
Abstract
Since the publication of the theory of island biogeography, ecologists have postulated that fragmentation of continuous habitat presents a prominent threat to species diversity. However, negative fragmentation effects may be artifacts; the result of species diversity declining with habitat loss, and habitat loss correlating positively with degree of fragmentation. In this study, we used butterfly assemblages on islands of Lake of the Woods, Ontario, Canada to decouple habitat fragmentation from habitat loss and test two competing hypotheses: (1) the island effect hypothesis, which predicts that decreasing fragment size and increasing fragment isolation reduces species diversity beyond the effects of habitat loss, and (2) the habitat amount hypothesis, which negates fragmentation effects and predicts that only total habitat area determines the diversity of species persisting on fragmented landscapes. Using eight independent size classes of islands (ranging from 0.1 to 8.0 ha) that varied in number of islands while holding total area constant, species diversity comparisons, species accumulation curves, and species–area relationship extrapolations demonstrated that smaller insular habitats contained at least as many butterfly species as continuous habitat. However, when highly mobile species occurring on islands without their larval food plants were excluded from analyses, island effects on potentially reproducing species became apparent. Similarily, generalized linear models suggested that effects of island isolation and vascular plant richness on insular butterfly richness were confounded by species of high mobility. We conclude that inter-fragment movements of highly mobile species may obscure important fragmentation effects on potentially reproducing populations, questioning support for the habitat amount hypothesis.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Within continuous habitats, island archipelagoes, and fragmented landscapes, species richness increases with total area surveyed (MacArthur and Wilson 1963, Wilson and MacArthur 1967; Wilson and Willis 1975). Indeed, the positive species–area relationship (SAR) is widely cited as the closest thing ecology has to a law (Schoener 1976; Lomolino 2000). As a corollary of the SAR, loss of habitat results in loss of species (He and Hubbell 2011); however, the configuration of remaining habitat is widely thought to also have an effect on species diversity (Mendenhall et al. 2014; Haddad et al. 2017). Linked to the process of habitat loss, variation in the size and isolation of habitat fragments may be described as degree of habitat fragmentation (Fahrig 2003). While some have gone so far as to assert that “habitat fragmentation is considered by many biologists to be the single greatest threat to biological diversity” (e.g., Noss 1991 p. 27), in many studies, habitat fragmentation has not been distinguished from habitat loss. After decoupling habitat fragmentation from habitat loss, effects of fragmentation appear to vary widely within and among both landscapes and taxa (Quinn and Harrison 1988; Debinski and Holt 2000; Fahrig 2003; Mendenhall et al. 2014).
Over the past half-century, ecologists have related a variety of theories to the fragmentation problem; perhaps the most prevalent and influential being the theory of island biogeography (MacArthur and Wilson 1963, Wilson and MacArthur 1967). Drawing on the heuristic power of island biogeography, ecologists have frequently likened the ecologies of oceanic archipelagos to those of fragmented landscapes (Haila 1986, 1990; Ovaskainen 2002; Fahrig 2013; Haddad et al. 2017). As with oceanic islands, species diversity within habitat fragments is predicted to reach equilibria between colonization and extinction rates, principally determined by fragment size and isolation. We refer to this application of equilibrium theory as the “island effect hypothesis,” which predicts that habitat fragmentation reduces species diversity below what is predicted based on habitat loss alone. A key assumption of this hypothesis is that fragment edges delimit species assemblages, such that population processes, including colonization and extinction, occur at the level of habitat fragments. If fragments are too small to support viable populations following their isolation, a gradual loss of species (faunal relaxation) will reduce species diversity at a rate inversely related to fragment size (Diamond 1972, 1975; MacArthur and Wilson 1963, Wilson and MacArthur 1967; Connor and McCoy 1979; Gonzalez 2000). Consequentially, slopes of species–area curves (z values) across isolated habitat fragments are predicted to be steeper than those within continuous habitats (Gonzalez 2000; Haddad et al. 2017). In the context of habitat fragmentation, the ultimate “steady-state” legacy of an island effect will be several small habitat fragments supporting fewer species than continuous habitat of equivalent area (Gonzalez 2000; Fahrig 2013).
In contrast to the island effect hypothesis, the recently proposed “habitat amount hypothesis” suggests that the size and isolation of habitat fragments have little effect on species diversity (Fahrig 2013). The central premise of this hypothesis is that fragment edges do not delimit populations, such that only the aggregate amount of habitat determines the number of species persisting on fragmented landscapes. It follows that negative relationships between habitat fragmentation and species diversity are best interpreted as artifacts; the result of species diversity declining with habitat loss, and habitat loss correlating positively with degree of fragmentation (Harrison and Bruna 1999; Fahrig 2003, 2013; Yaacobi et al. 2007). Faunal relaxation (or the gradual loss of species) following habitat loss is therefore predicted as a landscape-level process, unrelated to the configuration of remaining habitat. In sum, predictions of the habitat amount hypothesis are indistinguishable from the passive sampling hypothesis, developed in the context of oceanic islands (Connor and McCoy 1979). Both hypotheses predict that species richness increases with fragment/island size only because of the sample area effect: larger sample areas contain more individuals, which, for a given abundance distribution, belong to more species (Fahrig 2013). If the sample area effect best explains patterns of species diversity on fragmented landscapes, species–area curves across isolated habitats will approximate those of continuous habitats (Haddad et al. 2017). In consequence, fragmented and continuous habitat of equivalent total area should support equivalent numbers of species (Gonzalez 2000; Fahrig 2013).
While differences in species–area slopes between fragmented and continuous habitat have been interpreted as evidence against the habitat amount hypothesis (Haddad et al. 2017), Gotelli (1996 p. 227) states that, “…the most sensible view is that slopes of species–area curves are simply fitted constants, with little or no biological significance.” Indeed, slopes of species–area curves have been found to vary unpredictably within and between both sampling locations and taxa (Connor and McCoy 1979), with z values clustering in certain ranges by chance and because of reporting biases in the literature (Gotelli 1996; Gonzalez 2000). In contrast with z value interpretations, comparing species diversity across sets of habitat fragments (while controlling for total habitat area) is a proven, tractable method for assessing habitat fragmentation–species diversity relationships (Yaacobi et al. 2007; Gavish et al. 2012; Fahrig 2013). Such analyses bear on the long-standing SLOSS debate, addressing whether single large or several smaller fragments, equivalent in total area, contain (and therefore protect) more species (Diamond 1975; Abele and Connor 1979; Simberloff and Abele 1982; Ovaskainen 2002; Tjørve 2010). In a recent review of the fragmentation literature, Fahrig (2013) examined 14 studies addressing SLOSS directly. All studies reported equivalent or higher species richness within several smaller habitat fragments compared to fewer larger fragments or continuous habitat, suggesting that fragmentation does not reduce species diversity after habitat loss is controlled for. An additional study (Yaacobi et al. 2007) assessed patterns in species diversity on a fragmented agricultural landscape and reported similar findings.
