
Abstract
The nutrient concentrations and carbon:nitrogen:phosphorus stoichiometries of plants have been greatly altered by elevated CO2 concentrations and nitrogen (N) deposition. Studies of these changes, however, are mostly limited to tropical and subtropical forests. In this study, a C4 herbaceous grass (Bothriochloa ischaemum) was grown in a pot experiment at two CO2 concentrations (400 and 800 μmol mol−1) and three levels of N fertilization (0, 2.5 and 5 g N m−2 y−1) in a full factorial design. Plant biomass and the concentrations of non-structural carbohydrates (NSCs), N and phosphorus (P) in the shoots and roots were determined. The elevated CO2 concentration and N addition significantly increased total biomass, starch and NSC concentrations and root:shoot, starch:N, NSC:N, starch:P and NSC:P ratios in whole plants and roots. N addition alone decreased the soluble sugar (SS) concentration in whole plants and increased N concentration and decreased P concentration in whole plants and roots and thus decreased the SS:N ratio and increased the SS:P and N:P ratios. Our results suggested that SS, N and P concentrations and their stoichiometries responded more strongly to N addition than elevated CO2 concentration. Plant growth in this region suffers more from P than N limitation, and N addition would exacerbate the P limitation on plant growth.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Atmospheric CO2 concentrations have been increasing dramatically due to anthropogenic activities, from approximately 280 μmol mol−1 to the current level of 400 μmol mol−1, and are projected to exceed 800 μmol mol−1 by 2100 (NOAA 2013). Atmospheric nitrogen (N) deposition has also increased, by three- to five-fold within the last century, and is expected to continue to rise (Galloway et al. 2008). Elevated CO2 levels and N deposition have affected global biogeochemical cycles, particularly the carbon (C), N and phosphorus (P) cycles in terrestrial ecosystems (Heimann and Reichstein 2008; Gruber and Galloway 2008; Marklein and Houlton 2012). The altered ecosystemic nutrient cycles may lead to shifts in C:N:P stoichiometry in plant-soil systems (Yang et al. 2011; Huang et al. 2015a). The stoichiometric flexibility of elemental ratios, in return, may influence biogeochemical cycles by their effects on the material cycle in natural ecological systems (Huang et al. 2015b; Liu et al. 2015). Obtaining sufficient concentrations of nutrient elements and maintaining relatively stable stoichiometries in plant tissues is vital for health (Han et al. 2011). Our knowledge of the relationship between the global environmental changes and ecosystem stoichiometric responses is very important for understanding future biogeochemical cycles in terrestrial biosphere (Sardans et al. 2012; Huang et al. 2012; Liu et al. 2013a).
Increased atmospheric CO2 concentration stimulates photosynthesis and plant biomass, which are affected by most environmental stresses, such as nutrient limitation and water deficits (Newingham et al. 2013; Dong et al. 2016; Xiao et al. 2016). The sustainability of ecosystemic responses to CO2 maybe constrained by the progressive N limitation induced by the stimulation of growth from increased CO2 levels (Luo et al. 2004; Reich et al. 2006). The interaction of elevated CO2 levels and N addition would therefore synergistically increase plant biomass (Lee et al. 2010; Zhang et al. 2011; Novriyanti et al. 2012; Dong et al. 2016). A meta-analysis also showed that both grassland above- and belowground biomasses increased with a combination of CO2 elevation and N fertilization (Sillen and Dieleman 2012). Plant C:N and C:P ratios usually increase due to the dilution of N and P concentrations by the accumulation of carbohydrates (Novotny et al. 2007; Stiling and Cornelissen 2007). Elevated CO2 levels and N deposition can also alter the structures of soil microbial communities (Lee et al. 2015) and significantly increase soil respiration and nutrient (e.g. N and P) loss from decomposing litter (Deng et al. 2013; Liu et al. 2015). The availability of soil N and P can thus be increased under elevated CO2 levels in combination with N addition (Huang et al. 2014), thereby increasing plant N and P concentrations (Huang et al. 2015a). Plant N:P ratios, however, maybe not affected (Huang et al. 2015a), may increase (Huang et al. 2012) or may decrease (Liu et al. 2013a), depending on species (Liu et al. 2013a) and soil nutrient conditions (Huang et al. 2012, 2015a). Most studies until recently have focused on elemental-nutrient stoichiometries on the leaf-level rather than the level of whole plants (Liu et al. 2012; Huang et al. 2012). Mineral elements can respond to elevated CO2 levels and N addition more strongly in roots than leaves (Liu et al. 2013a; Huang et al. 2015c), and leaf-level responses may not be representative of whole-plant responses. The responses of different plant tissues to elevated CO2 levels and N addition, however, remain largely unexplored.
