
Abstract
Using automated and manual radio-telemetry and resightings of individual colour-ringed birds, we assessed the daily use of space of red knots Calidris canutus canutus at a tropical wintering area along the Sahara coast, the Banc d’Arguin in Mauritania. Confirming earlier suggestions, we found that birds were very faithful to their roosts and that the daily foraging range was small; in the course of several winter months birds used an area of only 2–16 km2 of intertidal area. We found no differences between their movements in daylight and at night. Additionally, individuals seem to return to exactly the same locations in subsequent winters. This pattern is very different from red knots wintering in the temperate Wadden Sea. Here, they readily change roost sites and easily cover areas of about 800 km2 in the course of weeks but, just as in Mauritania, no differences between day and night are apparent. In northern Patagonia and north-western Australia, red knots have range sizes closer to those on the Banc d’Arguin, but here they do show differences in space use between day and night. Ecological explanations for these contrasting patterns require further comparative data based on in-depth studies on the predictability of the food base and the presence of diurnal and nocturnal predators.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The way individual birds adjust their spatial relationships to conspecifics, competitors, predators, food and other features of their environment is fundamental to an animal’s ecology. Spacing behaviour comprises a broad class of behavioural variability across spatial scales as small as individual distances within a flock, to the global scales embodied by annual migrations (Myers 1984). Shorebirds are a particularly interesting group of birds on which to study space use as they are highly visible in the open habitats in which they usually live. Myers (1984) started to develop a comparative approach to spacing behaviour in nonbreeding shorebirds. He stated that spacing behaviour represents predictable responses to ecological and evolutionary processes. Traditionally, studies on spacing behaviour have focused on individuals at the local level, showing how behavioural traits such as territoriality, flocking behaviour, aggressiveness, and roost dispersion are adjusted to foraging opportunities, levels of raptorial predation, and other environmental conditions (cf. Myers et al. 1979; Piersma et al. 1993a; Cresswell 1994; Whitfield 2003; Bednekoff and Lima 2004).
This study is inspired by Myers’ (1984) comparative approach aimed at an evolutionary understanding of inter- and intraspecific variation of time- and space-use in nonbreeding shorebirds. In this paper, we limit ourselves to a study of contrasting space use patterns in a single species, the red knot Calidris canutus. Red knots are a suitable study species, as they always occur in coastal habitats, almost always use bivalves as their staple food, yet winter at a wide range of latitudes on all continents except Antarctica (Piersma and Davidson 1992; Piersma et al. 2005). Additionally, although separate flyways are inhabited by morphologically distinct subspecies, their genetic variability suggests that these subspecies are very closely related and have a very recent, post-Pleistocene, origin (Baker et al. 1994; Buehler and Baker 2005; Buehler and Baker, manuscript in preparation). This would mean that comparisons between subspecies and flyways are not hampered by deeply rooted genetic differences.
Extensive earlier work in the Dutch Wadden Sea and preliminary colour-ringing studies on the Banc d’Arguin, Mauritania, two major wintering sites on the East Atlantic Flyway, have provided evidence that spacing behaviour of red knots during the nonbreeding season shows considerable differences between the two sites (Piersma et al. 1993a) (Fig. 1). Home ranges of red knots in the temperate Wadden Sea cover an area of around 800 km2 in several weeks (van Gils and Piersma 1999; van Gils et al. 2000, 2005), whereas home ranges of red knots wintering on the tropical Banc d’Arguin were suspected to be as small as 10–15 km2 (Piersma et al. 1993a). Interestingly, there are also large differences in the fuelling rates of birds during northward migration, with red knots in tropical areas, including the Banc d’Arguin, consistently fuelling at lower rates than birds further south or north (Piersma et al. 2005). All these differences suggest the importance of specific ecological conditions in otherwise quite similar coastal intertidal areas shaping the movements and performance of shorebirds in space and time. Using telemetry and colour-ringing we here document home range size and site fidelity within and among years of red knots on the Banc d’Arguin, Mauritania.