Interestingly, studies supporting the habitat amount hypothesis (Yaacobi et al. 2007; Fahrig 2013) do not attempt to disentangle fragmentation effects on individual species, or differentiate between potentially reproducing species and transient species observed to occupy habitat fragments. While the sample area effect may best explain patterns of entire species assemblages on fragmented landscapes, assessments of fragmentation–species diversity relationships may be misleading if variation in fragmentation effects among species are not considered (Ewers and Didham 2006; Öckinger et al. 2009; Betzholtz and Franzén 2011; Franzén and Betzholtz 2012; Hanski 2015), and in particular, if potentially reproducing species are not distinguished from transient species temporarily occupying individual habitat fragments. Indeed, inter-fragment movements of highly mobile species from larger habitat fragments (supporting reproducing populations) to smaller fragments containing additional resources (e.g., Fretwell and Calver 1969; Dreisig 1995) have great potential to obscure fragmentation–species diversity relationships. Although several smaller and fewer (or single) large fragments may be observed to contain equivalent numbers of species, it should not be assumed that smaller fragments are capable of supporting viable populations in the absence of larger fragments present on the landscape.
In this study, we surveyed butterfly species diversity on lake islands in Sabaskong Bay, Lake of the Woods, Ontario, Canada. The tens of thousands of islands in Lake of the Woods represent an ideal system for decoupling habitat fragmentation from habitat loss to assess how habitat configuration relates to patterns of species diversity. Islands of Sabaskong Bay are remnant fragments of continuous habitat that was flooded 3000–4000 years ago in the early Subboreal period (Yang and Teller 2005), permitting an assessment of fragmentation effects on a landscape where insular biotas have likely relaxed to equilibria following habitat loss and fragment isolation (MacArthur and Wilson 1963, Wilson and MacArthur 1967; Haila 2002). Furthermore, akin to oceanic islands, habitat boundaries in this system are strictly delimited by water, and butterflies cannot utilize surrounding aquatic habitats at any life stage. This effectively controls for “matrix effects,” whereby the matrix of unsuitable habitat contributes to species diversity, or differentially facilitates the inter-fragment movements of individuals (Ricketts 2001; Haila 2002).
By differentiating between potential resident and transient butterfly species based on occurences of larval food plant species, we were able to investigate fragmentation effects on both the complete species assemblage and a subset of potential resident (reproducing) species. The detection of negative fragmentation effects in our study system would indicate that several smaller insular habitats do not support butterfly species diversity to the same extent as continuous habitat of equivalent area. Such a result may be attributed either to an island effect, or to decreases in habitat suitability or habitat diversity within smaller islands (Gotelli 1996). In contrast, support for the habitat amount hypothesis would indicate that insular and continuous habitats contribute equally to butterfly species diversity, and that fragment edges do not delimit butterfly populations at the scales addressed.
Methods
Study design and focal taxon
Thirty islands within Sabaskong Bay were selected to represent a nested-set sampling design that effectively decoupled habitat fragmentation from habitat loss (Table 1; Fig. 1). Specifically, islands were organized into two island sets that were used to assess fragmentation effects across two distinct ranges of island sizes. The first (small) island set contained 15 islands organized into four size classes, including eight 0.1-ha islands, four 0.2-ha islands, two 0.4-ha islands, and a single 0.8-ha island. The second (large) island set followed an identical pattern using islands ranging from 1.0 to 8.0 ha. By doubling the area of individual islands per twofold reduction in number of replicates, we were able to vary the degree of fragmentation across size classes while holding total habitat area constant. (See Electronic Supplementary Material [Appendix 1] for study island selection criteria.)

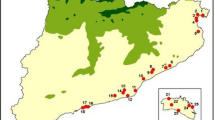
Map of the study area, located in Sabaskong Bay, Lake of the Woods, Ontario, Canada. All study islands were located within 20 km of the study camp, approximately 6 km northwest of Morson, Ontario
Butterflies were chosen as the focal taxon for this study because (1) they are commonly used as model organisms in ecology and data on their biology are widely available (Baguette and Van Dyck 2007), (2) the majority of butterfly species complete their lifecycles within relatively small patches of habitat (van Swaay et al. 2006; Nowicki et al. 2008), meaning butterfly diversity may serve as a proxy habitat suitability within islands, (3) butterflies cannot utilize aquatic habitats at any morphological stage, rendering the matrix of open water surrounding islands completely uninhabitable, and (4) butterfly occurrences representing potential resident (reproducing) and transient (non-reproducing) populations may be distinguished on individual islands by the presence or absence of their known larval food plants. Coupled with high detectability and established sampling methods (Pollard 1977), these traits make butterflies well-suited study organisms for assessing relationships between fragmentation and species diversity in our study system.
Survey methods
Butterfly species richness and abundance were estimated on each of the 30 islands through repeated full-island surveys, standardized to a survey time of 40 min per ha. This protocol ensured that sampling effort per unit area was consistent across islands of all sizes, eliminating the need for rarefaction, extrapolation, or other diversity corrections (Chao et al. 2014). Four rounds of butterfly surveys were completed by a single observer. Each island was visited at intervals between 10 and 14 days during peak flight season (from 01-June-2015 to 20-Aug-2015). Our survey protocol was similar to that outlined by Pollard (1977) to ensure that butterfly activity was optimal and consistent between surveys (see Electronic Supplementary Material [Appendix 1] for survey protocol details). Vascular plant species richness was surveyed by a second observer on all 30 islands using repeated vegetation surveys. Plant surveys were also standardized to a time of 40 min per ha, with four surveys completed per island (see Anderson et al. in review). Fourteen habitat classes, defined using vegetation and substrate characteristics, were used to quantify habitat diversity within each island (see Electronic Supplementary Material [Appendix 1; Table A1] for habitat class descriptions).
Analyses
Comparisons of species diversity
For each of the eight island size classes, we estimated the effective number of species using species richness, the exponential of the Shannon–Wiener index, and Simpson’s reciprocal index (Jost 2006; MacDonald et al. 2017; see Electronic Supplementary Material [Appendix 1] for index equations). We hypothesized that if fragmentation decreased butterfly diversity, diversity measures would be lowest within the smallest sizes class (representing higher degrees of fragmentation) and increase with larger size classes (representing lower degrees of fragmentation). Such a result would support the island effect hypothesis. Any other arrangement of species richness across size classes would suggest that fragmentation did not reduce species diversity, but would not necessarily support the habitat hypothesis. The habitat amount hypothesis specifically predicts that species diversity is unrelated to the number of habitat fragments when total habitat area is held constant. This prediction would equate to an even distribution of species diversity across size classes.
