such studies are rare, particularly in the tropics (Hättenschwiler et al. 2008). Plants can use different strategies to efficiently manage their nutrients such as external cycling, which relies on mineralization of nutrients from the surrounding environment or litter decomposition and internal cycling (Hättenschwiler et al. 2008). The latter, which refers to the withdrawal or resorption of nutrients from senescing leaves, is a common strategy used by plants growing in nutrient-poor environments (Hättenschwiler et al. 2008).
Besides studies of litterfall on nutrient cycling, global research on litterfall often focusses on the effects of climatic variables on litter production, notably temperature (maximum and minimum), precipitation and seasonality (Williams-Linera and Tolome 1996; Liu et al. 2004; Chave et al. 2010), as well as forest type (varying with latitude and altitude) (Bray and Gorham 1964; Miller 1984; Campanella and Bertiller 2008). In tropical forests, patterns of litterfall have been reported to be associated with precipitation, seasonality and temperature but large variations have been obtained between sites and researchers. These differences may be due to factors such as the placement of traps, types of litter sorted, frequency of litterfall collection and duration of the study. In a recent meta-analysis conducted on litterfall studies, Chave et al. (2010) supported Proctor’s (1983) claim that one year is sufficient to capture the annual litter production in a forest as they found that the inter-annual variability falls within 10 % of the mean litterfall.
In this study, we focussed on the nutrient-poor TPSF of SE Asia, where the dome-shaped topography prevents inflow of water which would otherwise supply an input of nutrients, as observed in other types of wetlands (Pahang Forestry Department 2005). Even during extreme rainfall or flooding events, nutrient input to the ecosystem is restricted to the edges or immediate riparian zones of the forest. Nutrient input in the central part of the forest is thus solely dependent upon rainfall, dusts, marine aerosols, litterfall and plant litter decomposition (Yule 2010). This may explain results obtained by Chave et al. (2010) which reported that flooded forests had the highest amount of annual litterfall as compared to other forest types. Although litterfall and its contribution to nutrient input is well documented across various ecosystems, our knowledge of flooded, disturbed, regenerating, or nutrient-poor environments such as TPSF is still limited.
Decomposition of leaf litter in tropical peat swamps is severely limited as a result of the extreme conditions (Yule and Gomez 2009; Yule 2010) and consequently they are of global importance in carbon sequestration, storing 2–3 % of total soil carbon (Page et al. 2006). Litter builds up as peat in layers up to 25 m deep, and when these forests are disturbed through logging, draining and agricultural conversion they are extremely susceptible to fire. Regional peat fires in 1997 resulted in the highest recorded increase in global carbon dioxide (CO2) levels since records began in the 1950s (Page et al. 2002; Aldhous 2004). Given the severe threats presently faced by these forests, further ecological research is crucial to fully understand their role in global climate change.
Two emergent tropical tree species were selected for this study, a dipterocarp Shorea uliginosa Foxw. (family Dipterocarpaceae) and a legume Koompassia malaccensis Magingay ex Benth. (family Fabaceae, subfamily Caesalpinioideae), which represent species with different adaptations to survive in such nutrient-limited environments. The commercial value of both plants is well known but little is known about their ecological importance especially in terms of effects on TPSF soil and subsequent sustainability of the forest. The most notable leaf physico-chemical properties involved in litterfall and nutrient cycling are the plant secondary compounds (especially lignin concentration), nutrient concentrations and leaf area (e.g. Campanella and Bertiller 2008).
In this study, we have examined the potential nutrient use mechanisms revolving around plant foliage and leaf litterfall that enable Koompassia and Shorea to persist in the nutrient-poor environment of the TPSF. Green leaves and leaf litter of the selected tree species were analysed with a focus on nutrient concentrations as well as physico-chemical properties. Litterfall patterns, and effects of climatic factors notably precipitation and temperature were also examined. The general working hypotheses of this study are: (1) Koompassia and Shorea will rely heavily on resorption of nutrients before litterfall as both tree species grow in a nutrient-poor environment, and (2) Koompassia will show greater nutrient retranslocation or resorption compared to Shorea as the former is a nitrogen-fixer and thus has a higher amount of nutrients in its living tissues. As a result, Shorea was predicted to produce lower quality, but a higher quantity of leaf litter as it was hypothesized to depend more on nutrient uptake from decomposing litter. (3) Lastly, litterfall patterns of the selected tree species were hypothesized to be affected by both leaf litter physico-chemical properties and climate.
