
Abstract
Aims
Nitrogen (N) addition could affect litter decomposition through its direct effects on soil N availability and indirect effects on initial litter chemistry. The aim of this study was to evaluate the relative contribution of these direct and indirect effects to the decomposition of fine roots with different diameter classes.
Methods
A two-year reciprocal replant–transplant field experiment was conducted in a Mongolian pine (Pinus sylvestris var. mongolica) plantation to examine the relative effect of exogenous and endogenous N enrichment induced by N addition (10 g N m−2 yr.−1) on the decomposition of fine roots with different diameter classes: < 0.5 mm (small fine root, SFR) and 0.5–2 mm (large fine root, LFR).
Results
The LFR had significantly higher decomposition rates (k: 0.315–0.397 yr.−1) than the SFR (0.245–0.274 yr.−1) after 2 years of incubation. Exogenous N (i.e., increased soil N availability due to N addition) had no significant effect on the decomposition rates of fine roots, whereas endogenous N (i.e. increased N concentration in litter due to N addition) inhibited and accelerated the decomposition of SFR and LFR, respectively. Endogenous N decreased the net release of N but both endogenous and exogenous N increased the net release of phosphorus (P) from SFR. By contrast, exogenous and endogenous N decreased the net release of N and P from LFR.
Conclusions
Our results suggest that N addition affected fine root decomposition indirectly by changing the chemical traits of fine roots rather than directly through changing soil N availability. Elevated input and decreased net N release of fine roots might be a potential mechanism explaining the increases of total organic carbon and total N in the semi-arid forest soil under N addition. Our study also suggests that SFR may be a more important source of stable soil organic matter relative to LFR.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Plant litter decomposition is an important pathway in carbon (C) and nutrient cycling in terrestrial ecosystems (Attiwill and Adams 1993). Although the field of litter decomposition has made great strides, many earlier studies focus on leaf litter decomposition. As such, far less attention has been paid to fine root decomposition (Sun et al. 2016; Lin and Zeng 2017). A recent estimation has shown that approximately 22% of the terrestrial net primary production is allocated to fine roots (≤ 2 mm in diameter) (McCormack et al. 2015). Previous studies found that the fluxes of C and nitrogen (N) from the decomposition of fine roots may exceed those from aboveground litter (Nadelhoffer and Raich 1992; Tu et al. 2015). Soil nutrient cycling would be underestimated by 20–80% if the production and decomposition of fine roots were neglected (Ostonen et al. 2005). Therefore, studying fine root decomposition is necessary to fully understand ecosystem C and nutrient cycling (Lin and Zeng 2017).
Many biotic and abiotic factors have been suggested to affect root litter decomposition (Silver and Miya 2001; Zhang et al. 2008). Among these factors, N availability effects have gained great attention in the context of increased atmospheric N deposition at the global scale (Galloway et al. 2004; Denman et al. 2007). During the past 100 years, N input into terrestrial ecosystems has increased from 34 to 100 Tg N yr.−1, and it is predicted to increase to 200 Tg yr.−1 by 2050 (Galloway et al. 2008). Moreover, South and East Asia are likely to experience the greatest increase in the rate of N deposition (Liu et al. 2013). Nitrogen deposition can increase soil N availability, which could considerably affect decomposer activities and then directly influence the decomposition of fine roots (Fang et al. 2007). On the other hand, the initial chemistry of fine roots could be affected by N deposition (Kozovits et al. 2007), indirectly affecting root litter decomposition. The rationale behind this indirect effect may be that root chemistry is one of the main factors controlling root decomposition (Silver and Miya 2001; Zhang et al. 2008). However, few studies have focused on the indirect effect of endogenous N (i.e., effects of N addition on litter chemistry) on root litter decomposition under N deposition. Furthermore, the relative effect of endogenous and exogenous N (i.e., effects of N addition on soil N availability) on fine root decomposition is still unclear (Zheng et al. 2017b).
Apart from N availability, the diameter of fine roots may also be an important factor influencing fine root decomposition, considering that many traits related to root decomposition exhibit large variations among root diameter classes within the fine root branching hierarchy (Lin and Zeng 2017). For example, fine roots with larger diameters might have higher C and lignin concentrations, which could inhibit root decomposition (Silver and Miya 2001). In contrast, small-diameter fine roots typically have a lower C:N ratio and lack secondary development compared to large-diameter fine roots (Pregitzer et al. 2002; Guo et al. 2008), and thus these small-diameter fine roots are expected to have a higher decomposition rate (Sun et al. 2016). However, mounting evidence suggests that the fine root decomposition rates increase with increasing root diameter (Fan and Guo 2010; Xiong et al. 2013; Sun et al. 2013). One of the mechanisms responsible for the lower decomposition rate of small-diameter fine roots may be their higher concentrations of the acid-unhydrolyzable fraction than large-diameter fine roots (Xiong et al. 2013; Sun et al. 2013). An alternative explanation for the higher decomposition rate of large-diameter fine roots may be their higher concentration of non-structural carbohydrates. These easily degradable labile C substrates may provide necessary energy for the decomposer community, promoting the degradation of many complex C compounds in large-diameter fine roots (Hättenschwiler et al. 2011; Tamura and Tharayil 2014). In addition, previous studies found that N released in the first several years in the fine root decomposition was limited (Goebel et al. 2011; Xiong et al. 2013). Although the mechanisms underlying the differences in decomposition rates among fine roots of different diameter classes are well known, decomposition of these fine roots responding to N deposition is still poorly understood.
