
Abstract
Eichhornia crassipes, commonly known as water hyacinth, is a free-floating perennial aquatic plant native to South America, which has been widely introduced on different continents, including Africa. E. crassipes is abundant in both the Congo (Africa) and Amazon (South America) River catchments. We performed a comparative analysis of the ostracod communities (Crustacea, Ostracoda) in the E. crassipes pleuston in the Amazon (South America) and Congo (Africa) River catchments. We also compared the ostracod communities from the invasive E. crassipes with those associated with stands of the native African macrophyte Vossia cuspidata. We recorded 25 species of ostracods associated with E. crassipes in the Amazon and 40 in the Congo River catchments, distributed over 31 ostracod species in E. crassipes and 27 in V. cuspidata. No South American invasive ostracod species were found in the Congolese pleuston. Diversity and richness of Congolese ostracod communities was higher in the invasive (Eichhornia) than in a native plant (Vossia). The highest diversity and abundance of ostracod communities were recorded in the Congo River. The result of principal coordinates analysis, used to evaluate the (dis)similarity between different catchments, showed significant differences in species composition of the communities. However, a dispersion homogeneity test (PERMDISP) showed no significant differences in the variability of the composition of species of ostracods (beta diversity) within Congo and Amazon River catchments. It appears that local ostracod faunas have adapted to exploit the opportunities presented by the floating invasive Eichhornia, which did not act as “Noah’s Ark” by introducing South American ostracods in the Congo River.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Several aquatic plants native to South America have successfully invaded other continents. E. crassipes (Mart.) Solms, commonly known as the water hyacinth, has invaded freshwater systems in over 50 countries on five continents (Villamagna and Murphy 2010). Eichhornia crassipes is a free-floating perennial aquatic plant and is especially pervasive throughout Southeast Asia, the southeastern United States, central and western Africa and Central America (see overview in Villamagna and Murphy 2010). The water hyacinth is a pest species in many waterbodies, this in spite of many efforts to eliminate or at least control its populations. Introduction of this plant species in different parts of the world might have been accidental and/or intentional. Indeed, intentional introductions to ponds have been common, as the water hyacinth is an ornamental plant that also reduces nutrient concentrations and algae blooms (Villamagna and Murphy 2010).
Floating vegetation (i.e. not rooted in the sediment) is a typical feature of South American floodplains, where its submerged system of mostly roots hosts rich communities of invertebrate (and fish) taxa; these communities are called pleuston.
Ostracods are small bivalved crustaceans inhabiting sediments (Pieri et al. 2007; Van der Meeren et al. 2010; Szlauer-Lukaszewska 2014) and aquatic macrophytes (Higuti et al. 2010; Liberto et al. 2012; Mazzini et al. 2014; Matsuda et al. 2015), which are common in inland waters. There are about 2000 species of non-marine Ostracoda described in approximately 200 genera in various non- marine environments in the world (Martens et al. 2008), of which about 275 species in 55 genera are known from South and Central America (Martens and Behen 1994; Martens et al. 2008), while nearly 500 species in 84 genera of ostracods have been described from African inland waters (Martens 1984, 1998; Martens et al. 2008).
Currently, 117 species represented by 37 genera have been reported from Brazil (Martens and Behen 1994; Martens et al. 2008; Würdig and Pinto 2001; Pinto et al. 2003, 2004, 2005a, b, 2008; Higuti et al. 2009, 2013; Higuti and Martens 2012a, b, 2014). In both Africa and South America, more than 80 % of all species are endemic to their continent and zoogeographical region.
Eichhornia crassipes, native to South America and invasive in Africa, is abundant in both the Congo (Democratic Republic of Congo) and Amazon (Brazil) River catchments. Many studies reported on the negative impact of invasive aquatic plants on their own communities and/or on the ecosystems in general. However, there is little information about the introduction of aquatic organisms (periphyton, invertebrates) together with these invasive plants. Some biological features of ostracods could facilitate their long-distance dispersal, such as the presence of drought-resistant eggs (Horne and Martens 1998) so that they might colonize a new habitat and successfully establish a new population. Furthermore, the reproduction of many species of ostracods happens by parthenogenesis (Martens et al. 2008), so that a new population can be formed by a single founder specimen or egg.