Carbohydrates are the main products of plant photosynthesis and are be partitioned mostly into structural carbohydrates (SCs) and non-structural carbohydrates (NSCs), based on their roles (Song et al. 2016). SCs, including lignin, cellulose, hemicelluloses and pectin, are mainly used for the structural growth of plants; NSCs, mainly soluble sugar (SS) and starch, provide substrates for plant growth and metabolism (Quentin et al. 2015; Hartmann and Trumbore 2016). SS is the main form for translating carbohydrates, and starch is the pivotal non-soluble longer-term storage compound. The concentrations of NSCs strongly influence plant growth and are sensitive to environmental changes (Ibrahim et al. 2011; de Souza et al. 2013). The proportional relationships between N, P and NSCs, to a large extent, reflect the amount of available C for plant growth and its use efficiency (Yin et al. 2009; Guo et al. 2015). Previous studies have mainly focused on the foliar stoichiometries of C, N and P, but the responses of NSC, N and P stoichiometries to elevated CO2 levels and N addition, and the ecological significance of these responses, are unclear, strongly hindering our understanding of the availability of C for plant growth and its relationships with elemental N and P.
Bothriochloa ischaemum (L.) Keng is a dominant species in natural grassland communities, which is widespread in the hilly-gully regions on the Loess Plateau. The current rate of N deposition in this area is 2.2 g N m−2 y−1 (Wei et al. 2010; Han et al. 2013) and is expected to increase in the future (Galloway et al. 2004). Elevated global levels of CO2 and N deposition would synergistically lead to changes in the nutrients cycles in this temperate area. The levels of both soil total N and P are low on the Loess Plateau (Liu et al. 2013b) and differ greatly from those in temperate and tropical forests, which are usually regarded as N- and P-poor, respectively (Huang et al. 2012). The response of perennial grass species to elevated CO2 levels in combination with N deposition has rarely been studied. The aim of the present research was to evaluate the concentrations and stoichiometries of NSCs, N, and P in whole plants, shoots and roots of B. ischaemum under elevated CO2 levels combined with N additions. We further hypothesized that (1) roots would respond more strongly than shoots to elevated CO2 concentration and N addition and (2) N addition would have a stronger effect than elevated CO2 concentration on the concentrations and stoichiometries of NSCs, N and P in this nutrient-poor area.
Materials and methods
Plant material and soil
Seeds of B. ischaemum, a C4 perennial herbaceous grass species, were collected in the autumn of 2013 from the experimental fields at the Ansai Research Station (ARS) (36°51′30″N, 109°19′23″E), Chinese Academy of Sciences, on the Loess Plateau of China. The station has a mean annual temperature of 8.8 °C, and receives a mean annual precipitation of 510 mm, most of which occurs from July to September. The rates of seed germination were >90% when germinated on moist filter paper in Petri dishes at 25 °C prior to the experiment.
The soil was collected from the upper 20 cm layer of a farmland at ARS. Soil gravimetric moisture content as a percentage of field capacity (FC) and the wilting point were 20 and 4%, respectively. The soil organic carbon, total nitrogen, and total phosphorus content were 1.50, 0.21, and 0.57 g kg−1, respectively. The soil was sieved through a 5 mm mesh before the experiment.
Experimental design
The cylindrical plastic pots (20 cm in depth, 15 cm in diameter) with sealed bottom were filled with 3.8 kg soil. Multiple seeds were sown in the pots on 1 June 2014. All pots were well watered in order to ensure seedling establishment. After emergence, seedlings were thinned to three plant per pot. The pots were transferred to two closed climate-controlled chambers (AGC-D001P, Qiushi Corp., Hangzhou, China) on 1 August 2014. The environmental conditions of the two chambers have been described detail elsewhere (Xiao et al. 2016). Briefly, one chamber received ambient CO2 concentration (400 μmol mol−1), the other received elevated CO2 concentration (800 μmol mol−1). The pots in each chamber received N fertilizer by spraying the seedlings twice a month at rates of total NH4NO3–N of 0, 2.5 and 5 g N m−2 y−1, respectively. All pots were regularly weighed, and water was added through plastic pipes to above 80% of FC until the end of the experiments. There were a total of six treatments were included: ambient CO2 concentration and no N fertilization (AN0), ambient CO2 concentration and low-N fertilization (AN1); ambient CO2 concentration and high-N fertilization (AN2); elevated CO2 concentration and no N fertilization (EN0), elevated CO2 concentration and low-N fertilization (EN1); elevated CO2 concentration and high-N fertilization (EN2). Each treatment had five replicates.