Arid and semi-arid land accounts for about one third of the global land area (Huang et al. 2015). Low soil N availability usually accompanies water limitation in arid and semi-arid ecosystems. The availability of soil N and other nutrients could be influenced by N deposition, thereby affecting the productivity of arid and semi-arid ecosystems (Brooks 2003). Studies on the effect of N deposition on the growth of roots and the decomposition of root litter have mainly been conducted in temperate and tropical forest ecosystems (McCormack et al. 2015; Lin and Zeng 2017). Mongolian pine (Pinus sylvestris var. mongolica), an important tree species for afforestation in semi-arid regions with sandy soils, has been planted since the mid-1950s to reduce wind erosion and to enhance sand-fixation in Keerqin sandy lands of northeast China (Mao et al. 2016). In those Mongolian pine plantations, soil nutrients, especially N, are often deficient. Considering the important role of root litter decomposition on the release of available nutrients into the soil, it is necessary to evaluate the influence of N deposition on the decomposition of and nutrient release from fine root litter in semi-arid regions of northeast China. In this study, we conducted a field decomposition experiment in a N-limited Mongolian pine plantation in semi-arid northeast China to test the following hypotheses: (1) N addition will enhance fine root decomposition through increasing soil N availability and reducing fine root C/N ratios; and (2) Small fine roots with <0.5 mm in diameter will have a slower decomposition rate than large fine roots with 0.5–2 mm in diameter. Results from this study will inform the ecological consequences of increased N deposition on belowground litter decomposition and related nutrient cycling in semi-arid regions.
Materials and methods
Site description
The study was conducted at Daqinggou Ecological Station (42°54’N, 122°21′E; 260 m. a.s.l.) of the Institute of Applied Ecology, Chinese Academy of Sciences, in the southeastern Keerqin sandy lands, northeast China. The study site has a dry sub-humid temperate monsoon climate. Mean annual precipitation is about 450 mm, with 70% of which falling from June to September. The mean annual temperature is 6.4 °C and the lowest and highest monthly mean air temperatures are −12.5 °C in January and 23.8 °C in July, respectively. The soil is a nutrient-poor sandy soil, with 90.9% sand, 5.0% silt, and 4.1% clay (Zeng et al. 2009).
Experimental design and sampling
In May 2011, eight 20 m × 30 m plots (four +N plots and four N0 plots) were randomly established in a 13-year old Mongolian pine plantation in the Daqinggou Ecological Station (Zheng et al. 2017a). There was a 5 m buffer zone between neighboring plots. The experimental plots were randomly allocated to two treatments: N0 (no N added) and +N (addition of 10 g N m−2 yr.−1). For the +N plots, urea was dissolved in water and spread monthly during the growing season (May to September, 2011–2016). 2 g N m−2 was spread every time, with a total of 10 g N m−2 yr.−1. The N0 plots received the same amount of water as the +N plots.
In April 2015, after 4 years of N addition, Mongolian pine fine roots (≤ 2 mm in diameter) were collected from the upper 50 cm mineral soil layer in all plots. All roots were washed with tap water and then air-dried at room temperature for 10 days. Fine roots are usually defined as ≤2 mm in diameter. A study reported that lateral roots <0.5 mm in diameter could account for 75% of the fine root biomass and root samples <0.5 mm in diameter were often used to determine mycorrhizal colonization (Pregitzer et al. 2002). Therefore, in this study, fine roots were operationally classified into <0.5 mm (small fine root, SFR) and 0.5–2 mm (large fine root, LFR) diameter classes. According to a study conducted in northeast China (Guo et al. 2008), fine roots <0.5 mm in diameter for Mongolian pine consist of first- to third-order roots, and fine roots of 0.5–2 mm in diameter consist of fourth- and fifth-order roots. Meanwhile, they found that first- and second-order roots as well as a portion of third-order roots lack secondary (wood) development, thus mainly representing absorptive roots, while fourth- and fifth-order roots have secondary xylem developed within the stele without mycorrhizal colonization, thus primarily serving functions such as transport, anchorage, and storage.
Living fine roots were used in our decomposition experiment because there is currently no perfect method to collect sufficient dead roots that naturally died, despite the fact that physiological and chemical characteristics of living roots may differ from those that naturally died (Eissenstat and Volder 2005; Resendes et al. 2008; Sun et al. 2016). Fine roots of the small and large diameter classes derived from the N0 and +N treatments are referred to as N0-SFR, N0-LFR, +N-SFR and +N-LFR, respectively. The air-dried fine roots from each plot were divided into two sub-samples. One sub-sample was used for analyzing water content (oven dried at 65 °C for 48 h) and chemical properties (C, N, P, and lignin concentrations). The other sub-sample was cut into pieces (about 1 cm in length) and was used for decomposition experiment.
After finishing the preparation of fine roots, we initiated the decomposition experiment of fine roots in a reciprocal replant–transplant design. In total, there were four types of fine roots (N0-SFR, N0-LFR, +N-SFR, +N-LFR). Air-dried roots (5 g) of each type were placed in litterbags (10 cm × 10 cm) made of a nylon mesh with 100-μm opening size. Litterbags were buried 15 cm beneath the soil surface in the eight plots in April of 2015. Thus, there were 48 litterbags (4 types of fine roots × 4 replicate samples × 3 sampling times) in each plot, with a total of 384 litterbags set up in this study. The litterbags were harvested in late July and October of 2015, and in October of 2016. Retrieved root samples were weighed after cleaning off soil and other debris and dried at 65 °C for 48 h.
