
Abstract
Renal afferent and efferent sympathetic nerves are involved in the regulation of blood pressure and have a pathophysiological role in hypertension. Additionally, several conditions that frequently coexist with hypertension, such as heart failure, obstructive sleep apnea, atrial fibrillation, renal dysfunction, and metabolic syndrome, demonstrate enhanced sympathetic activity. Renal denervation (RDN) is an approach to reduce renal and whole body sympathetic activation. Experimental models indicate that RDN has the potential to lower blood pressure and prevent cardio-renal remodeling in chronic diseases associated with enhanced sympathetic activation. Studies have shown that RDN can reduce blood pressure in drug-naïve hypertensive patients and in hypertensive patients under drug treatment. Beyond its effects on blood pressure, sympathetic modulation by RDN has been shown to have profound effects on cardiac electrophysiology and cardiac arrhythmogenesis. RDN can display anti-arrhythmic effects in a variety of animal models for atrial fibrillation and ventricular arrhythmias. The first non-randomized studies demonstrate that RDN may promote the maintenance of sinus rhythm following catheter ablation in patients with atrial fibrillation. Registry data point towards a beneficial effect of RDN to prevent ventricular arrhythmias in patients with heart failure and electrical storm. Further large randomized placebo-controlled trials are needed to confirm the antihypertensive and anti-arrhythmic effects of RDN. Here, we will review the current literature on anti-arrhythmic effects of RDN with the focus on atrial fibrillation and ventricular arrhythmias. We will discuss new insights from preclinical and clinical mechanistic studies and possible clinical implications of RDN.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Renal sensory afferent and efferent nerves allow bidirectional communication between the central nervous system and the kidney. Peripheral and central inputs alter efferent renal sympathetic nerve activity, influencing the innervated structural and functional components of the kidney, including vessels, glomeruli, and tubuli [1]. The physiological efferent renal sympathetic nerve activity regulates renal blood flow, glomerular filtration rate, tubular sodium and water handling, and stimulates renin release from the juxtaglomerular apparatus which regulates blood pressure and renal perfusion [1, 2]. Under pathophysiological conditions, abnormal efferent renal sympathetic nerve activity can contribute to the associated abnormalities of renal function which, in turn, are of importance in the pathogenesis of several disease conditions like hypertension, renal disease and heart failure [2]. This efferent signaling is further regulated by afferent input from sensory renal chemo- and mechano-receptors in the kidney. Afferent fibers from the kidney travel along with the sympathetic nerves at the level of the kidney and then enter the dorsal roots and project to regions of the brainstem involved in cardiovascular control [2,3,4,5]. Therefore, the kidney represents a source of increased sympathetic activation under certain pathophysiological conditions, such as renal ischaemia, hypoxia, and intrinsic renal disease [6,7,8]. The density of peri-arterial renal sympathetic nerve fibers is lower in distal segments and dorsal locations when compared with proximal superior locations. There is a clear predominance of efferent nerve fibers, with decreasing density of afferent nerves from proximal to distal peri-arterial and renal parenchyma [3].
These observations support the rationale to modulate autonomic innervation of the renal artery, arterioles and tubules in order to reduce norepinephrine spillover from the kidney and to influence sympathetically dependent pathophysiologies [9,10,11]. Renal denervation (RDN) substantially prevents progressive blood pressure rise and associated renal injury and cardiac remodeling in animal models of hypertension [9, 10] (Fig. 1). Factors such as withdrawal of sympathetic nerve activity to the kidney and subsequent changes in fluid mobilization and reduction in circulating angiotensin II, as well as removal of elevated afferent renal nerve activity as a consequence of a pathophysiological alteration in the kidney, may contribute to the antihypertensive effects of RDN. Interestingly, selective destruction of afferent renal nerves had limited antihypertensive effects in hypertensive Dahl salt-sensitive rats [12, 13].

Effects of renal denervation (RDN) on blood pressure, renal catecholamine levels and renal and cardiac interstitial fibrosis in spontaneously hypertensive obese rats (SHR-ob). a Mean arterial blood pressure measured by telemetry in control rats (Ctr), SHRs-ob, and SHRs-ob+renal sympathetic denervation (SHRs-ob+RDN) during 100 days after RDN performed at the age of 34 weeks. b Immunohistochemical renal tyrosine hydroxylase staining and c renal norepinephrine tissue content at the age of 48 weeks. d Interstitial left ventricular (LV) and e kidney fibrosis with representative histological images. (Adopted from [9])
In non-randomized studies [14, 15] and registries [16], RDN reduced blood pressure and partially reversed cardiac hypertensive end-organ damage in patients with drug-resistant hypertension [17,18,19]. A recent randomized sham controlled trial (SYMPLICITY HTN-3, ClinicalTrials.gov number, NCT01418261) could not confirm blood pressure reduction by RDN in drug-resistant hypertensive patients [20]. However, in another published, randomized, sham controlled trial, RDN by a new ablation catheter reduced blood pressure in medication naïve hypertensive patients [21]. Several factors such as problems with assessment of renal denervation efficacy, variable experience of the proceduralists and the complex issues of patient drug compliance, including improvement of drug adherence in initially poorly adherent patients (the Hawthorne effect), placebo effect and regression to the mean, makes the interpretation of the results difficult.
