
Abstract
Habitat selection is an important process in birds that influences individual survival and fitness, and ultimately shapes population dynamics. As a consequence, strong selective pressures apply to favor strategies allowing individuals to choose high-quality habitat for foraging while reducing predation risk and competition. In long-distance migratory birds, such as shorebirds, the non-breeding period is considered as a key period of their annual cycle with reported effects on individual survival and subsequent reproduction. Site selection by non-breeding shorebirds should depend on habitat quality for them to ensure survival until the next breeding season. More specifically, birds should distribute in space and time according to their resource availability and specialize on feeding habitats or/and prey to reduce intraspecific competition. To test this hypothesis, we studied Bar-tailed Godwits (Limosa lapponica) at one of their main non-breeding sites along the French coast. We first used GPS tracking to investigate their foraging and roosting home ranges as well as core sampling to define the diversity of available habitats and their quality as feeding resources. We then compared individual habitat selection in relation to feeding home range sizes and benthic macrofauna abundances. Our results provide the first fine scale definition of shorebird movements and distribution along the non-breeding period. We found that godwits showed an extreme fidelity to restricted feeding areas (about 3.4 km2) during winter, with low overlap between individual feeding home ranges. Each bird appeared to mainly use 4–6 of the 11 available foraging habitats, with a specialization on 1 to 2 main habitats and their associated prey. However, our results did not emphasize a clear relationship between home range sizes and the quality of bird feeding sites. This study thus demonstrates the high specialization of individual non-breeding Bar-tailed godwits relying on specific foraging grounds and habitats, with important implications for the conservation of this species.
Zusammenfassung
Stark diversifizierte Lebensräume und Ressourcen beeinflussen die Habitatwahl bei überwinternden Limikolen.
Die Habitatwahl ist ein wichtiger Prozess bei Vögeln, der das individuelle Überleben und die Lebensqualität beeinflusst und letztlich die Populationsdynamik prägt. Infolgedessen existiert ein starker Selektionsdruck, der Strategien begünstigt, die es den Individuen ermöglichen, qualitativ hochwertige Lebensräume für die Nahrungssuche zu wählen und gleichzeitig das Prädationsrisiko und die Konkurrenz zu reduzieren. Bei Langstreckenziehern, wie z. B. Limikolen, gilt die Nichtbrutzeit als eine Schlüsselperiode ihres Jahreszyklus, die sich auf das individuelle Überleben und die anschließende Reproduktion auswirkt. Die Standortwahl von nichtbrütenden Küstenvögeln sollte von der Habitatqualität abhängen, damit sie ihr Überleben bis zur nächsten Brutsaison sichern können. Genauer gesagt sollten sich die Vögel räumlich und zeitlich entsprechend ihrer Ressourcenverfügbarkeit verteilen und sich auf Nahrungshabitate oder/und Beute spezialisieren, um intraspezifische Konkurrenz zu reduzieren. Um diese Hypothese zu testen, untersuchten wir Uferschnepfen (Limosa lapponica) an einem ihrer wichtigsten Nichtbrutplätze an der französischen Küste. Zunächst untersuchten wir mit Hilfe von GPS-Ortungen ihre Nahrungs- und Schlafplatzbereiche sowie Bodenproben, um die Vielfalt der verfügbaren Habitate und deren Qualität als Nahrungsressourcen zu bestimmen. Anschließend verglichen wir die individuelle Habitatwahl in Abhängigkeit von der Größe des Nahrungshabitates und der Abundanz der benthischen Makrofauna. Unsere Ergebnisse liefern die erste feinräumige Darstellung der Ortsveränderungen einer Küstenvogelart und ihrer Verteilung außerhalb der Brurzeit. Wir fanden, dass Uferschnepfen während des Winters eine extreme Ortstreue zu begrenzten Nahrungsgebieten (ca. 3,4 km2) zeigten, mit geringer Überlappung zwischen den einzelnen Nahrungsgebieten. Jeder Vogel schien hauptsächlich vier bis sechs der 11 verfügbaren Nahrungshabitate zu nutzen, mit einer Spezialisierung auf ein bis zwei Haupthabitate und deren zugehörige Beute. Unsere Ergebnisse zeigten jedoch keinen eindeutigen Zusammenhang zwischen der Größe der genutzten Habitate und der Qualität der Nahrungsplätze der Vögel. Diese Studie zeigt die hohe Spezialisierung von Uferschnepfen außerhalb der Brutzeit auf bestimmte Nahrungsplätze und Habitate, was wichtige Folgerungen für den Schutz der Art hat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In spatially and temporally heterogeneous environments, habitat selection is a decision-making process where individuals are faced with choices not only resting upon habitat quality, but also on a trade-off between costs and benefits to acquire space for feeding (Fretwell and Lucas 1969; Kennedy and Gray 1994). Hence, the quality and availability of local habitats vary along with abiotic (e.g., feeding substrate), biotic (e.g., food resources, predators, parasites) and social (e.g., intraspecific competition) factors (Bruggeman et al. 2016; Philippe et al. 2016). Habitat selection ultimately influences the survival and reproduction of individuals (Hutto 1985; Block and Brennan 1993). Therefore, strong selective pressures apply to favor strategies that allow individuals to select high-quality habitats for foraging while avoiding predation and competition (Garabedian et al. 2019; Abdulwahab et al. 2019). The first step to study habitat selection is to define spatial habitat use, investigating animal movements and individual home ranges (van Moorter et al. 2016). The home range is defined as the area used by mobile animal during its regular activities (i.e., foraging, roosting, mating, caring for young; Burt 1943; Powell 2000). By affecting their physiology, energetics and behaviour, many factors are known to directly affect animal home ranges (size and shape; reviews in Mace et al. 1983; Rolando 2002; McGarigal et al. 2016), such as age, sex, body condition, habitat structure or weather conditions. Nonetheless, they are primarily driven by the distribution and availability of food resources (Brown 1975; Schoener 1983).
For long-distance migratory birds, such as many shorebird species, the non-breeding period is considered as a key period of their annual cycle with reported effects on individual fitness and population dynamics (Pienkowski and Evans 1984; Marra and Holmes 2001). Site selection by individuals during this period thus depends on habitat quality for them to ensure survival until the next breeding season (Evans 1976; Hutto 1985). At their coastal wintering sites, shorebirds are expected to face challenging weather conditions (Evans 1976; Clark 2009), high predation risks (van den Hout et al. 2008) and strong intraspecific competition (Beauchamp 2007). Most of them also have to deal with the characteristics of intertidal ecosystems where the tidal cycle reduces the time-window during which individuals can access their foraging grounds (Fonseca et al. 2017). In addition, birds must ensure their molt during this period, a particularly energy-demanding process (Murphy 1996). Consequently, wintering shorebirds have to balance their different energy gains and costs, especially the risk of starvation against the risk of predation (Evans 1976; Lima 1986), weighting the time spent in secured roosting sites against the time spent foraging in challenging feeding habitats (to acquire energy and maintain their body condition).