Comparison between the foraging range of nonbreeding red knots (Calidris canutus) in the Dutch Wadden Sea (islandica subspecies) and on the Banc d’Arguin, Mauritania (canutus subspecies). Adapted from Piersma et al. (1993a)
Materials and methods
Study site
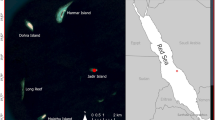
Twenty birds were captured at night with mist nets at two main roost sites near Iwik, Banc d’Arguin, Mauritania, West Africa, on 17 December 2003 (see Fig. 2a). Four of them (#1–#4) were caught at a high tide roost in the north-western part of Baie d’Aouatif (19°54′N, 16°17′W), while 16 individuals (#5–#20) were caught at the high tide roost at Ebel Kheaiznaya (19°54′N, 16°19′W). All birds were released at the beach in the southwest of the Baie d’Aouatif but returned to their original roost site at the latest 2 days after the catch (Fig. 3). For the analyses, the timing of the tides at Iwik were calculated on the basis of predicted times for high and low tide for Dakar, Senegal [admiralty tide tables (ATT), Vol. 2 Europe (excl. UK and Ireland), Mediterranean Sea and Atlantic Ocean], with both high and low tide on average 5 h later in Iwik than in Dakar (Smit et al. 1989; Wolff and Smit 1990). Day and night were separated by morning and evening civil twilight and time of civil twilight was calculated after Burnett (1999).

a Map of the study area in the north of the Parc National du Banc d’Arguin, Mauritania. Squares indicate the positions of the two automatic radio tracking stations (ARTS) at the main high tide roosts at Ebel Kheaiznaya and Baie d’Aouatif. Signals could be received by the ARTS within a range of a 1-km radius (circles). The dotted line indicates the area scanned regularly with portable receivers from December 2003 to April 2004. b Close-up of the intertidal areas around the village of Iwik (number 2a) in which previously colour-ringed individual red knots were intensively searched for in December 2004. Areas in light grey represent mudflats, areas in dark grey show water

Signals received from red knots at the Banc d’Arguin from the automatic radio tracking stations in December 2003 at the two high tide roosts at Baie d’Aouatif and Ebel Kheaiznaya. The dot size gives the number of signals received per hour. Knot #2, #3, and #4 were caught at Baie d’Aouatif, all others at Ebel Kheaiznaya
Biometric data and radio-telemetry
All birds were ringed with a metal ring from the Dutch ringing scheme and an individual combination of four colour rings and a red leg flag (see Piersma and Spaans 2004). Birds were aged according to Prater et al. (1977), distinguishing hatch-year birds (first calendar year or cy), born the year before (second cy) and older birds (adults). Standard biometric data were collected. Additionally, a drop of blood was collected from the brachial vein and stored in 95% ethanol. Using a standard molecular assay verified for red knots (Baker et al. 1999), we determined sex of all transmittered birds. Using superglue, small 1.8-g transmitters (BD2; 173 MHz, guaranteed lifetime of 6 weeks; Holohil Systems, Carp, Ontario, Canada) were attached to trimmed back feathers and the underlying skin (Warnock and Warnock 1993; Nebel et al. 2000). Radio signals were collected by two 24-h operating automated radio tracking stations (ARTS), supplemented by regular checks with mobile antennae and receivers. The ARTS (Telemetrics, Arnhem, The Netherlands; see Green et al. 2002; Battley et al. 2004) were erected at Ebel Kheaiznaya and Baie d’Aouatif (Fig. 2a) and were operating constantly from 18 to 27 and 18 to 28 December 2003, respectively. Each bird was scanned every 6.6 min. This receiver set-up records signals within a 1-km radius (van Gils et al. 2000, 2005; Battley et al. 2004). During the time that the automatic stations were operating signals were received from 14 individuals. For day and night time calculations only days with full 24-h recordings were included. Tide related patterns of roost use were calculated by dividing all signals received by the ARTS per hour by all signals that could possibly be received from the 3 (Baie d’Aouatif) and 11 (Ebel Kheaiznaya) tagged individuals at Baie d’Aouatif and Ebel Kheaiznaya, respectively, in the given hour.
Parallel to the automated recordings, scans were made with portable receivers (IC-R10, ICOM) connected to a three-element Yagi antenna. From 18 to 28 December 2003, all individuals were scanned daily along the beaches surrounding Baie d’Aouatif and Ebel Kheaiznaya (Fig. 2a). On 27 and 28 December, a sailing trip was made to the islands of Niroumi and Nair to check for the transmittered birds (Fig. 2a). There, we regularly checked during different times of the day and all phases of the tide, but no signals were received. Additional scans using portable receivers were made in the periods 11–16 January, 27–30 January, 13–15 February, 17 March, 27 March and 20 April, all in 2004. There were no discrepancies between the automatic receiving stations and the portable receivers. In total, we received signals from 17 birds until the last successful check on 15 February 2004 (see Fig. 4). Thus, only three radio-tagged birds were ‘lost’. Two individuals, one from each catching site, both of which were first year males with rather low body masses at capture, probably left the area and were never observed subsequent to the day of their release. The third individual that went missing was an adult female from Ebel Kheaiznaya that was released after several days of nursing for leg cramp (see Rogers et al. 2004); her signals were only received for two subsequent days. However, this bird was seen alive when it eventually returned to the area in the following year.