Although it is common to assess decomposition of fine roots by removing roots from the soil, air drying, and returning samples to the soil after being confined in litterbags, this is not the case that roots decompose in situ (Dornbush et al. 2002) and the litterbag method does not provide the actual rate of root decomposition (Li et al. 2015). The decay rates in our study were relative rates and were used to assess treatment effects.
To better understand the effects of exogenous and endogenous N on fine root decomposition, we divided the decomposition experiment into three components: (1) Litterbags with N0-root were incubated in the N0 and +N plots. This allows us to examine the direct effect of N addition on fine root decomposition (i.e., exogenous N effect due to altered soil properties by N addition). (2) Litterbags with N0-root and +N-root were incubated in the N0 plots. This allows us to quantify the indirect effect of N addition on fine root decomposition (i.e., endogenous N effect due to altered litter chemistry by N addition). (3) Litterbags with N0-root were incubated in the N0 plots and those with +N-root were incubated in the +N plots. This allows us to ascertain the coupled direct and indirect effects of N addition on fine root decomposition (i.e., overall effect of N addition).
To characterize the soil environment of fine root decomposition, 30 surface (0–10 cm) mineral soil samples were collected from each plot using a soil corer (2.5 cm in diameter) in June 2015. The samples from each plot were mixed to form a composite sample. Soil samples were sieved with a 2-mm mesh and then air-dried for the further determination of soil pH, soil total C, total N, and total P. In October 2015, 5 years after the initiation of N addition, Mongolian pine fine root density was determined by collecting soil samples using a soil corer (6 cm in diameter) from the top 60 cm of the soil in all eight plots. Eight cores were randomly collected from each plot. Roots were washed with tap water, dried at 65 °C for 48 h and then weighed (Powers et al. 2005).
Chemical analysis of soil and fine roots
For chemical analyses, the oven-dried fine roots were ground to a fine powder using a ball mill (GT200, Beijing Grinder Instrument Co., Ltd., China). Ash content in each root sample was determined by combusting a 0.5 g subsample in a muffle furnace at 550 °C for 4 h. Total organic C (TOC) concentration in soil and fine root samples was determined using the K2Cr2O7–H2SO4 wet oxidation method (Nelson and Sommers 1996). After digesting with H2SO4, total N (TN) and P concentrations in soil and fine root samples were determined with a continuous-flow auto-analyzer (AutoAnalyzer III, Bran+Luebbe GmbH, Germany). Soil pH was measured with a pH meter (PHS-3C, Leici, China) in a 1:2 mixture (w:w) of soil and water.
Lignin concentration in fine root samples was analyzed using a modified acetyl bromide method (Morrison 1972a, b). Briefly, litter sample (6 mg) was placed into a 25 mL test tube with a solution of 25% acetyl bromide (AcBr) in acetic acid (5 mL) and 0.2 mL HClO4 (70%). The graduated tubes were incubated at 70 °C in a water bath for 30 min, and then 10 mL 2 mol L−1 NaOH was added. Finally, acetic acid was added to the 25 mL mark on the graduated tube. The acetic acid solution was then measured by UV spectrophotometry at 280 nm to determine the concentration of lignin, which was calculated as follows:
where Abs is the sample solution lignin absorbance at 280 nm, V is the constant volume of sample solution (25 mL), Wsample is the sample weight (6 mg), and Astandard is the specific absorbance coefficient (SAC) value employed for estimating lignin concentration (20 L g−1 cm−1) (Iiyama and Wallis 1990).
Data analysis
The percent remaining (R) of mass, lignin and nutrient elements (N and P) of root litter for each incubation period (Xi) was determined and compared to the initial values (X0) using the formula: %R = (Xi/X0) × 100. The mass remaining, as a proportion of the initial mass, was plotted against time, using a single-exponent decomposition model, X = e–kt (Olson 1963), where X is the fraction of mass remaining at time t (yr.), and k (yr.−1) is the decomposition constant. The relationship between lignin remaining and mass remaining was analyzed using linear regression. Fine root biomass (g m−2) was calculated by: ρ = M/S × 10, where M (mg) is the total mass of fine roots in the soil corer to a depth of 60 cm, and S (cm2) is the area of the soil corer. Data on mass remaining and decomposition rate of litter were presented on ash-free mass basis.
Repeated measures ANOVA (N addition and root type as main effects and time as a within subject factor) was used to test the remaining of mass, lignin and elements (N, P) among the different N addition treatments and root diameters. When Mauchly’s test of sphericity was not passed (the variances were not equal), the data was adjusted by the Greenhouse-Geisser method (Tu et al. 2015). To investigate N addition and root diameter effects on fine root decomposition over 2 years, k value, mass remaining, lignin remaining and elements (N, P) remaining were compared among root diameter classes and N addition treatments using a two-way ANOVA. One-way ANOVA was used to test the differences of the initial chemical parameters among different root types. Correlation analysis was used to determine the relationship between the initial fine root chemistry and k value and mean value of N remaining during the whole decomposition period. A Fisher’s LSD test was used to test for significant differences among treatments with respect to root biomass, soil pH, soil TOC, soil TN, and soil C/N ratio. All statistical analyses were conducted using SPSS 18.0 (SPSS Inc. Chicago, USA). All statistical analyses were set at α = 0.05.