The objectives of the present study were to determine (1) if Congolese Eichhornia comprises exclusive Congolese faunas or if also invasive (South American) species occur; (2) if ostracods communities in native (Vossia cuspidata (Roxb.) Griff.) and invasive (E. crassipes) macrophyte stands in the Congo River catchment differ and (3) if ostracod biodiversity within and between the Amazon and Congo River catchments are significantly different.
Materials and methods
Study area
The Amazon River in South America is the second longest in the world and by far the largest by water flow with an average discharge of approximately 214 million liters per second. During the rainy season, the water discharged in the Atlantic Ocean travels approximately 160 km out into sea. The Amazon has the largest river basin in the world, occupying more than 6.8 million km2; rainfall is evenly distributed spatially and temporally, ranging from 1500 to 2500 mm annually, for about 6 months a year (Goulding et al. 2003). The Amazon River and its tributaries are accompanied along their middle and lower courses by large fringing floodplains that cover an area of about 300,000 km2, the flooded forest is known as várzea (Irion et al. 1997).
The Congo River has the second highest discharge (1325 km3/year or 42 million liters/s) of any river in the world after the Amazon (Coynel et al. 2005). It drains 3.7 million km2 of the African continent (Campbell 2005). It covers about 25 % of the total wet tropical zone and is responsible for almost 3.4 % of the freshwater inputs to the Atlantic Ocean (Probst and Tardy 1987). In the wet season, most of the forests are flooded, while in the dry season they dry out. The Congo Basin has a humid tropical climate: the mean annual precipitation, calculated between 1980 and 1990 is close to 1550 mm (Mahé 1993).
The Amazon and Congo systems, situated across the equator, are characterized by tropical rain forest and by extensive floodplains. However, there are major differences in catchment characteristics. For example, the runoff in the Amazon River is higher (Filizola 2003) because the headwaters of the Amazon catchment are located in high mountains in contrast with the plateaus or hills of the Congo catchment (Meybeck et al. 2001).
Collections and measurements
The ostracod collection of the Amazon floodplain used in the present study results from a project in the SISBIOTA program (National System of Biodiversity Research) funded by CNPq/Fundação Araucária. The project covered four Brazilian floodplains (in addition to the Amazon: Paraná, Araguaia and Pantanal). Two sampling campaigns were conducted in the Amazon floodplain, the first one was done in October 2011 (dry season) and the second one in May 2012 (rainy season). More than 20 lakes or lake-like habitats were sampled, mostly for their pleuston communities in Eichhornia stands.
The ostracod collection from the Congo River was effected during the large Congo-expedition in May and June 2010 (Boyekoli Ebala Congo 2010—http://www.congobiodiv.org/en). More than 50 samples were collected, either in the pleuston of the invasive species E. crassipes or amongst stands of the native V. cuspidata.
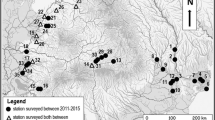
We used data from ostracods associated with E. crassipes and V. cuspidata of the same sampling period in both river catchments to compare the ostracod fauna. The localities sampled in the Amazon and Congo River catchments are generally located close to cities or village. Thirteen localities were selected for each of the river catchments (Fig. 1). In each locality, two replicate samples of each aquatic macrophyte (E. crassipes in Amazon, E. crassipes and V. cuspidata in Congo) were taken.

Location of the sampling stations of Congo River catchment (localities numbers 1–13) and Amazon River catchment (localities numbers 14–26). Source Original (see acknowledgments)
Eichhornia crassipes individuals of similar size were hand collected and were placed in plastic buckets to remove the ostracods; roots were separated from the leaves and were washed in the bucket. The residuals were filtered in a hand net (mesh size c 160 µm). Ostracods in Vossia stands were collected directly by moving the handnet through the rooted plants for c 5 min. Ostracods were killed by adding 97 % ethanol to the wet residual; all samples were washed again in the lab and were transferred to fresh ethanol (70 %). Samples were sorted under a microscope stereoscopic. Specimens were identified using valves (Scanning Electron Microscopy) and appendages (soft parts dissected in slides with light microscopy).