Methods
Study area
This study was conducted in the Sungai Karang TPSF (N 3°39′30.8″E 101°19′18.4″), a protected low-lying forest reserve on the west coast of Peninsular Malaysia with an area of approximately 23,486 Ha. It is the second largest contiguous peat forest block remaining in Peninsular Malaysia after the Pekan-Nenasi peat forest on the east coast. The forest is quite diverse with 107 tree species from 27 different families recorded including Aglaia odorata Pannell C.M., Campnosperma coriaceum [Jack] Halllier f., Cryptocarya impressa Miq, Ixora grandiflora Ker Gawl., Macaranga pruinosa Mull.Arg., Parartocarpus venosus [Becc.] Garrett, Pternandra galeata Korth., and Shorea platycarpa Heim. (Hahn-Schilling 1994), as well as ferns (Stenochlaena palustris [Burm.f.] Bedd, Nephrolepis biserrata [Sw.] Schott., Asplenium longissimum BI., Dicranopteris sp.), and palms (Pinanga sp., Ptychoraphis sp., Korthalsia sp., Pandanus helicopus Kurz ex Mig.). The main soil substrate of this forest is peat (histosols) of about 5 m in depth lying above marine alluvial clay (the area was previously probably covered by mangrove forest). The forest experiences a fairly stable tropical climate throughout the year but the monsoon brings heavier rainfall to the forest from September to December. The peat substrate is permanently waterlogged and the forest floor is submerged during wet periods. The water is acidic (pH 3–4), and dark brown in colour due to high concentrations of tannins and humic acids (DOC typically 80 mg L−1) (Yule and Gomez 2009). It is anaerobic because of lack of water flow. Inorganic nutrient concentrations are also low (nitrogen and phosphorus <0.1 mg L−1).
At the time of the study, the forest lies adjacent to Malaysian Federal Land Development Authority’s (FELDA) oil palm plantation and Integrated Agricultural Development Project’s (IADP) paddy fields (rice). During the study, a large area of peat forest adjacent to the forest reserve was drained and cleared for new oil palm development. This drainage would have impacted the hydrology of the remaining forest.
Litterfall and living leaves collection
Four sites within the forest with Koompassia and Shorea were chosen for the study. At each site, four 1 × 1.5 m2 litter traps were randomly established under Koompassia and Shorea canopy, respectively, with two traps per tree as placement of traps, in closed canopy relative to the cardinal directions and distance from tree trunks, does not affect the amount of leaf litter collected (Lowman 1988). The traps were made of non-degradable nylon mesh and were tied to adjacent tree trunks 1 m above the ground using nylon ropes. Litterfall was collected monthly from March 2011 to February 2012. All litter trap samples were retrieved and brought back to the laboratory immediately to be sorted according to the litter components of Koompassia leaf litter, Shorea leaf litter, wood litter of unidentified species and residual litter (reproductive organs and leaf litter of other unidentified plant species). Large woody materials such as branches were also included in the study. They were broken into pieces and brought back to the lab to be weighed. The dry weight (g) of all the samples was obtained after oven-drying of each litter component at 60 °C for 48 h. Mature green leaves of Koompassia and Shorea were also collected from the study site. As both are emergent tree species, six smaller trees (>2 m) of each species were sampled using a tree pruner.
Chemical analysis
In the laboratory, sub-samples of living leaves and leaf litter of Koompassia and Shorea were air-dried and ground for determination of total Kjeldahl nitrogen (N) and phosphorus (P). Total N was extracted using a block digester method whereby samples were digested using sulphuric acid before being converted to ammonium cation through the use of a copper sulphate catalyst and addition of potassium sulphate. The pH of the digested sample was then altered to a known basic pH to convert the ammonium cation to ammonia. Concentration of total N was determined colorimetrically using the Lachat Quikchem® flow-injection analysis system after heating the buffered sample with salicylate and hypochlorite to produce a blue-coloured solution. Sodium nitroprusside was used to intensify the colour of the solution. Total P was determined by converting the sample P to orthophosphates (PO4 3−) via digestion in the presence of sulphuric acid and persulphate. The concentration of orthophosphates was determined colorimetrically by pH-adjusted molybdate tartrate-ascorbic acid method using the Lachat Quikchem® flow-injection analysis system.
Sub-samples of the air-dried and ground living leaves and litter were also analysed for sodium (Na), magnesium (Mg), potassium (K) and calcium (Ca) contents. The leaves were dried at 105 °C, weighed and converted to ash via combustion at 550 °C in a furnace for 4 h. The ash was subsequently digested in 2 N hydrochloric acid and analysed for Na, Mg, K and Ca using an acetylene-air flame atomic absorption spectrophotometer (AAS, Perkin Elmer 3100).
Total phenols, tannins as well as proximate lignin, cellulose and fibre content of litter were also quantified following the methods in Graca et al. (2005). Briefly, successive removal of tissues using acid detergent, 72 % sulphuric acid (H2SO4) and ignition of samples at 550 °C for 3 h in a muffle furnace were done. Determination of remaining tissue weight after each step was conducted to calculate the proximate lignin, cellulose and fibre content. Total phenol and tannin contents were determined following Folin–Ciocalteu’s assay using 70 % acetone as extraction solution at 4 °C. For total tannin determination, polyvinylpolypyrrolidone (PVPP) was added to the sample to chelate the tannins before carrying out Folin–Ciocalteu’s assay.
Physical analysis
To determine the leaf area, leaves were placed on a grid paper (1 mm × 1 mm) and the number of squares intercepted by the leaves were counted. Leaf toughness was assessed using a penetrometer based on the protocol outlined in Graca et al. (2005).
Environmental variables
In this study, the environmental variables included monthly rainfall data, rain days and mean maximum/minimum temperatures. The data obtained from the nearest weather station—MARDI Tanjong Karang—were provided by the Malaysian Meteorological Department (MMD).