Results
Effects of N addition on root chemistry
Initial chemical traits of fine root litter were affected by N addition (Table 1). Nitrogen addition significantly increased fine root N concentrations by 16.3% in SFR and 14.7% in LFR, but reduced P concentrations by 25.5% in SFR and 44.0% in LFR. Nitrogen addition decreased fine root C:N ratio by 14.5% in SFR and 19.1% in LFR. Nitrogen addition had no effect on C and lignin concentrations in SFR but decreased them by 7.6% and 11.9% in LFR, respectively. Compared with SFR, LFR had significantly lower N but higher C, lignin concentration and C:N ratio (Table 1) under no N addition.
Mass remaining and decomposition constant k value of root litter
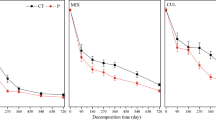
Fine roots lost 38.7–53.6% of their initial mass after 2 years of incubation. The SFR collected from the +N plots had significantly lower k-values (0.245 yr.−1) than that collected from the N0 plots (0.274 yr.−1) (Table 2). After 2 years of incubation, exogenous N had no impact on the mass remaining of SFR (Table 2, Fig. 1a), but endogenous N reduced the decomposition of SFR (p < 0.05) (Table 2, Fig. 1c), with no overall effect of N addition on the decomposition of SFR (Table 2, Fig. 1e).

Percent mass remaining of fine root over the decomposition period. Small fine root (SFR) (a) and large fine root (LFR) (b) were collected from the N0 plots and incubated in the N0 and +N plots. SFR (c) and LFR (d) were collected from the N0 and +N plots but incubated in the N0 plots. e, f In situ decomposition of fine roots under their respective fields. Error bars are means ± SE. Significant differences between treatments are indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
The LFR collected from the +N plots had significantly higher k-values (0.320–0.397 yr.−1) than that collected from the N0 plots (0.315 yr.−1) (Table 2). Exogenous N had no influence on the decomposition of LFR (Fig. 1b). Endogenous N inhibited the decomposition of LFR in the first year, but accelerated it after 2 years of incubation (Fig. 1d). Nitrogen addition showed a decreased overall effect on the decomposition rate of LFR in the first year but had no effect on it after 2 years of incubation (Table 2, Fig. 1f). The LFR had significantly higher k-values (0.315–0.397 yr.−1) than SFR (0.245–0.274 yr.−1) regardless of N addition (p < 0.05, Table 2).
Dynamics of lignin and C remaining during root decomposition
The dynamics of lignin remaining in the decomposing root litter was similar to that of mass remaining. For SFR, exogenous N increased lignin decomposition at half a year and then inhibited it (Fig. 2a). Endogenous N increased lignin decomposition in the first year and had no effect on the lignin decomposition rate in SFR after 2 years (Fig. 2c and e; Fig. S1). Similarly, N addition showed a stimulated overall effect on lignin decomposition in SFR in the first year but no such effect after 2 years. For LFR, N addition decreased lignin decomposition rate in the first year (Fig. 2b, d and f). After 2 years of incubation, endogenous N significantly increased LFR lignin decomposition but exogenous N had no influence on LFR lignin decomposition, with no overall effect of N addition on LFR lignin decomposition (Fig. S1).

Percent lignin remaining in fine roots over the decomposition period. Small fine root (SFR) (a) and large fine root (LFR) (b) were collected from the N0 plots and incubated in the N0 and +N plots. SFR (c) and LFR (d) were collected from the N0 and +N plots but incubated in the N0 plots. e, f In situ decomposition of fine roots under their respective fields. Error bars are means ± SE. Significant differences between treatments are indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
Residual C in all treatments decreased with time, similar to the patterns of mass remaining (Fig. 3). For SFR, N addition increased C decomposition rate in the first 6 months of incubation (Fig. 3a, c and e). After 2 years of incubation, N addition had no influence on C decomposition of SFR (Fig. S1). For LFR, exogenous N had no influence on C decomposition (Fig. 3b). Endogenous N inhibited C decomposition rate in LFR in the first year of incubation and then had a similar decomposition rate to LFR collected from the N0-plots after 2 years of incubation (Fig. 3d). An inhibited overall effect of N addition on C decomposition in LFR was found through the 2 years of incubation (Fig. 3f). The LFR had a faster C decomposition rate (53%–58%) than the SFR (46%–51%) after 2 years of incubation (Fig. S1).

Percent C remaining in fine roots over the decomposition period. Small fine root (SFR) (a) and large fine root (LFR) (b) were collected from the N0 plots and incubated in the N0 and +N plots. SFR (c) and LFR (d) were collected from the N0 and +N plots but incubated in the N0 plots. e, f In situ decomposition of fine roots under their respective fields. Error bars are means ± SE. Significant differences between treatments are indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
Dynamics of N and P remaining during root decomposition
Total N and P remaining exhibited different temporal patterns for different root diameter groups. Two distinct patterns were observed. First, the N remaining had similar temporal patterns in all treatments (Fig. 4). Endogenous and exogenous N inhibited N release (hereinafter representing a net process of uptake and release) from litter throughout the two-year study. The N release rate from LFR was 2–3 times higher than that from SFR. For SFR, exogenous N had no effect on N release (Fig. 4a). Endogenous N decreased the N release rate (14.5%) in SFR as compared with the control (17.5%) (Fig. 4c). Similarly, a decreased overall effect of N addition on N release rate was observed (12.7%, Fig. 4e). For LFR, the N release pattern was characterized by a rapid decrease followed by a gradual decrease (Fig. 4b, d and f), with more than 40% of the N released during the first 6 months of incubation. Subsequently, the N release rate slowed down throughout the rest of the decomposition period. Exogenous N decreased the N release rate (42.6%) as compared to the control (49.4%) (Fig. 4b). Similarly, a decreased overall effect of N addition on the N release rate (44.0%) was observed (Fig. 4f). Endogenous N had no influence on N release from LFR (Fig. 4d).