The environmental variables measured in the field included water temperature (°C) and dissolved oxygen (mg L−1) (oxymeter-YSI), pH and electrical conductivity (µS cm−1) (YSI 63). Some geographical, physical, and chemical features of the waterbodies of Amazon and Congo River catchments are listed in Table 1.
Data analyses
Richness, abundance, Shannon–Wiener diversity and evenness were calculated to assess differences in the ostracods communities in Amazon and Congo River catchments. A parametric analysis of variance (ANOVA) was performed to test for significance of differences in species richness, abundance, diversity and evenness of ostracods between plants species and between catchments. When the homogeneity assumption required for ANOVA was not fulfilled, a non-parametric Kruskal–Wallis test was used. Analyses of variance were performed in Statistica 7.1 (StatSoft 2005).
Rarefaction analysis was performed, because of differences in abundance amongst the samples, in order to compare species richness between the sampling units at comparable levels of density in both catchments. Rarefaction curves were computed using the BioDiversity Pro Programme (McAleece et al. 1997).
Principal coordinates analysis (PCoA) was used to evaluate the (dis)similarity of species composition between different catchments. Differences between the two catchments were tested by ANOVA applied to the scores of PCoA axes. The dispersion homogeneity test (PERMDISP) was performed to test the variability in the ostracods species composition (beta diversity) within Amazon and Congo River catchments. This analysis calculates a centroid for each catchment, and determines the Bray–Curtis distance of each sampling locality to the centroid. The higher average of Bray–Curtis distances to the centroid means greater dissimilarity in species composition, suggesting an increase in beta diversity. The significance (p < 0.05) in beta diversity between catchments was analyzed by a permutation test of least squares residuals, using 999 permutations. The analyses was performed using the R 3.0.1 software (R Development Core Team 2013) through vegan and permute packages (Oksanen et al. 2013).
Results
We recorded 25 species of ostracods associated to E. crassipes in the Amazon River catchment and 40 species in the Congo River catchment. Of these 40 Congolese species, 31 were found in the invasive E. crassipes and 27 ostracod species occurred in the native V. cuspidata (See appendix 1 and 2 in Electronic supplementary material). The most speciose ostracod subfamily was Cypricercinae (8 species in E. crassipes) in the Amazon; and Cypridopsinae (9 species in both E. crassipes and V. cuspidata) and Herpetocypridinae (7 species in E. crassipes and 6 species in V. cuspidata) in the Congo River catchment (Table 2). The only ostracod species present in both catchments was Stenocypris major, a circumtropical species. Several ostracod species in the Amazon and nearly all species in the Congo River catchment are new to science and remain to be described.
The highest diversity and abundance of ostracods were recorded in the Congo River catchment in both aquatic plants species, where richness and Shannon diversity values were higher in the invasive E. crassipes (Fig. 2a–c). However, no significant difference was observed regarding these attributes between E. crassipes and V. cuspidata. When comparing diversity of ostracods associated with E. crassipes between both catchments, significantly higher values of species richness (p < 0.01) and diversity (p < 0.05) were found in the invasive plant from the Congo River catchment. The evenness values were similar between Amazon and Congo River catchments (Fig. 2d).

Mean values and standard error of a richness, b abundance, c Shannon diversity and d evenness of ostracods community associated with E. crassipes (Ec) and V. cuspidata (Vc) in Amazon (AM) and Congo River (DRC) catchments
Rarefaction curves indicated higher number of ostracods species in the Congo River catchment, especially on the invasive E. crassipes, for similar numbers of individuals. The richness reached an asymptote in the E. crassipes samples from the Amazon River catchment, while it is still increasing in the E. crassipes and V. cuspidata samples of the Congo River catchment (Fig. 3), meaning that with more sampling most likely still more species could be found there.