Calculation
The % nutrient retranslocation efficiency (NRE) and nutrient use efficiency (NUE) were calculated according to Finzi et al. (2001):
-
1.
%NRE = [(A − B)/A] × 100, whereby A refers to the nutrient concentration in green leaves, while B refers to the nutrient concentration in leaf litter,
-
2.
NUE = annual litterfall mass (g m−2)/litterfall nutrient content (g m−2).
Data analysis
Kurtosis and skewness as well as Levene’s test of homogeneity with the critical value of 0.05 was applied to evaluate the normality of the collected data. Analysis of variance (ANOVA) was then used to examine potential significant differences in litterfall components between months and the physico-chemical properties between the sample types of each tree species. ANOVA was also used to examine differences in mean monthly maximum and minimum temperature. No statistical test was conducted on annual litterfall and rainfall data because total values for the year and month were used, respectively. Tukey’s post hoc test was used to determine differences between the different treatments as stated above. The relationship between litter physico-chemical properties of each tree species were also examined using principal component analysis (PCA). For tree species ordination according to these litter traits, the loading coefficients of each species were calculated with respect to the first two principal components. Lastly, Pearson’s correlation was used to evaluate the relationship between litterfall for each species and climatic variables. All statistical analyses were conducted using SPSS 16.0 and the accepted level of significance was set at 0.05.
Results
Litterfall of Koompassia and Shorea in Sungai Karang peat swamp forest, North Selangor
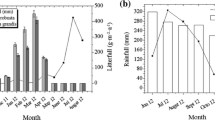
As shown in Fig. 1, leaf litterfall occurred throughout the year and varied across the months for both Koompassia (F 1,144 = 4.492, p < 0.01) and Shorea (F 1,144 = 4.492, p < 0.01). The timing of leaf litterfall differed between the two species whereby maximum leaf litter was observed in the month of June–July for Koompassia and December for Shorea. The data depicted single major leaf shedding events in the year for both tree species but the wood litter fraction did not vary much throughout the year for either tree species. The high variability observed for monthly wood litter collections was due to the inclusion of large branches collected occasionally in the litter traps. Residual litter (reproductive organs and unidentified leaf litter) collected from Koompassia traps recorded highest values in October and no major differences were observed throughout the remaining year. No major differences were observed for residual litter collected from the Shorea trap.

Monthly leaf, wood and residual (reproductive organs and leaf litter of other unidentified plant species) litter of Shorea and Koompassia. Monthly values are expressed as g m−2 ± SD (n = 8). Any two treatments sharing a lower case letter indicate amounts that are not significantly different
The total annual leaf litterfall for Koompassia and Shorea were 388.17 and 918.14 g m−2, respectively. Annual leaf litter production of Shorea was twofold higher as compared to that of Koompassia. For the wood and residual litterfall, Koompassia recorded 653.94 and 1,567.19 g m−2 while Shorea recorded 523.91 and 1,353.31 g m−2, respectively. In this study, residual litter accounted for the highest fraction of total litterfall for both tree species with a value of 60.10 and 48.31 % for Koompassia and Shorea, respectively. For litter traps placed under Koompassia, the smallest litter fraction was leaf litter at 14.83 % while wood litter recorded a higher fraction at 25.08 %. The opposite was observed for litter traps placed under Shorea where wood litter accounted for only 18.71 % while species leaf litter recorded a value of 32.98 %. Total annual litterfall of the selected plant species could not be determined as the research was conducted in a tropical forest of high diversity resulting in a mixture of plant components from many different plant species, which could not be accurately separated in the litter trap.
Nutrient quality and physico-chemical traits of green leaves and litter of Koompassia and Shorea
Table 1 shows the nutrient concentrations of the leaf litter collected. Significant differences were observed between all samples for N (F 3,20 = 60.257, p < 0.01), Ca (F 3,20 = 15.633, p < 0.01), Mg (F 3,20 = 26.575, p < 0.01), Na (F 3,20 = 24.340, p < 0.01) and K (F 3,20 = 75.001, p < 0.01). Koompassia green leaves had the highest concentration for all nutrient elements studied except for P as there were no differences (F 3,20 = 5.058, p = 0.120) observed for P concentrations among the four sample types collected. Koompassia litter had similar nutrient concentrations to Shorea litter except for N, which was significantly higher in Koompassia litter, and Na, which was significantly higher in the latter. However, the higher annual mass of Shorea litter will result in higher annual nutrient return per unit area to the forest when compared to Koompassia. When comparing green leaves to leaf litter, the data revealed consistently higher nutrient concentrations in Koompassia green leaves compared to its leaf litter. While Shorea leaf litter recorded higher N and Mg values compared to its green leaves, no differences were observed for the remaining nutrient elements.
Aside from nutrient concentration, it is also important to determine the leaf litter nutrient stoichiometry. Among all the nutrients listed in Table 1, N and P are the most common nutrients analysed as they are key nutrients required for plant growth. Therefore, only the leaf N:P ratios were reported. When leaf N:P ratio is <14, it indicates N limitation while values >16 indicate P limitation (Koerselma and Meuleman 1996). In this study, green leaf N:P ratios for Koompassia and Shorea are 102.0 and 35.5 which suggests P limitation for tree growth at the study site. Compared to green leaves, the leaf litter N:P ratios of both tree species are 62.27 and 50.23, respectively.