Percent N remaining in fine roots over the decomposition period. Small fine root (SFR) (a) and large fine root (LFR) (b) were collected from the N0 plots and incubated in the N0 and +N plots. SFR (c) and LFR (d) were collected from the N0 and +N plots but incubated in the N0 plots. e, f In situ decomposition of fine roots under their respective fields. Error bars are means ± SE. Significant differences between treatments are indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
Second, P remaining in fine roots decreased rapidly in the first year of incubation, and then decreased slowly (Fig. 5). All the N addition treatments increased the P release rate of SFR (by 23.0–33.7%) (Fig. 5a, c and e). Exogenous N had no influence on P release of LFR through the 2 years of incubation (Fig. 5b). Endogenous N decreased the P release of LFR (by 23.5%) (Fig. 5d). A decreased overall effect of N addition on the P release of LFR (by 13.2%) was observed (Fig. 5f). The LFR had a faster P release rate (60.0–73.5%) as compared to the SFR (47.5–63.5%) (Fig. S1).

Percent P remaining in fine roots over the decomposition period. Small fine root (SFR) (a) and large fine root (LFR) (b) were collected from the N0 plots and incubated in the N0 and +N plots. SFR (c) and LFR (d) were collected from the N0 and +N plots but incubated in the N0 plots. e, f In situ decomposition of fine roots under their respective fields. Error bars are means ± SE. Significant differences between treatments are indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
Root biomass and soil properties under N addition
Five years of N addition slightly increased fine root biomass by 5.2% (Fig. S2A, p > 0.05) but significantly decreased soil pH (Fig. S2B). Nitrogen addition significantly increased soil TOC and TN concentration (Fig. S2C, D, p < 0.05), but had no effect on soil total P concentrations and C:N ratio (Fig. S2E, F).
Discussion
Effects of N addition on decomposition of fine root of different diameter classes
During the two-year decomposition study, increased soil N availability (direct effect) and altered initial chemistry induced by N addition (indirect effect) inhibited SFR but increased LFR decomposition. For SFR, N addition stimulated its decomposition rate in the early stage, but ultimately lowered its decomposition rate. A similar pattern was observed in fine root decomposition in subtropical and temperate forests (Berg and Matzner 1997; Yang et al. 2004; Sun et al. 2016). For LFR, N addition inhibited its decomposition rate in the early stage, but ultimately increased its decomposition rate at the end of the experiment. In our study, two evident mass loss stages were observed (Fig. 1). In the first 6 months of decomposition, fine root mass loss ranged from 27% to 39%, while a very slow decomposition rate was observed in the following period. A similar root decomposition pattern was also observed in a subtropical bamboo forest (Tu et al. 2015). After 1 year of decomposition, the lignin remaining in fine roots was approximately 41.0–55.9% in all treatments. Furthermore, there was a significant positive linear relationship between lignin remaining and mass remaining across root types and N treatments (r = 0.62, p < 0.001; Fig. S3), which suggests that lignin may have played an important role in affecting the fine root mass loss. Many previous studies have found that N addition could inhibit lignin decomposition (Fog 1988; Berg and Matzner 1997; Magill and Aber 1998), and finally decrease the litter decomposition rate (Carreiro et al. 2000; Hobbie 2008). Compared with LFR, SFR had lower lignin concentration in N0 plots, suggesting that N addition-induced changes in lignin concentrations are likely to be the main reason responsible for increased SFR decomposition and decreased LFR decomposition in the first year (Knorr et al. 2005).
At the same time, N addition could reduce enzyme activities and increase the initial lignin concentration to inhibit litter decomposition (Sinsabaugh et al. 2005, 2008; Zak et al. 2008; Sinsabaugh 2010). The following mechanisms may be involved in the inhibitory effect of exogenous N on lignin decay. First, N addition decreased soil pH and then affected the activities of lignin peroxidase, manganese peroxidase and laccase, which are the three most important lignin-decaying enzymes (Janusz et al. 2013). White-rot (Basidiomycetes) and soft-rot microorganisms (Ascomycetes) are the main fungi that are related to lignin decay. The best pH range for the production of Basidiomycetes and Ascomycetes are 4.0–5.0 and 6.0–7.5, respectively (Sinsabaugh 2010). After 4 years of N addition, the pH of surface soil decreased from 6.48 (N0 plots) to 5.51 (+N plots) in this study. Therefore, Ascomycetes might be the main lignin-decaying fungi in the semi-arid ecosystem. Decreased soil pH induced by N addition may affect ligninolytic enzyme activities and then inhibit lignin degradation (Waldrop et al. 2004; Tu et al. 2014). Indeed, many studies have found that the inhibitory effects of high N environments on ligninolytic enzyme production are widespread (Berg and Matzner 1997; Carreiro et al. 2000; Waldrop et al. 2004; Tu et al. 2014, 2015). Second, exogenous N can react with the products from lignin degradation to form other recalcitrant compounds which are difficult to be degraded by microorganisms (Berg 1986). Our study found that SFR had a lower lignin decomposition rate in the late decomposition stage (year 2). Excessive exogenous and endogenous N in fine roots reacted with products of lignin and polyphenols degradation to form polymeric substances, which are more difficult to decompose (Berg and Matzner 1997). Therefore, N addition inhibited the decomposition of SFR in the 2 years of decomposition.