Beyond regulation of blood pressure, increased sympathetic activity is suggested to contribute to the progression of cardiac arrhythmias [22]. At the cellular level, norepinephrine, the neurotransmitter of the sympathetic nervous system, is released from postganglionic neurons in response to sympathetic stimuli activating cardiac beta-receptors. Subsequently, altered cardiac calcium handling and electrophysiology contribute to arrhythmogenic mechanisms including delayed afterdepolarization-related ectopic firing and re-entry [23, 24]. Sympathetic hyperinnervation is reported in the atria of dogs following rapid atrial pacing [25], and increased sympathetic and vagal nerve discharges can be documented before the onset of atrial fibrillation (AF) in dogs with pacing-induced congestive heart failure [26]. Additionally, recent data have shown that sympathetic activation promotes the development of the arrhythmogenic substrate via neurohumoral mechanisms, such as the renin–angiotensin–aldosterone system, leading to the upregulation of profibrotic pathways in the atria [23, 24]. Therefore, elevated sympathetic activity may contribute to both cardiac structural and electrical remodeling, thus predisposing to arrhythmia. Activation of the autonomic nervous system plays an important role in the initiation and the maintenance of AF, but may also be targeted for the maintenance of sinus rhythm. Beta-receptor blockade by metoprolol was effective in preventing recurrence of AF after successful cardioversion [27], and autonomic modulation by, for example, low-level baroreceptor stimulation can help to maintain sinus rhythm [22, 28].
These observations raise the question of whether modulation of the autonomic nervous system by targeting the renal sympathetic nerves might be an effective strategy to improve rhythm control in patients with cardiac arrhythmias. Here, we will review the current literature on the anti-arrhythmic effects of autonomic nervous system modulation through RDN, with a particular focus on atrial fibrillation and ventricular arrhythmias. We will discuss new insights from preclinical and clinical mechanistic studies and possible clinical implications.
Modulation of renal sympathetic innervation by renal denervation
In small animal models, RDN can be performed by combined surgical and chemical approaches, which modulate both efferent and afferent renal nerve activity [9, 11]. Both kidneys are approached through medial laparotomy, the visible nerves in the area of the renal hilus are removed and approximately 2–4 mm of the adventitia from the renal artery is stripped. The area is then moistened with a phenol/ethanol (10–20%) solution for 10–15 min. Combined surgical and chemical RDN reduces kidney tissue norepinephrine levels by > 90–95% in several animal models. The release of norepinephrine into the urine and renal venous blood, which is inducible by efferent sympathetic nerve stimulation, can be reduced by 80% after RDN in animal studies [5, 29]. Previous studies on RDN in normotensive rats by a 10% phenol/ethanol solution suggest renal reinnervation within 9 weeks [30]. In obese spontaneously hypertensive rats, RDN by a 20% phenol/ethanol solution resulted in a more sustained reduction in blood pressure, renal sympathetic nerve density and renal norepinephrine tissue content (Fig. 1) [9].
Additionally, RDN can prevent cardiac neural remodeling and nerve sprouting under pathophysiological conditions, suggesting reduction of afferent nerve activity. In spontaneously hypertensive rats, RDN by a 20% phenol/ethanol solution resulted in a reduced expression of the norepinephrine transporter in the kidney and in the heart (Fig. 2). In rats with ischemic heart failure, RDN preserves sympathetic nerve innervation in the ventricles, thus improving cardiac function [31], while in goats and dogs with atrial fibrillation induced by atrial tachypacing, RDN achieved a significant reduction in atrial sympathetic nerve sprouting [32,33,34].

a Representative image during a chemical renal denervation (RDN) procedure in a spontaneously hypertensive rat. Both kidneys were surgically denervated by cutting all visible nerves in the area of the renal hilus and by strippingapproximately 2–4 mm of the adventitia from the renal artery. The area was then moistened with a phenol/ethanol solution for 10–15 min using a brush. b Representative western blot to quantify protein expression of norepinephrine transporter in kidney and heart tissue of sham-denervated spontaneously hypertensive rat and of RDN spontaneously hypertensive rats
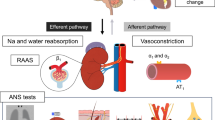
Most of the brainstem regions involved in cardiovascular control receive input from the renal afferent fibers. Afferent signals arising from the renal sympathetic nerves are able to influence nerve activity of several ganglia including those innervating the heart (Fig. 3). Studies suggest a direct link between renal sympathetic nerve activity and left stellate ganglion (LSG) activity [35, 36]. Renal sympathetic nerve stimulation is associated with LSG neuronal activity and upregulates the level of LSG nerve growth factor expression [36]. Importantly, the stimulation procedure not only activates the efferent sympathetic nerves but also the afferent fibers going from the kidney to the central nervous system. Additionally, the stimulation frequency used in these renal nerve activation studies are likely to impact renal hemodynamic or excretory renal function, which has not been sufficiently characterized and studied.

Effects of renal denervation (RDN) on efferent and afferent signaling of the kidney. NE Norepinephrine, MSNA muscle sympathetic nerve activity, LSG Left stellate ganglion
Besides modulation of circulating catecholamines and whole body sympathetic nerve activity, bilateral RDN caused significant central and peripheral sympathetic nerve remodeling, improved baroreflex sensetivity [37] and reduced stellate ganglion nerve activity in ambulatory dogs [36]. Catheter-based RDN can also affect cardiac sympathetic activity. Selective RDN significantly reduces cardiac sympathetic overdrive assessed by 123I-MIBG scintigraphy [39, 40].
A catheter-based approach for RDN has been developed for human use. RDN can result in an up to 50% reduction of renal norepinephrine spillover measured with a radiochemical tracer methodology using 3H-norepinephrine in humans [12, 13]. Additionally, firing of single sympathetic vasoconstrictor fibers (measured by single muscle sympathetic nerve activity), was reduced by 37% [38]. These findings indicate that sympathetic activation can be reduced by RDN beyond just the kidneys, suggesting a combined modulation of efferent and afferent signaling (Fig. 3).
The first-generation devices used radiofrequency pulses emitted from a monopolar electrode positioned under fluoroscopic guidance in each of the renal arteries. The latest generation devices are multi-electrode catheters allowing a more standardized and more intense ablation procedure [41]. During the development of RDN catheters, the ablation strategy changed from ablation limited to the main vessels to more extended ablation in the main and subsequent branch vessels [41]. This revised approach was associated with higher efficacy and lower variability in treatment effects in preclinical studies. Alternatively, ultrasound or chemical ablation have been investigated in clinical studies [41].