Sample collection and measurement
At the end of growing season, the three plants in each of pot were harvested. The shoots were severed from the roots at the crown. The roots were collected from the soil and carefully washed. Shoots and roots were dried at 80 °C for 48 h in oven to determine the dry weights (DW), respectively. Shoots and roots were ground using a mortar and pestle to fine powder prior to NSC, N, and P analyses.
NSCs were defined as the sum of SS and starch concentrations, which were measured using the anthrone method (Yemm and Willis 1954). The details of the extractions and measurements are described by Zhang et al. (2015). SS and starch concentrations were expressed as mg g−1 DW.
N and P concentrations were measured by Kjeldahl method and molybdenum blue method, respectively (Bao 2000). The N and P concentrations were presented in units of mg g−1 DW.
Statistical analysis
The SPSS 16.0 (SPSS Inc., Chicago, USA) was used for data analysis. Two-way analysis of variance (ANOVA) was conducted to evaluate the effects of CO2 and N treatments and their interactions on the concentrations and stoichiometries of NSCs, N and P based on whole plants, shoots and roots. Differences between each treatment were compared by Duncan’s multiple range tests at a probability level of 0.05.
Results
Plant Biomass
The elevated CO2 concentration and N addition had significantly positive effects on total, shoot and root biomasses and the root:shoot ratio (P <0.01 or <0.001) (Table 1). The elevated CO2 concentration increased total biomass by 47.14, 71.05 and 51.04% in the EN0, EN1 and EN2 treatments, respectively. N addition at the ambient CO2 concentration increased total biomass by 117.17 and 482.14% in the AN1 and AN2 treatments, respectively, and N addition at the elevated CO2 concentration increased total biomass by 152.43 and 497.57% in the EN1 and EN2 treatments, respectively. The interaction of the elevated CO2 concentration and N addition significantly affected total and root biomass (P < 0.01).
NSC, N and P concentrations
The elevated CO2 concentration significantly increased the starch and NSC concentrations in whole plants, shoots and roots (P <0.01 or <0.001) (Table 2; Fig. 1). N addition significantly decreased the SS concentration in whole plants and shoots and significantly increased the starch and NSC concentrations in whole plants, shoots and roots (P <0.01 or <0.001). The interaction of the elevated CO2 concentration and N addition significantly affected only the SS concentration in the shoots, with the highest and lowest values in the AN0 and EN2 treatments, respectively (P < 0.05) (Table 2; Fig. 1b).

SS (a–c), starch (d–f), NSC (g–i), N (j–l) and P (m–o) concentrations in whole plants, shoots and roots of B. ischaemum. Different letters indicate significant differences (P < 0.05) based on Duncan’s multiple range test
The elevated CO2 concentration had a significantly negative effect on the whole-plant P concentration (P < 0.05) (Table 2; Fig. 1) but not the P concentrations in the shoots and roots. N addition significantly increased the N concentrations in whole plants and roots and significantly decreased the P concentrations in whole plants, shoots and roots (P <0.01 or <0.001). The interaction of the elevated CO2 concentration and N addition significantly affected the P concentration in roots (P < 0.05) (Table 2).
Stoichiometries of NSCs, N and P
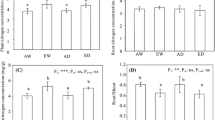
The elevated CO2 concentration significantly increased the starch:N and NSC:N ratios in whole plants, shoots and roots (Table 3; Fig. 2). N addition significantly decreased the SS:N ratio in whole plants, shoots and roots and significantly increased the starch:N and NSC:N ratios in whole plants and roots. The elevated CO2 concentration and N addition did not have a significant interactive effect on the SS:N, starch:N or NSC:N ratios.

