
Abstract
Reducing water salinity towards iso-osmotic conditions is a common practice applied in euryhaline fish farming to limit osmoregulation costs and enhance growth. In this respect, the present study investigated the timing of salinity reduction in an abrupt manner during European eel (Anguilla anguilla) larval culture by examining associated impacts on morphological and molecular levels. Larvae from 3 different parental combinations (families) were reared at constant 36 psu for 6 days (control) or subjected to a direct reduction to 18 psu on 1, 2, or 3 days post-hatch. Overall, salinity reduction enhanced growth and survival, resulting from more efficient energy resource utilization. In the control group, expression of growth-related igf2 remained constant, demonstrating a steady growth progression, while igf1 expression increased over time only for the salinity reduced treatments, potentially qualifying as a useful biomarker for growth performance. Even though each parental combination seems to have a different capacity to cope with salinity alterations, as observed by family-driven water-transport-related aquaporin (aqp1, aqp3) gene expression, it could be inferred that the abrupt salinity change is generally not stressful, based on non-upregulated heat shock proteins (hsp70, hsp90). However, the applied salinity reduction (irrespective of timing) induced the development of pericardial edema. As such, we conclude that despite the positive effect of salinity reduction on early growth and survival, the long-term benefit for eel larval culture lies in establishing a protocol for salinity reduction, at a precise developmental time point, without causing pericardial malformations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
To facilitate growth during fish larval development in culture, the high energy demand of osmoregulation can be reduced by adjusting the culture environment towards salinity conditions, which facilitate energy resources to be utilized with greater efficiency (reviewed in Bœuf and Payan 2001). Intermediate salinity regimes have been frequently employed, with positive results concerning larval survival and growth for several fish species (Gaumet et al. 1995; Tandler et al. 1995; Imsland et al. 2001; Rahmah et al. 2020). However, the mechanisms involved in the processes of decreasing osmoregulatory energy expenditure and acting beneficially can be multidimensional. Along these lines, better feed intake (Bœuf and Payan 2001) and feed conversion efficiency rates (Gaumet et al. 1995) have previously been reported. Overall, an aquatic environment with osmotic pressure matching the osmolality of larvae (iso-osmotic) is suggested to be less stressful (Deane et al. 2002), thus allowing marine fish larvae in culture to thrive at lowered salinity due to reduced energy costs for maintenance of homeostasis.
European eel (Anguilla anguilla) is a catadromous fish with a migration pattern connecting distant oceanic spawning habitats with continental juvenile on-growing habitats (Schmidt 1923; Righton and Metcalfe 2011). In these corresponding native environments, eels have adapted to different salinity regimes, developing elaborate osmoregulatory mechanisms (Cutler and Cramb 2001; McCormick 2001; Bone and Moore 2008). During early life history stages, eels are encountered exclusively in the ocean (Schmidt 1923; Tsukamoto 1992), where they experience hyperosmotic conditions (Lee et al. 2013). It is therefore anticipated that eel larvae must possess genetically pre-programmed mechanisms for osmoregulation, allowing to cope with osmotic and ionic gradients between their body fluid osmolality and the surroundings (Varsamos et al. 2005). Even though osmoregulatory organs are absent during the early developmental stages, eel larvae exhibit numerous ion-transporting ionocytes on their body surface already at hatch (Sasai et al. 1998, 2007; Seo et al. 2015; Kuroki et al. 2016). These mitochondrial-rich cells require a significant proportion of metabolic energy to be allocated for osmoregulation (Morgan and Iwama 1991; Bœuf and Payan 2001; Bone and Moore 2008).
In aquaculture, eel offspring are commonly reared in seawater at 36 psu (Sørensen et al. 2016a), but applying a reduction in salinity to half-strength seawater during larval rearing has proven beneficial (Okamura et al. 2009; Kurokawa et al. 2013; Kuroki et al. 2016; Politis et al. 2018a). However, the application techniques and timing of such a salinity reduction have received little attention. In European eel, it was shown that the earlier a salinity reduction is applied, the better survival and growth rates are achieved (Politis et al. 2018a). For the closely related Japanese eel, a stable full-strength salinity regime is suggested until at least 4 days post-hatch (dph), based on the emergence of morphological deformities after early exposure of pre-leptocephali to low-salinity conditions (Kurokawa et al. 2013). More specifically, malformations, such as pericardial edema and open lower jaw may occur, with high mortalities during early ontogeny often being attributed to such abnormal phenotypes (Okamura et al. 2007, 2011, 2016, 2018; Okamoto et al. 2009).