Species accumulation curves (SACs) and saturation index
For the two island sets separately and together, cumulative species richness was plotted against cumulative island area in two different ways: (1) increasing order of island size (small to large), and (2) decreasing order of island size (large to small). Slight variation in island area within size classes allowed for the sorting of islands in a reasoned manner. Data points were connected with straight lines to generate SACs, which were constrained to pass through the origin to allow for area-under-the-curve comparisons (Quinn and Harrison 1988; Gavish et al. 2012). Similar slopes between large-to-small and small-to-large SACs would suggest that species richness increased with cumulative area, irrespective of the number of fragments. Steeper slopes of large-to-small SACs would suggest that fewer/single larger islands contained more species, while steeper small-to-large SACs would indicate that several smaller islands contained more species.
Differences in slopes between large-to-small and small-to-large SACs were quantified using a saturation index (Quinn and Harrison 1988). This index was estimated as the area under the small-to-large SAC divided by that of the large-to-small SAC. To estimate area under the SACs, integrals were calculated using the trapezoidal rule. Saturation index estimates less than one would indicate negative fragmentation effects, which lend support to the island effect hypothesis. Index estimates equal to one would indicate that fragmentation did not affect species diversity, supporting the habitat amount hypothesis. Index estimates greater than one would indicate that fragmentation increased species diversity. This positive fragmentation effect is not predicted by either the island effect or habitat amount hypothesis, but may result from several smaller islands intersecting the distributions of more species than fewer or single larger island (Tjørve 2010; Fahrig 2013). Alternatively, several smaller islands may contain a higher diversity of habitat types than fewer or single larger island, which may support a higher diversity of species (Williams 1964; Nilsson et al. 1988).
SAR extrapolation and SLOSS index
For the two island sets separately and together, a log–log least-squares linear regression was applied to island area and species richness to attain a SAR (Yaacobi et al. 2007; Gavish et al. 2012). To allow the logarithmic transformation of a single 0.1-ha island with a species richness of zero, a constant of one was added to all richness values. Each SAR was extrapolated to the aggregate area of all islands used to build the SAR (small island set = 3.21 ha; large island set = 32.13 ha; both island sets together = 35.34 ha). Substituting this aggregate area into the SAR regression (and subtracting a constant of one to account for the original transformation) yielded a species richness estimate for continuous habitat equivalent in area to all study islands. This richness estimate (S sl) represents the single large conservation strategy, while the aggregate observed richness of study islands (S ss) represents the several small conservation strategy (Gavish et al. 2012).
To test whether estimated (S sl) and observed (S ss) richness values were significantly different, 95% confidence intervals were extrapolated for each of the SARs. If the SAR predicted a significantly higher number of species than observed (S sl > S ss), habitat fragmentation reduced species richness, supporting the island effect hypothesis. Conversely, if SAR regressions accurately predicted aggregate observed richness (S sl ≈ S ss), habitat configuration was not related to species richness, supporting the habitat amount hypothesis. If the SAR predicted a significantly lower number of species than observed (S sl < S ss), habitat fragmentation increased species richness. A SLOSS index, estimated as 100% × (S ss − S sl)/S ss, indicates the proportion of species richness of several small fragments relative to that of a single large fragment (Boecklen 1997; Gavish et al. 2012). For example, a SLOSS index value of 20% (or − 20%) would indicate that the study islands representing the several small conservation strategy (S ss) contained 20% more species (or 20% fewer species) from the species pool than continuous habitat representing the single large conservation strategy (S sl).
Area-independent effects on butterfly species diversity (GLMs)
Generalized linear models (GLMs) and an information theoretic approach (Burnham and Anderson 2004) were used to test for effects of island isolation, habitat diversity, vascular plant species richness, and island shape (relative habitat edge) on butterfly species richness. Negative binomial regressions were used to account for overdispersion within island butterfly species richness data (Ver Hoef and Boveng 2007). Variance inflation factors (VIFs) were used to test for collinearity among explanatory variables, with a value of 10 used as a maximum cutoff (Craney and Surles 2002). Models were ranked for support using the small sample size corrected Akaike’s Information Criterion (AIC c ), where smaller AIC c values indicate higher relative model support (Burnham and Anderson 2004). Coefficients from our best-supported GLMs were standardized to permit comparisons of the relative importance of island attributes in structuring butterfly species richness.
Island area (log-transformed) was included as a covariate in most models to control for the expected positive SAR. A univariate log (area) model therefore represented an “ecological null” model for assessing the relative effects of other island characteristics on butterfly species richness. To test for the effects of island isolation on butterfly species richness, we quantified the proportion of open water (1 − proportion landmass) within various buffer sizes calculated from island edges (250, 500, 1000, 2500, and 5000 m). This measure of isolation is independent from the area of specific study islands. Proportion of uninhabitable matrix surrounding fragments has been shown to be a stronger predictor of dispersal and fragment immigration than distance-based metrics, justifying this measure (Moilanen and Nieminen 2002; Tischendorf et al. 2003). Beyond the size and isolation of islands, habitat diversity and plant diversity may contribute to patterns in butterfly species richness. Habitat diversity is expected to positively relate to butterfly richness because butterfly species vary in their habitat requirements (i.e., the habitat diversity hypothesis, Williams 1964; Nilsson et al. 1988). Not unrelated, vascular plant species richness may serve as a proxy for larval food plant diversity, breadth and seasonal availability of nectar resources, and habitat diversity. Both habitat diversity and plant richness are expected to positively relate to island area (Nilsson et al. 1988; Gotelli 1996). However, habitat diversity and plant richness should make a statistical contribution to GLMs beyond the variation explained by island area if they contribute to patterns in butterfly richness (Gotelli 1996).
Within single islands, edges and interiors may differentially support butterfly populations. Edge effects therefore represent another causal mechanism that may affect patterns in butterfly species richness (Saunders et al. 1991; Murcia 1995; Stasek et al. 2008). To capture variability in the amount of island edge, independent of island area, a relative edge index was estimated as the perimeter of a given study island made relative to the perimeter of a theoretical island identical in size but perfectly circular in shape. Values approaching one represented islands with minimal habitat edge, with higher values indicating increased habitat edge. Negative relationships between the relative edge index and butterfly richness would indicate that the increased habitat edge associated with fragmentation reduced butterfly diversity.