Rarefaction curves of ostracods species associated with different aquatic plants in Amazon (AM) and Congo River catchments (DRC). Ec = E. crassipes and Vc = V. cuspidata. +: native E. crassipes of the Amazon River catchment, ●: invasive E. crassipes of the Congo River catchment, -: native V. cuspidata of the Congo River catchment
The result of the PCoA, used to evaluate the (dis)similarity between different catchments, showed significant differences in species composition, assessed for the scores of PCoA, axis 1 (F = 170.98; p < 0.00) and axis 2 (F = 5.39; p < 0.05). The dispersion homogeneity test (PERMDISP) showed no significant differences (F = 0.31; p = 0.59) in the variability of ostracod species composition (beta diversity) within Amazon and Congo River catchments. The highest average distance to the centroid was recorded in the Amazon River catchment (0.58) compared to the Congo River catchment (0.56) (Fig. 4).

Multivariate permutation analysis diagram showing the variability in ostracods species composition associated with E. crassipes between different catchments. Full square Amazon River catchment, circle: Congo River catchment
Discussion
Composition of ostracod communities
Ostracods are amongst the most abundant invertebrates associated with macrophytes and are themselves a group that have a high diversity of species (Thomaz et al. 2008; Higuti et al. 2007, 2010; Liberto et al. 2012; Mazzini et al. 2014; Matsuda et al. 2015).
The Cyprididae are the most species ostracod family in the Amazon and Congo River catchments. This is in line with the fact that this family comprises c 50 % of all known Recent non-marine ostracod species and more than 80 % of all species in most tropical regions (Martens 1998). The patterns of species composition are different between both catchments: Cypricercinae (32 % of species) and Cypridopsinae (20 %) are the most dominant subfamilies in the Amazon River catchment, Cypridopsinae (30 %) and Herpetocypridinae (20 %) in the Congo River catchment. These findings are in agreement with those of Martens (1998), who compared the diversity of non-marine Ostracoda from Africa and South America.
The comparison of the ostracod communities between the two catchments clearly shows that the pleuston in the Amazon River catchment only contained South American species, while those in the Congo River catchment only contained Congolese ostracod species, or in any case did not have any identifiable South American (or other non-African) ostracod species, with Stenocypris major being the only shared, circumtropical, ostracod species. The invasive E. crassipes therefore did not act as a “Noah’s Arc” by transporting also invasive ostracod species into that part of the Congo River catchment. This can be either because the invasive plant was introduced as seeds or propagules, in which case no pleuston was associated with it yet, or because the invasive pleuston in adult plants failed to establish viable communities in the new environment.
However, several ostracod species do have the capacity to invade new areas, and have already done so in the past (see Table 3).
Comparison between ostracod communities in native (Amazon) and invasive (Congo) E. crassipes
Our results showed higher ostracod diversity and abundance in the invasive E. crassipes in the Congo River catchment than in the native E. crassipes specimens from the Amazon River catchment. This is a rather unexpected result, as one could predict to have a greater diversity of ostracod species in E. crassipes of the Amazon River catchment, where this plant is native and where ostracod species are expected to be better adapted to life in the root system of this (and other) floating plants. The fact that the collections were sampled in a small part of the Amazon floodplain relatively near to the city of Manaus might be part of the reason. However, the rarefaction analyses showed that the cumulative curve of ostracod species in E. crassipes from the Amazon River catchment had reached the maximum plateau with our collections. The results are thus representative for at least that part of the Amazon River catchment. Moreover, size of root systems in this macrophyte species can vary considerably and is known to depend at least in part on water chemistry (Kobayashi et al. 2008). Observations showed that the root systems of this plant in the Congo River catchment samples were much smaller than those from the Amazon River catchment, down to less than half the volume (results not shown). This makes the higher values of richness, diversity and abundance of the ostracod communities in the Congolese samples even more surprising.
The present results indicate that invasive species such as E. crassipes do not necessarily always have a fully negative effect on the invaded communities. Invasive E. crassipes root systems obviously offer novel substrates for native animals, such as ostracods, that can adapt to life in pleuston. However, this is clearly only one side of a broader picture, where other native communities such as zooplankton or whole community levels might be negatively impacted.