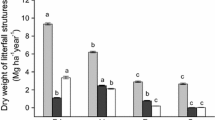
Table 2 shows the leaf litter physico-chemical traits of both tree species. These differed for leaf area (F 1,38 = 505.26, p < 0.01), total phenols (F 1,38 = 23.49, p < 0.05), total tannins (F 1,8 = 12.19, p = 0.013), fibre concentration (F 1,8 = 20.33, p < 0.05) and cellulose concentration (F 1,8 = 507.77, p < 0.01). Shorea had higher leaf area and concentration of these litter chemicals compared to Koompassia except for phenol concentration where it was higher in the latter. No differences were observed for leaf toughness and lignin concentration between litters of the two species.
Annual changes in nutrient use efficiency and nutrient retranslocation efficiency of Koompassia and Shorea
Percentage nutrient retranslocation efficiency for Koompassia showed that it was capable of retranslocating its nutrients (except P) before leaf fall while negative values of % NRE for Shorea indicated that its nutrients (except for K) were retained in the senescing leaves (Table 3). However, this should be interpreted with care as only N and Mg concentrations differed between the green leaves and litter of Shorea (Table 2). This suggests that only N and Mg were retained in the senescing leaves of Shorea and the tree is not capable of retranslocating the rest of its nutrients except for K. Nutrient use efficiency depends on the ability of plants to reabsorb nutrients from the soil and also on their ability to store and use the nutrients (Biswas and Khan 2011). In this study, the NUE of Koompassia is low compared to the values reported by Biswas and Khan (2011) which ranged 103–5,428 but the % NRE was higher. This suggests that the nutrient retranslocation in Koompassia is high and it also suggests that the tree is suffering from nutrient limitation; thus it is minimizing the loss of nutrients to the environment. The NUE of the different nutrient elements in Koompassia was ranked as follows: Mg > Ca > N > K > Na > P.
Relationships among the leaf litter nutrient quality, physico-chemical traits and leaf litter types
Replicates of leaf litter samples from both tree species formed two separate clusters (Fig. 2) indicating that both tree species produced leaf litter with significantly different physicochemical properties. The data also indicate that Koompassia produces higher quality litter compared to Shorea. Koompassia litter exhibited higher nutrient content (total nitrogen, total phosphorus, calcium, magnesium and potassium) and also higher % lignin (Fig. 2). Shorea litter had low amounts of nutrients but higher total tannin content, % cellulose, % fibre and leaf area. PC1 was shown to explain 75.3 % of the variations observed between the two species and both PCs combined account for 84.8 % of the variations observed. This indicates that the variables measured are sufficient to explain the differences observed between the leaf litters of the two species.

Ordination of plant physico-chemical properties of K. malaccensis and S. uliginosa with respect to the first two principal components of the correlation matrix. Plots that cluster together have similar physico-chemical properties
Climatic measurements
During the study period, the total precipitation was 2,002 mm, which was in the range of annual rainfall in Peninsular Malaysia (2,000–3,000 mm). According to MMD, there are two main rainfall peaks in a year, which generally occur during October to November and April to May. Results obtained showed that the highest rainfall occurred in November followed by August and May. The study period showed a similar rainfall pattern to the general annual rainfall pattern in Peninsular Malaysia but a peak was also observed in August. Minimum rainfall generally occurs in January to February and June to July. However, during the study period minimum rainfall was observed in December and April. Mean maximum and minimum temperatures at the study site were stable throughout the study period with values of 31.6 ± 0.5 and 23.8 ± 0.2 °C, respectively (Fig. 3).

Monthly records of total rainfall, minimum temperature and maximum temperature at Sungai Karang, North Selangor region, from March 2011 to February 2012. Temperature values are expressed as mean ± SE
Relationship between litterfall and climatic variables
When the data between litterfall and climatic variables were analysed, significant correlation was only observed between monthly litterfall of Shorea and rainfall (r = −0.644, p = 0.024) (Table 4). The negative correlation obtained indicates higher leaf litterfall during the dry season, as shown by the highest recorded value of Shorea leaf litterfall (Fig. 1) in the driest month (December 2012) of the study period (Fig. 3). No correlations were observed between monthly litterfall and temperature which is as expected because temperature showed little variation throughout the study period.
Discussion
Litterfall production of Shorea and Koompassia
In this study, the annual leaf litterfall mass of Shorea (918.14 g m−2) and that of Koompassia (388.17 g m−2) were comparable to data obtained from other tropical regions where annual species leaf litterfall ranged between 210 to 970 g m−2 (Cuevas and Lugo 1998; Hansen et al. 2009; Chave et al. 2010). Broadleaf forests generally have high litterfall, and among these, tropical moist broadleaf forests produce the highest litterfall, which can be attributed to the high productivity and high allocation of biomass to leaves in tropical regions (Liu et al. 2004).