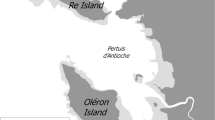
A large number of shorebirds winter along the French Coasts (Gaudard et al. 2018), and particularly in the Pertuis Charentais (Central Atlantic French Coast, Fig. 1), a region offering large areas of intertidal mudflats, essential feeding habitats for these species (Bocher et al. 2014; Robin et al. 2015). The Pertuis Charentais is the most important wintering area for shorebirds in France and is recognized as a site of international importance for these species (Delany et al. 2009). The Bar-tailed Godwit Limosa lapponica is one of them with a maximum of c. 4000 individuals recorded in January (F. Robin, com. Pers.). Two subspecies of Bar-tailed Godwit are present in western Europe but only L. l. lapponica overwinters in this region (Duijns et al. 2012), from the Netherlands and British Islands to the Iberian Peninsula, with a population estimated to 150,000 individuals (Wetlands International 2017). The species predominantly feeds on polychaetes whatever their wintering site in Europe during non-breeding period (Duijns et al. 2013). L. l. lapponica is characterized by stable population trends at the European scale. Nonetheless, important decreases of wintering populations were observed in France during 1980s and 1990s (Triplet et al. 2010), likely due to degraded wintering habitats and increasing human disturbance during this sensitive period (Verger 2005; Delany et al. 2009; BirdLife International 2017), when godwits are known to occupy a few small areas (Delany et al. 2009). The creation of Nature Reserves in the Pertuis Charentais (Fig. 1) highlighted positive effects on Bar-tailed Godwits by reducing disturbance at their high tide roosts, resulting in a significant increase in bird numbers over the 1985–2009 period on Ré Island, Yves Bay and Marennes–Oléron Bay, reaching national or international importance threshold (Triplet et al. 2010). In that context, understanding the spatial distribution of wintering Bar-tailed Godwits is essential to refine conservation measures (Morris 2003; Klar et al. 2008). Likewise, characterizing their habitat use is crucial, not only for a complete understanding of ecosystem functioning, but also to develop appropriate management strategies bound to protect this near-threatened species (BirdLife International 2017). In the present study, we tested the hypotheses that in a context of high bird densities, (1) female Bar-tailed Godwits specialize on feeding habitats and prey to avoid conspecific competition during the non-breeding period (Duijns and Piersma 2014), and (2) individuals using habitats with higher food abundances are predicted to have smaller home ranges. We first defined individual home ranges and more specifically feeding home ranges using tracking devices. Second, we determined available foraging habitats and benthic prey availability on accessible mudflats at low tide and their respective surfaces by in situ sampling. Finally, we compared individual habitat selection in relation to feeding home ranges size and prey abundance.

Map of the Pertuis Charentais (Central French Atlantic coast) and localisation of the study areas on Ré Island. Dark grey corresponds to the mainland and light grey to the intertidal area
Methods
Study site
The study was carried out on Ré Island (Central French Atlantic Coast) during non-breeding periods (from August to March) of 2015–2016 and 2016–2017. Ré Island is a part of the Pertuis Charentais area (Fig. 1), a region including the largest surface of intertidal mudflats on estuarine systems in France (Verger 2005). Two main feeding areas are available for Bar-tailed Godwits (hereafter godwits) on soft substrates on the Island (Aubouin 2014; Duijns et al. 2014). The first site is a mudflat inside a semi-enclosed bay, the ‘Fier d’Ars’ (46° 13′ 18′′ N; 1° 30′ 29′′), and the second site is sandflat on an exposed coast in the north of the Island, ‘La Loge’ (46° 14′ 25′′ N; 1° 28′ 42′′ W).
The ‘Fier d’Ars’ is a bay of about 800 ha bordered by 1600 ha of saltpans or brackish marshes. This wetland has been classified according to the Ramsar Convention since 2003, and part of it (the western part of the bay and the high tide roosts in saltmarshes) is part of the National Nature Reserve ‘Lilleau des Niges’ since 1980. The ‘Fier d’Ars’ is divided by a central creek, with a mainly bare muddy substrate on the edges or covered by seagrass beds (Zostera noltei) at the center, and a restricted sandy area along the stream to the north.
‘La Loge’ extends over c. 100 ha. The foreshore remains uncovered by the tide for about 4 h only, and the period of food availability is, therefore, limited for birds compared to the ‘Fier d’Ars’. ‘La Loge’ is part of the Ramsar site ‘Marsh of the Fier d’Ars’ as the only protection status, and tourist activities on the beach during summer are frequent during daylight.
Coastal shorebird survey data
Counts of shorebirds were carried out by the staff of the National Nature Reserve, year-round, around the 15th of each month on the whole Ré Island from 2000 to 2016. The census targeted all the known high tide roosts during the highest water levels during spring tide. Birds were counted with telescopes, simultaneously by five observers distributed on the main spots of Ré Island, from 2 h before and until the high tide.
Godwit captures and tracking
Godwits were captured using mist nets on high tide roosts between October 2015 and November 2016. Catching sessions took place during non-moonlight nights inside the National Nature Reserve. Individuals were marked with a metal ring and a unique color rings combination. The sex of individuals was determined based on their body mass (230–383 g for males and 280–455 g for females) and bill length (69–90 mm for males and 86–110 mm for females; Demongin 2016); and the age defined according to their plumage pattern (Demongin 2016).
A total of 15 godwits (11 in 2015 and 4 in 2016) were fitted with a GPS-VHF logger (STERNA UHF-SRD with solar charger, Ecotone, Gdánsk, Poland; 35 × 16 × 10 mm, 7.5 g). Birds were chosen so that the mass of the logger was less than 3% of the bird body mass. Consequently, only adult females (bill length > 86 mm) were selected for the study, considering that males were too small and that juvenile females were possibly not faithful to the study site during winter. Loggers were attached on the lower part of the back with a 2 mm Teflon harness according to the ‘leg-loop’ method (Mallory and Gilbert 2008). Tags recorded GPS locations (tested mean accuracy of ± 10 m) every 30 min. Out of the 15 females tagged, 6 recorded a sufficient number of positions during all the winter period. Among the others, four birds never contacted after the logger deployment and probably moved outside of the area, four birds had a tag which progressively stopped working because of misfunctioning in battery recharging and gave less than 300 locations in total over a short period of the winter, and one bird was found predated shortly after release. Among the six females, three (BTG01, BTG02 and BTG03) recorded locations for two consecutive winters (2015–2016 and 2016–2017). The three others birds provided only one winter of data in 2015–2016 (BTG04 and BTG05) or 2016–2017 (BTG06).