SS:N (a–c), starch:N (d–f) and NSC:N (g–i) ratios in whole plants, shoots and roots of B. ischaemum. Different letters indicate significant differences (P < 0.05) based on Duncan’s multiple range test

SS:P (a–c), starch:P (d–f), NSC:P (g–i) and N:P (j–l) ratios in whole plants, shoots and roots of B. ischaemum. Different letters indicate significant differences (P < 0.05) based on Duncan’s multiple range test
The elevated CO2 concentration significantly increased the SS:P ratio in roots and the starch:P and NSC:P ratios in whole plants, shoots and roots (Table 3; Fig. 3). N addition significantly increased the SS:P ratio in whole plants and shoots and the starch:P and NSC:P ratios in whole plants, shoots and roots. The elevated CO2 concentration and N addition had interactive effects on the starch:P and NSC:P ratios in roots.
The elevated CO2 concentration significantly increased the N:P ratio in whole plants (Table 3; Fig. 3), and N addition significantly increased the N:P ratio in whole plants, shoots and roots. The elevated CO2 concentration and N addition did not have significant interactive effects on plant N:P ratios.
Discussion
Plant biomass
Soil nutrient availability modifies the responses of plant species to elevated CO2 levels. Soil total N content is low on the Loess Plateau (Liu et al. 2013b), so N addition would be better for plant growth. The growth and biomass allocation of plants in response to elevated CO2 levels and N addition are species-specific (Zhang et al. 2010, 2011). The significant positive effect in our study of the elevated CO2 concentration and N addition on the root:shoot ratio implies that root growth in B. ischaemum is more sensitive than shoot growth to elevated CO2 levels and N addition, which supports our first hypothesis.
NSC, N and P concentrations
NSCs are products of photosynthesis and provide energy for plant growth and metabolism. NSCs play a central role in the response of plants to environmental changes (Quentin et al. 2015). Elevated CO2 levels and N addition can synergistically enhance plant photosynthesis (Zhang et al. 2011; Chen et al. 2016) and thus increase NSC accumulation (Zhang et al. 2008; Zhu et al. 2016). Other studies have reported that elevated CO2 levels and/or N addition had no influence on, or even decreased, NSC concentrations due to higher carbohydrate use and the constraints of soil environmental stresses (Kakani et al. 2011; Pelletier et al. 2009; Alderman et al. 2011; Farrer et al. 2013; Sullivan et al. 2015; Liu et al. 2016). In the present study, the elevated CO2 concentration and N addition synergistically increased NSC accumulation, mainly due to the increase in starch. The SS content did not change significantly and even decreased under the elevated CO2 concentration and/or N addition. The elevated CO2 concentration and N addition significantly increased root biomass, so less energy was used for assimilating soil water and nutrients, and more SS, the main direct energy source for metabolism, was converted to starch (Latt et al. 2001), which was stored in the plant tissues.
N and P are important nutrients for plant growth (Huang et al. 2012). They are commonly considered as limiting factor for primary production and other ecosystemic processes. N and/or P concentrations generally decrease in plants with elevated CO2 levels because of nutrient dilution by the accumulation of NSCs (Loladze 2002; Deng et al. 2015). Elevated CO2 levels, though, can stimulate soil microbial processes and increase nutrient mineralization, which contribute to the uptake of more available N and P by the plants (Huang et al. 2012; Sardans et al. 2012). Our results showed that plant N concentration was little affected by the elevated CO2 concentration but significantly decreased the whole-plant P concentration, suggesting that B. ischaemum on the Loess Plateau is more limited by P than N under elevated CO2 levels. N deposition directly increases soil N availability and can increase the capacity of plants to take up P by stimulating the activity of phosphatase in the rhizosphere (Phoenix et al. 2004; Fujita et al. 2010). Increased N deposition thus usually increases N and/or P concentrations in plant tissues. In the present study, N addition significantly increased N concentrations in whole plants and roots, supporting our first hypothesis that roots would respond more strongly than shoots to N deposition. P concentrations, however, decreased significantly with N addition in whole plants, shoots and roots, indicating that the limiting effect of P was enhanced by N addition.