Despite great advantages of catheter techniques, there is no universally accepted measure by which the completeness of RDN can be properly assessed intraprocedurally. Although periprocedural changes in circulating norepinephrine or neuropeptide Y [42], nerve stimulation-induced blood pressure changes [43, 44] and anatomy-guided lesion placement [45] have been introduced to improve ablation outcome, a standardized protocol to directly measure renal sympathetic nerve activity in humans is still lacking. In sedated and unrestrained conscious rodent models, assessment and quantification of renal nerve activity is feasible [46, 47]. The development of endovascular recording of renal nerve action potentials may provide a useful accessory tool to assess successful RDN. Innovation in this area will be crucial to predict and monitor the therapeutic value of RDN [48].
Effects of RDN on cardiac arrhythmias
Beyond hypertension and hypertensive end-organ damage, cardiac rhythm disorders, namely atrial fibrillation and ventricular arrhythmias, have been identified as a promising and emerging target of RDN [49].
Atrial electrophysiological effects of renal denervation
In experimental data, RDN resulted in a reduction in heart rate and AV-conduction velocity in pigs [50]. In chloralose/urethane anasthetized pigs, neither atrial effective refractory period nor P-wave duration was influenced by acute RDN, thus excluding a direct impact of RDN on atrial refractoriness or atrial conduction during sinus rhythm [50]. Additionally, the sensitivity of ganglionated plexi was not modulated by RDN. In an AF pig model with rapid atrial pacing, RDN reduced the duration of pacing-induced AF [51]. However, AF-induced electrical remodeling was not attenuated [50]. Reduced AV-conduction velocity was associated with lower ventricular heart rate during AF [50].
In a goat model of persistent AF induced by atrial tachypacing [32], RDN reduced renal norepinephrine concentrations, blunted atrial neural remodeling and reduced AF complexity along with structural remodeling (Fig. 4). Moreover, increased AF-inducibility, shortening and dispersion of atrial refractoriness as well as elevated plasma norepinephrine levels were almost completely prevented by RDN in animals subjected to stimulation of left stellate ganglion and rapid atrial pacing for three hours [52, 53].

Effects of renal denervation (RDN) compared to sham intervention (SHAM) in goats with atrial fibrillation induced by an implanted pace maker. a Representative X-ray images of the kidney. Locations of ablation delivery are indicated by superimposed small black points in the vessels. b Renal tissue norepinephrine concentrations in the left (white bars) and right (black bars) kidneys determined after the sacrifice experiments. c Representative perivascular tyrosine hydroxylase (TH) staining (brown twigs) of cardiac sympathetic nerves in SHAM and RDN goats with 6 weeks AF (magnification, ×1000). d Quantification of TH-positive (indicating sympathetic nerve structures) fraction of the perivascular area in the anterior left atrium (LAant), posterior left atrium (LApost) and right atrium (RA). e Quantification of AF complexity: representative spatial and temporal distribution of AF activation pattern during one AF cycle length of the right and left atrium in SHAM (top) and RDN (bottom) goats with 6 weeks. Isochronal maps: time between isochrones 10 ms, red earliest, blue latest activation. (Adopted from [32])
In a pig model of obstructive sleep apnea (OSA), shortened atrial refractoriness [54, 55] acutely induced by applied negative thoracic pressure was primarily mediated by combined sympathovagal activation, since it could be influenced by atropine, bilateral vagotomy or beta-receptor blockade [54, 56]. Compared to beta-blocker treatment, RDN resulted in more pronounced attenuation of the shortening in atrial refractoriness during OSA maneuvers, which might explain the superior anti-arrhythmic effect of RDN compared with beta-blocker therapy in this animal model [55]. Importantly, anti-arrhythmic drugs such as amiodarone or sotalol displayed a considerably less pronounced anti-arrhythmic effect compared to RDN in the same model [55]. In pigs undergoing repetitive OSA maneuvers over 4 h, RDN inhibited spontaneous atrial premature beats, and reduced the number AF episodes as well as AF duration [56]. The observed reduction in spontaneous atrial extra beats by RDN may reduce the trigger for AF in OSA [56]. Additionally, RDN has been shown to reduce susceptibility to AF in a canine models of renal impairment, induced by embolization of small branches of the renal artery in the right kidney using gelatin sponge granules [57], and tachycardiomyopathy, induced by ventricular tachypacing [58]. Taken together, these studies provide strong preclinical evidence towards an anti-arrhythmogenic effect of RDN.
Clinical data
RDN can reduce heart rate in patients with resistant hypertension [59]. In persistent AF, RDN can improve rate control [60], which might improve clinical symptoms and outcomes in patients with AF. RDN has been shown to prevent or even reverse atrial remodeling determined by echocardiography [61] or by electroanatomical mapping [62]. In a small study of patients with symptomatic AF and resistant hypertension, the atrial anti-arrhythmic effects of circumferential pulmonary vein isolation (PVI) combined with RDN were investigated [63,64,65]. Patients who received the combined procedures showed significant reductions in average systolic and diastolic blood pressure, whereas those in the PVI-only group did not show any significant improvement in blood pressure. Other electrophysiological parameters were not obviously changed by RDN. At 1-year follow-up, 69% of patients who received both procedures maintained sinus rhythm, compared to 29% of those in the PVI-only group [63]. In chronic kidney disease patients, the addition of RDN to pulmonary vein isolation reduced recurrence of paroxysmal atrial fibrillation [66]. In a case report, even RDN without PVI reduced blood pressure and attenuated paroxysmal AF episodes, which were symptomatic and drug-resistant before RDN [67]. It remains unknown whether sympathetic modulation of the autonomic nervous system by RDN can display anti-arrhythmic effects in hypertensive AF patients independent from its blood pressure-lowering effects. This important clinical and pathophysiological question should be addressed in a randomized sham controlled trial, in which RDN is compared to aggressive and adopted up-titration of antihypertensive drugs in the control group.