Days when signals of radio-tagged red knots were received during checks with portable receivers at the study site near Iwik, Banc d’Arguin, from December 2003 to February 2004. Dark grey shaded areas show periods when scans were made, light grey shaded area show the guaranteed transmitter lifetime. For area scanned see Fig. 2a
Calculating site fidelity by analysing resightings of individually colour-marked birds
During December 2004, intensive ring reading was carried out at nine different areas inside the study area around Iwik (Fig. 2b). With the help of a telescope (magnification 20–60×) we identified individual red knots by their colour-ring combination, which could be read up to a distance of about 300 m. Close to the high tide roosts, flocks were scanned either when they had just left the roost or when they were about to come back. Then, high numbers of birds are concentrated on a comparatively small area, yet the individuals are widespread enough to be able to check most of the birds’ legs. During low tide we followed the birds to their foraging areas, either by boat or on foot. Red knots are easy to observe on their foraging grounds as it is possible to approach quite close to a flock without disturbing them. Site fidelity was analysed by calculating the probability of either an Ebel Kheaiznaya or a Baie d’Aouatif ringed bird being seen in one of the nine areas observed. For all calculations, only red knots ringed at the Banc d’Arguin before 2004 were taken into account. In 2002 and 2003, 31 and 9 red knot were ringed at Baie d’Aouatif, and 224 and 197 at Ebel Kheaiznaya, respectively. Of these ringed birds, 70% were expected to be alive and return the following winters (estimated adult survival of the canutus knots is 0.7; B. Spaans, unpublished data). This results in totals of 21 birds captured at Baie d’Aouatif and 248 at Ebel Kheaiznaya to be expected to be present in the study area in early December 2004, assuming that all birds are site faithful. Note that of the initial 20 radio-tagged birds, 3 were seen again in December 2004. Supposing an annual adult survival of 0.7 and a resighting probability of 0.4 [calculated with MARK (White and Burnham 1999), B. Spaans, unpublished data], the observation of three individuals may be a bit less than the expected five or six, but does not differ significantly. And additionally, in April and May 2005 another three of these formerly radio-tagged birds were seen back in the area (B. Spaans, unpublished data).
Results
The 17 focal birds included one first cy female, one second cy female and two second cy males, and four female and nine male adults. The mean body mass of the three focal transmitter birds from Baie d’Aouatif was 121 g (±11 SD), mean body mass of the 14 focal transmitter birds from Ebel Kheaiznaya was 125±7 g (no difference between the two sites: Mann-Whitney U-test P>0.05; Z=−0.51, n=17). All individuals had full winter plumage and wing moult was fully completed. The three birds (two second cy and an adult) from Baie d’Aouatif were all males. At Ebel Kheaiznaya, there were six females (one first cy, one second cy, four adults) and eight males (one second cy, seven adults).
All but one of the 14 birds that were monitored constantly by the ARTS remained faithful to the roost site where they were caught (Fig. 3). The one exception (#19, a second cy male caught at Ebel Kheaiznaya) switched roosts twice between Ebel Kheaiznaya and Baie d’Aouatif. After its return to Ebel Kheaiznaya 4 days after catching, its signal was received 2 days later at Baie d’Aouatif where it stayed for 2 days before it eventually returned to Ebel Kheaiznaya. Results from the hand-held scans in January and February revealed further switching between the two sites as the bird was detected at Ebel Kheaiznaya in mid-January and back in Baie d’Aouatif in late January. At the last successful scan in mid-February, it was back at Ebel Kheaiznaya.
Most birds showed no changes in the use of roosts between day and night in either Baie d’Aouatif or Ebel Kheaiznaya, except for two birds. One second cy female (#5) used the roost at Ebel Kheaiznaya more during the daylight than at the night-time high tides (Wilcoxon-test P<0.05, Z=−2.52, n=8) and a first cy female (#6) used to roost at Ebel Kheaiznaya more at night than during daytime high tides (Wilcoxon-test P<0.05, Z=−2.52, n=8). We did not discover where these individuals were when out of reach of the ARTS at the roosts.