Regarding the aforementioned considerations, on the one hand, the timing of application should accommodate early life development, while on the other, salinity change should be in line with culture efficiency. Politis et al. (2021) showed that a stepwise salinity reduction close to iso-osmotic levels (18 psu) improves larval growth and survival. However, such an approach challenges technical aspects of recirculating aquaculture system (RAS) technology, by the need of either multiple RAS units stabilized to different fixed salinities or by dynamically altering salinity in each operating RAS. The latter scenario substantially compromises RAS biofilter stability (Kinyage et al. 2019), where particularly in saltwater systems, abrupt lowered salinity negatively affects biofilter function and reduces vital nitrification processes (Lyssenko and Wheaton 2006). Considering these aspects, the above approaches do not serve as sustainable solutions, calling for more understanding regarding physiological tolerances during larviculture. Interestingly though, eel larvae are able to tolerate a drastic change of salinity, performed by moving the larval rearing tanks from one stable RAS unit at 36 psu to another stable RAS unit at 18 psu (Politis et al. 2021), which represents a cost-efficient solution. Nonetheless, the biologically most suited timing for applying this salinity reduction is yet to be elucidated.
The present study aimed to investigate the application timing of such a salinity decrease and the biological significance for eel larvae from a morphometric (survival, biometrics, and deformity) and molecular (gene expression) perspective. For this, European eel larvae from three parental combinations (families) were reared either at constant 36 psu for 6 days (control) or subjected to a direct salinity reduction to 18 psu on 1, 2, or 3 dph.
Materials and methods
Broodstock management and gamete production
Female broodstock were collected from nature (Saltbæk Vig, 55°44′48.8"N 11°09′09.5"E, Denmark; and Lake Vandet, 57°00′50.1"N 8°33′16.5"E, Denmark), while male broodstock originated from glass eels raised at a commercial Danish eel farm (Royal Danish Fish, Hanstholm, Denmark). Males and females were transported to the EEL-HATCH facility in Hirtshals (Denmark), where they were reared in closed recirculation systems with a flow rate of 10–15 L min−1 and acclimated to 36 psu and 20 °C over the course of 2 weeks. Stimulation of spermatogenesis and milt collection were performed according to previously described assisted reproduction protocols (Tomkiewicz et al. 2011; Butts et al. 2014; Sørensen et al. 2016b), while stimulation of vitellogenesis and ovulation was performed using salmon pituitary extract and dihydroprogesterone, respectively (Tomkiewicz et al. 2019; Kottmann et al. 2020). The reproductively mature males (n = 14) had a mean standard length of 39.1 ± 3.8 cm and a body weight of 132.0 ± 23.6 g, while females (n = 3) had a mean standard length of 70.3 ± 5.7 cm and a body weight of 695.3 ± 221.3 g, respectively.
Egg fertilization and incubation
The eggs of each female (n = 3) were strip-spawned, mixed with milt originating from a combination of 3–5 males (Butts et al. 2014; Benini et al. 2018) and fertilized at an effective salinity of 36 psu and 20 °C (Sørensen et al. 2016a), to create the 3 family crosses used in this study. The buoyant eggs/embryos were transferred to 60-L conical flow-through incubators, as described in Sørensen et al. (2016b) and Politis et al. (2018b). Light was kept dimmed (Politis et al. 2014), temperature set at ~ 18 °C (Politis et al. 2017), and gentle aeration was applied until ~ 52 h post fertilization (hpf), while bottom purging was regularly applied for removal of dead eggs. Air diffusers were removed prior to hatching, which occurred at ~ 56 hpf.
Experimental design and conditions
The experimental setup included two identical RAS units (Fig. 1), each consisting of a ~ 1-m3 reservoir, 0.2-m3 header tank, 0.2-μm cartridge filter for particle removal (Cuno/3 M, 10-in. PDA 0.2-μm cartridge, Silhorko-Eurowater A/S, Skanderborg, Denmark), and UV treatment (UltraAqua, Aalborg, Denmark). The systems did not contain a biofilter and did not receive returning water from the 2-L acrylic experimental rearing jars. One system was kept at 36 ± 0.3 psu, while the other was adjusted to 18 ± 1.8 psu using municipal tap water (non-chlorinated groundwater). Temperature in both systems was maintained at 18.8 ± 0.4 °C (Politis et al. 2017).

Schematic representation of the experimental setup. European eel (Anguilla anguilla) larvae were reared either at a constant salinity of 36 psu (control) over 6 days post-hatch (dph) or experienced an abrupt salinity reduction on 1, 2, or 3 dph by moving larval rearing tanks from the 36-psu system to the 18-psu system
The experiment was repeated 3 times, each time using offspring from a different family (4 salinity treatments × 3 families × 3 replicates = 36 experimental units). For each family, newly hatched larvae (day 0) were randomly distributed (~ 800 individuals per replicate) into 12 acrylic 2-L flow-through jars (drz400sm hank, JugDesk Type, Taipei, Taiwan) with custom-designed bottom inflow and top outflow. All larval rearing tanks were initially connected to the 36 psu system. Thereafter, 3 jars were connected to the 18 psu system on day 1 (treatment 1), another 3 jars on day 2 (treatment 2), and finally another 3 jars on day 3 (treatment 3). The last 3 jars (control) remained connected to the 36-psu system the entire period (Fig. 2). Water exchange rate was adjusted to ~ 0.1 L min−1 for all jars. Eel larvae were reared under these experimental regimes for 6 days under low light intensity (Politis et al. 2014).