Stoichiometries of NSCs, N and P
NSCs are the main energy sources for plant growth, reproduction and survival under stress (Hartmann and Trumbore 2016; Song et al. 2016). The ratio of NSCs to N or P, to a large extent, reflects the relationship between the input of N or P and the output of NSCs and their use efficiencies (Li et al. 2008a, b; Guo et al. 2015). In accordance with our second hypothesis, the elevated CO2 concentration did not significantly influence the SS:N or SS:P ratios, and N addition significantly decreased the SS:N ratio and increased the SS:P ratio, implying that each unit of N produces less SS, and each unit of P produces more SS. The elevated CO2 concentration and N addition both significantly increased the starch:N, NSC:N, starch:P and NSC:P ratios, indicating that each unit of N and P produced more starch and NSCs, and the use efficiencies of N and P both increased.
N:P ratios can be used as diagnostic tool for evaluate nutrient limitation in terrestrial ecosystems under future environmental scenarios (Liu et al. 2013a; Huang et al. 2015a). The elevated CO2 concentration increased the whole-plant N:P ratio, and N addition significantly increased the whole-plant, shoot and root N:P ratios, supporting our second hypothesis. The higher N:P ratios under the elevated CO2 concentration and N addition were mainly due to the lower P concentrations. This result is consistent with those by Huang et al. (2012), who reported that elevated CO2 levels and N addition increased foliar N:P ratio in a subtropical model forest. Elevated CO2 levels and N addition can decrease N:P ratios but are mainly related to the increases P concentrations (Liu et al. 2013a), consistent with the results of a previous study reporting strong control of elevated CO2 levels and N addition on soil nutrient availability, especially P availability (Huang et al. 2014). The available-N:available-P ratio is much higher than the N:P ratio on the Loess Plateau (Jiao et al. 2013), and P mineralization cannot provide enough available P for plant growth under elevated CO2 levels and N addition. Plant growth in this region would thus suffer more from P than N limitation (Jiao et al. 2013; Xu et al. 2016), and N addition would exacerbate the P limitation on plant growth (An et al. 2011).
Conclusions
The elevated CO2 concentration and N addition significantly increased total biomass, starch and NSC concentrations and the root:shoot, starch:N, NSC:N, starch:P and NSC:P ratios in whole plants and roots of B. ischaemum. N addition alone decreased SS concentration in whole plants, increased N concentration and decreased P concentration in whole plants and roots and thus decreased the SS:N ratio and increased the SS:P and N:P ratios. Our results suggest that plant growth on the Loess Plateau suffers more from P than N limitation and that N addition would exacerbate the P limitation on plant growth.
References
Alderman PD, Boote KJ, Sollenberger LE, Coleman S (2011) Carbohydrate and nitrogen reserves relative to regrowth dynamics of ‘Tifton 8’ bermudagrass as affected by nitrogen fertilization. Crop Sci 51:1727–1738
An Z, Niu DC, Wen HY, Yang Y, Zhang HR, Fu H (2011) Effects of N addition on nutrient resorption efficiency and C:N:P stoichiometric characteristics in Stipa bungeana of steppe grasslands in the Loess Plateau, China. Chin J Plant Ecol 38:801–807
Bao SD (2000) Soil and agricultural chemistry analysis, 3rd edn. China Agriculture Press, Beijing, pp 268–270
Chen BB, Zou DH, Ma JH (2016) Interactive effects of elevated CO2 and nitrogen-phosphorus supply in the physiological properties of Pyropia haitanensis (Bangiales, Rhodophyta). J Appl Phycol 28:1235–1243
de Souza AP, Arundale RA, Dohleman FG, Long SP, Buckeridge MS (2013) Will the exceptional productivity of Miscanthus x giganteus increase further under rising atmospheric CO2? Agric Forest Meteorol 171–172: 82–92