When comparing litterfall mass between the two selected tree species, Shorea produced higher mass of leaf litter compared to Koompassia. Litterfall mass has been particularly noted to be associated with leaf lignin content. This point has been highlighted in a study conducted by Campanella and Bertiller (2008) in Patagonian Monte, a northeastern region of Chubut Province (Argentina), whereby high leaf litterfall masses of Chuquiraga avellanedae, Chuquiraga hystrix and Atriplex lampa were found to be associated with their high leaf lignin concentration. In this study, both leaf litter species had comparable lignin concentrations but the leaf size of Shorea was markedly greater than Koompassia, which could possibly be the main factor that explains the difference in litterfall masses between the two species. Nevertheless, it should be noted that the lignin concentration for a collection of other plants in published datasets is much lower (4–13 %) than the lignin concentration of Koompassia and Shorea (≈40 %). The high lignin concentrations could be an adaptation to reduce herbivory in the low nutrient environment of the peat swamp (Cornelissen and Thompson 1997). Trees growing in nutrient-poor ecosystems are also known to produce resistant litter to reduce the rate of litter decomposition and thus, nutrient mineralization in order to prevent surrounding fast-growing plants from outcompeting them.
Regulation of nutrients via litterfall and retranslocation within plant
The chemistry of the green leaves of the selected tree species was also compared to their leaf litter to detect any possible mechanism that could be used by the tree to conserve nutrients. The N and P concentration of green leaves and leaf litter of both tree species were comparable to other evergreen tropical plants but the concentrations of other nutrients were much lower (Cuevas and Lugo 1998, Campanella and Bertiller 2008). To examine possible mechanisms that may be involved in nutrient management, NUE and %NRE have been commonly used to describe the phenomenon. Results showed that the Koompassia had positive %NRE and NUE values while mostly negative values were obtained for Shorea. This suggests that Koompassia is capable of reabsorbing or retranslocating its nutrients from its leaves before leaf abscission while Shorea is not. It also suggests that Koompassia is capable of using the nutrient elements more efficiently, which enables it to survive in the extreme condition of the TPSF. Recovery of foliar nutrients from senescing leaves is a common phenomenon in plants growing on nutrient-poor soils (Milla et al. 2005) as this allows the plant to gain some independence from the soil nutrients. Aside from nutrient uptake, recovery of nutrients is also an important component in nutrient conservation as it increases the residence time of nutrients within plants and reduces loss of nutrients to the surrounding environment. The results are in accordance with the study done by Milla et al. (2005) whereby plants with higher nutrient status are capable of recovering more nutrients from their senescing leaves as they have a higher amount of retranslocable nutrients as compared to low nutrient status plants. This explains the higher difference in nutrient content observed between Koompassia green leaves and its leaf litter as compared to Shorea, and this could be related to the fact that, as a legume, Koompassia is able to fix nitrogen. However, care should be taken in making conclusions using the results obtained as a solid conclusion can only be made about the selected tree species when a larger study involving the entire area of the forest for an extended period of time is conducted. Such studies should also include the study of the selected tree species in other TPSF.
The higher N and Mg content in Shorea litter as compared to its green leaves may be due to nutrient assimilation in the senescing leaves of Shorea. Similar results of negative %NRE have also been obtained in other studies involving Phyllostachys pubescens Mazel ex J. Houz (Lin et al. 2004) as well as mangrove trees such as Kandelia candel (L.) Druce, Rhizophora stylosa Griff., Bruguiera gymnorrhiza (L.) Lamk (Lin and Wang 2001), Rhizophora harrisonii Leechm (Ricardo 1989) and Arctostaphylos patula Greene (Schlesinger et al. 1989). As Shorea did not display an ability to recover N from its leaves, this suggests that it is dependent on available N from the soil and subsequent uptake of N by its roots. For total P content, data obtained differ from the remaining plant nutrients measured whereby no differences were observed between the green leaves and leaf litter of all samples. This can be an indication that both Koompassia and Shorea do not recover P from their senescing leaves but rely more on uptake of P by their roots. Another possible explanation for this is the acidic conditions of the forest itself (pH ≤4) and the interaction with peat microorganisms. According to Riswan (1989), acidic conditions facilitate the chelation of phosphate complexes which then enables plant extraction and uptake of P from peat.
Aside from foliar nutrient recuperation, trees are also known to use leaf chemicals and litterfall mass to regulate the nutrient return to the forest substrate via litterfall. The main nutrient cycling process that is affected by this is litter decomposition. Leaf chemical compounds such as phenols, tannins, fibre, cellulose and lignin have been reported to negatively affect decomposition by inhibiting processes such as N fixation and mineralization (Kraus et al. 2004; Wurzburger and Hendrik 2007) as well as acting as toxins to neighbouring plants and microorganisms (Kuiters 1990; Hattenschwiler and Vitousek 2000; Fierer et al. 2001; Bais et al. 2003; Meier and Bowman 2008; Ushio et al. 2008). In this study, Shorea leaf litter had higher concentrations of plant defensive chemicals such as tannins, fibre and cellulose while Koompassia had higher concentration of phenols. Overall, Koompassia produces higher quality litter with higher nutrient content and lower concentrations of secondary compounds compared to Shorea (Fig. 2). Despite the lower mass of Koompassia malaccensis litter, its higher quality can provide an important pulse of nutrients to its surrounding environment, which will enable it to survive in the harsh nutrient-poor environment of the TPSF. Shorea may compensate for this by producing larger mass of litter. Therefore, Koompassia and Shorea both regulate the nutrient return via litterfall through different mechanisms whereby Koompassia may rely on high quality litter despite low litterfall mass while the opposite was observed for Shorea. These particular mechanisms have also been observed and described in other studies (Lugo et al. 1990). In order to better understand the mechanisms involved in litterfall and nutrient return of the selected tree species, a study that involves the identification and analysis of all the litter components (flowers, seeds, fruits, etc.) belonging to the selected tree species would be ideal. However, this proves to be a difficult process in a tropical forest due to the high diversity of plant species.