Potential resident and transient butterfly occurrences
While multiple studies addressing entire species assemblages support the habitat amount hypothesis (Fahrig 2013), responses to fragmentation will vary between species (Henle et al. 2004; Ewers and Didham 2006). Within single taxa, such variability in responses is often linked to species’ mobility (Roland and Taylor 1997; Lens et al. 2002; Ewers and Didham 2006; Öckinger et al. 2009). In the case of lake islands, butterfly species of high mobility may utilize resources on small islands that do not contain their larval food plants, thereby temporarily contributing to observed species diversity without constituting reproducing populations.
To test this hypothesis, we first used GLMs (logit link) to relate the probability of observing a butterfly species on at least one island where their larval food plant was not detected to (1) species' wingspans (mm; Burke et al. 2011; Hall et al. 2014) and (2) a species mobility index generated by Burke et al. (2011). Species’ wingspans were log-transformed to improve model fit (Burke et al. 2011). The prevalence (number of occurrences) of both butterfly species and their larval food plants was controlled for in GLMs as covariates. We then classified butterfly species occurrences on all 30 study islands as either “potential resident” or “transient” based on the presence or absence of known larval food plants [larval food plant associations were compiled from records summarized by Hall et al. (2014) and Acorn and Sheldon (2017)]. We were not able to distinguish between potential resident and transient populations of Feniseca tarquinius based on food plant occurrences because larvae are known only to feed on woolly aphids (Eriosomatinae; Hall et al. 2014; Acorn and Sheldon 2017). Feniseca tarquinius was therefore excluded from the subset of potential resident species. Danaus plexippus, Vanessa virginiensis, V. cardui, and V. atalanta are migratory species and are not known to complete their life cycles within our study area (Hall et al. 2014; Acorn and Sheldon 2017), so were also excluded.
We proceeded to repeat our analyses (“comparisons of species diversity,” “species accumulation curves [SACs] and saturation index,” “SAR extrapolation and SLOSS index,” and “area-independent effects on butterfly species diversity [GLMs]”) using only the potential resident species subset of the complete species assemblage. Through this reanalysis, we were able to investigate whether inter-island movements of highly mobile species obscured fragmentation effects on potentially reproducing populations.
Spatial patterns in species turnover
To further investigate ecological mechanisms structuring butterfly diversity, 28 pairwise comparisons of butterfly species turnover (using the complete species assemblage), habitat turnover, plant species turnover, and inter-island distance were made between islands within the 0.1- and 1.0-ha size classes. This isolated relationships between the complete butterfly assemblage, the vascular plant assemblage, habitat composition, and the spatial distribution of islands, while holding habitat area constant. Butterfly species turnover, plant species turnover, and habitat turnover were estimated using the Jaccard pairwise dissimilarity index (Baselga and Orme 2012). We tested for relationships between butterfly species turnover and inter-island distance because several small islands spread over a larger spatial extant may intersect the distributions of more species than fewer or single larger island (sensu Tjørve 2010; Fahrig 2013). This effect of several small islands “sampling” a larger species pool due to their wider spatial distribution may therefore obscure fragmentation effects using SLOSS-based analyses. Failure to detect positive relationships between species turnover and inter-island distance would indicate that the spatial distribution of small islands did not confound SLOSS-based analyses. Mantel tests (999 permutations) were used to assess whether relationships were significant. All statistical analyses were performed in the program R (R Core Team 2017).
Results
Complete species assemblage: A total of 82 butterflies belonging to 13 species and 786 butterflies belonging to 33 species were observed on islands within the small and large island set, respectively (Table 2). Butterfly diversity of the small island set was almost perfectly nested within that of the large island set, with 12 of the 13 species observed in the small fragment also observed in the large island set. (See Electronic Supplementary Material for species occurrence/abundance data by island [Online Resource 1] and the documented range expansion of Euphyes dion [Appendix 1].)
Potential resident species subset: A total of 53 butterflies belonging to ten species and 684 butterflies belonging to 29 species were observed on islands within the small and large island sets, respectively. Potential resident butterfly diversity of the small island set was perfectly nested within that of the large.
Comparisons of species diversity
Complete species assemblage: Each of the two smallest size classes contained more butterfly species than the two largest size classes in the small island set (Table 2). While the exponential of the Shannon–Wiener index and Simpson’s reciprocal index did not exhibit this relationship, MacDonald et al. (2017) demonstrate that these indices may fail to capture variation in butterfly species diversity and caution their interpretation. Overall, effective numbers of species for the complete species assemblage showed no clear relationship to degree of fragmentation in the large island set.
Potential resident species subset: Effective numbers of species generally showed no clear relationship to island size class in either the small or large island set; the exception being the relatively high number of species in the largest size class (single 8-ha island; S = 21) compared with other size classes in the large island set.
Species accumulation curves (SACs) and saturation index
Complete species assemblage: When species accumulation was plotted against cumulative fragment area for the small island set, the small-to-large SAC lay considerably above the large-to-small SAC (saturation = 1.22; Fig. 2). Incongruently, slopes of SACs were quite similar for the large island set (saturation = 0.96) and for both island sets together (saturation = 0.98). These results indicate that fragmented habitat contained more butterfly species than continuous habitat in the small island set. Incongruently, fragmentation effects were neutral to weakly negative in the large island set. Most interestingly, the positive fragmentation effect observed between the small island set’s SACs was not preserved when both island sets were aggregated for SAC analysis.

Cumulative species richness relative to cumulative island area for the complete species assemblage (a, c, e) and the potential resident species subset (b, d, f). Accumulation of species richness occurs from the smallest to the largest island (small-to-large SAC; represented by closed circles connected by solid lines) and from the largest to smallest island (large-to-small; represented by closed triangles connected by dashed lines). Saturation index values are estimated as the area under the small-to-large SAC divided by that of the large-to-small SAC
Potential resident species subset: The small-to-large and large-to-small SACs were overlapping for the small island set (saturation = 1.03). This result indicates that fragmented and continuous habitat contained equivalent numbers of potential resident butterfly species at this scale. In contrast with this pattern, large-to-small SACs lay above small-to-large SACs for the large island set (saturation = 0.92) and both island sets together (saturation = 0.89). Such results indicate a negative fragmentation effect on potentially reproducing butterfly populations. Overall, saturation was lower for the resident species subset than for the complete species assemblage at all scales (small, large, and both island sets together).
SAR extrapolation and SLOSS index
Complete species assemblage: Compared to SAR species richness estimates for continuous habitat, we observed higher aggregate richness across study islands in the small island set (S sl = 6.14; S ss = 13; SLOSS index = 53%) and lower aggregate richness across study islands in the large island set (S sl = 64.42; S ss = 33; SLOSS index = − 95%; Fig. 3). When all 30 islands were combined for SAR extrapolation (both island sets), the SAR richness estimate for continuous habitat was very close to the aggregate richness observed across all study islands (S sl = 33.33; S ss = 34; SLOSS index = − 1%). For all three SAR extrapolations, aggregate richness observed across study islands fell within the extrapolated SAR’s 95% confidence intervals.