All 17 focal birds were detected during manual radio scans up to 15 February 2004 (Fig. 4) at their normal locations. This is a few weeks longer than the guaranteed transmitter lifetime of 6 weeks. It is quite likely that birds remained in the area much longer.
In December 2004, a total of 119 individuals colour-ringed locally in late 2002 and 2003 were observed in different parts of the study area (see Fig. 2b). Of the 102 colour-ringed individuals that were seen at Ebel Kheaiznaya and the adjacent mudflats (areas 1, 8, and 9 in Fig. 2b), two were originally caught and ringed at Baie d’Aouatif and 100 at Ebel Kheaiznaya, representing correspondingly 10% (2 out of 21) and 40% (100 out of 248) of the birds ringed at the respective roost. Thus, at Ebel Kheaiznaya, the resighting probability of a bird caught at the Ebel Kheaiznaya roost was 4 times higher than the probability of seeing a bird caught at Baie d’Aouatif. In the areas along the western and northern fringes of Baie d’Aouatif (areas 2, 2a, 3, and 4), the observed fraction of birds caught at both Ebel Kheaiznaya and Baie d’Aouatif was about the same, with 5% of the birds originally captured at Baie d’Aouatif (1 of 21) and also 5% from Ebel Kheaiznaya (13 of 248). In the Baie d’Aouatif (areas 6 and 7), the fraction of birds resighted was 10 times higher for Baie d’Aouatif birds (2 of 21, =10%) than for Ebel Kheaiznaya birds (2 of 248, =1%). As most of the resightings of individually marked birds were made close to their original catching site both in the course of one winter and between years, red knots wintering on the Banc d’Arguin tend to return to the familiar roosts and feeding areas that they have been faithful to in previous years.
Radio-tagged red knots at Baie d’Aouatif could be detected throughout the tidal cycle, suggesting that the birds remained within a 1-km radius of the ARTS during high and low tide (Fig. 5). We thus calculated that red knots roosting at Baie d’Aouatif use an intertidal area of 1.6 km2, as only half of the area inside the listening array consisted of mudflats (Fig. 2a). At Ebel Kheaiznaya, the birds used the area inside the listening array of the ARTS intensively during the outgoing tide, but left the 1-km radius 1 h after low tide, to return to the area only 1 h before high tide (Fig. 5). Although these birds left the Ebel Kheaiznaya coastline, they apparently did not go as far as the mudflats surrounding the islands of Niroumi, Nair, and Northern Tidra, up to 9–11 km away (Fig. 2a). The observations of colour-ringed individuals in December 2004 confirmed that red knots roosting at Ebel Kheaiznaya foraged on mudflats west of the roost (minimum distance to coast approximately 700 m), just outside the receiving area of the ARTS at the roost (Fig. 2b). We deduced that these birds use a home range covering approximately 16 km2 (comprising areas 1, 8, and 9, Fig. 2b).

Percentage of signals received during the tidal cycle at Baie d’Aouatif (upper graph) and Ebel Kheaiznaya (lower graph) as received by the given automatic radio tracking station. Percentages were calculated by dividing all signals received by the ARTS per hour by all signals that could possibly be received from the 3 (Baie d’Aouatif) and 11 (Ebel Kheaiznaya) tagged individuals at Baie d’Aouatif and Ebel Kheaiznaya, respectively, in the given hour
Discussion
Red knots of the nominate canutus subspecies wintering at the Banc d’Arguin show a high degree of site fidelity within and between years. Moreover, as the regular detection of the birds in the area over a period of more than 2 months suggests, this high degree of site fidelity is sustained throughout the whole winter. In contrast to this, islandica red knots wintering in temperate British estuaries have shown no or only very little site fidelity between years or within wintering seasons (Symonds et al. 1984; Rehfisch et al. 2003).
The size of the area used by red knots wintering at the tropical Banc d’Arguin was 16 km2 or less, much smaller than the home ranges of wintering red knots elsewhere in the world (Table 1). For example, red knots of the islandica subspecies wintering in temperate areas like the Wadden Sea covered an area as large as 800 km2 in the course of some weeks (Piersma et al. 1993a; van Gils and Piersma 1999; van Gils et al. 2000, 2005). At other temperate European wintering areas in British estuaries such as Morecambe Bay (Prater 1972) and the Firth of Forth (Symonds et al. 1984), individual islandica red knots also roam over areas of several tens of square kilometres in the course of the winter (see also Rehfisch et al. 2003). Furthermore, they even move quite often between estuaries (Evans 1981, 1984), extending the area used to several hundreds of kilometres. The rufa subspecies wintering in the southern hemisphere in temperate Patagonia, Argentina, likewise covers areas of 100–200 km2 (Sitters et al. 2001). Consequently, both temperate wintering subspecies cover areas up to 50 times larger than the wintering red knots at the tropical Banc d’Arguin. Home ranges of the piersmai subspecies at tropical Roebuck Bay, north-west Australia, are as large as 20 km2 (Rogers et al. 2006), which is in the same order of magnitude, although still slightly larger than at the Banc d’Arguin. Our comparison thus confirms the idea that home ranges of temperate wintering red knots are much larger than of those wintering in the tropics (Piersma et al. 1993a).