Deng Q, Cheng XL, Zhou GY, Liu JX, Liu SZ, Zhang QF, Zhang DQ (2013) Seasonal responses of soil respiration to elevated CO2 and N addition in young subtropical forest ecosystems in southern China. Ecol Eng 61:65–73
Deng Q, Hui DF, Luo YQ, Elser J, Wang YP, Loladze I, Zhang QF, Dennis S (2015) Down-regulation of tissue N:P ratios in terrestrial plants to elevated CO2. Ecology 96:3354–3362
Dong JL, Li X, Duan ZQ (2016) Biomass allocation and organs growth of cucumber (Cucumis sativus L.) under elevated CO2 and different N supply. Arch Agron Soil Sci 62:277–288
Farrer EC, Herman DJ, Franzova E, Pham T, Suding KN (2013) Nitrogen deposition, plant carbon allocation, and soil microbes: changing interactions due to enrichment. Am J Bot 100:1458–1470
Fujita Y, Robroek BJM, de Ruiter PC, Heil GW, Wassen MJ (2010) Increased N affects P uptake of eight grassland species: the role of root surface phosphatase activity. Oikos 119:1665–1673
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland CC, Green PA, Holland EA, Karl DM, Michaels AF, Porter JH, Townsend AR, Vöosmarty CJ (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Gruber N, Galloway JN (2008) An earth-system perspective of the global nitrogen cycle. Nature 451:293–296
Guo ZW, Hu JJ, Yang QP, Li YC, Chen SL, Chen WJ (2015) Influence of mulching management on the relationships between foliar non-structural carbohydrates and N, P concentrations in Phyllostachys violascens stand. Chinese. J Appl Ecol 26:1064–1070
Han WX, Fang JY, Reich PB, Woodward FI, Wang ZH (2011) Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol Lett 14:788–796
Han XW, Tsunekawa A, Tsubo M, Shao HB (2013) Responses of plant-soil properties to increasing N deposition and implications for large-scale eco-restoration in the semiarid grassland of the northern Loess Plateau, China. Ecol Eng 60:1–9
Hartmann H, Trumbore S (2016) Understanding the roles of nonstructural carbohydrates in forest trees-from what we can measure to what we want to know. New Phytol 211:386–403
Heimann M, Reichstein M (2008) Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 451:289–292
Huang WJ, Zhou GY, Liu JX, Zhang DQ, Xu ZH, Liu SZ (2012) Effects of elevated carbon dioxide and nitrogen addition on foliar stoichiometry of nitrogen and phosphorus of five tree species in subtropical model forest ecosystems. Environ Pollut 168:113–120
Huang WJ, Zhou GY, Liu JX, Duan HL, Liu XZ, Fang X, Zhang DQ (2014) Shifts in soil phosphorus fractions under elevated CO2 and N addition in model forest ecosystems in subtropical China. Plant Ecol 215:1373–1384
Huang WJ, Houlton BZ, Marklein AR, Liu JX, Zhou GY (2015a) Plant stoichiometric responses to elevated CO2 vary with nitrogen and phosphorus inputs: evidence from a global-scale meta-analysis. Sci Rep 5:18225
Huang WJ, Zhou GY, Deng XF, Liu JX, Duan HL, Zhang DQ, Chu GW, Liu SZ (2015b) Nitrogen and phosphorus productivities of five subtropical tree species in response to elevated CO2 and N addition. Eur J Forest Res 134:845–856
Huang WJ, Zhou GY, Liu JX, Zhang DQ, Liu SZ, Chu GW, Fang X (2015c) Mineral elements of subtropical tree seedlings in response to elevated carbon dioxide and nitrogen addition. PLoS ONE 10:e0120190
Ibrahim MH, Jaafar HZE, Rahmat A, Rahman ZA (2011) The relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules 16:162–174
Jiao F, Wen ZM, An SS, Yuan Z (2013) Successional changes in soil stoichiometry after land abandonment in Loess Plateau, China. Ecol Eng 58:249–254
Kakani VG, Vu JCV, Allen LH Jr, Boote KJ (2011) Leaf photosynthesis and carbohydrates of CO2 enriched maize and grain sorghum exposed to a short period of soil water deficit during vegetative development. J Plant Physiol 168:2169–2176
Latt CR, Nair PKR, Kang BT (2001) Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate. For Ecol Manag 146:145–158
Lee M, Manning P, Rist J (2010) A global comparison of grassland biomass responses to CO2 and nitrogen enrichment. Philos Trans R Soc B 365:2047–2056