Schematic representation of the experimental treatments, where European eel (Anguilla anguilla) larvae were reared over 6 days post-hatch (dph) under constant salinity of 36 psu (control) or experiencing a salinity reduction to 18 psu on 1 (treatment 1), 2 (treatment 2), or 3 (treatment 3) dph
Data collection
Mortality and biometry
Larval sampling was conducted on 0, 1, 2, 3 and 6 dph. Here, ~ 10 larvae from each replicate (n = 3), family (n = 3), and treatment (n = 4) were randomly sampled, anesthetized using ethyl 3-aminobenzoate methanesulfonate (MS-222, Sigma-Aldrich Chemie, Steinheim, Germany), and photographed using a zoom stereomicroscope (SMZ1270i fitted DS-Fi2 Camera Head, Nikon Corporation, Tokyo, Japan). Subsequently, larval images were analyzed for total body area, oil-drop area, and pericardial edema using the NIS-Elements-D analysis software (Nikon Corporation, Tokyo, Japan). Based on the derived measurements, the following estimates were calculated:
To study the severity of heart malformation, the ratio of edema height relative to neurocranium height was discriminated and categorized as either minor (0.5–0.8) or severe (> 0.8) according to Kurokawa et al. (2008) and Okamoto et al. (2009). Dead larvae were counted and removed from all experimental units on a daily basis to minimize microbial interference (Sørensen et al. 2014). At the completion of the experiment, all larvae were enumerated and survival was estimated taking into account all the sampled larvae throughout the experimental process. Larval cumulative mortality was calculated as a percentage from hatch until 6 dph.
Gene expression
For molecular analysis, a pool of ~ 20 larvae from each replicate (n = 3), family (n = 3), and treatment (n = 4) were randomly sampled on 0, 1, 2, 3, and 6 dph. These larvae were recorded, euthanized using MS-222, preserved in RNAlater Stabilization Reagent, and kept at − 20 °C following the procedures suggested by the supplier (Qiagen, Germany). RNA was extracted using the NucleoSpin® RNA Kit (Macherey–Nagel, Germany) following the manufacturer’s instructions. RNA concentration and purity were determined by spectrophotometry using Nanodrop® ND-1000 (Peqlab, Germany) and then reverse-transcribed using the qScript™ cDNA Synthesis Kit (Quantabio, Germany) according to the manufacturer’s instructions, including an additional gDNA wipe out step (PerfeCta® DNase I Kit (Quantabio, Germany)). The expression levels of target genes were determined by quantitative real-time PCR (RT-qPCR), using specific primers (Table 1). Primers were designed to target all known potential isoforms with an amplification size ranging from 75 to 200 nucleotides and optimal Tm of 60 °C, using primer 3 software (http://frodo.wi.mit.edu/primer3/) based on cDNA sequences available in GenBank databases.
Expressions of genes in each larval sample from 2 randomly selected replicates, from each family (n = 3), treatment (n = 4), and larval age (0, 1, 2, 3, and 6 dph) were analyzed in two technical replicates using the qPCR Biomark™ HD technology (Fluidigm, USA) based on dynamic arrays (GE chips). The chip was run according to the Fluidigm protocol with a Tm of 60 °C. The relative quantity of target gene transcripts was normalized (ΔCT) to the geometric mean of the 2 (reference) genes (ef1a and npr3), after qBase + software revealed that these mRNA levels were stable throughout analyzed samples (Hellemans et al 2007). Coefficient of variation (CV) of technical replicates was calculated and checked. Further analysis of gene expression was carried out according to the 2−ΔΔCt method (Livak and Schmittgen 2001).
Statistical analysis
All data were analyzed using SAS software (v.9.1; SAS Institute Inc., Cary, NC, USA). Residuals were evaluated for normality (Shapiro–Wilk test) and homoscedasticity (plot of residuals vs. predicted valves) to ensure they met model assumptions. Data were log10 or arcsine square root transformed, when necessary. Alpha was set at 0.05 for main effects and interactions. Treatment means were contrasted using Tukey’s honest significance test. Cumulative mortality data were analyzed using a mixed model repeated measures ANOVA, where the main model variables were salinity treatment (fixed factor), age (fixed factor), and the salinity treatment × age interaction (fixed factor). Body area, oil droplet area, growth rate, oil droplet utilization, growth efficiency, and pericardial edema ratio were analyzed using a series of mixed model factorial ANOVAs. Additionally, a series of mixed effects models were run to investigate how salinity treatment impacts gene expression at each age and how gene expression changes throughout early larval ontogeny for each salinity regime. In these above models, family and the family × salinity treatment interaction were considered random factors. Variance components (VC) for random effects were generated using the restricted maximum likelihood (REML) estimation method and expressed as a percentage.
Results
Mortality and biometry
The earlier the salinity reduction was induced, the smaller the larval size in terms of (final) body area reached on 6 dph, while larvae raised in full-strength salinity water (Control) developed the significantly smallest body area compared to any of the salinity reduction treatments (p = 0.021; Fig. 3A). Accordingly, larval growth rate was lower for the control group, with no significant differences observed among the rest of the treatments (p = 0.021; Fig. 3B).