Comparison between ostracod communities in native (V. cuspidata) and invasive (E. crassipes) plant species in the Congo River catchment
We observed that the ostracod communities associated with an invasive plant species, E. crassipes, have a higher richness, diversity and abundance than those associated to a native plant species, V. cuspidata, in the Congo River catchment. This is a clear example of an invasive species physically facilitating communities of native species [see review in Rodriguez (2006)].
Also Mormul et al. (2010) observed higher values of these attributes of ostracod communities in an invasive plant, Hydrilla verticillata when compared to a native macrophyte, Egeria najas, in the Paraná River floodplain, although these differences were not significant. However, cumulative curves did indicate higher diversity of ostracod species on the invasive H. verticillata. The invasive plants might thus provide favorable habitats for native ostracod communities. The experiment by Mormul et al. (2010) was performed to test the effect of an invasive plant on the ostracod communities, using two plants with similar architecture (as measured in fractal heterogeneity). In the present case, E. crassipes and V. cuspidata do not have similar architecture: Eichhornia is a floating plant with normally dense root systems with high heterogeneity, whereas Vossia is a grass-like rooted plant, with low individual heterogeneity but occurring in dense stands. This density of the macrophyte stands will to a certain degree compensate for the lower individual heterogeneity.
Figueiredo et al. (2015) conducted an experiment using a native (E. najas) and an invasive (H. verticillata) macrophyte species to test the preference of a native fish prey species for either macrophyte as a refuge in the presence of an invasive voracious predator. It appeared that the prey fish did not show preference for one plant over the other, and they concluded that habitat structure affects choice more than evolutionary history between fish and macrophytes and this is in line with our results.
The effects of invasive plants on aquatic communities, however, is more complex and depends on more factors than on heterogeneity of the habitat only. Villamagna and Murphy (2010) showed that abundance and diversity of aquatic invertebrates generally increase in response to increased habitat heterogeneity and structural complexity provided by E. crassipes, but can also decrease due to decreased planktonic algae (food) availability. Stiers et al. (2011) compared macrophytes and macroinvertebrates communities of 32 Belgium ponds, of which 22 included invasive plants. Negative impact on native plants and macroinvertebrates was recorded for three invasive plants, reducing the richnees of both communities. Schultz and Dibble (2012) summarized the role of invasive plants on freshwater fish and macroinvertebrate communities. They found that positive effects of invasive macrophytes on fish and macroinvertebrate communities were associated with characteristics held in common with native macrophytes such as photosynthesis, increasing habitat heterogeneity, and stabilizing substrate. The authors found that three other traits of invasive plants can largely be responsible for negative effects on fish and macroinvertebrate communities: increased growth rate, allelopathic chemical production, and phenotypic plasticity that allow for greater adaptation to environmental conditions and resource utilization than native species.
Conclusions
Ostracod communities in Congo and Amazon River catchments are highly dissimilar, indicating that the invasive E. crassipes root systems in the Congo River catchment were colonized by African ostracods, and that the arrival of invasive E. crassipes did not bring along successful invasions of South American ostracod species, nor species from other parts of the world from where the plants may have been introduced into the Congo basin. Also, richness, diversity and abundance of ostracod communities in the invasive Congolese plants are higher than in the native Amazonian plants. Finally, richness, diversity and abundance is also higher in the invasive E. crassipes than in the native V. cuspidata in the Congo River catchment, but this could in part be owing to lower complexity of the native plant. It appears that local African ostracod faunas have adapted to exploit the habitat opportunities presented by the floating invasive E. crassipes.