Lee SH, Kim SH, Ding WX, Kang H (2015) Impact of elevated CO2 and N addition on bacteria, fungi, and archaea in a marsh ecosystem with various types of plants. Appl Microbiol Biotechnol 99:5295–5305
Li MH, Xiao WF, Shi PL, Wang SG, Zhong YD, Liu XL, Wang XD, Cai XH, Shi ZM (2008a) Nitrogen and carbon source-sink relationships in trees at the Himalayan treelines compared with lower elevations. Plant Cell Environ 31:1377–1387
Li MH, Xiao WF, Wang SG, Cheng GW, Cherubini P, Cai XH, Liu XL, Wang XD, Zhu WZ (2008b) Mobile carbohydrates in Himalayan treeline trees. I. Evidence for carbon gain limitation but not for growth limitation. Tree Physiol 28:1287–1296
Liu JX, Zhang DQ, Zhou GY, Duang HL (2012) Changes in leaf nutrient traits and photosynthesis of four tree species: effects of elevated CO2, N fertilization and canopy positions. J Plant Ecol 5:376–390
Liu JX, Huang WJ, Zhou GY, Zhang DQ, Liu SZ, Li YY (2013a) Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests. Global Change Biol 19:208–216
Liu ZP, Shao MA, Wang YQ (2013b) Spatial patterns of soil total nitrogen and soil total phosphorus across the entire loess plateau region of China. Geoderma 197–198:67–78
Liu JX, Fang X, Deng Q, Han TF, Huang WJ, Li YY (2015) CO2 enrichment and N addition increase nutrient loss from decomposing leaf litter in subtropical model forest ecosystems. Sci Rep 5:7952
Liu J, Wu NN, Wang H, Sun JF, Peng B, Jiang P, Bai E (2016) Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 97:1796–1806
Loladze I (2002) Rising atmospheric CO2 and human nutrition: toward globally imbalanced plant stoichiometry? Trends Ecol Evol 17:457–461
Luo YQ, Su B, Currie WS, Dukes JS, Finzi A, Hartwig U, Hungate B, McMurtrie RE, Oren R, Parton WJ, Pataki DE, Shaw RM, Zak DR, Field CB (2004) Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 54:731–739
Marklein AR, Houlton BZ (2012) Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol 193:696–704
Newingham BA, Vanier CH, Charlet TN, Ogle K, Smith SD, Nowak RS (2013) No cumulative effect of 10 years of elevated CO2 on perennial plant biomass components in the Mojave Desert. Glob Change Biol 19:2168–2181
NOAA (2013) http://www.esrl.noaa.gov/gmd/ccgg/trends/
Novotny AM, Schade JD, Hobbie SE, Kay AD, Kyle AD, Reich PB, Elser JJ (2007) Stoichiometric response of nitrogen-fixing and non-fixing dicots to manipulations of CO2, nitrogen, and diversity. Oecologia 151:687–696
Novriyanti E, Watanabe M, Kitao M, Utsugi H, Uemura A, Koike T (2012) High nitrogen and elevated CO2 effects on the growth, defense and photosynthetic performance of two eucalypt species. Environ Pollut 170:124–130
Pelletier S, Tremblay GF, Lafreniere C, Bertrand A, Belanger G, Castonguay Y, Rowsell J (2009) Nonstructural carbohydrate concentrations in timothy as affected by N fertilization, stage of development, and time of cutting. Agron J 101:1372–1380
Phoenix GK, Booth RE, Leake JR, Read DJ, Grime JP, Lee JA (2004) Simulated pollutant nitrogen deposition increases P demand and enhances root-surface phosphatase activities of three plant functional types in a calcareous grassland. New Phytol 161:279–289
Quentin AG, Pinkard EA, Ryan MG, Tissue DT, Baggett LS, Adams HD, Maillard P, Marchand J, Landhäusser SM, Lacointe A, Gibon Y, Anderegg WR, Asao S, Atkin OK, Bonhomme M, Claye C, Chow PS, Clément-Vidal A, Davies NW, Dickman LT, Dumbur R, Ellsworth DS, Falk K, Galiano L, Grünzweig JM, Hartmann H, Hoch G, Hood S, Jones JE, Koike T, Kuhlmann I, Lloret F, Maestro M, Mansfield SD, Martínez-Vilalta J, Maucourt M, McDowell NG, Moing A, Muller B, Nebauer SG, Niinemets Ü, Palacio S, Piper F, Raveh E, Richter A, Rolland G, Rosas T, Saint Joanis B, Sala A, Smith RA, Sterck F, Stinziano JR, Tobias M, Unda F, Watanabe M, Way DA, Weerasinghe LK, Wild B, Wiley E, Woodruff DR (2015) Non-structural carbohydrates in woody plants compared among laboratories. Tree Physiol 35:1146–1165