Prey availability and habitat description
The availability and quality of trophic resources at the feeding areas used by tagged godwits (distribution, density and biomass of benthic macrofauna) were investigated by sediment core sampling. Sampling was performed at the centroid of each predefined individual Feeding Core Area (hereafter FCA), based on GPS locations recorded over the first 2 months of deployment. Indeed, to determine the benthic macrofauna potentially available for birds during winter, the sampling of sediment was carried out during the course of the winter. Given the high fidelity of godwits to foraging sites, these sampling stations based on birds’ FCA after 2 months were considered to be representative of the entire winter. The number of core sampling stations per bird ranged from two to nine, depending on the size of its FCA and the number of core areas which composed it, to apply the same sampling pressure per feeding surface between the different birds. These samples were collected between January and February, by foot and at low tide according to methods described in Bocher et al. (2007) and Kraan et al. (2007). At each point, a 15 cm diameter sediment core (0.018 m2) was collected at a depth of 15 cm (depth of sediment containing almost all polychaetes available for foraging birds according to mean bill length of females). These samples were sieved over a 1-mm mesh size on site. Annelids were separated and preserved in 70 °C ethanol. Molluscs were stored at − 20 °C until sorted at the laboratory. Although mud snails Hydrobia ulvae are rare in the godwits diet, they were sampled by taking one additional core (70 mm diameter) of 0.0037 m2 to a depth of 5 cm and sieved over a 0.5 mm mesh, to differentiate between present and available abundance. Mud snails were a potentially abundant and small prey that would require too long processing time for a larger sediment core sampling. In the laboratory, all organisms were identified to the species level, if possible, and the molluscs were measured to an accuracy of 0.1 mm. The Dry Mass (DM) of flesh for each prey was measured after 72 h of drying at 55 °C. Dried specimens were then incinerated at 550 °C for 5 h to determine their ash mass and then a proxy of their energy content: the ash free dry mass (AFDM; Zwarts and Wanink 1993). For annelids damaged or cut during sieving, anterior part of the body, when remaining, was measured and DM and AFDM estimates were determined from allometric relationships specific to each species and established in a related study (Bocher et al., in prep.).
A sediment sample was collected to a maximum depth of 5 cm for granulometric characteristics. Medium Grain Size (mm) and the percentage of silt (fraction < 0.063 mm) were determined using a Malvern Mastersizer 2000 diffraction laser (particle sizes analyzed from 0.04 to 2000 mm). Results on granulometric characteristics of sediment were used as complement data to define the habitat type at each sampling station and to draw the map of habitats available to godwits.
A fine scale cartography of intertidal habitats was drawn by combining benthic macrofauna and sediment sample results with field observations, and using the EUNIS classification of coastal habitats as a reference Bajjouk et al. (2015). Seagrass beds delimitations were achieved during summer 2012 on field with GPS (Trimble GeoXH).
Home ranges and habitat selection
Space use during the winter period was analyzed through the estimation of the Utilization Distribution (UD), i.e., the probability density to found each bird at any place according to the location of this place (Calenge 2015). The UDs were computed with the Kernel density estimates (KDE; van Winkle 1975; Worton 1989) using the function ‘kernelUD’ (package ‘adehabitatHR’) on the statistical software R (3.6.1; Calenge 2020). KDE takes into account the spatial distribution of locations, and corresponding time spent by birds in an area, to estimate, at the individual scale, their home range (at 95% isopleth) and core area (at 50% isopleth; Worton 1989). We used a fixed-kernel method with a smoothing factor of 70% the minimum reference value computed by the ‘ad-hoc’ bandwidth over all individuals, a compromise between under-smoothing of least-square cross-validation (LSCV) and over-smoothing of reference bandwidth (Kie et al. 2010; Schuler et al. 2014). Using a single value of smoothing factor allowed us to compare individuals. Cell size was set to 20 m considering the accuracy of GPS positions.
As the number of recorded positions differed between birds, we checked that data quantity had no effect on the estimation of home ranges size. To this end, we calculated wintering home range size (95% isopleth) of each individual using the kernel method for position samples successively increased by 50 locations chronologically, from n = 50 to n = max number of positions collected (Supplementary material Fig. S1). For most of birds, the size of the home range showed a significant increase until c. 1000 GPS fixes, beyond, it no longer changed significantly and then reached a plateau. This confirmed that the number of GPS fixes recorded was sufficient for all individuals to provide reliable estimates of home ranges size.
The UDs were analyzed at the global frame for each bird to estimate the individual home range (HR) and core area (CA) using all GPS locations. Then, we calculated separately UDs on feeding positions (i.e., when birds were foraging on intertidal areas at low tide, below a water height of 3.3 m relative to the hydrographic zero) and roosting positions (i.e., when birds were roosting on salt marshes, upper intertidal areas or pond when the water height exceeded 3.3 m). We thus estimated the complete home range (HR, 95% kernel density contour) and core area (50% kernel density contour) during Feeding (FHR and FCA) and during roosting (RHR and RCA). For the three birds for which two consecutive wintering periods were recorded, we investigated the inter-annual fidelity to their wintering feeding areas. The high fidelity observed in these three individuals allowed us to assume the absence of a ‘year’ effect on the feeding UDs, and thus to compare the six individuals by retaining the winter 2015–2016 data for BTG01, BTG02, BTG03, BTG04, BTG05, and winter 2016–2017 data for BTG06.
In another study on Ré Island (Bocher et al., in prep.), 64 droppings of godwits were sorted of and there were found almost exclusively remains from polychaetes, and very anecdotally shell fragments of molluscs. Hence, considering that godwits predominantly feed on polychaetes whatever their wintering sites in Europe (Duijns et al. 2013), including Ré Island (Bocher et al., in prep.), we tested the hypothesis that the size of individual FCA was negatively related to the abundance of polychaete biomass (excluding small species with individual biomass < 0.001 g AFDM) inside the FCA. To investigate the use and importance of foraging habitats, a habitat selection analysis was then performed with the R package ‘adehabitatHS’ (Calenge 2011). To this end, we considered the estimated Minimum Convex Polygon (MCP) of foraging locations from all equipped godwits to define the available foraging areas. Habitat selection analysis was performed overlaying the FCA and the habitat typology using the method of selection ratios (Manly et al. 2002) for design II (second-order selection), where the availability of habitats is the same for all six monitored birds and the habitat selection is analyzed at the individual level (Johnson 1980). To explain variations in habitat selection among individuals, we used an Eigen analysis of selection ratios (Calenge and Dufour 2006), an extension of principal component analysis were birds are projected in the factorial plane describe by the habitat types and which best explains the heterogeneity of the selection. This multifactorial method is suitable for the exploration of habitat selection and identification of groups of animals using habitats in a same or different way.
Results
Phenology and abundance of godwits
The wintering period at the study site spanned over a maximum of 8 months from August to March, with a maximum number of birds recorded in January with a mean of 641 ± 85 individuals over 2000–2016 (Supplementary material Fig. S2). The last birds arrived in late-October/early-November and most of the birds left the site for pre-nuptial migration during March. Very few birds were present from April to July, i.e., during the migration period.
Home range variability
HR of the six birds over the two winters of study varied between 248 and 581 ha (mean = 461 ± 48 ha, n = 6). Despite a different number of total recorded locations, the size evolution of the HR of most of birds showed a plateau beyond which the additional locations bring no significant variation (Table 1, Supplementary material Fig. S1).
Inter-annual fidelity to feeding areas
For the three birds displaying two successive non-breeding periods, FHR overlapped by 83%, 74% and 71% between both winters (Supplementary material Table S1), while the average inter-individual overlap of FHR was 43% (Supplementary material Table S2). Individual BTG01 had the largest FHR (478 ha in 2015–2016; 408 ha in 2016–2017) and used both winters a large part of the bay with multiple FCA (Fig. 2). BTG02 foraged mainly in the ‘Fier d’Ars’, with only a few visits on ‘La Loge’. The difference in FHR between winters for BTG02 (363 ha in 2015–2016; 232 ha in 2016–2017) was explained by the sporadic visit of a supplementary area during the first winter located outside the study site (‘Fossé de Loix’ not represented on Fig. 2). The FHR of BTG03 during both periods were very similar (287 ha in 2015–2016; 297 ha in 2016–2017). The estimated size of FCA between both winters was also close for BTG01 (69 ha in 2015–2016; 73 ha in 2016–2017), BTG02 (22 ha in 2015–2016; 34 ha in 2016–2017) and BTG03 (51 ha in 2015–2016; 49 ha in 2016–2017), but their location slightly changed with an overlap of 55%, 16% and 63%, respectively.