SAR extrapolations for the complete species assemblage (a, c, e) and the potential resident species subset (b, d, f). Solid and dashed lines represent log–log least-squares linear SAR regressions and their 95% confidence intervals, respectively. Area coefficients of log–log SAR regressions are reported as z values, approximating exponents of the species–area power model (S = cA z).Closed and open circles represent species richness for individual islands and their aggregate richness, respectively (a constant of one was added to all richness values to allow for log-transformations). Axes were back-transformed from logarithmic to linear scales for straightforward interpretation of species richness and area values. SLOSS index values were estimated as 100% × (S ss − S sl)∕S ss, where S ss represents the aggregate observed richness of study islands and S sl represents the SAR’s richness estimate for continuous habitat of equivalent areal extent
Potential resident species subset: Compared to SAR species richness estimates for continuous habitat, we observed approximately equivalent aggregate richness across study islands in the small island set (S sl = 9.55; S ss = 10; SLOSS index = 5%), lower aggregate richness across study islands in the large island set (S sl = 73.04; S ss = 29; SLOSS index = − 152%), and lower aggregate richness when both island sets were considered together (S sl = 40.20; S ss = 29; SLOSS index = − 40%). Again, however, aggregate richness observed across study islands fell within the extrapolated SAR’s 95% confidence intervals for all thee SAR extrapolations. Of particular interest, SLOSS index estimates were lower for the potential resident species subset than for the complete assemblage at all scales (small, large, and both island sets together). Furthermore, slopes of SARs (z values) were greater for the potential resident species subset than for the complete species assemblage at all scales.
Area-independent effects on butterfly species diversity (GLMs)
Complete species assemblage: The best-supported model explaining patterns of butterfly species richness across study islands accounted for island area and vascular plant richness, with both variables relating positively to butterfly richness (Tables 3, 4). Standardized coefficients indicate that island area had a greater effect than vascular plant richness on butterfly richness. The inclusion of a log (area) × plant richness interaction term decreased model support, indicating the relationship was consistent across islands sizes. After controlling for the positive relationship between island area and butterfly species richness, the inclusion of island isolation measures (proportion of open water within 250-, 500-, 1000-, 2500-, and 5000-m buffers) decreased model support in all cases. However, three isolation measures (250-, 5000-, and 500-m buffers, in descending order of support) decreased model support by less than two AIC c points, indicating isolation effects were uncertain. As both the island effect and habitat amount hypothesis predict, the proportion of open water within the best-supported buffer size (250 m) was negatively related butterfly richness. Model support was reduced with inclusion of habitat richness and the relative island edge index, suggesting that habitat diversity and edge effects did not contribute to patterns in butterfly species richness beyond the variation explained by island area.
Potential resident species subset: When transient butterfly species were excluded from the complete assemblage, island area and island isolation (250-m buffer) were found to best explain variation in potential resident butterfly richness. In this model, island isolation was negatively related to butterfly richness and had a greater effect than island area. Models accounting for other isolation buffers decreased model support by more than two AIC points, indicating they were not well-supported. Vascular plant richness decreased model support wherever included, suggesting it was not related to insular patterns in richness of potential resident species. The standardized effects of both island area and isolation on butterfly richness were always greater for the resident species than for the complete species assemblage. VIFs were less than 10 for all models, suggesting collinearity was not problematic (Craney and Surles 2002).
Potential resident and transient butterfly occurrences
Logistic regressions indicate that the probability of observing a butterfly species on at least one island without their larval food plants was positively related to wingspan (P = 0.011) and species mobility (P = 0.0075; Fig. 4). These relationships remained significantly positive after accounting for the prevalence (number of occurrences) of both butterfly species and their food plants.

The probability of observing butterfly species on at least one island where their larval food plant was not detected relative to (a) average wingspan (mm) and (b) the species mobility index (Burke et al. 2011). Species’ wingspans were log-transformed to improve model fit. Solid lines represent GLMs (logit link) used to assess relationships between variables. Relationships were significant for both average wingspan (P = 0.011) and species mobility (P = 0.0075)
Spatial patterns in species turnover
In accordance with GLMs addressing the complete species assemblage, Mantel tests indicated that butterfly species turnover was weakly positively related to plant species turnover in the 0.1- and 1.0-ha size classes (r = 0.26, P = 0.18 and r = 0.32, P = 0.13, respectively); although, these relationships were not significant. Contrasting with GLMs, butterfly species turnover and habitat turnover were significantly positively related in the 1.0-ha size class (r = 0.41, P = 0.045), but showed no strong relationship in the 0.1-ha size class (r = 0.10, P = 0.33). Butterfly species turnover was unrelated to inter-island distance in both the 0.1- and 1.0-ha size classes (r = 0.10, P = 0.35 and r = − 0.019, P = 0.55, respectively), indicating that the spatial distribution of several small islands did not contribute to their aggregate species richness.
Discussion
Fragmentation and species diversity
As predicted by the positive SAR, smaller islands were depauperate relative to larger islands at all scales addressed in this study. However, controlling for total habitat area demonstrated that this positive SAR is largely an artifact of the sample area effect: smaller sample areas contain fewer individuals, which for a given abundance distribution, belong to fewer species (Connor and McCoy 1979; Fahrig 2013). Direct comparisons of species richness across island size classes and SAC analyses of the complete species assemblage within the small island set suggest that several smaller islands actually contained more butterfly species than fewer larger islands, or a single large island of equivalent total area. Although this positive fragmentation effect was apparent in SAR extrapolation (S sl < S ss), the effect was not statistically significant, questioning the statistical power of the analysis. Most interestingly, this positive fragmentation effect was neutralized when transient butterflies were removed from analyses. Direct comparisons of species richness, SAC analysis, and SAR extrapolation all support the directionality of this pattern. This suggests that inter-island movements of highly mobile butterfly species, from larger island or mainland habitats with larval food plants to small islands without larval food plants, inflated the number of species small islands were observed to support. Positive relationships between both wingspan and mobility and the probability of observing butterfly species on islands without their larval food plants support this hypothesis.