Final body area (A), growth day−1 (B), final oil droplet area (C), oil droplet utilization day−1 (D), growth efficiency (E), and pericardial edema severity ratio (F) of European eel (Anguilla anguilla) larvae reared from 0 to 6 days post-hatch (dph) under constant salinity of 36 psu (Control) or exposed to a salinity reduction to 18 psu on 1 (treatment 1), 2 (treatment 2), and 3 (treatment 3) dph. Values represent means of 3 families (3 replicates × 10 individuals per family) and error bars represent least square means standard error. Values with the same letters are not significantly different at alpha of 0.05
Significantly less energy reserves, in terms of (final) oil droplet area on 6 dph, were observed in larvae experiencing the abrupt salinity reduction on 3 dph (treatment 3) compared to treatments 1 and 2, while the lowest energy reserves were observed in larvae reared in full-strength seawater (control) (p = 0.001; Fig. 3C). Furthermore, significantly lower oil droplet utilization was observed when salinity reduction was implemented on 1 and 2 dph (p = 0.001; Fig. 3D). Consequently, this coincides with the growth efficiency values, which were highest for larvae experiencing a salinity reduction on 1 and 2 dph (p < 0.001; Fig. 3E), while the stable salinity regime of 36 psu (control) displayed the lowest growth efficiency among all treatments.
Concerning the condition of pericardial edema, it was exclusively observed in larvae from the three salinity reduced treatments (pericardial edema severity ratio > 0.8). Nevertheless, the developmental stage at which salinity reduction was introduced had no significant implications on the magnitude of the specific malformation (p = 0.005; Fig. 3F).
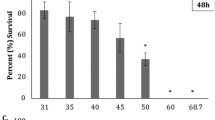
The cumulative mortality (Fig. 4) was significantly (p < 0.05) higher for larvae reared in full-strength seawater (control), while no differences were detected among the salinity reduction treatments. Moreover, no significant salinity treatment × age interaction was observed.

Cumulative mortality (%) of European eel (Anguilla anguilla) larvae reared from 0 to 6 days post-hatch (dph) under constant salinity of 36 psu (control) or exposed to a salinity reduction to 18 psu on 1 (treatment 1), 2 (treatment 2), or 3 (treatment 3) dph. Values represent means of 3 families (3 replicated tanks per family). Values with the same letters are not significantly different at alpha of 0.05
Gene expression
As revealed by the VC analysis (Table 2), the expression patterns of genes related to water transport (aqp1, aqp3) were significantly (p < 0.05) family driven on 1, 2, and 3 dph, while they were driven by a significant genome (family) × environment (salinity treatment) interaction on 6 dph (aqp1: p = 0.037; aqp3: p = 0.015). The family effect was also significant (p < 0.05) for stress-related genes (hsp70, hsp 90) on 2 and 3 dph, where it explained up to 47.1% of the observed variability. Similarly, expression patterns of growth-related genes were found to be predominantly influenced by family on 2 dph for igf1 and on 1 and 2 dph for igf2.
The expression levels of neither aquaporin nor heat-shock protein groups were affected (p > 0.05) by larval age and remained steady throughout ontogeny. Likewise, no significant fluctuations were noted for the expression of igf2 among developmental stages. Still, igf1 expression was affected by developmental age (Fig. 5), where expression increased over time in all reduction treatments (treatment 1: p = 0.002; treatment 2: p = 0.046; treatment 3: p < 0.0001), while it remained stable throughout ontogeny for larvae reared at constant 36 psu.

Effect of age on European eel (Anguilla anguilla) larval relative expression of insulin-like growth factor 1 (igf1). Each panel (A–D) illustrates a different salinity scenario under which European eel (Anguilla anguilla) larvae were reared over 6 days post-hatch (dph): (A) constant salinity at 36 psu (control) or experiencing a salinity reduction to 18 psu on (B) 1 (treatment 1), (C) 2 (treatment 2), or (D) 3 (treatment 3) dph. Values represent means of 3 families (2 replicates × 20 individuals per family × 2 technical replicates) and error bars represent least square means standard error. Values with the same letters are not significantly different at alpha of 0.05
Discussion
Today, production of European eel offspring is deemed essential to support sustainable aquaculture development and alleviate pressure from the depleted wild eel stocks (ICES 2020). Despite the notion that farming should imitate the conditions corresponding to the species- and stage-specific natural habitat, the employment of rather abnormal rearing environments may in fact facilitate production. The present study investigated the application timing of unnatural and abrupt salinity changes from 36 to 18 psu during eel larviculture, to elucidate the biological significance for eel larvae from a morphometric and molecular point of view. At the end of the experiment (6 dph), growth in body area was significantly increased by salinity reduction towards iso-osmotic conditions. As such, the assumption of energy saving due to lower osmoregulation costs was here confirmed by the higher availability of internal energy reserves (in terms of yolk and oil-droplet) in larvae experiencing a salinity reduction, which were more efficiently utilized and invested into development instead of maintaining ionic and water balance. However, larvae experiencing this salinity change already on day 1 did not grow as big as larvae switching to 18 psu later in development. This indicates that newly hatched larvae might have restricted ability to cope with abrupt salinity changes, which could be compromising their growth potential.