Feeding home range (FHR) of three Bar-tailed Godwits during two successive wintering periods on Ré Island. Home ranges were calculated as 50% (core area) and 95% (Home Range) kernel density contours
Feeding and roosting home ranges
For each bird, the number of GPS fixes at roost and on feeding areas was similar, allowing us to compare the size of FHR and RHR (Table 1). The number of GPS locations between birds was not comparable both for roosting and for feeding fixes, but our dataset is large enough not to bias the estimation of the utilization distributions between birds (see above). The mean size of RHR (kde 95%) and RCA (kde 50%) were 211 ± 35 ha (min–max 161–250 ha) and 33 ± 12 ha (min–max 19–48 ha), respectively (Fig. 3). The birds used between six and ten different roosts all around the bay according to individual. During neap tide, they remained in the upper part of the intertidal area but during spring tide, they only roosted in marshes within the Nature Reserve or in saltpans. The mean size (± SE) of FHR (kde 95%) and FCA (kde 50%) were 340 ± 105 ha (min–max 167–478 ha) and 43 ± 21 ha (min–max 13–69 ha), respectively (Fig. 2). The FHR and RHR overlapped only on the upper part of the intertidal area during neap tide, particularly on an elevated sandflat in front of the Nature Reserve.

Feeding home range (FHR) and roosting home range (RHR) of six Bar-tailed Godwits monitored during the non-breeding period on Ré Island. Home ranges were calculated as 50% (core area) and 95% (home range) kernel density contours
Rhythm of feeding activity
According to GPS fixes located on potential feeding areas, godwits fed during approximately 6–7 h by tide cycle (Fig. 4). The presence time of birds on the mudflats was not symmetrical around low tide, with a feeding period of 2–3 h before low tide and 3–4 h after. Beyond this general pattern, the duration of use of the mudflats seems to differ between individuals, with an arrival of birds on the feeding areas generally more synchronous (between − 4 and − 2 h in tidal cycle) than the return towards roosting places (between 2 and 5 h in tidal cycle). BTG05 spent less time than other birds on feeding areas, with a maximum value of 80–85% of its time on intertidal areas during the low tide.

Proportion of time spent by six Bar-tailed Godwits on mudflats, i.e., feeding, during the tidal cycle
Prey availability and feeding home ranges
A total of 19 stations located in FCA were retained to describe habitat quality within the FHR (between two and nine stations per FCA per bird). A total of 16 bivalve, five gastropod, 32 annelid and 1 nemert species or taxa were identified for all sampled stations (Table S4). Three bivalve species were largely abundant: Cerastoderma edule (mean minimal and maximal densities for individual FCA: 98–274 ind m−2), Ruditapes spp. (12–62 ind m−2) and Scrobicularia plana (0–154 ind m−2); but differed between habitats. Among gastropods, only Hydrobia ulvae was largely abundant (22–468 ind m−2). Among annelids, the most abundant species were Capitellidae sp. (0–180 ind m−2), Cirriformia tentaculata (0–25 ind m−2), Hediste diversicolor (0–117 ind m−2), Nephtys hombergii (11–41 ind m−2), Notomastus latericeus (2–82 ind m−2), Owenia fusiformis (8–46 ind m−2), Scoloplos armiger (0–462 ind m−2) and Arenicola marina (2–10 ind m−2). The size of bird FCA was not related to the mean abundance of polychaete biomass inside predefined core areas (t = 0.80, df = 4, p value = 0.47, Fig. 5).

Size of individual feeding core area (FCA—core area) as a function of the mean polychaete biomass at each individual core area
Foraging habitat selection
Eleven habitats were identified and delimited on the intertidal areas of both study areas, with marked differences between ‘La Loge’ and ‘Fier d’Ars’ (Fig. 6). On ‘La Loge’, two habitats of intertidal muddy sand (A2.242 and A2.231; EUNIS Typology) and one habitat of barren littoral coarse sand (A2.221) were described. Outside the ‘Fier d’Ars’, a last type of intertidal habitat, mainly composed of rocky elements, was represented on the area of ‘La Loge’ composed by littoral rockpool communities: A1.41 (Fig. 6a). The ‘Fier d’Ars’ was muddy prevailing facies, largely dominated by seagrass beds on muddy sand (A2.6111) and oyster parks on muddy foreshore (A2.32). The rest of intertidal areas on ‘Fier d’Ars’ was characterized by bare muddy sand habitats (A2.242 and A2.313) and rocky habitats (A1.11 and A1.4; Fig. 6b).

Habitat typology at a Ré Island, b ‘La Loge’ and c the ‘Fier d’Ars’
There was a strong habitat selection by the six godwits (χ2 = 86,441.52, df = 54.0, p < 0.001) and habitat selection was not identical among all individuals (χ2 = 9618.85, df = 45.0, p < 0.001; Supplementary material Table S3). Three habitats were preferentially used by females: seagrass beds in muddy sand (A2.6111) dominated by polychaetes, oligochaetes and molluscs; sandflat dominated by C. edule and polychaetes (A2.242); and Mudflat dominated by H. diversicolor and S. plana (A2.313; Global Selection Ratios > 1; Fig. 7, Supplementary material Fig. S3). Conversely, the habitats of muddy sand dominated by polychaetes Nephtys cirrosa and S. armiger (A2.231), fucoids on sheltered marine shores (A1.31) and oyster parks (A2.32) were globally avoided (Global Selection Ratios < 1), but confidence intervals indicate their marginal use by some few birds (Fig. 7, Supplementary material Fig. S3). The four remaining habitats were clearly avoided (Global Selection Ratios and IC < 1).

Proportion of habitat types in feeding core area (FCA) of each Bar-tailed Godwit. The FCA estimated through the kernel method, was crossed with the typology of habitats, and give the surface proportion (%) of each habitat in this main spatial delimitation of foraging. This description of represented habitats inside the FCA is a proxy of the foraging habitat selection by each godwit
The Eigen analysis highlighted that two factors mainly explained the habitat selection, with 90.2% of information explained by the first axis and 6.2% by the second one. The results show that birds much more frequently used habitats A2.6111, A2.242 and A2.313. BTG01, BTG03 and BTG06 used preferentially the seagrass beds (A2.6111; Fig. 8), BTG04 favored sandflat (A2.242), and BTG02 and BTG05 did not selected one specific habitat but combined the use of A2.611, A2.242 and A2.313 (intertidal muddy sand dominated by H. diversicolor, M. balthica and S. plana). Without highlighting groups of birds, the Eigen analysis underlies a variability in habitat selection between individuals, with a ‘continuum’ along the second axis between habitats A2.6111, A2.242 and A2.313. Differences in the proportion of each habitat in the FCA confirmed a specific pattern of habitats combination for each individual, all-different from each other (Fig. 7).