To explain positive fragmentation effects in the context of the habitat amount hypothesis, Fahrig (2013) points out that several small fragments spread over a larger area are more likely to intersect the distributions of more species than single large fragments (e.g., Tjørve 2010). Given this relationship, the habitat amount hypothesis and underlying sample-area effect would predict positive relationships between degree of fragmentation and species diveristy when SLOSS-based analyses are used. However, pairwise comparisons of butterfly species turnover across the smallest islands in both island sets (0.1- and 1.0-ha islands) showed no relationship to pairwise comparisons of distance. It is therefore reasonable to conclude that islands within our study area “sampled” a spatially consistent species pool, and that the cumulative species richness of several small habitat fragments is generally unrelated to their spatial distribution at the scales addressed. A more likely explanation for positive fragmentation effects involves the inter-fragment movements of highly mobile species from larger habitat fragments, supporting reproducing populations, to smaller fragments, containing additional resources. Such movements are indeed predicted by ideal free distribution theory for nectar-feeding insects if smaller fragments contain higher densities of nectar resources (Dreisig 1995). Alternatively, several smaller islands may contain a higher diversity of habitat types compared with fewer or single larger islands, supporting a higher diversity of species (Williams 1964; Nilsson et al. 1988). However, GLMs accounting for both habitat diversity and habitat area do not support this latter hypothesis.
In contrast with the positive fragmentation effect observed in the small island set, the complete species assemblage shows no clear trend in species richness across island size classes in the large island set, supporting the habitat amount hypothesis. In partial conflict with this pattern, SAC analysis and SAR extrapolation both suggest a neutral to weakly negative fragmentation effect. Excluding transient species from the analyses revealed stronger negative fragmentation effects on potentially reproducing butterfly populations for all analyses. The largest island in the large island set (8 ha of continuous habitat) contained a higher number of potential resident species than other size classes (8 ha of fragmented habitat). Similarly, SAC analyses show that fewer or single larger islands contained more potential resident species than several smaller islands summing to an equivalent area. Excluding transient species from SAR extrapolation in the large island set made negative fragmentation effects more apparent, although still not statistically significant.
Two principal conclusions may be drawn: (1) negative fragmentation effects are more apparent in the large island set compared to the small, suggesting scale dependency, and (2) negative fragmentation effects are more apparent when excluding transient species that do not represent reproducing populations. This latter conclusion also holds true when both island sets were combined for SAC analysis and SAR extrapolation. We suspect that negative fragmentation effects observed across all 30 islands are likely a combined result of stochastic extinctions of populations isolated to smaller islands, as predicted by the island effect hypothesis, and decreased habitat suitability within these islands, such as the exclusion of potential larval food plants.
Implications of scale dependency
While SAC analysis may be the simplest method for archipelago- or landscape-wide comparisons of species diversity (Quinn and Harrison 1988), scale separation in our study shows that SACs have potential to obscure important aspects of fragmentation-species diversity relationships. We observed a considerable difference between small-to-large and large-to-small SACs for the complete species assemblage in the small island set, indicated by a saturation index estimate of 1.22. However, this pattern of species accumulation across small islands was not preserved when both island sets were aggregated for analyses, where the saturation index was estimated at 0.98.
Although the small and large island sets contained equivalent numbers of islands, cumulative island area was tenfold greater in the large island set than in the small island set. Similarly, aggregate species richness was approximately twice as high in the large island set than in the small island set for the complete species assemblage. When all 30 islands were combined for SAC analysis, constrained integrals show that the small island set contributed only 3.09% to the area under the small-to-large SAC and 12.32% to the area under the large-to-small SAC. This demonstrates that patterns of species accumulation across larger fragments have potential to dominate those across smaller fragments, particularly when the range of fragment sizes is great and the abundance of small fragments is high—an arrangement common to many datasets. Within single archipelagos and landscapes, island/fragment areas may vary by several orders of magnitude, with smaller islands/fragments typically more abundant than larger ones (Quinn and Harrison 1988; Lomolino and Weiser 2001; Fahrig 2003, 2013). In the context of habitat fragmentation, it should not be assumed that ecological patterns and processes are consistent across these scales (Johnson 1980; Lomolino and Weiser 2001).
Scale separation and the SAR
SAR richness estimates for continuous habitat did not significantly differ from the aggregate observed richness of study islands for any of the six SAR extrapolations. Such a result may be attributed to either neutral fragmentation effects, or lack of statistical power to resolve relationships. While previous studies report P-values for SAR slope estimates (e.g., Yaacobi et al. 2007; Gavish et al. 2012), this method does not provide for the meaningful discrimination of insignificant results. Extrapolating SAR confidence intervals to infer the significance of fragmentation effects is a novel approach that explicitly accounts for variation in statistical power between SAR extrapolations. As we observed in this study, confidence interval discrimination may increase the probability of a type II error (concluding that fragmentation was not related to diversity when in fact it was). However, this method reduces the possibility of type I error, because it accounts for uncertainty in SAR extrapolations stemming from small sample sizes or “noisy” species richness data. Reduced regression confidence and the subsequent broadening of confidence intervals likely explain why SAR extrapolation failed to detect (1) the positive fragmentation effect observed in the small island set for the complete species assemblage, and (2) the negative fragmentation effect observed in the large island set for the resident species subset. With other factors held constant, regression confidence increases with sample size—a relationship observed when all 30 islands were pooled for SAR extrapolation. However, when all 30 islands were included in the SAR, negative fragmentation effects were still not significant for the potential resident species subset. Contrasting with this result, SACs over all 30 islands demonstrate clearly that single or fewer larger islands contained more potential resident species than several small islands summing to an equivalent area. Accounting for regression confidence therefore brings to question the SAR extrapolation method’s statistical power to resolve fragmentation effects.
Further questioning the viability of the SAR extrapolation method, the aggregation of fragment sizes required to raise SAR regression confidence to levels sufficient for resolving fragmentation effects has the inherent potential to obscure scale-dependent relationships. Scale separation should be considered necessary when the range of fragment sizes within datasets is great, such that fragmentation–species diversity relationships shift in relation to fragment size. However, many datasets do not contain a sufficient number of fragments to allow for scale separation while maintaining an adequate sample size [e.g., Quinn and Harrison (1988) and Boecklen (1997), where datasets reviewed contained as few as six and five fragments, respectively]. Moreover, small sample sizes are often paired with broad ranges in fragment size [e.g., Rosin et al. (2011), where only 31 fragments ranged in size by over two orders of magnitude]. Such relationships also question whether z values should be used to infer fragmentation effects, as these analyses inherently assume uniformity in both SARs and fragmentation–species diversity relationships across broad ranges of fragment sizes.
Examples of shifts in SARs across island sizes are made clear by the small island effect (Lomolino and Weiser 2001), which states that insular species richness may not predictably increase with area below a threshold island or fragment size (Triantis et al. 2006). Below this threshold, species richness is largely determined by area-independent variables, such as intraspecific and interspecific interactions, stochastic events, island isolation, and habitat diversity (Nilsson et al. 1988; Lomolino 2000; Lomolino and Weiser 2001; Schoener et al. 2001; Triantis et al. 2006; Rosin et al. 2011). Area-independent variables influencing species richness add uncertainty to species–area regressions, further decreasing the probability of detecting fragmentation effects in SAR extrapolations or z values comparisons. Future studies should test for shifts in the SAR (e.g., Lomolino and Weiser 2001) before proceeding with such analyses. Theoretically, islands or fragments below the small island effect threshold should be excluded.