Litterfall pattern
Another important aspect of the litterfall is the annual litterfall pattern. Both tree species were observed to shed their leaves once a year albeit at different times. Koompassia and Shorea are evergreen or semi-evergreen tropical moist forest tree species, which typically have long leaf life spans due to the low resource costs to support leaf life span and by producing leaves that have a longer duration of photosynthetic activity (Singh and Kushwaha 2005). The leaf life span of these types of tree is usually around 1 year (Singh and Kushwaha 2005), which supports the data obtained from this study. The chemistry of the leaf litter may also explain the single shedding event observed in this study. Leaves that have high amounts of secondary metabolites and have high tensile strength such as those of Koompassia and Shorea will usually have long leaf life spans as this reduces the amount of resources required for drought or herbivory tolerance. Long leaf life span is also a common characteristic for plants growing in water and N-stressed ecosystems such as the TPSF, which allows the plant to utilize N for an extended period of time (Campanella and Bertiller 2008). Furthermore, the leaf nutrient profile of both tree species also showed that they are both late successional tree species, which typically have long leaf life spans.
Interaction between litterfall and climate
When litterfall was tested for interactions with climatic variables, no interactions were observed except for a weak negative correlation between Shorea litterfall and rainfall. Litterfall events during the drier part of the year have been frequently observed and described in many studies especially in tropical regions as water stress during the dry season promotes litterfall (Zhang et al. 2014). This has previously been reported as a survival strategy of Shorea trees that shed their old leaves of different ages during the dry season to prevent water loss via transpiration (Singh and Kushwaha 2005). However, given that the TPSF is permanently waterlogged, the trees would never actually experience water stress during dry periods; instead the lowering of the water table would decrease the size of pools and provide an increase in area of the forest floor that is above the water table. Further studies (Ong in prep.) showed that litter decomposition in Sungai Karang PSF is faster in leaves exposed on hummocks than those submerged in pools. Consequently, increased litterfall in the dry season could be an adaptation to promote more rapid litter decomposition and remineralization of nutrients.
Despite a weak negative correlation between Shorea litterfall and rainfall, this is an indication that Shorea may be more sensitive to weather or climate changes as compared to Koompassia. With the increasing rainfall variability along with longer periods of drought as projected by climate studies, Shorea may increase its annual leaf litterfall in response to that, as it has been documented to shed its leaves during the dry period. However, more studies will be required to verify this. Aside from leaf litter, seed production may also be affected by climate change whereby germination and growth of certain seeds may be positively affected by dry periods or infrequent rainfall while others may not. In a study conducted by O’Brien et al. (2013), infrequent rainfall was found to positively affect germination of large seeds but negatively affect their growth while the opposite was observed for small seeds, which are also known to prefer growing in wetter areas. In general, large-seeded tree species tend to produce leaves with greater area and vice versa. In this study, Shorea produces larger leaves compared to Koompassia, which may suggest that its seeds have a greater mass as compared to the latter. Hence, higher seed production may be observed for Shorea during the dry season as compared to Koompassia. This can be another indication that Shorea is more affected by drought as compared to Koompassia, which is a good point to be considered for future studies on litterfall in TPSF.
Aside from dry periods, lower radiation paired with higher number of cloudy days during the rainy season also reduces tree photosynthetic rate and litterfall. In other tropical studies, mean minimum and maximum temperature as well as distribution of rainfall (number of rainy days) have been associated with litterfall (Williams-Linera and Tolome 1996; Liu et al. 2004; Chave et al. 2010) but similar interactions were not observed in this study due to the relatively aseasonal climate, with no seasonal changes in temperature.
The limited relationship observed between litterfall and climatic variables could also be due to the slightly unusual climatic pattern observed during the particular study period. The mean annual minimum and maximum rainfall events observed at the study site differed slightly from the mean annual pattern observed on the west coast of Peninsular Malaysia. Shorea litterfall occurred just before the normal dry period while Koompassia litterfall occurred during the typical, dry period. There is a possibility that the trees were adapted to the more typical annual rainfall pattern. Furthermore, recent studies span unusual or unstable climatic periods that resulted in drought in 2005 or extremely long rainy season such as the La Niña from 2008 to 2009 (Chave et al. 2010). Such data may not represent the long-term seasonality pattern and thus, this study may not be comparable to those datasets. Besides that, we should not disregard the fact that litterfall can also respond to variations in solar radiation (Wright and van Schaik 1994), rate of litter decomposition (Cueves and Medina 1988), stand age, soil fertility (Vitousek 1984), heavy winds or rain (Cuevas and Lugo 1998) as well as interactions between plant species as well as other organisms such as pollinators and herbivores in an ecosystem.