Results of the Eigen analysis of selection ratios highlighting habitat selection by six Bar-tailed Godwits on 11 habitat types. a Habitat types projected along the first two factorial axes and b bird scores on the first factorial plane. The analysis produced two factors (the two axis) to explain the heterogeneity of habitat selection. These two factors constitute the first factorial plane on which the habitats, which compose it, are projected, and birds are positioned in this plane described by habitats
Discussion
This study on Bar-tailed Godwits, enabled by GPS-tracking technology recently adapted to medium size shorebirds, highlighted the individual use of foraging patches during the almost entire non-breeding period, sometimes consistent from year to year, and showing a specific combination of habitats used among all suitable ones. Although our analysis was based on six birds, thus calling for further research to strengthen our conclusions, it underlines the high fidelity of some individuals of godwits to their feeding grounds during the non-breeding period. Indeed, on a small wintering site, tracked godwits were restricted to small foraging areas and did not exploit all suitable areas. In addition, at least three of the six monitored birds used the wintering site for two consecutive year, and with a high fidelity for their respective foraging home ranges. It also demonstrates a specific combination of foraging habitats used by each bird monitored, supporting the idea of a specialization on preys at the individual scale (Sutherland et al. 1996; Durell 2000). Previous studies showed that during winter, godwits feed predominantly on worms, wherever their distribution along the European costs (Evans 1976; Scheiffarth 2001; Duijns et al. 2013). On Ré Island, other studies confirmed a diet largely composed of worms, whether on the site of ‘La Loge’ or ‘Fier d'Ars’ (Aubouin 2014; Bocher et al. 2014; Duijns et al. 2014). Consequently, the spatial distribution of birds on mudflats could be linked to a specialization on feeding habitats, and more precisely on particular species of annelids, the largely dominant prey in their diet.
Adult godwits were overwintering on Ré Island between August and March. The peak number of birds at the study site was observed in mid-January, with a mean of 641 ± 85 individuals (over the period 2000–2016). Considering a total feeding functional area estimated to 900 ha on Ré Island (Aubouin 2014), the density of godwits on the island is then estimated to ca. 0.7 ind ha−1 in the middle of winter. This density is the highest (by 2–7 times) of the four main wintering sites located inside the Pertuis Charentais area: Aiguillon Bay (0.3 ind ha−1), Yves Bay (0.1 ind ha−1) and Marennes–Oléron Bay (0.3 ind ha−1). The mean godwit densities measured in the Dutch sectors of Wadden Sea was estimated to 9.5 ± 1.5 ind ha−1 in May (Duijns and Piersma 2014) and around 1.5–4.5 ind ha−1 between July and September (Folmer et al. 2010, van den Hout and Piersma 2013). During these pre- and post-migration periods, both subspecies are present and denser than during the winter period. Therefore, although the carrying capacity appears lower than observed at other times of the year in the Wadden Sea, the site of Ré Island seems to offer habitats of good quality for godwits at the local scale. More specifically, despite the restricted surface of sand and mudflat areas, quality and availability of trophic resources on Ré Island appears high enough to allow high bird densities. The observed fidelity of individuals to their feeding grounds on such a small site with a patchwork of diversified habitats suggests that birds specialize in prey species to maintain sufficient energy intake rates and ensure their survival (Catry et al. 2014; Ceia and Ramos 2015). Indeed, such a specialization could reduce intraspecific competition and restrict individuals to specific FCA, explaining the low overlap observed (48%) between our tagged birds (Kouba et al. 2017). A higher number of birds monitored would most likely have resulted in an increased overlap between individual FHR. This is supported by the density of birds observed on site during the winter, especially on ‘La Loge’ were godwits are usually observed in flocks, unlike in the ‘Fier d’Ars’ (P. Bocher, com. Pers.). Nevertheless, the distinct feeding distribution observed on six birds using the same roosts confirms the non-gregarious pattern of foraging godwits, at least inside the ‘Fier d’Ars’.
In addition to use the same restricted feeding area during the non-breeding period, godwits which provided locations for two consecutive winters (n = 3) showed a high inter-annual overlap in their FHR. Previous studies have reported fidelity of shorebirds to their non-breeding site but only a few highlighted faithfulness to specific feeding and roosting sites, both within and between winters (Conklin and Battley 2011; Coleman and Milton 2012). Based on GPS telemetry, our study help to fill this gap at the patch scale. Indeed, in this study, we show a high fidelity of godwits to their roosting and feeding home ranges, with a less marked but still important inter-annual fidelity to feeding core areas. This highlights the abilities of godwits to specialize on particular set of foraging habitats, and calls for future research studies to explore the high degree of individual specialization on preys, all along their life in long-lived species such as shorebirds.
Our results also highlight that the surface of wintering home ranges for godwits wintering on Ré Island was small (< 600 ha) in comparison to others shorebirds species as Dunlin Calidris alpina and Red Knot Calidris canutus. For instance, the home range of dunlin ranged from 1080 to 56,470 ha according to the study area (Sanzenbacher and Haig 2002; Shepherd and Lank 2004; Taft et al. 2008; Choi et al. 2014) and the home range of red knot could range from 1000 to 80,000 ha (Piersma et al. 1993; Leyrer et al. 2006). This large home range size variability among dunlin and red knot are due to differences in habitats and feeding conditions (Piersma et al. 1993; Choi et al. 2014). Nevertheless, these species foraged in large flocks and birds moved according to group decision from one place to another (van Gils et al. 2015). Among close related species of godwit, the feeding home range of the Marbled Godwit Limosa feoda was estimated to 672 ha on Larnack Reef, a stable island near a highly dynamic intertidal area (Gulf of Mexico; Gabbard et al. 2001). In this study, the home range estimation was produced with convex polygons, which differ from our method (kernel home range), but the order of magnitude is more similar to our results. FHR of godwits wintering on Ré Island thus appeared spatially restricted, reinforcing the idea of bird fidelity to their foraging areas and a supposed specialization on foraging habitats and prey species at the individual scale at Ré Island.
Estimation of FHR also showed a strong variability between individuals with low overlaps highlighting distinct areas prospected by godwits, while RHR showed a large mean inter-individual overlap (69%). Roost sites located at Western side of the study site mainly corresponded to former saltpans inside the Nature Reserve (‘Lilleau des Niges’), known to hold most of the shorebirds at spring high tide (J-C. Lemesle, com. Pers.). Roosting areas located at the Eastern side are not protected, and birds no longer use them during high water levels of spring tide. The alternating use of these roosts likely depends on the accessibility during the tidal cycle, as the roosts at Eastern side are located closer to the foraging sites. The short distances between roosting and feeding grounds could be one of the main factors driving the spatial distribution of wintering shorebirds (Morrison and Harrington 1979), reducing flight distance and associated loss of energy and predator exposure (Si et al. 2011; Novaes and Cintra 2013). Our results suggest similar processes in godwits on Ré Island. Feeding patterns identified in our study confirm that godwits start to feed between two and three hours after high tide, present on feeding areas for four to six hours before to join roosting areas (Lindisfarne, Northumberland—England; Smith and Evans 1973). Despite spatially restricted home ranges, godwits devote almost all of their time for foraging (> 85%) during the complete access to mudflat, both day and night. This result suggests that birds have to optimize prey collection throughout their accessibility to achieve a sufficient intake rate and satisfy their energy needs.