Similar to growth, survival was lowest for the control group, where larvae were reared in full strength salinity. Interestingly, mortality increased the longer the larvae remained in high-salinity conditions, confirming previous observations by Politis et al. (2021), where applying such a drastic salinity change on 6 dph did not improve eel larval survival compared to larvae constantly reared in full-strength seawater. As such, the present study confirms that the early establishment of iso-osmotic conditions generally accommodates the survival of weak larvae that do not survive in full-strength salinity. However, it needs to be mentioned that in the current study, all salinity reduction treatments, irrespective of the timing the salinity reduction was induced, caused deformities in terms of pericardial edema. This pathology seems to be directly provoked by low salinity since almost all individuals reared in reduced salinity were diagnosed with severe heart edema on 6 dph. There is no evidence that pericardial edema is directly lethal, at least until yolk absorption (Kurokawa et al. 2008), but negative consequences, especially during the first-feeding stages, are most likely. In order to prevent this deformity in Japanese eel culture, it is suggested to maintain full-strength seawater during the early yolk sac stages (Kurokawa et al. 2013) and shift to low salinity conditions on 6 dph (Okamura et al., 2016), enabling feeding procedures on the bottom of so-called Kreiseltanks, while positive effects on survival are sustained in the long term. However, it is worth mentioning that Japanese eel larvae are reared at higher temperatures than European eel larvae; and thus, the same age (6 dph) corresponds to a later developmental stage (first-feeding). In this regard, it still remains to be clarified whether salinity preferences and tolerance limits, as well as respective physiological responses to environmental alterations, are developmental stage (reviewed in Varsamos et al. 2005) and/or species-dependent.
Moreover, in this study, we investigated the mechanism of cellular protein repair, which is mainly driven by heat shock proteins (HSP) and activated when cellular protein damage occurs under osmotic and/or heat stress (Deane et al. 2002). As such, a hyper- or hypo-osmotic shock is expected to upregulate the expression of hsps compared to iso-osmotic conditions (Deane et al. 2002). However, in this study, the expression patterns of genes (hsp70, hsp90) relating to this mechanism were not directly affected by salinity, supporting the assumption that either the mechanism has not gained full functionality yet to respond to such extreme physiological changes, or that the salinity regimes applied in the current study are within the physiological tolerance threshold limits of this species. Additionally, the expression patterns of hsp70 and hsp90 were not affected by the age or stage of eel larvae, as they were constantly expressed throughout development, but differed among batches on 2 and 3 dph, indicating that each family might have a different capacity to handle environmental (salinity) changes. In this regard, the sensitivity of the relevant molecular mechanism could be inheritable, rendering eel larvae genetically pre-programmed to respond according to the “good genes” hypothesis (Neff and Pitcher 2005).
We also targeted processes such as growth, metabolism, and development, which are regulated by underlying molecular mechanisms involving the somatotropic axis, interlinking insulin-like growth factors (IGF) and growth hormones (GH) (Reinecke et al 2005). Here, we attempted to molecularly trace growth, associated to salinity treatments, by following expression patterns of two genes (igf1 and igf2), which are an integral part of the abovementioned mechanism. So far, it has been reported that iso-osmotic conditions enhance growth and trigger the igf1 transcription (Deane et al. 2002). However, we did not detect differences in expression of neither igf1 nor igf2 between the salinity treatments. This insinuates that it is either developmentally too early to clearly decipher the sensitivity of this mechanism to environmental parameters (such as salinity), or that growth is mediated post-translationally (Scott et al. 2004). With respect to the latter, similar results have been obtained for stunted coho salmon, Oncorhynchus kisutch, when osmoregulation was out of control and igf1 production was hampered despite the high GH plasma levels (reviewed in Bœuf and Payan 2001). In this regard, further research possibly examining the larval dry weight and protein levels under different salinity scenarios and/or over a longer period may shed more light on the above theory. Moreover, similar to hsps, growth-related expression of igf1 and igf2 was influenced by family (on 1 and 2 dph), indicating the importance of maternally supplied yolk and the oil droplet, as growth of larvae in the present study was exclusively dependent on internal energy reserves. These internal sources are commonly utilized throughout the endogenous feeding phase, until larvae become dependent on external dietary factors throughout the following exogenous feeding stage (which was not explored in this study). Therefore, it was presumable to experience a sustained expression of igf2 over time, demonstrating a steady growth progression. On the contrary, expression of igf1 increased over time, at least for the salinity reduction treatments, indicating that igf1 might be a useful future biomarker to molecularly trace growth throughout development.