One of the main factors influencing habitat use by shorebirds is the availability of their prey (van de Kam et al. 2004). For shorebirds wintering in the Wadden Sea the food supply may be characterised as unpredictable. Erratic large drops in temperature make benthic food unavailable to birds, therefore causing them to leave the site in search of warmer areas (e.g. the British estuaries; Evans 1976). Contrarily, benthic food may suddenly be highly available at some spots when, e.g. after periods of frost, moribund shellfish with gaping valves provide a rich food resource (Zwarts and Wanink 1993). In other years, prey harvestable by red knots may be highly variable due to unpredictable spat fall (Zwarts et al. 1992; Beukema et al. 1993). Consequently, temperate wintering birds have to keep track with ever changing feeding conditions (Evans et al. 1980; Evans 1981). Tropical wetlands lack climatic extremes such as cold spells, and no cyclones occur off the West African coast. Moreover, they appear to lack the seasonal synchronisation of the reproductive cycle of the invertebrate prey (Wolff 1991; Piersma et al. 1993b; Dittmann 2002; de Goeij et al. 2003). Thus, prey living close to the surface, and without peaked seasonal occurrences, provide more or less constant feeding conditions throughout the year (Piersma 1980; Piersma et al. 1993a). Additionally, in contrast to temperate wintering islandica knots, canutus lacks a mass peak (Piersma 1994) and has a lower basic metabolic rate in winter (Piersma et al. 1996). We propose that this allows birds to confine themselves to very well-known small areas. This even seems to be the case during the period of fuelling for northward migration (P.J. van den Hout and L. van Kooten, personal communication), a season when feeding time is stretched to the maximum at Banc d’Arguin (Zwarts et al. 1990), but rates of mass increase are relatively low (Piersma et al. 2005).
In our comparison of winter spacing behaviour at different wintering sites we came across differences in diurnal and nocturnal habitat choice at some sites. Such differences are often reported in shorebird studies (Mouritsen 1994; Burton and Armitage 2005) and are frequently related to differences in predation risk as the birds are exposed to a different group of predators at night than during the day. Rogers et al. (2006) showed that roosting behaviour of wintering red knots in north-west Australia differed considerably between day and night because of an increased predation risk at the favourite daytime roosts during the night. To decrease predation risk, the birds appeared to accept the extra costs of time and energy expenditure by choosing roost sites further away from the feeding habitats. In wintering red knots in Patagonia, Argentina, predator avoidance has been suggested to lead to the complete avoidance of profitable foraging sites at night (Sitters et al. 2001). These two examples suggest that sites may be so dangerous that increased costs of commuting between foraging areas and roost sites, and even the avoidance of good feeding sites, may be accepted (Cresswell 1994; Hilton et al. 1999; van Gils et al. 2000). If the lack of differences between the day and night time use of roosts and feeding areas is anything to go by, red knots wintering on the Banc d’Arguin did not seem to encounter differences in predation pressure on the roosts between day and night. Note that daytime predation by raptors is well documented (Bijlsma 1990), just as in the Wadden Sea (van Gils and Piersma 1999; van Gils et al. 2000, 2005). However, from our results, we cannot exclude small scale differences in feeding site choice between day and night.
In conclusion, we suggest that the combination of food predictability and the absence of day/night differences in predation pressure both at high tide roosts and at foraging areas, allow canutus red knots wintering at the Banc d’Arguin to have very small and predictable home ranges compared with red knots wintering in other areas. True to Myers’ (1984) research agenda, we must now assemble further comparative data based on in-depth studies on the predictability of the food base and the presence of diurnal and nocturnal predators to fully exploit the comparative potential provided by red knots world-wide.