Beyond the time spent foraging and according to the optimal foraging theory, godwits are predicted to optimize their prey intake rate by selecting specific feeding habitats of higher quality (Evans 1976). Godwits preferentially used 3 of the 11 identified habitats in our study site. Three birds preferentially selected habitat associated with eelgrass beds (A2.6111) where annelid abundance is high. Habitat composed mainly of C. edule and polychaetes (A2.242), and habitat composed mainly of H. diversicolor, M. balthica and S. plana (A2.313), were also selected by most of the birds. The inter-individual variability observed in habitat selection reinforces the hypothesis that individuals may specialize in particular polychaete prey species to limit competition (Durell 2000) and/or optimize intake rate by experience on catching abilities on particular prey species (Bolnick et al. 2003). Selection of specific prey or substrate could also explain the differences observed in the size of feeding core areas. However, and conversely to our hypothesis, our results did not show a significant negative relationship between the biomass of polychaetes and the size of bird feeding areas. An alternative hypothesis is that polychaete intake rates could not directly reflect worm abundance but rather the availability and catchability of certain species in the sediment. Thus, it could be more difficult for birds to extract polychaetes in some habitats due to the worm depth or the substrate hardness (Finn et al. 2008), or due to prey behavior (Duijns and Piersma 2014), which would lead to an increased foraging and prospecting effort to achieve sufficient intake rates to fulfill bird daily energy requirements.
This study was carried out on female adults only, and results cannot be extrapolated to males. Indeed, a large sexual dimorphism exists in godwits with females being on average 1.25 times larger than males, leading to contrasting foraging strategies (Pierre 1994; Duijns et al. 2014). Similarly, juveniles show a lack of experience when arriving for the first time on unknown wintering site which might affect their foraging behavior and habitat use (Durell 2000; van den Hout et al. 2014). Further investigations performed on different stages and including both sexes will thus be required when the miniaturization of GPS data logger will be improved, to have a complete understanding of wintering strategies in this near-threatened species (BirdLife International 2017).
Bird specialization on particular prey or habitats implies knowledge of their distribution and the way to exploit them (Bolnick et al. 2003). Individuals can optimize their movement and maximize their foraging gain ratio. Thus, important changes in the quality and distribution of trophic resources, under the effect of anthropogenic disturbances, could modify bird energy intakes and jeopardize their winter survival and subsequent migration and reproduction (Gunnarsson et al. 2006). In a context of rapid environmental modifications of coastal habitats (Ivajnšič et al. 2018; Leo et al. 2019), expected degradations could result in an increased competition for the remaining resources. Consequently, any increase in bird density may have a marked effect on these habitats and individuals with specialized diet and feeding methods will be the first impacted, particularly if they are of lower social status (Durell 2000). This study thus confirms the importance of maintaining a good quality of wintering habitats, especially on intertidal habitats facing high human pressures such as professional or recreative on-foot fishing, roost-disturbing hunting activities, nautical activities and tourism.
References
Abdulwahab UA, Osinubi ST, Abalaka J (2019) Risk of predation: a critical force driving habitat quality perception and foraging behavior of granivorous birds in a Nigerian forest reserve. Avian Res 10:33. https://doi.org/10.1186/s40657-019-0173-6
Aubouin N (2014) Etude de la distribution et des stratégies de survie hivernale de populations migratrices d’oiseaux limicoles en vue d’actions de conservation en région Poitou Charentes: exemple de la barge rousse Limosa lapponica. Msc Thesis, Université de Montpellier, p 64
Bajjouk T, Guillaumont B, Michez N et al (2015) Classification EUNIS, système d’information Européen sur la nature : traduction française des habitats benthiques des régions Atlantique et Méditerranée. vol. 1, Habitats Littoraux, IFREMER/DYNECO/AG/15‐02/TB1. https://archimer.ifremer.fr/doc/00271/38222/
Beauchamp G (2007) Competition in foraging flocks of migrating semipalmated sandpipers. Oecologia 154:403–409. https://doi.org/10.1007/s00442-007-0818-8
BirdLife International (2017) Limosa lapponica (amended version of 2016 assessment). The IUCN Red List Threat Species. https://doi.org/10.2305/IUCN.UK.2017-1.RLTS.T22693158A111221714.en
Block WM, Brennan LA (1993) The habitat concept in ornithology: theory and applications. In: Power DM (ed) Current ornithology. Springer, Boston, pp 35–91
Bocher P, Piersma T, Dekinga A et al (2007) Site-and species-specific distribution patterns of molluscs at five intertidal soft-sediment areas in Northwest Europe during a single winter. Mar Biol 151:577–594. https://doi.org/10.1007/s00227-006-0500-4
Bocher P, Robin F, Kojadinovic J et al (2014) Trophic resource partitioning within a shorebird community feeding on intertidal mudflat habitats. J Sea Res 92:115–124. https://doi.org/10.1016/j.seares.2014.02.011
Bolnick DI, Svanbäck R, Fordyce JA et al (2003) The ecology of individuals: incidence and implications of individual specialization. Am Nat 161:1–28. https://doi.org/10.1086/343878
Brown JL (1975) The evolution of behavior. W.W Norton and Co, New York
Bruggeman JE, Swem T, Andersen DE et al (2016) Multi-season occupancy models identify biotic and abiotic factors influencing a recovering Arctic peregrine falcon Falco peregrinus tundrius population. Ibis 158:61–74. https://doi.org/10.1111/ibi.12313
Burt WH (1943) Territoriality and home range concepts as applied to mammals. J Mamm 24:346. https://doi.org/10.2307/1374834
Calenge C (2011) Exploratory analysis of the habitat selection by the wildlife in R: the adehabitatHS package. Office national de la chasse et de la faune sauvage, Saint Benoist 78610, Auffargis, France. Available at: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.473.208&rep=rep1&type=pdf
Calenge C (2015) Home range estimation in R: the adehabitatHR package. Office national de la classe et de la faune sauvage Saint Benoist, Auffargis
Calenge C (2020) Package ‘adehabitatHR’. R package version 0.4.18. https://cran.r-project.org/web/packages/adehabitatHR/adehabitatHR.pdf. Accessed 21 Jun 2020
Calenge C, Dufour AB (2006) Eigenanalysis of selection ratios from animal radio-tracking data. Ecology 87(9):2349–2355. https://doi.org/10.1890/0012-9658(2006)87[2349:EOSRFA]2.0.CO;2
Catry T, Alves JA, Gill JA et al (2014) Individual specialization in a shorebird population with narrow foraging niche. Acta Oecol 56:56–65. https://doi.org/10.1016/j.actao.2014.03.001