Ventricular arrhythmias, heart failure and renal denervation
In dogs, 3 h of renal sympathetic nerve stimulation increased LSG neuronal activity, and facilitated the incidence of ventricular arrhythmias during acute myocardial ischemia. Interestingly, the increase in ventricular arrhythmias could be attenuated by LSG ablation [34]. In different pig models with acute ischemia/reperfusion [66] (Fig. 5) or with myocardial infarction induced by a permanent coronary occlusion [69,70,71], RDN has been shown to reduce ventricular ectopic activity and ventricular fibrillation. Also in a dog model of heart failure induced by atrial tachycardiomyopathy induced by ventricular high-rate pacing, RDN attenuated the ventricular remodeling process [72, 73]. The occurrence of spontaneous premature ventricular contractions and the subsequent ventricular dysfunction could be prevented by sympathetic modulation by the procedure [74]. Similarly, arrhythmogenic prolongation of ventricular repolarization (QT-interval) induced by simulated sleep apnea [75] or by cesium [76] could be attenuated by RDN.

Effects of renal denervation (RDN) on ventricular fibrillation (VF) in a pig model for ventricular ischemia and reperfusion. a Representative view of the left ventricular during ischemia reperfusion experiments. Atrial electrophysiology was recorded by an epicardial catheter. b Incidence of VF during ischemia and the reperfusion phase in RDN-treated compared to SHAM-treated pigs. c Representative hemodynamics and electrocardiographic (ECG) tracings during 20 min of left anterior descending coronary artery ligation followed by reperfusion in a SHAM-treated and a RDN-treated animal. (Adopted from [68])
Clinical data
In patients with heart failure with reduced ejection fraction, circulating norepinephrine concentrations predict mortality [77]. Interestingly, cardiac and renal norepinephrine spillover is increased in mild to moderate and severe chronic heart failure compared to healthy subjects, with the absolute renal norepinephrine spillover higher than the cardiac spillover suggesting that the kidney contributes more to total norepinephrine spillover than the heart in heart failure [78, 79].
A small case series has provided evidence that, in patients with dilated cardiomyopathy and an electrical storm, RDN was able to reduce ICD shocks and ventricular ectopic activity [80]. These anti-arrhythmic effects could be confirmed in several other case series [81, 82] and in an international multicenter registry [83]. RDN may be most beneficial for patients with heart failure with recurrent, refractory arrhythmias that cannot tolerate maximal beta-blockade and are not eligible for anti-arrhythmic catheter-based ablation of ventricular tachycardia. Alternatively, RDN may also be used as an adjunct strategy in patients undergoing catheter ablation. In patients with heart failure, RDN reduced NT-proBNP and was safe without any adverse event deterioration of other indices of cardiac and renal function [84]. The role of autonomic modulation by RDN in patients with heart failure and ventricular arrhythmias needs to be further investigated in randomized, sham-controlled trials.
Conclusions
RDN is a promising and safe strategy to modulate the autonomic nervous system activity. Beyond its blood pressure-lowering effects, animal models of atrial fibrillation have demonstrated favorable electrophysiological changes, reverse remodeling and potential anti-arrhythmic effects of sympathetic modulation by RDN. Interestingly, adjunct RDN improved outcome of catheter ablation in atrial fibrillation patients in small non-randomized clinical trials; these findings warrant further clinical trials. Additionally, animal experiments and early registry data also suggest promising ventricular anti-arrhythmic effects by RDN in the setting of heart failure. Further studies are needed to investigate the anti-arrhythmic effects of RDN.
References
DiBona GF, Kopp UC (1997) Neural control of renal function. Physiol Rev 77:75–197
Böhm M, Linz D, Ukena C, Esler M, Mahfoud F (2014) Renal denervation for the treatment of cardiovascular high risk-hypertension or beyond? Circ Res 115:400–409
Sakakura K, Ladich E, Cheng Q, Otsuka F, Yahagi K, Fowler DR, Kolodgie FD, Virmani R, Joner M (2014) Anatomic assessment of sympathetic peri-arterial renal nerves in man. J Am Coll Cardiol 64:635–643
Stella A, Zanchetti A (1991) Functional role of renal afferents. Physiol Rev 71:659–682
Bradley T, Hjemdahl P (1986) Influence of afferent renal nerve activity on contralateral renal overflow of noradrenaline and dopamine to plasma in the dog. Acta Physiol Scand 128:119–120
Larsen R, Thorp A, Schlaich M (2014) Regulation of the sympathetic nervous system by the kidney. Curr Opin Nephrol Hypertens 23:61–68
Patel KP, Knuepfer MM (1986) Effect of afferent renal nerve stimulation on blood pressure, heart rate and noradrenergic activity in conscious rats. J Auton Nerv Syst 17:121–130
Rogenes PR (1982) Single-unit and multiunit analyses of renorenal reflexes elicited by stimulation of renal chemoreceptors in the rat. J Auton Nerv Syst 6:143–156