Furthermore, we molecularly explored the mechanism of water transport, mainly regulated by aquaporins (AQP). These membrane proteins function as water channels, which control body fluid homeostasis (Borgnia et al. 1999). Different homologues have been described depending on their selectivity. Among them, aqp1 is exclusively permeable to water, whereas aqp3 is additionally permeable to glycerol and urea (Borgnia et al. 1999). In adult European eel, their expression varies depending on the tissue, with renal aqp1 being downregulated (Martinez et al. 2005a) and intestinal aqp1 being upregulated (Martinez et al. 2005b) during seawater acclimation. On the other hand, gill aqp3 levels have been shown to decrease under similar conditions (Tse et al. 2006). In this study of eel larvae, deciphering tissue specific expression was not possible due to size limitations. Thus, whole larvae were used instead, where aqp1 and aqp3 were both expressed at constant levels within the ontogenetic window investigated (not affected by age/stage), potentially revealing to be of basal importance throughout eel early larval ontogeny. Moreover, expression of both aqp1 and aqp3 was family driven during the earliest stages, but on 6 dph, expression patterns were controlled by the genome (family) × environment (salinity) interaction instead. As such, these results reveal a family-specific phenotypic plasticity translating into a variable sensitivity of each batch to salinity, which should be taken into consideration in future larviculture of this species.
Conclusion
The present study showed that applying drastic salinity changes in European eel larviculture, where larvae are directly transferred from full strength seawater (36 psu) to 18 psu, can be realized, since this technique requires only two stable RAS units “balanced” at the desired salinity levels. Even though, the capacity of eel larvae to adapt to salinity changes was found to be batch-specific, the abrupt salinity changes applied in this study were within the threshold tolerance limits of eel larvae, indicating no physiological injury by the osmotic stress applied and supported by the even improved growth and survival. However, larvae suffer from pericardial edema, with no observed negative consequence during the endogenous pre-feeding period, but with potential sub-chronical implications during the later exogenous feeding stage. As such, we conclude that despite the positive effect of salinity reduction on early survival (probably by saving weak individuals), the long-term benefit for eel larviculture lies in the production of healthy offspring. Thus, the challenge ahead is finding the balance between this trade-off process and establishing a protocol for salinity reduction, at a developmental time point, without causing heart edema, while still benefiting feeding procedures and survival throughout the feeding culture.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Benini E, Politis SN, Sørensen SR et al (2018) Effect of parental origin on early life history traits of European eel. Reprod Dom Anim 53:1149–1158. https://doi.org/10.1111/rda.13219
Bœuf G, Payan P (2001) How should salinity influence fish growth? Comp Biochem Physiol Part C 130:411–423
Bone Q, Moore RH (2008) Osmoregulation and ion balance. In: Owen E (ed) Biology of fishes: Third Edition. Taylor & Francis Group In, pp 161–207
Borgnia M, Nielsen S, Engel A, Agre P (1999) Cellular and molecular biology of the aquaporin water channels. Annu Rev Biochem 68:425–458
Butts IAE, Sørensen SR, Politis SN et al (2014) Standardization of fertilization protocols for the European eel, Anguilla anguilla. Aquaculture 426–427:9–13. https://doi.org/10.1016/j.aquaculture.2014.01.020
Cutler CP, Cramb G (2001) Molecular physiology of osmoregulation in eels and other teleosts: the role of transporter isoforms and gene duplication. Comp Biochem Physiol Part A 130:551–564
Deane EE, Kelly SP, Luk JCY, Woo NYS (2002) Chronic salinity adaptation modulates hepatic heat shock protein and insulin-like growth factor I expression in Black Sea Bream. Mar Biotechnol 4:193–205. https://doi.org/10.1007/s1012602-0091-5
Gaumet F, Boeuf G, Sever A et al (1995) Effects of salinity on the ionic balance and growth of juvenile turbot. J Fish Biol 47(5):865–876
Hellemans J, Mortier G, Paepe A De et al (2007) qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol 8.https://doi.org/10.1186/gb-2007-8-2-r19
ICES (2020) European eel (Anguilla anguilla) throughout its natural range. In: Report of the ICES Advisory Committee, 2020. ICES Advice 2020
Imsland AK, Foss A, Gunnarsson S et al (2001) The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus). Aqua 198:353–367
Kinyage JPH, Pedersen PB, Pederse L-F (2019) Effects of abrupt salinity increase on nitrification processes in a freshwater moving bed biofilter. Aquac Eng 84:91–98. https://doi.org/10.1016/j.aquaeng.2018.12.005
Kottmann JS, Jørgensen MGP, Bertolini F et al (2020) Differential impacts of carp and salmon pituitary extracts on induced oogenesis, egg quality, molecular ontogeny and embryonic developmental competence in European eel. PLoS ONE 15:e0235617. https://doi.org/10.1371/journal.pone.0235617