Zusammenfassung
Kleine Home Ranges und hohe Ortstreue bei überwinternden Knutts (Calidris c. canutus) an der Banc d’Arguin, Mauretanien
Mit Hilfe von mit Radiosendern und individuellen Farbringkombinationen markierter Knutts (Calidris canutus canutus) haben wir das tägliche Raumnutzungsmuster in einem tropischen Überwinterungsgebiet entlang der westafrikanischen Atlantikküste, der Banc d’Arguin, Mauretanien, erfasst. Frühere Annahmen bestätigend, fanden wir eine sehr hohe Ortstreue sowohl im Verlauf eines Winters als auch zwischen den Jahren und sehr kleine Home Ranges von 2 – 16 km² im Verlauf von mehreren Monaten. Überwinternde Knutts an der Banc d’Arguin zeigten keine Unterschiede im Raumnutzungsverhalten zwischen Tag und Nacht. Diese Untersuchungsergebnisse unterschieden sich grundlegend von Knutts der islandica Unterart, die in gemäßigten Breiten, z.B. dem Wattenmeer, überwintern. Dort sind die Home Ranges mit ca. 800 km² im Verlauf von ein paar Wochen deutlich größer, jedoch existieren ebenfalls keine Unterschiede in den Bewegungsmustern zwischen Tag und Nacht. Im Vergleich dazu besitzen überwinternde rufa Knutts in Nordpatagonien und piersmai Knutts in Nordwestaustralien vergleichsweise kleinere Home Ranges als islandica Knutts im Wattenmeer, zeigen allerdings unterschiedliche Raumnutzungsmuster zwischen Tag und Nacht. Zur Klärung der ökologischen Ursachen dieser unterschiedlichen Muster sind weiterführende Studien erforderlich, die den Einfluss der Vorhersagbarkeit der Nahrungsgrundlage und von unterschiedlichem Predationsdruck bei Tag und Nacht beleuchten.
References
Baker AJ, Piersma T, Rosenmeier L (1994) Unravelling the intraspecific phylogeography of Knots Calidris canutus: a progress report on the search for genetic markers. J Ornithol 135:599–608
Baker AJ, Piersma T, Greenslade AD (1999) Molecular versus phenotypic sexing in Red Knots Calidris canutus. Condor 101:887–893
Battley PF, Piersma T, Rogers DI, Dekinga A, Spaans B, van Gils J (2004) Do body condition and plumage during fuelling predict northwards departure dates of Great Knots (Calidris tenuirostris) from north-west Australia? Ibis 146:46–60
Bednekoff PA, Lima SL (2004) Risk allocation and competition in foraging groups: reversed effects of competition if group size varies under risk of predation. Proc R Soc Lond B 271:1491–1496
Beukema JJ, Essink K, Michaelis H, Zwarts L (1993) Year-to-year variability in the biomass of macrobenthic animals on tidal flats of the Wadden Sea: how predictable is this food source for birds? Neth J Sea Res 31:319–330
Bijlsma RG (1990) Predation by large falcons on wintering waders on the Banc d’Arguin, Mauritania. Ardea 78:75–82
Buehler DM, Baker AJ (2005) Population divergence times and historical demography in Red Knots and Dunlins. Condor 107:497–513
Burnett K (1999) Sun rise and set, and twilight. http://www.xylem.f2s.com/kepler/sunrise.html
Burton NHK, Armitage MJS (2005) Differences in the diurnal and nocturnal use of intertidal feeding grounds by Redshanks Tringa totanus. Bird Study 52:120–128
Cresswell W (1994) Age-dependent choice of redshank (Tringa totanus) feeding location: profitability or risk? J Anim Ecol 63:589–600
Dittmann S (2002) Benthic fauna in tropical tidal flats of Hitchinbrook Channel, NE Australia: diversity, abundance, and their spatial and temporal variation. Wetlands Ecol Manage 10:323–333
Evans PR (1976) Energy balance and optimal foraging strategies in shorebirds: some implications for their distributions and movements in the non-breeding season. Ardea 64:117–139
Evans PR (1981) Migration and dispersal of shorebirds as a survival strategy. In: Jones NV, Wolff WJ (eds) Feeding and survival strategies in estuarine organisms. Plenum Press, New York, pp 275–290
Evans PR (1984) The British Isles. In: Evans PR, Goss-Custard JD, Hale WG (eds) Coastal waders and waterfowl in winter. Cambridge University Press, Cambridge, pp 261–275
Evans PR, Brearey DM, Goodyer LR (1980) Studies on Sanderlings at Teesmouth, NE England. Wader Study Group Bull 30:18–20