Linz D, Hohl M, Schütze J, Mahfoud F, Speer T, Linz B, Hübschle T, Juretschke HP, Dechend R, Geisel J, Rütten H, Böhm M (2015) Progression of kidney injury and cardiac remodeling in obese spontaneously hypertensive rats: the role of renal sympathetic innervation. Am J Hypertens 28:256–265
Mahfoud F, Moon LB, Pipenhagen CA, Jensen JA, Pathak A, Papademetriou V, Ewen S, Linz D, Böhm M (2016) Catheter-based radio-frequency renal nerve denervation lowers blood pressure in obese hypertensive swine model. J Hypertens 34:1854–1862
Hohl M, Linz D, Fries P, Müller A, Stroeder J, Urban D, Speer T, Geisel J, Hummel B, Laufs U, Schirmer SH, Böhm M, Mahfoud F (2016) Modulation of the sympathetic nervous system by renal denervation prevents reduction of aortic distensibility in atherosclerosis prone ApoE-deficient rats. J Transl Med. 14:167
Foss JD, Fink GD, Osborn JW (2016) Differential role of afferent and efferent renal nerves in the maintenance of early- and late-phase Dahl S hypertension. Am J Physiol Regul Integr Comp Physiol 310:R262–267
Osborn JW, Foss JD (2017) Renal nerves and long-term control of arterial pressure. Compr Physiol. 7:263–320
Krum H, Schlaich M, Whitbourn R, Sobotka PA, Sadowski J, Bartus K, Kapelak B, Walton A, Sievert H, Thambar S, Abraham WT, Esler M (2009) Catheter-based renal sympathetic denervation for resistant hypertension: a multicentre safety and proof-of-principle cohort study. Lancet 373:1275–1281
Esler MD, Krum H, Sobotka PA, Schlaich MP, Schmieder RE (2010) Böhm M for the Symplicity HTN-2 Investigators. Renal sympathetic denervation in patients with treatment resistant hypertension (the Symplicity HTN-2 trial): a randomised controlled trial. Lancet 376:1903–1909
Böhm M, Ukena C, Ewen S, Linz D, Zivanovic I, Hoppe U, Narkiewicz K, Ruilope L, Schlaich M, Negoita M, Schmieder R, Williams B, Zeymer U, Zirlik A, Mancia G, Mahfoud F (2016) Global SYMPLICITY registry investigators. Renal denervation reduces office and ambulatory heart rate in patients with uncontrolled hypertension: 12-month outcomes from the global SYMPLICITY registry. J Hypertens 34:2480–2486
Linz D, Mancia G, Mahfoud F, Narkiewicz K, Ruilope L, Schlaich M, Kindermann I, Schmieder RE, Ewen S, Williams B, Böhm M (2017) Global SYMPLICITY Registry Investigators. Renal artery denervation for treatment of patients with self-reported obstructive sleep apnea and resistant hypertension: results from the Global SYMPLICITY Registry. J Hypertens 35:148–153
Mahfoud F, Urban D, Teller D, Linz D, Stawowy P, Hassel JH, Fries P, Dreysse S, Wellnhofer E, Schneider G, Buecker A, Schneeweis C, Doltra A, Schlaich MP, Esler MD, Fleck E, Böhm M, Kelle S (2014) Effect of renal denervation on left ventricular mass and function in patients with resistant hypertension: data from a multi-centre cardiovascular magnetic resonance imaging trial. Eur Heart J 35:2224–2231
Schirmer SH, Sayed MM, Reil JC, Ukena C, Linz D, Kindermann M, Laufs U, Mahfoud F, Böhm M (2014) Improvements of left-ventricular hypertrophy and diastolic function following renal denervation—Effects beyond blood pressure and heart rate reduction. J Am Coll Cardiol 63:1916–1923
Bhatt DL, Kandzari DE, O’Neill WW, D’Agostino R, Flack JM, Katzen BT, Leon MB, Liu M, Mauri L, Negoita M, Cohen SA, Oparil S, Rocha-Singh K, Townsend RR, Bakris GL (2014) SYMPLICITY HTN-3 investigators. A controlled trial of renal denervation for resistant hypertension. N Engl J Med 370:1393–1401
Townsend RR, Mahfoud F, Kandzari DE, Kario K, Pocock S, Weber MA, Ewen S, Tsioufis K, Tousoulis D, Sharp ASP, Watkinson AF, Schmieder RE, Schmid A, Choi JW, East C, Walton A, Hopper I, Cohen DL, Wilensky R, Lee DP, Ma A, Devireddy CM, Lea JP, Lurz PC, Fengler K, Davies J, Chapman N, Cohen SA, DeBruin V, Fahy M, Jones DE, Rothman M (2017) Böhm M; SPYRAL HTN-OFF MED trial investigators*. Catheter-based renal denervation in patients with uncontrolled hypertension in the absence of antihypertensive medications (SPYRAL HTN-OFF MED): a randomised, sham-controlled, proof-of-concept trial. Lancet. https://doi.org/10.1016/S0140-6736(17)32281-X
Linz D, Ukena C, Mahfoud F, Neuberger HR, Böhm M (2014) Atrial autonomic innervation: a target for interventional antiarrhythmic therapy? J Am Coll Cardiol 63:215–224
Linz D, van Hunnik A, Ukena C, Ewen S, Mahfoud F, Schirmer SH, Lenski M, Neuberger HR, Schotten U, Böhm M (2014) Renal denervation: effects on atrial electrophysiology and arrhythmias. Clin Res Cardiol 103:765–774
Ripplinger CM, Noujaim SF, Linz D (2016) The nervous heart. Prog Biophys Mol Biol 120:199–209
Jayachandran JV, Sih HJ, Winkle W, Zipes DP, Hutchins GD, Olgin JE (2000) Atrial fibrillation produced by prolonged rapid atrial pacing is associated with heterogeneous changes in atrial sympathetic innervation. Circulation 101:1185–1191
Ogawa M, Zhou S, Tan AY, Song J, Gholmieh G, Fishbein MC, Luo H, Siegel RJ, Karagueuzian HS, Chen LS, Lin SF, Chen PS (2007) Left stellate ganglion and vagal nerve activity and cardiac arrhythmias in ambulatory dogs with pacing-induced congestive heart failure. J Am Coll Cardiol 50:335–343