Kurokawa T, Okamoto T, Gen K et al (2008) Influence of water temperature on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. J World Aquac Soc 39:726–735
Kurokawa T, Shibahara H, Gen K et al (2013) Determination of periods of sensitivity to low-salinity and low-temperature conditions during the early development of cultured Japanese eel Anguilla japonica larvae with respect to the rate of morphological deformity at completion of yolk resorption. Fish Sci 79:673–680. https://doi.org/10.1007/s12562-013-0636-3
Kuroki M, Seo MY, Okamura A et al (2016) Morphofunctional features of ionocytes in Japanese eel Anguilla japonica leptocephali acclimated to half-diluted and full-strength seawater. Ichthyol Res 63:487–495. https://doi.org/10.1007/s10228-016-0520-0
Lee KM, Yamada Y, Okamura A et al (2013) Hyposmoregulatory ability and ion- and water-regulatory mechanisms during the leptocephalus stages of Japanese eel Anguilla japonica. Fish Sci 79:77–86. https://doi.org/10.1007/s12562-012-0576-3
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real- time quantitative PCR and the 2−ΔΔCT method. Methods 25:402–408. https://doi.org/10.1006/meth.2001.1262
Lyssenko C, Wheaton F (2006) Impact of rapid impulse operating disturbances on ammonia removal by trickling and submerged-upflow biofilters for intensive recirculating aquaculture. Aquacult Eng 35(1):38–50
Martinez A-S, Cutler CP, Wilson GD et al (2005a) Cloning and expression of three aquaporin homologues from the European eel (Anguilla anguilla): effects of seawater acclimation and cortisol treatment on renal expression. Biol Cell 97:615–627
Martinez A-S, Cutler CP, Wilson GD et al (2005b) Regulation of expression of two aquaporin homologs in the intestine of the European eel: effects of seawater acclimation and cortisol treatment. Am J Physiol Regul Integr Comp Physiol 288:1733–1743. https://doi.org/10.1152/ajpregu.00747.2004
McCormick SD (2001) Endocrine control of osmoregulation in teleost fish. Am Zool 41:781–794
Morgan JD, Iwama GK (1991) Effects of salinity on growth, metabolism, and ion regulation in juvenile rainbow and steelhead trout (Oncorhynchus mykiss) and fall Chinook salmon (Oncorynchus tshawytscha). Can J Fish Aquat Sci 48:2083–2094
Neff BD, Pitcher TE (2005) Genetic quality and sexual selection: an integrated framework for good genes and compatible genes. Mol Ecol 14:19–38. https://doi.org/10.1111/j.1365-294X.2004.02395.x
Okamoto T, Kurokawa T, Gen K et al (2009) Influence of salinity on morphological deformities in cultured larvae of Japanese eel, Anguilla japonica, at completion of yolk resorption. Aquaculture 293:113–118. https://doi.org/10.1016/j.aquaculture.2009.04.005
Okamura A, Yamada Y, Horie N et al (2007) Effects of water temperature on early development of Japanese eel Anguilla japonica. Fish Sci 73:1241–1248. https://doi.org/10.1111/j.1444-2906.2007.01461.x
Okamura A, Yamada Y, Mikawa N et al (2009) Growth and survival of eel leptocephali (Anguilla japonica) in low-salinity water. Aquaculture 296:367–372. https://doi.org/10.1016/j.aquaculture.2009.08.039
Okamura A, Yamada Y, Mikawa N et al (2011) Notochord deformities in reared Japanese eel Anguilla japonica larvae. Aquaculture 317:37–41. https://doi.org/10.1016/j.aquaculture.2011.04.024
Okamura A, Yamada Y, Mikawa N et al (2016) Effect of salinity on occurrence of notochord deformities in Japanese eel Anguilla japonica larvae. Aquac Int 24:549–555. https://doi.org/10.1007/s10499-015-9944-1
Okamura A, Horie N, Mikawa N et al (2018) Influence of temperature and feeding regimes on growth and notochord deformity in reared Anguilla japonica leptocephali. Fish Sci 84:505–512. https://doi.org/10.1007/s12562-018-1188-3
Politis SN, Butts IAE, Tomkiewicz J (2014) Light impacts embryonic and early larval development of the European eel, Anguilla anguilla. J Exp Mar Bio Ecol 461:407–415. https://doi.org/10.1016/j.jembe.2014.09.014
Politis SN, Mazurais D, Servili A et al (2017) Temperature effects on gene expression and morphological development of European eel, Anguilla anguilla larvae. P L O S One 12:e0182726. https://doi.org/10.1371/journal.pone.0182726
Politis SN, Syropoulou E, Benini E et al (2021) Performance thresholds of hatchery produced European eel larvae reared at different salinity regimes. Aquaculture 539:736651. https://doi.org/10.1016/j.aquaculture.2021.736651
Politis SN, Mazurais D, Servili A et al (2018a) Salinity reduction benefits European eel larvae: insights at the morphological and molecular level. PLOS One 13.https://doi.org/10.1371/journal.pone.0198294
Politis SN, Sørensen SR, Mazurais D et al (2018b) Molecular ontogeny of first-feeding European eel larvae. Front Physiol 9.https://doi.org/10.3389/fphys.2018b.01477
Rahmah S, Liew HJ, Napi N, Rahmat SA (2020) Metabolic cost of acute and chronic salinity response of hybrid red tilapia Oreochromis sp. larvae. Aquac Rep 16:100233. https://doi.org/10.1016/j.aqrep.2019.100233