Conclusion
Koompassia malaccensis and Shorea uliginosa in Sungai Karang TPSF showed marked differences with respect to litterfall mass produced despite the homogeneous TPSF environment. They exhibited differences in terms of internal and external nutrient regulation whereby the former was suggested to rely on nutrient retranslocation to living tissues from senescing leaves as well as a higher litter quality despite low litterfall mass while the latter used higher litterfall mass to compensate for low litter quality and, was also suggested to depend more on uptake of nutrients from the soil. Both tree species were shown to have a single shed annually and little or no relationship was observed between litterfall and climatic variables due to the relatively aseasonal climate. The data obtained contribute to the knowledge of litterfall process in the tropics where it is poorly understood especially in the unique TPSF of Southeast Asia. A better understanding of this process is vital as it aids in the appropriate management of the rapidly vanishing peat swamp forests and the conservation of the selected emergent tree species which are heavily logged in the region.
Author contribution
The first author, Cheryl S. P. Ong, was involved in designing the research, acquiring research data, analysis as well as interpretation of data and drafting of the manuscript. Assoc. Prof. Yule CM and Assoc. Prof. Juan JC are the first author’s main supervisor and co-supervisor, respectively, who were involved in the study conception and design as well as the revision of the manuscript.
References
Aldhous P (2004) Land remediation: Borneo is burning. Nature 432:144–146. doi:10.1038/432144a
Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM (2003) Allelopathy and exotic plant invasion from molecules and genes to species interaction. Sci. 301:1377–1380
Biswas S, Khan D (2011) Major nutrient dynamics of two plant species at matha protected forest of Purulia, West Bengal, India. Int J Environ Sci 2(1):222–231
Bray JR, Gorham E (1964) Litter production in forests of the world. Adv Ecol Res 2:101–157
Campanella MV, Bertiller MB (2008) Plant phenology, leaf traits and leaf litterfall of contrasting life forms in the arid Patagonian Monte, Argentina. J Veg Sci 19:75–85
Chave J, Navarette D, Almeida S, Alvarez E, Aragao LEOC, Bonal D, Chatelet P, Silva-Espejo JE, Goret JY, von Hildebrand P, Jimenez E, Patino S, Penuela MC, Phillips OL, Stevenson P, Malhi Y (2010) Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 7:43–55
Cornelissen JHC, Thompson K (1997) Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol 135:109–114
Cuevas E, Lugo AE (1998) Dynamics of organic matter and nutrient return from litterfall in stands of ten tropical tree plantation species. For Ecol Manage 112(3):263–279
Cueves E, Medina E (1988) Nutrient dynamics within Amazonian forests II. Fine root growth, nutrient availability and leaf litter decomposition. Oecologia 76:222–235
Fierer N, Schimel JP, Cates RG, Zou J (2001) Influence of balsam poplar tannin fractions on carbon and nitrogen dynamics in Alaskan taiga floodplain soils. Soil Bio Biochem 33:1827–1839
Finzi AC, Allen AS, DeLucia EH, Ellsworth DS, Schlesinger WH (2001) Forest litter production, chemistry and decomposition following two years of free-air CO2 enrichment. Ecology 82(2):470–484
Graca MAS, Barlocher F, Gessner MO (eds) (2005) Methods to study litter decomposition: a practical guide. Springer, pp 115–120
Hahn-Schilling B (1994) Struktur, sukzessionale Entwicklung und Bewirkschaftung selektiv genuzter Moorwälder in Malaysia. Göttinger beiträge zur land- und forstwirtschaft in den tropen und subtropen Verlag Erich Goltze GmbH & Co, Gottingen
Hansen K, Vesterdal L, Schmidt IK, Gundersen P, Sevel L, Bastrup-Birk A, Pedersen LB, Bille-Hansen J (2009) Litterfall and nutrient return in five tree species in a common garden experiment. For Ecol Manag 257(10):2133–2144
Hattenschwiler S, Vitousek PM (2000) The role of polyphenols in terrestrial ecosystem nutrient cycling. Trees 15(6):238–243
Hättenschwiler S, Aesclimann B, Coûteaux M, Roy J, Bonal D (2008) High variation in foliage and leaf litter chemistry among 45 tree species of a neotropical rainforest community. New Phytol 179(1):165–175
Koerselma W, Meuleman AFM (1996) The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation. J Appl Ecol 33:1441–1450
Kraus ECT, Zasoski RJ, Dahlgren RA, Horwath WR, Preston CM (2004) Carbon and nitrogen dynamics in a forest soil amended with purified tannins from different plant species. Soil Biol Biochem 36:309–321
Kuiters AT (1990) Role of phenolic substances from decomposing forest litter in plant-soil interactions. Acta Bot Neerl 39:329–348
Lin P, Wang WQ (2001) Changes in the leaf composition, leaf mass and leaf area during leaf senescence in three species of mangroves. Ecol Eng 16(3):415–424
Lin YM, Peng ZQ, Lin P (2004) Dynamics of leaf mass, leaf area and element retranslocation efficiency during leaf senescence in Phyllostachys pubescens. Acta Bot Sin 46(11):1316–1323