Area-independent relationships
The habitat amount hypothesis predicts that fragment isolation and species diversity will negatively correlate when mean isolation inversely relates to the amount of habitat on a landscape: if fragment edges do not delimit populations, negative relationships between fragment isolation and species richness may be an artifact of local species pools decreasing with habitat amount (Fahrig 2013). However, the habitat amount hypothesis would not predict strong relationships between isolation and species richness if the spatial distribution of fragments is small, such that there is little spatial variation in the composition or abundances of species within a study area. This appears to be the case in our study system, as Mantel tests show no relationship between inter-island distances and differences in composition of the complete butterfly assemblage. Lack of strong relationships between island isolation and species richness in GLMs using the complete species assemblage is therefore best interpreted as support for the habitat amount hypothesis. Incongruently, when only considering the potential resident species subset, island isolation (250-m buffer) was found to be the most important factor structuring patterns of species richness in our best-supported model. This result suggests that when highly mobile, transient butterfly species occurring on islands without their food plants are excluded from fragmentation analyses, island effects on potentially reproducing populations become apparent. This result brings into question the neutral to positive fragmentation effects reported by multiple studies cited to support the habitat amount hypothesis (Fahrig 2013), as well as those concluded by Yaacobi et al. (2007).
Our best-supported GLM explaining variation in butterfly species richness within the complete species assemblage included both island area and vascular plant species richness. As previously documented (e.g., Erhardt 1985; Sparks and Parish 1995; Simonson et al. 2001; Croxton et al. 2005; Kitahara et al. 2008; but see Hawkins and Porter 2003), plant richness and butterfly richness were found to positively relate. Plant diversity may positively relate to butterfly diversity through the intermediate variables of food plant availability (Hawkins and Porter 2003), nectar resource availability (Kitahara et al. 2008), or habitat diversity (sensu Williams 1964; Nilsson et al. 1988). Interestingly, when excluding transient butterfly species occurring on islands without their larval food plants, vascular plant richness was a poor predictor of butterfly richness. This difference suggests that the positive relationship between plant and butterfly richness within the complete species assemblage was driven by inter-island movements of highly mobile butterfly species to islands of particularly high plant diversity.
Conclusions
When considering the complete species assemblage, habitat fragmentation did not reduce butterfly species diversity in our study system. This result suggests that habitat configuration has little effect on the number of butterfly species persisting on fragmented landscapes, supporting the habitat amount hypothesis. However, butterfly species vary widely in mobility (Burke et al. 2011), and are therefore likely to vary widely in their responses to habitat fragmentation (Ewers and Didham 2006; Dover and Settele 2009). Our study shows that differentiating between potentially reproducing species and highly mobile, transient species observed within individual habitat fragments yields critical insight into the negative effects of habitat fragmentation on species diversity.
References
Abele L, Connor EF (1979) Application of island biogeography theory to refuge design: Making the right decision for the wrong reasons. In: Proceedings of the first conference of scientific research in the National Parks I, Washington, pp 89–94
Acorn J, Sheldon I (2017) Butterflies of Ontario and Eastern Canada. Partners Publishing, Fredericton
Baguette M, Van Dyck H (2007) Landscape connectivity and animal behavior: functional grain as a key determinant for dispersal. Landsc Ecol 22:1117–1129
Baselga A, Orme CDL (2012) Betapart: an R package for the study of beta diversity. Methods Ecol Evol 3:808–812
Betzholtz P, Franzén M (2011) Mobility is related to species traits in noctuid moths. Ecol Entomol 36:369–376
Boecklen WJ (1997) Nestedness, biogeographic theory, and the design of nature reserves. Oecologia 112:123–142
Burke RJ, Fitzsimmons JM, Kerr JT (2011) A mobility index for Canadian butterfly species based on naturalists’ knowledge. Biodivers Conserv 20:2273–2295
Burnham KP, Anderson DR (2004) Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res 33:261–304
Chao A, Gotelli NJ, Hsieh T, Sander EL, Ma K, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67
Connor EF, McCoy ED (1979) The statistics and biology of the species-area relationship. Am Nat 113:791–833
Craney TA, Surles JG (2002) Model-Dependent Variance Inflation Factor Cutoff Values. Quality Eng 14(3):391–403
Croxton P, Hann J, Greatorex-Davies J, Sparks T (2005) Linear hotspots? the floral and butterfly diversity of green lanes. Biol Conserv 121:579–584
Debinski DM, Holt RD (2000) A survey and overview of habitat fragmentation experiments. Conserv Biol 14:342–355
Diamond JM (1972) Biogeographic kinetics: estimation of relaxation times for avifaunas of southwest pacific islands. Proc Natl Acad Sci USA 69:3199–3203
Diamond JM (1975) The island dilemma: lessons of modern biogeographic studies for the design of natural reserves. Biol Conserv 7:129–146
Dover J, Settele J (2009) The influences of landscape structure on butterfly distribution and movement: a review. J Insect Conserv 13:3–27
Dreisig H (1995) Ideal free distributions of nectar foraging bumblebees. Oikos 161–172
Erhardt A (1985) Diurnal lepidoptera: sensitive indicators of cultivated and abandoned grassland. J Appl Ecol 22:849–861
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:117–142
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40:1649–1663
Franzén M, Betzholtz P (2012) Species traits predict island occupancy in noctuid moths. J Insect Conserv 16:155–163
Fretwell SD, Calver JS (1969) On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheor 19:37–44
Gavish Y, Ziv Y, Rosenzweig ML (2012) Decoupling fragmentation from habitat loss for spiders in patchy agricultural landscapes. Conserv Biol 26:150–159
Gonzalez A (2000) Community relaxation in fragmented landscapes: the relation between species richness, area and age. Ecol Lett 3:441–448
Gotelli NJG (1996) Null models in ecology. Smithsonian Institution Press, Washington, D.C.