Reich PB, Hobbie SE, Lee T, Ellsworth DS, West JB, Tilman D, Knops JM, Naeem S, Trost J (2006) Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440:922–925
Sardans J, Rivas-Ubach A, Penuelas J (2012) The C:N:P stoichiometry of organisms and ecosystems in a changing word: a review and perspectives. Perspect Plant Ecol 14:33–47
Sillen WMA, Dieleman WIJ (2012) Effects of elevated CO2 and N fertilization on plant and soil carbon pools of managed grasslands: a meta-analysis. Biogeosciences 9:2247–2258
Song XZ, Peng CH, Zhou GM, Gu HH, Li Q, Zhang C (2016) Dynamic allocation and transfer of non-structural carbohydrates, a possible mechanism for the explosive growth of Moso bamboo (Phyllostachys heterocycla). Sci Rep 6:25908
Stiling P, Cornelissen T (2007) How does elevated carbon dioxide (CO2) affect plant–herbivore interactions? A field experiment and meta-analysis of CO2-mediated changes on plant chemistry and herbivore performance. Glob Change Biol 13:1823–1842
Sullivan PF, Ellison SBZ, McNown RW, Brownlee AH, Sveinbjörnsson B (2015) Evidence of soil nutrient availability as the proximate constraint on growth of treeline tress in northwest Alaska. Ecology 96:716–727
Wei Y, Tong YA, Qiao L, Liu XJ, Duan M, Li J (2010) Preliminary estimate of the atmospheric nitrogen deposition in different ecological regions of Shaanxi province. J Agro-Environ Sci 29:795–800
Xiao L, Liu GB, Xue S (2016) Elevated CO2 concentration and drought stress exert opposite effects on plant biomass, nitrogen, and phosphorus allocation in Bothriochloa ischaemum. J Plant Growth Regul 35:1088–1097
Xu BC, Gao ZJ, Wang J, Xu WZ, Palta JA, Chen YL (2016) N:P ratio of the grass Bothriochloa ischaemum mixed with the legume Lespedeza davurica under varying water and fertilizer supplies. Plant Soil 1–2:67–79
Yang YH, Luo YQ, Lu M, Schädel C, Han WJ (2011) Terrestrial C:N stoichiometry in response to elevated CO2 and N addition: a synthesis of two meta-analyses. Plant Soil 343:393–400
Yemm E, Willis A (1954) The estimation of carbohydrates in plant extracts by anthrone. Biochem J 57:508–514
Yin JJ, Guo DL, He SY, Zhang L (2009) Non-structural carbohydrate, N, and P allocation patterns of two temperate tree species in a semi-arid region of Inner Mongolia. Acta Sci Nat Univ Pekin 45: 519–527
Zhang YB, Duan BL, Qiao YZ, Wang KK, Korpelainen H, Li CY (2008) Leaf photosynthesis of Betula albosinensis seedlings as affected by elevated CO2 and planting density. For Ecol Manag 255:1937–1944
Zhang L, Yang YX, Zhan XY, Zhang CJ, Zhou SX, Wu DX (2010) Responses of a dominant temperate grassland plant (Leymus chinensis) to elevated carbon dioxide and nitrogen addition in China. J Environ Qual 39:251–259
Zhang L, Wu DX, Shi HQ, Zhang CJ, Zhan XY, Zhou SX (2011) Effects of elevated CO2 and N addition on growth and N2 fixation of a legume subshrub (Caragana microphylla Lam.) in temperate grassland in China. PLoS ONE 6:e26842
Zhang T, Cao Y, Chen YM, Liu GB (2015) Non-structural carbohydrate dynamics in Robinia pseudoacacia saplings under three levels of continuous drought stress. Trees 29:1837–1849
Zhu CW, Xu X, Wang D, Zhu JG, Liu G, Seneweera S (2016) Elevated atmospheric CO2 stimulates sugar accumulation and cellulose degradation rates of rice straw. GCB Bioenergy 8:579–587
Acknowledgements
This research was funded by the Western Young Scholars of the Chinese Academy of Sciences (XAB2015A05), and the National Natural Science Foundation of China (41371510, 41371508, 41471438, and 41330858).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Xiao, L., Liu, G., Li, P. et al. Nitrogen addition has a stronger effect on stoichiometries of non-structural carbohydrates, nitrogen and phosphorus in Bothriochloa ischaemum than elevated CO2 . Plant Growth Regul 83, 325–334 (2017). https://doi.org/10.1007/s10725-017-0298-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-017-0298-8