Long-term N deposition could increase the concentrations of N in the fine roots, which could affect fine root decomposition rate (Xia et al. 2018). In our study, N addition decreased initial lignin and C concentration of LFR. N concentrations in the fine roots in +N plots were higher than those in N0 plots (Table 1). A similar phenomenon was also found in a long-term N deposition experiment in the Harvard Forest in the United States (Micks et al. 2004). After 1 year of decomposition, N addition increased LFR decomposition due to the lower initial lignin and C concentrations in our study.
Effects of root diameter on fine root decomposition
In our study, the decomposition rate of SFR was lower than that of LFR, supporting our second hypothesis and was consistent with other studies (Xiong et al. 2013; Sun et al. 2013). Some studies found that higher concentrations of the acid-unhydrolyzable fraction (AUF) in SFR may be the main reason for its lower decomposition rate (Sun et al. 2016). Compared with the SFR, the LFR has higher non-structural carbohydrate concentrations (McCormack et al. 2015; Sun et al. 2016). In the early decomposition stage, the root mass loss was mainly derived from the leaching and fragmentation of root tissues (Tu et al. 2014). These easily degradable labile C substrates may provide necessary energy for the decomposer community, promoting the degradation of complex C compounds in LFR. Therefore, LFR with higher concentrations of non-structural carbohydrates has a faster decomposition rate compared to SFR with higher concentrations of recalcitrant compounds during the 2 years of decomposition. Furthermore, recent studies suggest that the labile components of root litter are the main precursor of mineral-associated organic matter (Cotrufo et al. 2013; Haddix et al. 2016). Nevertheless, given that the lower order fine roots (first- and second-order roots) are the dominant contributors to root mortality in various tree species due to their larger biomass and faster turnover (Pregitzer et al. 2002; Fan and Guo 2010), SFR is the main source of stable soil organic matter (Fan and Guo 2010; Kramer et al. 2010; Prescott 2010).
Nitrogen release patterns
Similar to the mass loss, N released more slowly from SFR than from LFR. Nitrogen release occurred mainly in LFR during the 2 years of decomposition, in contrast with other studies (Goebel et al. 2011; Sun et al. 2013). There was a significantly negative correlation between N remaining and initial C concentration (Table 3). The LFR with initial C:N ratios >50 generally released N consistently (Fig. 3). For SFR, there are three phases of N dynamics over the two-year period of decomposition under N addition: initial leaching, immobilization and release. One possible reason may be that exogenous N reacted with intermediates (such as polyphenols) to form anti-decomposing substances in fine roots, leading to more N accumulation in SFR (Sun et al. 2013). Another reason is that fine roots of different diameter classes are decomposed by different microbial communities (Hobbie and Horton 2007). Additionally, our result showed that SFR has a lower rate of N release (13–24%) during the 2 years of decomposition, which is in agreement with some other studies (Xiong et al. 2013; Sun et al. 2013). Parton et al. (2007) and Sun et al. (2013) showed that the majority of root N was released in the fourth year of decomposition. The major reason is that a large part of N in fine roots may interact with recalcitrant C component (i.e., AUF) and thus release of recalcitrant N compound occurs only when these recalcitrant C components are utilized by decomposer communities (Xiong et al. 2013).
After 5 years of N addition, fine root biomass and soil TOC in the Mongolian pine plantation increased, consistent with observations in some other forest ecosystems (King et al. 2002; Cleveland and Townsend 2006; Tu et al. 2015). Fine root and aboveground litter decomposition are main sources of soil organic carbon (Van Groenigen et al. 2006; Xiong et al. 2013). Thus, all the changes associated with fine root production and decomposition rate may impact the C pool of forest soil.
Conclusions
Nitrogen addition affected the decomposition rate of fine roots over 2 years of incubation, indirectly by changing the chemical traits of fine roots rather than directly through changing soil N availability. Our study provided evidence that litter quality rather than environmental factors (soil N availability), though both were altered by N addition, is an important factor influencing fine root decomposition rate. The lower decomposition rate of SFR than LFR is likely linked to the higher recalcitrant C fraction in the SFR than in the LFR. We also conclude that increasing atmosphere N deposition may increase soil TN and TOC. However, the effect of N deposition on microbial composition and enzyme activity and their linkage with fine root decomposition in such a semi-arid area should be further studied.