Liu C, Westman CJ, Bergs B, Kutsch W, Wang GZ, Man R, Ilvesiemi H (2004) Variation in litterfall–climate relationships between coniferous and broadleaf forests in Eurasia. Global Ecol Biogeogr 13(2):105–114
Lowman MD (1988) Litterfall and leaf decay in three Australian rainforest transformations. J Ecol 76:451–465
Lugo AE, Cuevas E, Sanchez MJ (1990) Nutrients and mass in litter and top soil of 10 tropical tree plantations. Plant Soil 125:263–280
Meier CL, Bowman WD (2008) Phenolic-rich leaf carbon fractions differentially influence microbial respiration and plant growth. Oecologia 158:95–107
Milla V, Kuban V, Kledjus B, Osdtrcilova V, Nerusil P (2005) Phenolic compounds as chemical markers of low taxonomic levels in the family Poaceae. Plant Soil Environ 51:506–512
Miller HG (1984) Dynamics of nutrient cycling in plantation ecosystems. In: Bowen GD, Nambiar EKS (eds) Nutrition of plantation forests. Academic Press, London, pp 53–78
O’Brien MJ, Philipson CD, Tay J, Hector A (2013) The influence of variable rainfall frequency on germination and early growth of shade-tolerant dipterocarp seedlings in Borneo. PLoS ONE 8(7):e70287
Page SE, Siegert F, Rieley JO, Boehm H-DV, Jaya A, Limin SH (2002) The amount of carbon released from peat and forest Wres in Indonesia during 1997. Nature 420:61–65
Page SE, Rieley JO, Wüst R (2006) Lowland tropical peatlands of southeast Asia. In: Martini IP, Martinez Cartizas A, Chesworth W (eds) Peatlands: evolution and records of environmental and climate changes. Elsevier, The Netherlands, pp 145–172
Pahang Forestry Department (2005) Pekan peat swamp forest, Pahang. Malaysia. The role of water in conserving peat swamp forests. Pahang Forestry Department, UNDP/GEF, Danida
Proctor J (1983) Tropical forest litterfall. I. Problems of data comparison. In: Sutton SL, Whitemore TC, Chadwick AC (eds) Tropical rain forest: ecology and management. Blackwell Scientific Publications, Oxford, pp 267–273
Ricardo SS (1989) Nutrient concentration and retranslocation in coastal vegetation and mangroves from the Pacific Coast of Costa Rica. Brenesia 37:33–50
Riswan S (1989) Kerangas forest at Gunung Pasir, Samboja, East Kalimantan. II. Nutrient status in the leaves. Paper presented at the Fourth-Round Table Conference on Dipterocarps. Bogor, Indonesia
Schlesinger WH, DeLucia EH, Billings WD (1989) Nutrient-use efficiency of woody plants on contrasting soils in the Western Great Basin Nevada. Ecology 70(1):105–113
Singh KP, Kushwaha CP (2005) Paradox of leaf phenology: Shorea robusta is a semi-evergreen species in tropical dry deciduous forests in India. Curr Sci 88(11):1820–1824
Ushio M, Wagai R, Balser TC, Kitayama K (2008) Variations in the soil microbial community composition of a tropical montane forest ecosystem: does tree species matter? Soil Biol Biochem 40:2699–2702
Vitousek PM (1984) Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 65:285–298
Williams-linera AG, Tolome J (1996) Litterfall, temperate and tropical dominant trees, and climate in a Mexican lower montane forest. Biotropica 28(4):649–656
Wright SJ, van Schaik CP (1994) Light and the phenology of tropical trees. Am Nat 143:192–199
Wurzburger N, Hendrik RL (2007) Rhododendron thickets alter N cycling and soil extracellular enzyme activities in southern Appalachian hardwood forests. Pedogiologia 50:563–576
Yule CM (2010) Loss of biodiversity and ecosystem functioning in Indo-Malayan peat swamp forests. Biodiv Conserv 19:393–409
Yule CM, Gomez LN (2009) Leaf litter decomposition in a tropical peat swamp forest in Peninsular Malaysia. Wet Ecol Manage 17(13):231–241
Zhang H, Yuan W, Dong W, Liu S (2014) Seasonal patterns of litterfall in forest ecosystem worldwide. Complex, Ecol. doi:10.1016/j.ecocom.2014.01.003
Acknowledgments
This research was supported by and conducted in Monash University Malaysia Campus. The authors are also grateful to the Ministry of Education and University of Malaya for financial support (FP054-2013B and RP025A-14AET). Recognition is also given to Selangor Forestry Department who allowed us to access the study area at Sungai Karang, North Selangor tropical peat swamp forest. We would also like to thank Ms. Chou Lee Yiung from Monash University Malaysia for her help in the field as well as Mr. Jamil Omar and Ms. Nor Asma in the Department of Land Management, Universiti Putra Malaysia for their assistance with foliar total N and P processing and determination.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Hampp.
Rights and permissions
About this article

Cite this article
Ong, C.S.P., Juan, J.C. & Yule, C.M. Litterfall production and chemistry of Koompassia malaccensis and Shorea uliginosa in a tropical peat swamp forest: plant nutrient regulation and climate relationships. Trees 29, 527–537 (2015). https://doi.org/10.1007/s00468-014-1130-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-014-1130-y