Ceia FR, Ramos JA (2015) Individual specialization in the foraging and feeding strategies of seabirds: a review. Mar Biol 162:1923–1938. https://doi.org/10.1007/s00227-015-2735-4
Choi C, Gan X, Hua N et al (2014) The habitat use and home range analysis of dunlin (Calidris alpina) in Chongming Dongtan, China and their conservation implications. Wetlands 34:255–266. https://doi.org/10.1007/s13157-013-0450-9
Clark JA (2009) Selective mortality of waders during severe weather. Bird Study 56:96–102. https://doi.org/10.1080/00063650802648465
Coleman JT, Milton DA (2012) Feeding and roost site fidelity of two migratory shorebirds in Moreton Bay, South-Eastern Queensland, Australia. Sunbird J Qld Ornithol Soc The 42:41
Conklin JR, Battley PF (2011) Impacts of wind on individual migration schedules of New Zealand bar-tailed godwits. Behav Ecol 22:854–861. https://doi.org/10.1093/beheco/arr054
Delany S, Scott D, Dodman T, Stroud D (2009) The wader atlas: an atlas of wader populations in Africa and Western Eurasia. Br Birds 102:639–642
Demongin L (2016) Identification guide to birds in the hand. Cambridge University Press, Cambridge, p 392, ISBN 978-2-9555019-0-0
Duijns S, Piersma T (2014) Interference competition in a sexually dimorphic shorebird: prey behaviour explains intraspecific competition. Anim Behav 92:195–201. https://doi.org/10.1016/j.anbehav.2014.04.007
Duijns S, Jukema J, Spaans B et al (2012) Revisiting the proposed leap-frog migration of bar-tailed godwits along the East-Atlantic flyway. Ardea 100:37–43. https://doi.org/10.5253/078.100.0107
Duijns S, Hidayati NA, Piersma T (2013) Bar-tailed godwits Limosa l. lapponica eat polychaete worms wherever they winter in Europe. Bird Study 60:509–517. https://doi.org/10.1080/00063657.2013.836153
Duijns S, van Gils JA, Spaans B et al (2014) Sex-specific winter distribution in a sexually dimorphic shorebird is explained by resource partitioning. Ecol Evol 4:4009–4018. https://doi.org/10.1002/ece3.1213
Durell SEALVD (2000) Individual feeding specialisation in shorebirds: population consequences and conservation implications. Biol Rev 75:503–518. https://doi.org/10.1111/j.1469-185X.2000.tb00053.x
Evans PR (1976) Energy balance and optimal foraging strategies in shorebirds: some implications for their distributions and movements in the non-breeding season. Ardea 55:117–139. https://doi.org/10.5253/arde.v64.p117
Finn PG, Catterall CP, Driscoll PV (2008) Prey versus substrate as determinants of habitat choice in a feeding shorebird. Estuar Coast Shelf Sci 80:381–390. https://doi.org/10.1016/j.ecss.2008.09.001
Folmer EO, Olff H, Piersma T (2010) How well do food distributions predict spatial distributions of shorebirds with different degrees of self-organization? J Anim Ecol 79:747–756. https://doi.org/10.1111/j.1365-2656.2010.01680.x
Fonseca J, Basso E, Serrano D, Navedo JG (2017) Effects of tidal cycles on shorebird distribution and foraging behaviour in a coastal tropical wetland: insights for carrying capacity assessment. Estuar Coast Shelf Sci 198:279–287. https://doi.org/10.1016/j.ecss.2017.09.016
Fretwell SD, Lucas HL (1969) On territorial behavior and other factors influencing habitat distribution in birds. Theor Dev Acta Biotheor 19:16–36. https://doi.org/10.1007/BF01601953
Gabbard C, Sprandel G, Cobb D (2001) Home range analyses of shorebirds wintering along the Gulf of Mexico, Florida, USA. Bull-Wader Study Group 96:79–84
Garabedian JE, Moorman CE, Peterson MN, Kilgo JC (2019) Effects of group size and group density on trade-offs in resource selection by a group-territorial central-place foraging woodpecker. Ibis 162:477–491. https://doi.org/10.1111/ibi.12733
Gaudard C, Quaintenne G, Ward A et al (2018) Synthèse des dénombrements d’oiseaux d’eau en France à la mi-janvier 2017. https://www.lpo.fr/images/actualites/2018/bilan_2017_comptages_wetlands/rapport_oiseauxdeau_2017_web.pdf. Accessed 26 Jun 2019
Gils JAV, Geest MVD, Meulenaer BD et al (2015) Moving on with foraging theory: incorporating movement decisions into the functional response of a gregarious shorebird. J Anim Ecol 84:554–564. https://doi.org/10.1111/1365-2656.12301
Gunnarsson TG, Gill JA, Atkinson PW et al (2006) Population-scale drivers of individual arrival times in migratory birds. J Anim Ecol 75:1119–1127
Hutto RL (1985) Habitat selection by nonbreeding, migratory land. In: Habitat selection in birds, p 455
Ivajnšič D, Kaligarič M, Fantinato E et al (2018) The fate of coastal habitats in the Venice Lagoon from the sea level rise perspective. Appl Geogr 98:34–42. https://doi.org/10.1016/j.apgeog.2018.07.005
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71. https://doi.org/10.2307/1937156
Kennedy M, Gray RD (1994) Agonistic interactions and the distribution of foraging organisms: individual costs and social information. Ethology 96:155–165. https://doi.org/10.1111/j.1439-0310.1994.tb00891.x
Kie JG, Matthiopoulos J, Fieberg J et al (2010) The home-range concept: are traditional estimators still relevant with modern telemetry technology? Philos Trans R Soc B Biol Sci 365:2221–2231. https://doi.org/10.1098/rstb.2010.0093
Klar N, Fernández N, Kramer-Schadt S et al (2008) Habitat selection models for European wildcat conservation. Biol Cons 141:308–319. https://doi.org/10.1016/j.biocon.2007.10.004
Kouba M, Bartoš L, Tomášek V et al (2017) Home range size of Tengmalm’s owl during breeding in Central Europe is determined by prey abundance. PLoS ONE 12:e0177314. https://doi.org/10.1371/journal.pone.0177314
Kraan C, Piersma T, Dekinga A et al (2007) Dredging for edible cockles (Cerastoderma edule) on intertidal flats: short-term consequences of fisher patch-choice decisions for target and non-target benthic fauna. ICES J Mar Sci 64:1735–1742. https://doi.org/10.1093/icesjms/fsm153
Leo KL, Gillies CL, Fitzsimons JA et al (2019) Coastal habitat squeeze: a review of adaptation solutions for saltmarsh, mangrove and beach habitats. Ocean Coast Manag 175:180–190. https://doi.org/10.1016/j.ocecoaman.2019.03.019
Leyrer J, Spaans B, Camara M, Piersma T (2006) Small home ranges and high site fidelity in red knots (Calidris c. canutus) wintering on the Banc d’Arguin. Maurit J Ornithol 147:376–384. https://doi.org/10.1007/s10336-005-0030-8
Lima SL (1986) Predation risk and unpredictable feeding conditions: determinants of body mass in birds. Ecology 67:377–385. https://doi.org/10.2307/1938580
Mace GM, Harvey PH, Clutton-Brock T (1983) Vertebrate home-range size and energetic requirements. The ecology of animal movement. Clarendon, Oxford, pp 32–53
Mallory ML, Gilbert CD (2008) Leg-loop harness design for attaching external transmitters to seabirds. Mar Ornithol 36:183–188
Manly BF, McDonald L, Thomas DL et al (2002) Resource selection by animals: statistical design and analysis for field studies. Springer Science & Business Media, Berlin