Kühlkamp V, Schirdewan A, Stangl K, Homberg M, Ploch M, Beck OA (2000) Use of metoprolol CR/XL to maintain sinus rhythm after conversion from persistent atrial fibrillation: a randomized, double-blind, placebo-controlled study. J Am Coll Cardiol 36:139–146
Linz D, Hohl M, Khoshkish S, Mahfoud F, Ukena C, Neuberger HR, Wirth K, Böhm M (2016) Low-level but not high-level baroreceptor stimulation inhibits atrial fibrillation in a pig model of sleep apnea. J Cardiovasc Electrophysiol 27:1086–1092
Oliver JA, Pinto J, Sciacca RR, Cannon PJ (1980) Basal norepinephrine overflow into the renal vein: effect of renal nerve stimulation. Am J Physiol 239:F371–F377
Mulder J, Hökfelt T, Knuepfer MM, Kopp UC (2013) Renal sensory and sympathetic nerves reinnervate the kidney in a similar time-dependent fashion after renal denervation in rats. Am J Physiol Regul Integr Comp Physiol 304:R675–682
Pinkham MI, Loftus MT, Amirapu S, Guild SJ, Quill G, Woodward WR, Habecker BA, Barrett CJ (2017) Renal denervation in male rats with heart failure improves ventricular sympathetic nerve innervation and function. Am J Physiol Regul Integr Comp Physiol 312:R368–R379
Linz D, van Hunnik A, Hohl M, Mahfoud F, Wolf M, Neuberger HR, Casadei B, Reilly SN, Verheule S, Böhm M, Schotten U (2015) Catheter-based renal denervation reduces atrial nerve sprouting and complexity of atrial fibrillation in goats. Circ Arrhythm Electrophysiol. 8:466–474
Wang X, Zhao Q, Huang H, Tang Y, Xiao J, Dai Z, Yu S, Huang C (2013) Effect of renal sympathetic denervation on atrial substrate remodeling in ambulatory canines with prolonged atrial pacing. PLoS ONE 8:e64611
Huang B, Yu L, Scherlag BJ, Wang S, He B, Yang K, Liao K, Lu Z, He W, Zhang L, Po SS, Jiang H (2014) Left renal nerves stimulation facilitates ischemia-induced ventricular arrhythmia by increasing nerve activity of left stellate ganglion. J Cardiovasc Electrophysiol 25:1249–1256
Yu L, Huang B, Wang Z, Wang S, Wang M, Li X, Zhou L, Meng G, Yuan S, Zhou X, Jiang H (2017) impacts of renal sympathetic activation on atrial fibrillation: the potential role of the autonomic cross talk between kidney and heart. J Am Heart Assoc 6:e004716
Tsai WC, Chan YH, Chinda K, Chen Z, Patel J, Shen C, Zhao Y, Jiang Z, Yuan Y, Ye M, Chen LS, Riley AA, Persohn SA, Territo PR, Everett TH, Lin SF, Vinters HV, Fishbein MC, Chen PS (2017) Effects of renal sympathetic denervation on the stellate ganglion and brain stem in dogs. Heart Rhythm 14:255–262
Tsai WC, Chan YH, Chinda K, Lai WT, Lin SF, Chen PS (2014) Renal sympathetic denervation decreases the incidence of atrial tachycardia and improves baroreflex sensitivity in ambulatory dogs. Heart Rhythm 11:236
Hering D, Lambert EA, Marusic P, Walton AS, Krum H, Lambert GW, Esler MD, Schlaich MP (2013) Substantial reduction in single sympathetic nerve firing after renal denervation in patients with resistant hypertension. Hypertension 61:457–464
Berukstis A, Vajauskas D, Gargalskaite U, Misonis N, Burneikaite G, Zakarkaite D, Miglinas M, Laucevicius A (2016) Impact of renal sympathetic denervation on cardiac sympathetic nerve activity evaluated by cardiac MIBG imaging. EuroIntervention 11:1070–1076
Donazzan L, Mahfoud F, Ewen S, Ukena C, Cremers B, Kirsch CM, Hellwig D, Eweiwi T, Ezziddin S, Esler M, Böhm M (2016) Effects of catheter-based renal denervation on cardiac sympathetic activity and innervation in patients with resistant hypertension. Clin Res Cardiol 105:364–371
Mahfoud F, Schmieder RE, Azizi M, Pathak A, Sievert H, Tsioufis C, Zeller T, Bertog S, Blankestijn PJ, Böhm M, Burnier M, Chatellier G, Durand Zaleski I, Ewen S, Grassi G, Joner M, Kjeldsen SE, Lobo MD, Lotan C, Felix Lüscher T, Parati G, Rossignol P, Ruilope L, Sharif F, van Leeuwen E, Volpe M, Windecker S, Witkowski A, Wijns W (2017) Proceedings from the 2nd European Clinical Consensus Conference for device-based therapies for hypertension: state of the art and considerations for the future. Eur Heart J. https://doi.org/10.1093/eurheartj/ehx215
Dörr O, Ewen S, Liebetrau C, Möllmann H, Gaede L, Linz D, Hohl M, Troidl C, Bauer T, Böhm M, Hamm C, Mahfoud F, Nef H (2015) Neuropeptide Y as an indicator of successful alterations in sympathetic nervous activity after renal sympathetic denervation. Clin Res Cardiol 104:1064–1071
de Jong MR, Adiyaman A, Gal P, Smit JJ, Delnoy PP, Heeg JE, van Hasselt BA, Lau EO, Persu A, Staessen JA, Ramdat Misier AR, Steinberg JS, Elvan A (2016) Renal nerve stimulation-induced blood pressure changes predict ambulatory blood pressure response after renal denervation. Hypertension 68:707–714
Chinushi M, Suzuki K, Saitoh O, Furushima H, Iijima K, Izumi D, Sato A, Sugai M, Iwafuchi M (2016) Electrical stimulation-based evaluation for functional modification of renal autonomic nerve activities induced by catheter ablation. Heart Rhythm 13:1707–1715
Mahfoud F, Tunev S, Ewen S, Cremers B, Ruwart J, Schulz-Jander D, Linz D, Davies J, Kandzari DE, Whitbourn R, Böhm M, Melder RJ (2015) Impact of lesion placement on efficacy and safety of catheter-based radiofrequency renal denervation. J Am Coll Cardiol 66:1766–1775