References
Attiwill PM, Adams MA (1993) Nutrient cycling in forests. New Phytol 124:561–582
Berg B (1986) Nutrient release from litter and humans in coniferous forest soils: a mini review. Scand J For Res 1:359–369
Berg B, Matzner E (1997) Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environ Rev 5:1–25
Brooks ML (2003) Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. J Appl Ecol 40:344–353
Carreiro MM, Sinsabaugh RL, Report DA, Parkhurst DF (2000) Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 81:2359–2365
Cleveland CC, Townsend AR (2006) Nutrient additions to tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proc Natl Acad Sci U S A 103:10316–10321
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2013) The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Chang Biol 19:988–995
Denman KL, Brasseur G, Chidthaisong A, Ciais P, Cox PM, Dickinson RE, Hauglustaine D, Heinze C, Holland E, Jacob D, Lohmann U, Ramachandran S, da Silva Dias PL, Wofsy SC, Zhang XY (2007) Couplings between changes in the climate system and biogeochemistry. Cambridge University Press, pp 499–587
Dornbush ME, Isenhart TM, Raich JW (2002) Quantifying fine-root decomposition: an alternative to buried litterbags. Ecology 83:2985–2990
Eissenstat DM, Volder A (2005) The efficiency of nutrient acquisition over the life of a root. Nutrient acquisition by plants: an ecological perspective. Ecol Stud 181:185–220
Fan PP, Guo DL (2010) Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soil. Oecologia 163:509–515
Fang H, Mo J, Peng S, Li Z, Wang H (2007) Cumulative effects of nitrogen additions on litter decomposition in three tropical forests in southern China. Plant Soil 297:233–242
Fog K (1988) The effect of added nitrogen on the rate of decomposition of organic matter. Biol Rev 63:433–462
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland CC, Green PA, Holland EA, Karl DM, Michaels AF, Porter JH, Townsend AR, Vorosmarty CJ (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Goebel M, Hobbie SE, Bulaj B, Zadworny M, Archibald DD, Oleksyn J, Reich PB, Eissenstat DM (2011) Decomposition of the finest root branching orders: linking belowground dynamics to fine-root function and structure. Ecol Monogr 81(1):89–102
Guo DL, Xiao MX, Wei X, Chang WJ, Liu Y, Wang ZQ (2008) Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol 180:673–683
Haddix M, Paul E, Cotrofo MF (2016) Dual, differential isotope labeling shows the preferential movement of labile plant constituents into mineral-bonded soil organic matter. Glob Chang Biol 22:2301–2312
Hättenschwiler S, Coq S, Barantal S, Handa IT (2011) Leaf traits and decomposition in tropical rainforests: revisiting some commonly held views and towards a new hypothesis. New Phytol 189:950–965
Hobbie SE (2008) Nitrogen effects on litter decomposition: a five-year experiment in eight temperate grassland and forest sites. Ecology 89:2633–2644
Hobbie SE, Horton TR (2007) Evidence that saprotrophic fungi mobilise carbon and mycorrhizal fungi mobilise nitrogen during litter decomposition. New Phytol 173:447–449
Huang JP, Yu HP, Guan XD, Wang GY, Guo RX (2015) Accelerated dryland expansion under climate change. Nat Clim Chang 6:166–172
Iiyama K, Wallis AFA (1990) Determination of lignin in herbaceous plants by an improved acetyl bromide procedure. J Sci Food Agric 51:145–161
Janusz G, Kucharzyk KH, Pawlik A, Staszczak M, Paszczynskic AJ (2013) Fungal laccase, manganese peroxidase and lignin peroxidase: gene expression and regulation. Enzym Microb Technol 52:1–12
King JS, Albaugh TJ, Allen HL, Buford M, Strain BR, Dougherty P (2002) Below-ground carbon input to soil is controlled by nutrient availability and fine root dynamics in loblolly pine. New Phytol 154:389–398
Knorr M, Frey SD, Curtis PS (2005) Nitrogen additions and litter decomposition: a meta-analysis. Ecology 86:3252–3257
Kozovits AR, Bustamante MMC, Garofalo CR, Bucci S, Franco AC, Goldstein G, Meinzer FC (2007) Nutrient resorption and patterns of litter production and decomposition in a Neotropical savanna. Funct Ecol 21:1034–1043
Kramer C, Trumbores S, Froberg M, Cisneros Dozal LM, Zhang D, Xu X, Santos GM, Hanson PJ (2010) Recent (<4 year old) leaf litter is not a major source of microbial carbon in a temperate forest mineral soil. Soil Biol Biochem 42:1028–1037
Li A, Fahey TJ, Pawlowska TE, Fisk MC, Burtis J (2015) Fine root decomposition, nutrient mobilization and fungal communities in a pine forest ecosystem. Soil Biol Biochem 83:76–83
Lin GG, Zeng DH (2017) Heterogeneity in decomposition rates and annual litter inputs within fine-root architecture of tree species: implications for forest soil carbon accumulation. For Ecol Manag 389:386–394
Liu XJ, Zhang Y, Han WX, Tang AH, Shen JL, Cui ZL, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang FS (2013) Enhanced nitrogen deposition over China. Nature 494:459–462
Magill AH, Aber JD (1998) Long-term effects of experimental nitrogen additons on foliar litter decay and humans formation in forest ecosystems. Plant Soil 203:301–311
Mao B, Mao R, Hu YL, Huang Y, Zeng DH (2016) Decomposition of Mongolian pine litter in the presence of understory species in semi-arid Northeast China. J For Res 27:329–337
McCormack ML, Dickie IA, Eissenstat DM, Fahey TJ, Fernandez CW, Guo DL, Helmisaari H-S, Hobbie EA, Iversen CM, Jackson RB, Leppälammi-Kujansuu J, Norby RJ, Phillips RP, Pregitzer KS, Pritchard SG, Rewald B, Zadworny M (2015) Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol 207:505–518
Micks P, Aber JD, Boone RD, Davidson EA (2004) Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. For Ecol Manag 196:57–70
Morrison IM (1972a) A semi-micro method for the determination of lignin and its use in predicting the digestibility of forage crops. J Sci Food Agric 23:455–463