References
Bruton MN, Merron SV (1985) Alien and translocated aquatic animals in Southern Africa: a general introduction, checklist and bibliography. South African National Scientific Programmes Report no. 113. Republic of South Africa, 71 pp
Campbell D (2005) The Congo River basin. In: Fraser LH, Keddy PA (eds) The world’s largest wetlands: ecology and conservation. Cambridge University Press, Cambridge, pp 149–164
Coynel A, Seyler P, Etcheber H, Meybeck M, Orange D (2005) Spatial and seasonal dynamics of total suspended sediment and organic carbon species in the Congo River. Global Biogeochem Cycles 19:1–17
Figueiredo BRS, Mormul RP, Thomaz SM (2015) Swimming and hiding regardless of the habitat: prey fish do not choose between a native and a non-native macrophyte species as a refuge. Hydrobiologia 746:285–290. doi:10.1007/s10750-014-2096-x
Filizola N (2003) Transfert sédimentaire actuel par les fleuves amazoniens. Thesis, University Paul Sabatier Toulouse III
Goulding M, Barthem R, Ferreira E (2003) The Smithsonian atlas of the Amazon. Smithsonian Institution Press, Washington
Higuti J, Martens K (2012a) On a new cypridopsine genus (Crustacea, Ostracoda, Cyprididae) from the Upper Paraná River Floodplain (Brazil). Zootaxa 3391:23–38
Higuti J, Martens K (2012b) Description of a new genus and species of Candonopsini (Crustacea, Ostracoda, Candoninae) from the alluvial valley of the Upper Paraná River (Brazil, South America). Eur J Taxon 33:1–31. doi:10.5852/ejt.2012.33
Higuti J, Martens K (2014) Five new species of Candoninae (Crustacea, Ostracoda) from the alluvial valley of the Upper Paraná River (Brazil, South America). Eur J Taxon 106:1–36. doi:10.5852/ejt.2014.106
Higuti J, Velho LFM, Lansac-Tôha FA, Martens K (2007) Pleuston communities are buffered from regional flood pulses: the example of ostracods in the Paraná River floodplain, Brazil. Freshw Biol 52:1930–1943. doi:10.1111/j.1365-2427.2007.01821.x
Higuti J, Meisch C, Martens K (2009) On Paranacypris samambaiensis n.gen. n.sp. (Crustacea, Ostracoda), the first South American psychrodromid, from the alluvial valley of the Upper Paraná River, Brazil. J Nat Hist 43:769–783. doi:10.1080/00222930802702506
Higuti J, Declerck SAJ, Lansac-Tôha FA, Velho LFM, Martens K (2010) Variation in ostracod (Crustacea, Ostracoda) communities in the alluvial valley of the upper Paraná River (Brazil) in relation to substrate. Hydrobiologia 644:261–278. doi:10.1007/s10750-010-0122-1
Higuti J, Schön I, Audenaert L, Martens K (2013) On the Strandesia obtusata/elliptica - lineage (Crustacea, Ostracoda, Cyprididae) in the aluvial valley of the upper Paraná River (Brazil), with the description of three new species. Crustaceana 86:182–211. doi:10.1163/15685403-00003160
Horne DJ, Martens K (1998) An assessment of the importance of resting eggs for the evolutionary success of non-marine Ostracoda (Crustacea). In: Brendonck L, De Meester L, Hairston N (eds) Evolutionary and ecological aspects of crustacean diapause. Adv Limnol 52:549-561
Irion G, Junk WJ, Mello JASN (1997) The large central Amazonian river floodplains near Manaus. Geological, climatological, hydrological and geomorphological aspects. In: Junk WJ (ed) The central Amazon floodplain: ecology of a pulsing system. Springer, Berlin, pp 23–46
Kobayashi JT, Thomaz SM, Pelicice FM (2008) Phosphorus as a limiting factor for Eichhornia crassipes growth in the upper Paraná River floodplain. Wetlands 28:905–913
Liberto R, Mesquita-Joanes F, César I (2012) Dynamics of pleustonic ostracod populations in small ponds on the Island of Martín García (Río de la Plata, Argentina). Hydrobiologia 688:47–61. doi:10.1007/s10750-011-0600-0
Mahé G (1993) Modulation annuelle et fluctuations interannuelles des précipitations sur le bassin versant du Congo. In: Boulègue J, Olivry JC (eds) Grands Bassins Fluviaux Périatlantiques: Congo, Niger, Amazone. ORSTOM, Paris, pp 13–26