Stocker SD, Muntzel MS (2013) Recording sympathetic nerve activity chronically in rats: surgery techniques, assessment of nerve activity, and quantification. Am J Physiol Heart Circ Physiol 305:H1407–1416
Hamza SM, Hall JE (2012) Direct recording of renal sympathetic nerve activity in unrestrained, conscious mice. Hypertension 60:856–864
Barber-Chamoux N, Esler MD (2017) Predictive factors for successful renal denervation: should we use them in clinical trials. Eur J Clin Invest. https://doi.org/10.1111/eci.12792
Linz D, Hunnik Av, Ukena C, Mahfoud F, Ewen S, Verheule S, Böhm M, Schotten U (2014) Effects of renal denervation on atrial arrhythmogenesis. Future Cardiol 10:813–822
Linz D, Mahfoud F, Schotten U, Ukena C, Hohl M, Neuberger HR, Wirth K, Böhm M (2013) Renal sympathetic denervation provides ventricular rate control but does not prevent atrial electrical remodeling during atrial fibrillation. Hypertension 61:225–231
Linz D, Mahfoud F, Schotten U, Ukena C, Neuberger HR, Wirth K, Böhm M (2012) Renal sympathetic denervation suppresses postapneic blood pressure rises and atrial fibrillation in a model for sleep apnea. Hypertension 60:172–178
Hou Y, Hu J, Po SS, Wang H, Zhang L, Zhang F, Wang K, Zhou Q (2013) Catheter-based renal sympathetic denervation significantly inhibits atrial fibrillation induced by electrical stimulation of the left stellate ganglion and rapid atrial pacing. PLoS ONE 8:e78218
Zhou Q, Zhou X, TuEr-Hong ZL, Wang H, Yin T, Li Y, Zhang L, Lu Y, Xing Q, Zhang J, Yang Y, Tang B (2016) Renal sympathetic denervation suppresses atrial fibrillation induced by acute atrial ischemia/infarction through inhibition of cardiac sympathetic activity. Int J Cardiol 203:187–195
Linz D, Schotten U, Neuberger HR, Böhm M, Wirth K (2011) Negative tracheal pressure during obstructive respiratory events promotes atrial fibrillation by vagal activation. Heart Rhythm. 8:1436–1443
Linz D, Schotten U, Neuberger HR, Böhm M, Wirth K (2011) Combined blockade of early and late activated atrial potassium currents suppresses atrial fibrillation in a pig model of obstructive apnea. Heart Rhythm. 8:1933–1939
Linz D, Hohl M, Nickel A, Mahfoud F, Wagner M, Ewen S, Schotten U, Maack C, Wirth K, Böhm M (2013) Effect of renal denervation on neurohumoral activation triggering atrial fibrillation in obstructive sleep apnea. Hypertension 62:767–774
Liang Z, Shi XM, Liu LF, Chen XP, Shan ZL, Lin K, Li J, Chen FK, Li YG, Guo HY, Wang YT (2015) Renal denervation suppresses atrial fibrillation in a model of renal impairment. PLoS ONE 10:e0124123
Wang X, Zhao Q, Deng H, Wang X, Guo Z, Dai Z, Xiao J, Wan P, Huang C (2014) Effects of renal sympathetic denervation on the atrial electrophysiology in dogs with pacing-induced heart failure. Pacing Clin Electrophysiol 37:1357–1366
Ukena C, Mahfoud F, Spies A, Kindermann I, Linz D, Cremers B, Laufs U, Neuberger HR, Böhm M (2013) Effects of renal sympathetic denervation on heart rate and atrioventricular conduction in patients with resistant hypertension. Int J Cardiol 167:2846–2851
Qiu M, Shan Q, Chen C, Geng J, Guo J, Zhou X, Qian W, Tang L, Yin Y (2016) Renal sympathetic denervation improves rate control in patients with symptomatic persistent atrial fibrillation and hypertension. Acta Cardiol 71:67–73
Schirmer SH, Sayed MM, Reil JC, Lavall D, Ukena C, Linz D, Mahfoud F, Böhm M (2015) Atrial remodeling following catheter-based renal denervation occurs in a blood pressure-and heart rate-independent manner. JACC Cardiovasc Interv 8:972–980
McLellan AJ, Schlaich MP, Taylor AJ, Prabhu S, Hering D, Hammond L, Marusic P, Duval J, Sata Y, Ellims A, Esler M, Peter K, Shaw J, Walton A, Kalman JM, Kistler PM (2015) Reverse cardiac remodeling after renal denervation: atrial electrophysiologic and structural changes associated with blood pressure lowering. Heart Rhythm 12:982–990
Pokushalov E, Romanov A, Corbucci G, Artyomenko S, Baranova V, Turov A, Shirokova N, Karaskov A, Mittal S, Steinberg JS (2012) A randomized comparison of pulmonary vein isolation with versus without concomitant renal artery denervation in patients with refractory symptomatic atrial fibrillation and resistant hypertension. J Am Coll Cardiol 60:1163–1170
Pokushalov E, Romanov A, Katritsis DG, Artyomenko S, Bayramova S, Losik D, Baranova V, Karaskov A, Steinberg JS (2014) Renal denervation for improving outcomes of catheter ablation in patients with atrial fibrillation and hypertension: early experience. Heart Rhythm 11:1131–1138
Romanov A, Pokushalov E, Ponomarev D, Strelnikov A, Shabanov V, Losik D, Karaskov A, Steinberg JS (2017) Pulmonary vein isolation with concomitant renal artery denervation is associated with reduction of both arterial blood pressure and atrial fibrillation burden: data from implantable cardiac monitor. Cardiovasc Ther. https://doi.org/10.1111/1755-5922.12264
Kiuchi MG, Chen S (2017) E Silva GR, Rodrigues Paz LM, Kiuchi T, de Paula Filho AG, Lima Souto GL. The addition of renal sympathetic denervation to pulmonary vein isolation reduces recurrence of paroxysmal atrial fibrillation in chronic kidney disease patients. J Interv Card Electrophysiol 48:215–222