Reinecke M, Björnsson BT, Dickhoff WW et al (2005) Growth hormone and insulin-like growth factors in fish: where we are and where to go. Gen Comp Endocrinol 142:20–24. https://doi.org/10.1016/j.ygcen.2005.01.016
Righton DA, Metcalfe JD (2011) Eel migrations. In: Encyclopedia of fish physiology. Elsevier Inc., pp 1937–1944
Sasai S, Kaneko T, Tsukamoto K (1998) Extrabranchial chloride cells in early life stages of the Japanese eel, Anguilla japonica. Ichthyol Res 45:95–98
Sasai S, Katoh F, Kaneko T, Tsukamoto K (2007) Ontogenic change of gill chloride cells in leptocephalus and glass eel stages of the Japanese eel, Anguilla japonica. Mar Biol 150:487–496. https://doi.org/10.1007/s00227-006-0355-8
Schmidt J (1923) The breeding places of the eel. Philos Trans R Soc Lond Ser B 211:179–208
Scott GR, Richards JG, Forbush B et al (2004) Changes in gene expression in gills of the euryhaline killifish Fundulus heteroclitus after abrupt salinity transfer. Am J Physiol Cell Physiol 287:300–309
Seo MY, Kuroki M, Okamura A et al (2015) Occurrence of larval and adult types of ion-secreting ionocytes in Japanese eel Anguilla japonica. Ichthyol Res 62:487–494. https://doi.org/10.1007/s10228-015-0463-x
Sørensen SR, Skov PV, Lauesen P et al (2014) Microbial interference and potential control in culture of European eel (Anguilla anguilla) embryos and larvae. Aquaculture 426–427:1–8. https://doi.org/10.1016/j.aquaculture.2014.01.011
Sørensen SR, Anthony I, Butts E et al (2016a) Effects of salinity and sea salt type on egg activation, fertilization, buoyancy and early embryology of European eel, Anguilla anguilla. Zygote 24:121–138. https://doi.org/10.1017/S0967199414000811
Sørensen SR, Tomkiewicz J, Munk P et al (2016b) Ontogeny and growth of early life stages of captive-bred European eel. Aquaculture 456:50–61. https://doi.org/10.1016/j.aquaculture.2016.01.015
Tandler A, Anav FA, Choshniak I (1995) The effect of salinity on growth rate, survival and swimbladder inflation in gilthead seabream, Sparus aurata, larvae. Aqua 135:343–353
Tomkiewicz J, Kofoed TM, Pedersen JS (2011) Assessment of testis development during induced spermatogenesis in the European eel Anguilla anguilla. Mar Coast Fish 3(1):106–118. https://doi.org/10.1080/19425120.2011.556902
Tomkiewicz J, Politis SN, Sørensen SR, Butts IAE, Kottmann JS (2019) European eel - an integrated approach to establish eel hatchery technology in Denmark. In: Don A, Coulson P (eds) Eels - biology, monitoring, management, culture and exploitation: Proceedings of the First International Eel Science Symposium. 5M Publishing, pp 340–374
Tse WKF, Au DWT, Wong CKC (2006) Characterization of ion channel and transporter mRNA expressions in isolated gill chloride and pavement cells of seawater acclimating eels. Biochem Biophys Res Commun 346(346):1181–1190. https://doi.org/10.1016/j.bbrc.2006.06.028
Tsukamoto K (1992) Discovery of the spawning area for Japanese eel. Nature 356:789–791
Varsamos S, Nebel C, Charmantier G (2005) Ontogeny of osmoregulation in postembryonic fish: a review. Comp Biochem Physiol Part A 141:401–429. https://doi.org/10.1016/j.cbpb.2005.01.013
Acknowledgements
We would like to thank Paraskevas Koumpiadis and Annika Toth for broodstock husbandry and provision of gametes for the experiment, Eftychia Maria Goniou for embryonic rearing, and Eugenia Capatina, Joanna Miest, and Adrian Loh for assistance in molecular work.
Funding
This study received funding from the Innovation Fund Denmark under grant agreement no. 7076-00125B (ITS-EEL) and from “ENV-Fonden.”
Author information
Authors and Affiliations
Contributions
JT, SP, SS, and IB provided funding, while JT, SP, and SS designed the study. JT established the assisted broodstock protocols providing gametes for the experiment. SS constructed and tailored the larval systems and rearing tanks. ES, SP, and EB conducted the experiment and collected samples. SP and EB carried out the gene expression analysis. IB performed the statistical analysis. SP and SS made illustrations. ES wrote original manuscript draft. All the authors contributed to data interpretation and manuscript revision.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval
All fish were handled according to the European Union regulations concerning the protection of experimental animals (Dir 86/609/EEC). The experimental protocol for the present study was approved by the Danish Ministry of Food, Agriculture and Fisheries (permit number: 2015–15-0201–00696). In particular, broodstook used were anesthetized with ethyl p-aminobenzoate (benzocaine) prior to any handling, while efforts were made to minimize animal handling stress.
Consent for publication
All the authors approved the submitted version of this manuscript.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Syropoulou, E., Benini, E., Sørensen, S.R. et al. Early and abrupt salinity reduction impacts European eel larval culture. Fish Physiol Biochem 48, 355–366 (2022). https://doi.org/10.1007/s10695-022-01056-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-022-01056-6