Martens K (1984) Annotated checklist of non-marine ostracods (Crustacea, Ostracoda) from African inland waters. Koninklijk Museum voor Midden-Afrika, Tervuren-Belgie. Zool Dok 20, 51 pp
Martens K (1998) Diversity and endemicity of recent non-marine ostracods (Crustacea, Ostracoda) from Africa and South America: a faunal comparison. Verh Internat Verein Limnol 26:2093–2097
Martens K, Behen F (1994) A checklist of the non-marine ostracods (Crustacea, Ostracoda) from the inland waters of South America and adjacent islands. Trav Sci Mus Hist Nat Luxemb 22:1–81
Martens K, Schön I, Meisch C, Horne DJ (2008) Global biodiversity of non-marine Ostracoda (Crustacea). Hydrobiologia 595:185–193. doi:10.1007/s10750-007-9245-4
Matsuda JT, Lansac-Tôha FA, Martens K, Velho LFM, Mormul RP, Higuti J (2015) Association of body size and behavior of freshwater ostracods (Crustacea, Ostracoda) with aquatic macrophytes. Aquat Ecol 49:321–331. doi:10.1007/s10452-015-9527-2
Matzke-Karasz R, Nagler C, Hofmann S (2014) The ostracod springtail- camera recordings of a previously undescribed high-speed escape jump in the genus Tanycypris (Ostracoda, Cypridoidea). Crustaceana 87:1072–1094. doi:10.1163/15685403-00003343
Mazzini I, Ceschin S, Abati S, Gliozzi E, Piccari F, Rossi A (2014) Ostracod communities associated to aquatic macrophytes in an urban park in Rome, Italy. Int Rev Hydrobiol 99:425–434. doi:10.1002/iroh.201301728
McAleece N, Lambshead J, Patterson G, Gage J (1997) BioDiversity Pro, version 2. The Natural History Museum, London, and The Scottish Association of Marine Science, Oban, Scotland
McKenzie KG, Moroni A (1986) Man as an agent of crustacean passive dispersal via useful plants exemplified by Ostracoda Ospiti Esteri of the Italian ricefields ecosystem and implications arising therefrom. J Crustacean Biol 6:181–198
Meybeck M, Green P, Vörösmarty C (2001) A new typology for mountains and other relief classes: an application to global continental water resources and population distribution. Mt Res Dev 21:34–45
Mormul RP, Thomaz SM, Higuti J, Martens K (2010) Ostracod (Crustacea) colonization of one native and one non-native macrophyte species of Hydrocharitaceae: an experimental evaluation. Hydrobiologia 644:185–193. doi:10.1007/s10750-010-0112-3
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2013) vegan: Community Ecology Package. R package version 2.0-8. http://CRAN.R-project.org/package=vegan
Pieri V, Caserini C, Gomarasca S, Martens K, Giampaolo R (2007) Water quality and diversity of the Recent ostracod fauna in lowland springs from Lombardy (northern Italy). Hydrobiologia 585:79–87. doi:10.1007/s10750-007-0630-9
Pinto RL, Rocha CEF, Martens K (2003) On two new species of the genus Vestalenula Rossetti & Martens, 1998 (Crustacea, Ostracoda, Darwinulidae) from semi-terrestrial habitats in São Paulo State (Brazil). Zool J Linn Soc 139:305–313. doi:10.1046/j.1096-3642.2003.00070.x
Pinto RL, Rocha CEF, Martens K (2004) On the genus Penthesilenula Rossetti & Martens, 1998 (Crustacea, Ostracoda, Darwinulidae) from (semi-) terrestrial habitats in São Paulo State (Brazil), with the description of a new species. J Nat Hist 38:2567–2589. doi:10.1080/00222930310001647424
Pinto RL, Rocha CEF, Martens K (2005a) On the evolution of the genus Microdarwinula Danielopol, 1968 (Ostracoda, Darwinulidae) with the description of a new species from semi-terrestrial habitats in São Paulo State (Brazil). Crustaceana 78:975–986. doi:10.1163/156854005775197244
Pinto RL, Rocha CEF, Martens K (2005b) On new terrestrial ostracods (Crustacea, Ostracoda) from Brazil, primarily from São Paulo State. Zool J Linn Soc 145:145–173. doi:10.1111/j.1096-3642.2005.00185.x