Morrison IM (1972b) Improvements in the acetyl bromide technique to determine lignin and digestibility and its application to legumes. J Sci Food Agric 23:1463–1469
Nadelhoffer KJ, Raich JW (1992) Fine root production estimates and belowground carbon allocation in forest ecosystems. Ecology 73:1139–1147
Nelson DW, Sommers LE (1996) Total carbon, organic carbon and organic matter. Part 3: chemical methods. Wisconsin: Soil Science Society of America and American Society of Agronomy, pp 961–1010
Olson JS (1963) Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44(2):322–331
Ostonen I, Lõhmus K, Pajuste K (2005) Fine root biomass, production and its proportion of NPP in a fertile middle-aged Norway spruce forest: comparison of soil core and ingrowth core methods. For Ecol Manag 212:264–277
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Carol Adair E, Brandt LA, Hart SC, Fasth B (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–364
Powers JS, Treseder KK, Lerdau MT (2005) Fine roots, arbuscular mycorrhizal hyphae and soil nutrients in four neotropical rain forests: patterns across large geographic distances. New Phytol 165:913–921
Pregitzer KS, DeForest JL, Burton AJ, Allen MF, Ruess RW, Hendrick RL (2002) Fine root architecture of nine north American trees. Ecol Monogr 72:293–309
Prescott CE (2010) Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101:133–149
Resendes ML, Bryla DR, Eissenstat DM (2008) Early events in the life of apple roots: variation in root growth rate is linked to mycorrhizal and nonmycorrhizal fungal colonization. Plant Soil 313:175–186
Silver WL, Miya RKS (2001) Global patterns in root decomposition: comparisons of climate and litter quality effects. Oecologia 129:407–419
Sinsabaugh RL (2010) Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol Biochem 42:391–404
Sinsabaugh RL, Gallo ME, Lauber C, Waldrop MP, Zak DR (2005) Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 75:201–215
Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD, Crenshaw C, Contosta AR, Cusack D, Frey S, Gallo ME, Gartner TB, Hobbie SE, Holland K, Keeler BL, Powers JS, Stursova M, Takacs-Vesbach C, Waldrop MP, Wallenstein MD, Zak DR, Zeglin LH (2008) Stoichiometry of soil enzyme activity at global scale. Ecol Lett 11:1252–1264
Sun T, Mao Z, Han Y (2013) Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in four temperate tree species. Plant Soil 372:445–458
Sun T, Dong LL, Wang ZW, Lü XT, Mao ZJ (2016) Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperature forests. Soil Biol Biochem 93:50–59
Tamura M, Tharayil N (2014) Plant litter chemistry and microbial priming regulate the accrual, composition and stability of soil carbon in invaded ecosystem. New Phytol 203:110–124
Tu LH, Hu HL, Chen G, Peng Y, Xiao YL, Hu TX, Zhang J, Li XW, Liu L, Tang Y (2014) Nitrogen addition significantly affects forest litter decomposition under high levels of ambient nitrogen deposition. PLoS One 9:e88752
Tu LH, Peng Y, Chen G, Hu HL, Xiao YL, Hu TX, Liu L, Yi T (2015) Direct and indirect effects of nitrogen additions on fine root decomposition in a subtropical bamboo forest. Plant Soil 389:273–288
Van Groenigen KJ, Six J, Hungate BA, de Graadd MA, van Breemen N, van Kessel C (2006) Element interactions limit soil carbon storage. Proc Natl Acad Sci U S A 103:6571–6574
Waldrop MP, Zak DR, Sinsabaugh RL, Gallo M, Lauber C (2004) Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol Appl 14:1172–1177
Xia MX, Tdhelm AF, Pregitzer KS (2018) Long-term simulated atmospheric nitrogen deposition alters leaf and fine root decomposition. Ecosystems 21:1–14
Xiong YM, Fan PP, Fu SL, Zeng H, Guo DL (2013) Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant Soil 363:19–31
Yang YS, Chen GS, Guo JF, Lin P (2004) Decomposition dynamic of fine roots in a mixed forest of Cunninghamia lanceolata and Tsoongiodendron odorum in mid-subtropics. Ann For Sci 61:65–72
Zak DR, Holmes WE, Burton AJ, Pregitzer KS, Talhelm AF (2008) Simulated atmospheric NO3 − deposition increases soil organic matter by slowing decomposition. Ecol Appl 18:2016–2027
Zeng DH, Hu YL, Chang SX, Fan ZP (2009) Land cover change effects on soil chemical and biological properties after planting Mongolian pine (Pinus sylvestris var. mongolica) in sandy lands in Keerqin, Northeast China. Plant Soil 317:121–133
Zhang DQ, Hui DF, Luo YQ, Zhou GY (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93
Zheng LL, Zhao Q, Yu ZY, Zhao SY, Zeng DH (2017a) Altered leaf functional traits by nitrogen addition in a nutrient-poor pine plantation: a consequence of decreased phosphorus availability. Sci Rep 7:7415
Zheng ZM, Mamuti M, Liu HM, Shu YQ, Hu SJ, Wang XH, Li BB, Lin L, Li X (2017b) Effects of nutrient additions on litter decomposition regulated by phosphorus-induced changes in litter chemistry in a subtropical forest, China. For Ecol Manag 400:123–128
Acknowledgments
This study was funded by the National Natural Science Foundation of China (31870603, 41877341). We thank Gui-Yan Ai for help with the laboratory work and the CAS Key Laboratory of Forest Ecology and Management for funding Qun Gang’s visit to Canada. We also thank three anonymous reviewers for their comments and suggestions that greatly improved an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Hans Lambers.
Electronic supplementary material
ESM 1
(DOC 712 kb)
Rights and permissions
About this article

Cite this article
Gang, Q., Chang, S.X., Lin, G. et al. Exogenous and endogenous nitrogen differentially affect the decomposition of fine roots of different diameter classes of Mongolian pine in semi-arid northeast China. Plant Soil 436, 109–122 (2019). https://doi.org/10.1007/s11104-018-03910-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-03910-0