Marra PP, Holmes RT (2001) Consequences of dominance-mediated habitat segregation in American redstarts during the nonbreeding season. Auk 118:92–104. https://doi.org/10.1642/0004-8038(2001)118[0092:CODMHS]2.0.CO;2
McGarigal K, Wan HY, Zeller KA et al (2016) Multi-scale habitat selection modeling: a review and outlook. Landsc Ecol 31:1161–1175. https://doi.org/10.1007/s10980-016-0374-x
Morris DW (2003) How can we apply theories of habitat selection to wildlife conservation and management? Wildl Res 30:303–319. https://doi.org/10.1071/WR02028
Morrison H (1979) Critical shorebird resources in James Bay and Eastern North America. Trans N Am Wildl Nat Resour Conf 44:498–507
Murphy ME (1996) Nutrition and metabolism. Avian energetics and nutritional ecology. Springer, Boston, pp 31–60
Novaes WG, Cintra R (2013) Factors influencing the selection of communal roost sites by the black vulture Coragyps atratus (Aves: Cathartidae) in an urban area in central Amazon. Zoologia (Curitiba) 30:607–614. https://doi.org/10.1590/S1984-46702013005000014
Philippe AS, Pinaud D, Cayatte M-L et al (2016) Influence of environmental gradients on the distribution of benthic resources available for shorebirds on intertidal mudflats of Yves Bay, France. Estuar Coast Shelf Sci 174:71–81. https://doi.org/10.1016/j.ecss.2016.03.013
Pienkowski MW, Evans PR (1984) Migratory behavior of shorebirds in the western Palearctic. In: Burger J et al (eds) Shorebirds: migration and foraging behavior. Behavior of Marine Animals: Current Perspectives in Research, 6, pp 73–123
Pierre JP (1994) Effect of sexual dimorphism on feeding behaviour of the bar-tailed godwit Limosa lapponica at a southern hemisphere wintering site. N Z Nat Sci 21:109–112
Piersma T, Hoekstra R, Dekinga A et al (1993) Scale and intensity of intertidal habitat use by knots Calidris canutus in the Western Wadden Sea in relation to food, friends and foes. Neth J Sea Res 31:331–357. https://doi.org/10.1016/0077-7579(93)90052-T
Powell RA (2000) Animal home ranges and territories and home range estimators. Res Tech Anim Ecol Controv Conseq 442:65–110
Robin F, Meunier F, Corre F et al (2015) Site and prey selection by wintering black-tailed godwit Limosa limosa islandica feeding on seagrass beds and bare mudflats on the central Atlantic coast of France. Revue d’Écologie (Terre et Vie) 70:134–147
Rolando A (2002) On the ecology of home range in birds. Revue d’Écologie (Terre et Vie) 57:53–73
Sanzenbacher PM, Haig SM (2002) Residency and movement patterns of wintering dunlin in the Willamette Valley of Oregon. The Condor 104:271–280
Scheiffarth G (2001) The diet of bar-tailed godwits Limosa lapponica in the Wadden Sea: combining visual observations and faeces analyses. Ardea 89:481–494
Schoener TW (1983) Field experiments on interspecific competition. Am Nat 122:240–285. https://doi.org/10.1086/284133
Schuler KL, Schroeder GM, Jenks JA, Kie JG (2014) Ad hoc smoothing parameter performance in kernel estimates of GPS-derived home ranges. Wildl Biol 20:259–266. https://doi.org/10.2981/wlb.12117
Shepherd PCF, Lank DB (2004) Marine and agricultural habitat preferences of dunlin wintering in British Columbia. J Wildl Manag 68:61–73. https://doi.org/10.2193/0022-541X(2004)068[0061:MAAHPO]2.0.CO;2
Si Y, Skidmore AK, Wang T et al (2011) Distribution of barnacle geese Branta leucopsis in relation to food resources, distance to roosts, and the location of refuges. Ardea 99:217–226. https://doi.org/10.5253/078.099.0212
Smith PC, Evans PR (1973) Studies of shorebirds at Lindisfarne, Northumberland. 1. Feeding ecology and behaviour of the bar-tailed godwit. Wildfowl 24:135–140
Sutherland W, Ens B, Goss-Custard J, Hulscher J (1996) Specialisation. Oxf Ornithol Ser 7:56–76
Taft OW, Sanzenbacher PM, Haig SM (2008) Movements of wintering dunlin Calidris alpina and changing habitat availability in an agricultural wetland landscape. Ibis 150:541–549. https://doi.org/10.1111/j.1474-919x.2008.00816.x
Triplet P, Le Dréan Quénec’hdu S, Mahéo R (2010) La barge rousse Limosa lapponica hivernant en France (1977–2009). Alauda 78:207–216
van den Hout PJ, Piersma T (2013) Low-tide distribution of shorebirds in the Wadden Sea. Limosa 86:25–30
van den Hout PJ, Spaans B, Piersma T (2008) Differential mortality of wintering shorebirds on the Banc d’Arguin, Mauritania, due to predation by large falcons: falcon-induced mortality of shorebirds in a major wintering area. Ibis 150:219–230. https://doi.org/10.1111/j.1474-919X.2008.00785.x
van den Hout PJ, van Gils JA, Robin F et al (2014) Interference from adults forces young red knots to forage for longer and in dangerous places. Anim Behav 88:137–146. https://doi.org/10.1016/j.anbehav.2013.11.020
van Winkle W (1975) Comparison of several probabilistic home-range models. J Wildl Manag 39:118–123. https://doi.org/10.2307/3800474
van Moorter B, Rolandsen CM, Basille M, Gaillard J-M (2016) Movement is the glue connecting home ranges and habitat selection. J Anim Ecol 85:21–31
Verger F (2005) Marais maritimes et estuaires du littoral français. Belin, Paris, p 335
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967–2015. Ede, The Netherlands: Wetlands International. URL: https://www.iwc.wetlands.org/index.php/aewatrends. Accessed 3 Dec 2019
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168. https://doi.org/10.2307/1938423
Zwarts L, Wanink JH (1993) How the food supply harvestable by waders in the Wadden Sea depends on the variation in energy density, body weight, biomass, burying depth and behaviour of tidal-flat invertebrates. Neth J Sea Res 31:441–476. https://doi.org/10.1016/0077-7579(93)90059-2
Acknowledgements
We thank Jérôme Jourde for his help in determining the benthic macrofauna, Tania Damany and Sébastien Comarmond for their help with sample processing; Nicolas Lachaussée and Philippe Pineau for the sediment core sampling; Françoise Amélineau and Chloé Tanton for their help during bird capture/marking sessions, Cécilia Pignon-Mussaud who help to arrange the database. We also thank all the volunteers having participating to the bird capture and sediment sampling. This work has been supported by the project ECONAT funded by the Contrat de Plan Etat-Région and the CNRS and the European Regional Development Fund (QUALIDRIS project). This work was also a contribution to the project ANR PAMPAS (ANR-18 CE32-0006) and the Ligue pour la Protection des Oiseaux. We thank Christine Dupuy and Christel Lefrançois for the logistic support. We also thank Hans J. Hartmann for the translation of the summary in German. All work adheres to the legal requirements of the countries in which it was carried out and meets the ethical and animal welfare guidelines.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Gottschalk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Jourdan, C., Fort, J., Pinaud, D. et al. Highly diversified habitats and resources influence habitat selection in wintering shorebirds. J Ornithol 162, 823–838 (2021). https://doi.org/10.1007/s10336-021-01873-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-021-01873-1