Pinto RL, Rocha CEF, Martens K (2008) On the first terrestrial ostracod of the Superfamily Cytheroidea (Crustacea, Ostracoda): description of Intrepidocythere ibipora n. gen. n. sp. from forest leaf litter in São Paulo State, Brazil. Zootaxa 1828:29–42
Probst JL, Tardy Y (1987) Long-range streamflow and world continental runoff fluctuations since the beginning of this century. J Hydrol 94:289–311
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rodriguez LF (2006) Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biol Invasions 8:927–939. doi:10.1007/s10530-005-5103-3
Rossi V, Benassi G, Veneri M, Bellavere C, Menozzi P, Moroni A, McKenzie KG (2003) Ostracoda of the Italian ricefields thirty years on: new synthesis and hypothesis. J Limnol 62:1–8
Schultz R, Dibble E (2012) Effects of invasive macrophytes on freshwater fish and macroinvertebrate communities: the role of invasive plant traits. Hydrobiologia 684:1–14. doi:10.1007/s10750-011-0978-8
StatSoft, Inc. (2005) STATISTICA (data analysis software system), version 7.1. www.statsoft.com
Stiers I, Crohain N, Jones G, Triest L (2011) Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biol Invasions 13:2715–2726. doi:10.1007/s10530-011-9942-9
Szlauer-Lukaszewska A (2014) The dynamics of seasonal ostracod density in groyne fields of the Oder River (Poland). J Limnol 73:298–311. doi:10.4081/jlimnol.2014.865
Thomaz SM, Dibble ED, Evangelista LR, Higuti J, Bini LM (2008) Influence of aquatic macrophyte habitat complexity on invertebrate abundance and richness in tropical lagoons. Freshw Biol 53:358–367. doi:10.1111/j.1365-2427.2007.01898.x
Van der Meeren T, Almendinger JE, Ito E, Martens K (2010) The ecology of ostracodes (Ostracoda; Crustacea) in western Mongolia. Hydrobiologia 641:253–273. doi:10.1007/s10750-010-0089-y
Villamagna AM, Murphy BR (2010) Ecological and socio-economic impacts of invasive water hyacinth (Eichhornia crassipes): a review. Freshw Biol 55:282–298. doi:10.1111/j.1365-2427.2009.02294.x
Würdig NL, Pinto ID (2001) Pseudocandona pumilis sp. nov. (Ostracoda). Ecological data and distribution in the Tramandai lagunar system, RS, Southern Brazil. Nauplius 7:39–51
Acknowledgments
JH is grateful to Belgian Science Policy (BELSPO) for the postdoctoral scholarship in the Royal Belgian Institute of Natural Sciences (RBINS, Brussels, Belgium) as well as to the Global Taxonomic Initiative (GTI) for several grants that allowed taxonomic work in RBINS. KM gratefully acknowledges the International Office of the State University of Maringá (UEM) for a travel grant. We thank Ministry of Science and Technology (MCT)/National Council for Scientific and Technological Development (CNPq)/Fundação Araucária for funding the SISBIOTA (National System of Biodiversity Research) Project and BELSPO as well as other sponsors for funding the 2010 Boyekole Ebale Congo expedition and the Project COBAFISH. Dr. Fábio Amodêo Lansac Tôha [Nucleus of Research in Limnology, Ichthyology and Aquaculture (Nupelia), UEM, Brazil] is the Principle Investigator of the SISBIOTA project, while Dr. Luiz Felipe Machado Velho (Nupelia, UEM, Brazil) led the expedition to the Amazon in 2011. Dr. Erik Verheyen (RBINS, Brussels, Belgium) is coordinator of the COBAFISH project and also led the 2010 Boyekole Ebale Congo expedition. Dr. Papy Mongindo and Mr Ernest Tambwe (Kisangani, Congo) greatly assisted in collecting the Congolese samples. Dr. Roger Paulo Mormul and Jaime Luiz Lopes Pereira (Nupelia, UEM, Brazil) are thanked for statistical support and production of the map, respectively.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Higuti, J., Martens, K. Invasive South American floating plants are a successful substrate for native Central African pleuston. Biol Invasions 18, 1191–1201 (2016). https://doi.org/10.1007/s10530-016-1061-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-016-1061-1