Vollmann D, Sossalla S, Schroeter MR, Zabel M (2013) Renal artery ablation instead of pulmonary vein ablation in a hypertensive patient with symptomatic, drug-resistant, persistent atrial fibrillation. Clin Res Cardiol 102:315–318
Linz D, Wirth K, Ukena C, Mahfoud F, Pöss J, Linz B, Böhm M, Neuberger HR (2013) Renal denervation suppresses ventricular arrhythmias during acute ventricular ischemia in pigs. Heart Rhythm 10:1525–1530
Huang B, Yu L, He B, Lu Z, Wang S, He W, Yang K, Liao K, Zhang L, Jiang H (2014) Renal sympathetic denervation modulates ventricular electrophysiology and has a protective effect on ischaemia-induced ventricular arrhythmia. Exp Physiol 99:1467–1477
Jackson N, Gizurarson S, Azam MA, King B, Ramadeen A, Zamiri N, Porta-Sánchez A, Al-Hesayen A, Graham J, Kusha M, Massé S, Lai PF, Parker J, John R, Kiehl TR, Nair GK, Dorian P, Nanthakumar K (2017) Effects of renal artery denervation on ventricular arrhythmias in a postinfarct model. Circ Cardiovasc Interv 10:e004172
Chang S-N, Chang S-H, Yu CC et al (2017) Renal denervation decreases susceptibility to arrhythmogenic cardiac alternans and ventricular arrhythmia in a rat model of post–myocardial infarction heart failure. J Am Coll Cardiol Basic Trans Sci 2:184–193
Guo Z, Zhao Q, Deng H, Tang Y, Wang X, Dai Z, Xiao J, Wan P, Wang X, Huang H, Huang C (2014) Renal sympathetic denervation attenuates the ventricular substrate and electrophysiological remodeling in dogs with pacing-induced heart failure. Int J Cardiol 175:185–186
Dai Z, Yu S, Zhao Q, Meng Y, He H, Tang Y, Wang X, Xiao J, Wang X, Huang C (2014) Renal sympathetic denervation suppresses ventricular substrate remodelling in a canine high-rate pacing model. EuroIntervention 10:392–399
Yamada S, Lo LW, Chou YH, Lin WL, Chang SL, Lin YJ, Liu SH, Cheng WH, Tsai TY, Chen SA (2017) Beneficial effect of renal denervation on ventricular premature complex induced cardiomyopathy. J Am Heart Assoc 6:e004479
Linz D, Denner A, Illing S, Hohl M, Ukena C, Mahfoud F, Ewen S, Reil JC, Wirth K, Böhm M (2016) Impact of obstructive and central apneas on ventricular repolarisation: lessons learned from studies in man and pigs. Clin Res Cardiol 105:639–647
Yu L, Huang B, Zhou X, Wang S, Wang Z, Wang M, Li X, Zhou L, Meng G, Yuan S, Wang Y, Jiang H (2017) Renal sympathetic stimulation and ablation affect ventricular arrhythmia by modulating autonomic activity in a cesium-induced long QT canine model. Heart Rhythm 14:912–919
Cohn JN, Levine TB, Olivari MT, Garberg V, Lura D, Francis GS, Simon AB, Rector T (1984) Plasma norepinephrine as a guide to prognosis in patients with chronic congestive heart failure. N Engl J Med 311:819–823
Hasking GJ, Esler MD, Jennings GL, Burton D, Johns JA, Korner PI (1986) Norepinephrine spillover to plasma in patients with congestive heart failure: evidence of increased overall and cardiorenal sympathetic nervous activity. Circulation 73:615–621
Petersson M, Friberg P, Eisenhofer G, Lambert G, Rundqvist B (2005) Long-term outcome in relation to renal sympathetic activity in patients with chronic heart failure. Eur Heart J 26:906–913
Ukena C, Bauer A, Mahfoud F, Schreieck J, Neuberger HR, Eick C, Sobotka PA, Gawaz M, Böhm M (2012) Renal sympathetic denervation for treatment of electrical storm: first-in-man experience. Clin Res Cardiol 101:63–67
Armaganijan LV, Staico R, Moreira DA, Lopes RD, Medeiros PT, Habib R, Melo Neto J, Katz M, Armaganijan D, Sousa AG, Mahfoud F, Abizaid A (2015) 6-month outcomes in patients with implantable cardioverter-defibrillators undergoing renal sympathetic denervation for the treatment of refractory ventricular arrhythmias. JACC Cardiovasc Interv 8:984–990
Remo BF, Preminger M, Bradfield J, Mittal S, Boyle N, Gupta A, Shivkumar K, Steinberg JS, Dickfeld T (2014) Safety and efficacy of renal denervation as a novel treatment of ventricular tachycardia storm in patients with cardiomyopathy. Heart Rhythm 11:541–546
Ukena C, Mahfoud F, Ewen S, Bollmann A, Hindricks G, Hoffmann BA, Linz D, Musat D, Pavlicek V, Scholz E, Thomas D, Willems S, Böhm M, Steinberg JS (2016) Renal denervation for treatment of ventricular arrhythmias: data from an International Multicenter Registry. Clin Res Cardiol 105:873–879
Hopper I, Gronda E, Hoppe UC, Rundqvist B, Marwick TH, Shetty S, Hayward C, Lambert T, Hering D, Esler M, Schlaich M, Walton A, Airoldi F, Brandt MC, Cohen SA, Reiters P, Krum H (2017) Sympathetic response and outcomes following renal denervation in patients with chronic heart failure: 12-month outcomes from the symplicityhf feasibility study. J Card Fail. https://doi.org/10.1016/j.cardfail.2017.06.004
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
DL, FM, PS and MB received research grants, speaker honoraria, and consultancy fees from Medtronic/Ardian, St. Jude, Boston Scientific, and Cordis. The other authors report no conflicts.
Rights and permissions
About this article

Cite this article
Linz, D., Hohl, M., Elliott, A.D. et al. Modulation of renal sympathetic innervation: recent insights beyond blood pressure control. Clin Auton Res 28, 375–384 (2018). https://doi.org/10.1007/s10286-018-0508-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10286-018-0508-0