van Gils J, Piersma T (1999) Day- and nighttime movements of radiomarked Knots, Calidris canutus, staging in the western Wadden Sea in July–August 1995. Wader Study Group Bull 89:36–44
van Gils J, Piersma T, Dekinga A, Spaans B (2000) Distributional ecology of individually radio-marked Knots Calidris canutus in the western Dutch Wadden Sea in August–October 1999. Limosa 73:29–34
van Gils J, Dekinga A, Spaans B, Vahl W K, Piersma T (2005) Digestive bottleneck affects foraging decisions in Red Knots (Calidris canutus). II. Patch choice and length of working day. J Anim Ecol 74:120–130
de Goeij P, Lavaleye M, Pearson GB, Piersma T (2003) Seasonal changes in the macro-zoobenthos of a tropical mudflat. NIOZ-Report 2003–2004, Texel
Green M, Piersma T, Jukema J, de Goeij P, Spaans B, van Gils J (2002) Radio-telemetry observations of the first 650 km of the migration of bar-tailed godwits Limosa lapponica from the Wadden Sea to the Russian Arctic. Ardea 90:71–80
Hilton GM, Ruxton GD, Cresswell W (1999) Choice of foraging area with respect to predation risk in Redshanks: the effects of weather and predator activity. Oikos 87:295–302
van de Kam J, Ens BJ, Piersma T, Zwarts L (2004) Shorebirds. An illustrated behavioural ecology. KNNV, Utrecht
Mouritsen KN (1994) Day and night feeding in dunlins Calidris alpina: choice of habitat, foraging technique and prey. J Avian Biol 25:55–62
Myers JP (1984) Spacing behavior of non-breeding shorebirds. In: Burger J, Olla BL (eds) Shorebirds—migration and foraging behaviour. Plenum Press, New York, pp 271–321
Myers JP, Connors PG, Pitelka FA (1979) Territory size in wintering sanderlings: the effects of prey abundance and intruder density. Auk 96:551–561
Nebel S, Piersma T, van Gils J, Dekinga A, Spaans B (2000) Length of stopover, fuels storage and a sex-bias in the occurrence of Red Knots Calidris c. canutus and C.c.islandica in the Wadden Sea during southward migration. Ardea 88:165–176
Piersma T (1980) Macrobenthic fauna of the intertidal flats. In: Altenburg W, Engelmoer M, Mes R, Piersma T (eds) Wintering waders on the Banc d’Arguin, Mauritania. Stichting Veth tot steun aan Waddenonderzoek, Leiden, pp 50–66
Piersma T (1994) Close to the edge: energetic bottlenecks and the evolution of migratory pathways in Knots. Uitgeverij Het Open Boek, Den Burg, Texel
Piersma T, Davidson NC (1992) The migration and annual cycles of five subspecies of Knots in perspective. Wader Study Group Bull 64 [Suppl]:187–197
Piersma T, Hoekstra R, Dekinga A, Koolhaas A, Wolf P, Battley PF, Wiersma P (1993a) Scale and intensity of intertidal habitat use by Knots Calidris canutus in the Western Wadden Sea in relation to food, friends and foes. Neth J Sea Res 31:331–357
Piersma T, de Goeij P, Tulp I (1993b) An evaluation of intertidal feeding habitats from a shorebird perspective: towards relevant comparisons between temperate and tropical mudflats. Neth J Sea Res 31:503–512
Piersma T, Bruinzeel L, Drent R, Kersten M, Van der Meer J, Wiersma P (1996) Variability in basal metabolic rate of a long-distance migrant shorebird (Red Knot, Calidris canutus) reflects shifts in organ sizes. Physiol Zool 69:191–217
Piersma T, Spaans B (2004) The power of comparison: ecological studies on waders worldwide. Limosa 77:43–54
Piersma T, Rogers DI, Gonzalez PM, Zwarts L, Niles LJ, de Lima Serrano do Nacimento I, Minton CDT, Baker AJ (2005) Fuel storage rates in Red Knots worldwide: facing the severest ecological constraint in tropical intertidal conditions? In: Greenberg R, Marra PP (eds) Birds of two worlds: the ecology and evolution of migration. Johns Hopkins University Press, Baltimore, Md., pp 262–273
Prater AJ (1972) The ecology of Morecambe Bay. III. The food and feeding habits of Knot (Calidris canutus, L.) in Morecambe Bay. J Appl Ecol 9:179–194
Prater AJ, Marchant JH, Vuorinen J (1977) Guide to the identification and ageing of Holoarctic Shorebirds. BTO, Tring, Herts
Rehfisch MM, Insley H, Swann B (2003) Fidelity of overwintering shorebirds to roosts on the Moray Basin, Scotland: implications for predicting impacts of habitat loss. Ardea 91:53–70
Rogers DI, Battley PF, Sparrow J, Koolhaas A, Hassell CJ (2004) Treatment of capture myopathy in shorebirds: a successful trial in North-West Australia. J Field Ornithol 75:157–164
Rogers DI, Battley PF, Piersma KG, van Gils J, Rogers KG (2006) High-tide habitat choice: insights from modelling roost selection by shorebirds around a trophical bay. Anim Behav (in press)
Sitters HP, Gonzalez PM, Piersma T, Baker AJ, Price DJ (2001) Day and night feeding habitat of Red Knot in Patagonia: profitability versus safety? J Field Ornithol 72:86–95
Smit CJ, Blomert AM, Meijboom A, Wolff WJ (1989) Hydrographical measurements. In: Ens BJ, Piersma T, Wolff WJ, Zwarts L (eds) Report of the Dutch-Mauritanian project Banc d’Arguin 1985–1986, pp 43–56 (WIWO-report 25/RIN-rapport 89/6, Texel)
Symonds FL, Langslow DR, Pienkowski MW (1984) Movements of wintering shorebirds within the Firth of Forth: species differences in usage of an intertidal complex. Biol Conserv 28:187–215
Warnock N, Warnock S (1993) Attachment of radio-transmitters to sandpipers: review and methods. Wader Study Group Bull 70:28–30
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study [Suppl] 46:S120–S139
Whitfield DP (2003) Redshank Tringa totanus flocking behaviour, distance from cover and vulnerability to Sparrowhawk Accipiter nisus predation. J Avian Biol 34:163–169
Wolff WJ (1991) The interaction of benthic macrofauna and birds in tidal flat estuaries: a comparison of the Banc d’Arguin, Mauritania, and some estuaries in The Netherlands. In: Elliott M, Ducrotoy J-P (eds) Estuaries and coasts: spatial and temporal intercomparisons. Olsen & Olsen, Fredensbord, pp 299–306
Wolff WJ, Smit CJ (1990) The Banc d’Arguin, Mauritania, as an environment for coastal birds. Ardea 78:17–38
Zwarts L, Wanink JH (1993) How the food supply harvestable by shorebirds in the Wadden Sea depends on the variation in energy density, body weight, biomass, burying depth and behaviour of tidal-flat invertebrates. Neth J Sea Res 31:441–476
Zwarts L, Blomert A-M, Hupkes R (1990) Increase of feeding time in waders preparing for spring migration from the Banc d’Arguin, Mauritania. Ardea 78:237–256
Zwarts L, Blomert A-M, Wanink JH (1992) Annual and seasonal variation in the food supply harvestable by knot Calidris canutus staging in the Wadden Sea in late summer. Mar Ecol Prog Ser 83:129–139
Acknowledgements
We thank the co-participants of the 2003 expedition (Jan van Dijk, Piet van den Hout, Laurens van Kooten, Bob Loos, and Jeroen Reneerkens) for their valuable help in catching and radio-tracking the birds and the co-participants of the 2004 expedition (Pieter Honkoop, Harry Horn, Piet van den Hout, Jan van de Kam, Jan Lewis, Bob Loos, Jeroen Reimerink, Jeroen Reneerkens, and Jaap Strikwerda) for helping to relocate previously marked individuals. Authorities of the Parc National du Banc d’Arguin gave permission to work in Iwik. We also thank the collaborators of the Parc National du Banc d’Arguin, especially Sall Mamadou Alassane and Ba Amadou Galo and the other inhabitants of the Biological Station in Iwik for their cheerful contributions and co-operation. Antonio Araujo gave key support throughout the work in Mauritania. We also would like to thank Gregor Scheiffarth and an anonymous referee for valuable comments on an earlier version of the manuscript. Finally, many thanks to Dick Visser for preparing the graphs. The two expeditions were financially supported by the Prins Bernhard Cultuurfondsprijs voor Natuurbehoud 2004 to TP.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein
Rights and permissions
About this article
Cite this article
Leyrer, J., Spaans, B., Camara, M. et al. Small home ranges and high site fidelity in red knots (Calidris c. canutus) wintering on the Banc d’Arguin, Mauritania. J Ornithol 147, 376–384 (2006). https://doi.org/10.1007/s10336-005-0030-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-005-0030-8