Haddad NM, Gonzalez A, Brudvig LA, Burt MA, Levey DJ, Damschen EI (2017) Experimental evidence does not support the habitat amount hypothesis. Ecography 40:48–55
Haila Y (1986) On the semiotic dimension of ecological theory: the case of island biogeography. Biol Philos 1:377–387
Haila Y (1990) Toward an ecological definition of an island: a northwest European perspective. J Biogeogr 17:561–568
Haila Y (2002) A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecol Appl 12(2):321–334
Hall PW, Jones CD, Guidotti A, Hubley B (2014) The ROM field guide to butterflies of Ontario. Royal Ontario Museum, Toronto
Hanski I (2015) Habitat fragmentation and species richness. J Biogeogr 42:989–993
Harrison S, Bruna E (1999) Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography 22:225–232
Hawkins BA, Porter EE (2003) Does herbivore diversity depend on plant diversity? The case of California butterflies. Am Nat 161:40–49
He F, Hubbell SP (2011) Species-area relationships always overestimate extinction rates from habitat loss. Nature 473:368–371
Henle K, Davies KF, Kleyer M, Margules C, Settele J (2004) Predictors of species sensitivity to fragmentation. Biodivers Conserv 13:207–251
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71
Jost L (2006) Entropy and diversity. Oikos 113:363–375
Kitahara M, Yumoto M, Kobayashi T (2008) Relationship of butterfly diversity with nectar plant species richness in and around the Aokigahara primary woodland of Mount Fuji, central Japan. Biodivers Conserv 17:2713–2734
Lens L, Van Dongen S, Norris K, Githiru M, Matthysen E (2002) Avian persistence in fragmented rainforest. Science 298:1236–1238
Lomolino MV (2000) Ecology’s most general, yet protean pattern: the species-area relationship. J Biogeogr 27:17–26
Lomolino M, Weiser M (2001) Towards a more general species-area relationship: Diversity on all islands, great and small. J Biogeogr 28:431–445
MacArthur RH, Wilson EO (1963) An equilibrium theory of insular zoogeography. Evolution 17:373–387
MacDonald ZG, Nielsen SE, Acorn JH (2017) Negative relationships between species richness and evenness render common diversity indices inadequate for assessing long-term trends in butterfly diversity. Biodivers Conserv 26:617–629
Mendenhall CD, Karp DS, Meyer CF, Hadly EA, Daily GC (2014) Predicting biodiversity change and averting collapse in agricultural landscapes. Nature 509:213–217
Moilanen A, Nieminen M (2002) Simple connectivity measures in spatial ecology. Ecology 83:1131–1145
Murcia C (1995) Edge effects in fragmented forests: implications for conservation. Trends Ecol Evol 10:58–62
Nilsson SG, Bengtsson J, As S (1988) Habitat diversity or area per se? Species richness of woody plants, carabid beetles and land snails on islands. J Anim Ecol 57:685–704
Noss RF (1991) Landscape connectivity: different functions at different scales. In: Hudson WE (ed) Landscape linkages and biodiversity. Island Press, Washington, DC, pp 27–39
Nowicki P, Settele J, Henry P, Woyciechowski M (2008) Butterfly monitoring methods: the ideal and the real world. Isr J Ecol Evol 54:69–88
Öckinger E, Franzén M, Rundlöf M, Smith HG (2009) Mobility-dependent effects on species richness in fragmented landscapes. Basic Appl Ecol 10:573–578
Ovaskainen O (2002) Long-term persistence of species and the SLOSS problem. J Theor Biol 218:419–433
Pollard E (1977) A method for assessing changes in the abundance of butterflies. Biol Conserv 12:115–134
Quinn JF, Harrison SP (1988) Effects of habitat fragmentation and isolation on species richness: evidence from biogeographic patterns. Oecologia 75:132–140
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158:87–99
Roland J, Taylor PD (1997) Insect parasitoid species respond to forest structure at different spatial scales. Nature 386:710
Rosin ZM, Skórka P, Lenda M, Moron D, Sparks TH, Tryjanowski P (2011) Increasing patch area, proximity of human settlement and larval food plants positively affect the occurrence and local population size of the habitat specialist butterfly Polyommatus coridon (Lepidoptera: Lycaenidae) in fragmented calcareous grasslands. Eur J Entomol 108:99–106
Saunders DA, Hobbs RJ, Margules CR (1991) Biological consequences of ecosystem fragmentation: a review. Conserv Biol 5:18–32
Schoener TW (1976) The species-area relation within archipelagos: models and evidence from island land birds. In: Proceedings of the sixteenth international ornithology congress, pp 629–642
Schoener TW, Spiller DA, Losos JB (2001) Natural restoration of the species-area relation for a lizard after a hurricane. Science 294:1525–1528
Simberloff D, Abele LG (1982) Refuge design and island biogeographic theory: effects of fragmentation. Am Nat 120:41–50
Simonson SE, Opler PA, Stohlgren TJ, Chong GW (2001) Rapid assessment of butterfly diversity in a montane landscape. Biodivers Conserv 10:1369–1386
Sparks T, Parish T (1995) Factors affecting the abundance of butterflies in field boundaries in Swavesey fens, Cambridgeshire, UK. Biol Conserv 73:221–227
Stasek DJ, Bean C, Crist TO (2008) Butterfly abundance and movements among prairie patches: the roles of habitat quality, edge, and forest matrix permeability. Environ Entomol 37:897–906
Tischendorf L, Bender DJ, Fahrig L (2003) Evaluation of patch isolation metrics in mosaic landscapes for specialist vs. generalist dispersers. Landscape Ecol 18:41–50
Tjørve E (2010) How to resolve the SLOSS debate: lessons from species-diversity models. J Theor Biol 264:604–612
Triantis K, Vardinoyannis K, Tsolaki E, Botsaris I, Lika K, Mylonas M (2006) Re-approaching the small island effect. J Biogeogr 33:914–923
van Swaay C, Warren M, Loïs G (2006) Biotope use and trends of European butterflies. J Insect Conserv 10:189–209
Ver Hoef JM, Boveng PL (2007) Quasi-poisson vs. negative binomial regression: how should we model overdispersed count data? Ecology 88:2766–2772
Williams CB (1964) Patterns in the balance of nature. Academic Press, London
Wilson EO, MacArthur RH (1967) The theory of island biogeography. Princeton University Press, Princeton
Wilson EO, Willis EO (1975) Applied biogeography. Ecol Evol Communities 522:534
Yaacobi G, Ziv Y, Rosenzweig ML (2007) Habitat fragmentation may not matter to species diversity. Proc Biol Sci 274:2409–2412
Yang Z, Teller JT (2005) Modeling the history of Lake of the Woods since 11,000 cal yr BP using GIS. J Paleolimnol 33:483–497
Acknowledgements
We wish to extend special thanks to the staff and owners of Redwing Lodge at Lake of the Woods, Ontario for their hospitality and facilitation of field activates. We also thank the staff at J & J General Store, Morson, Ontario for their hospitality and assistance with field communications. For assistance with field preparations, study analyses, and manuscript review, we thank Meghan Dauphinee, Andreas Hamann, Sylvia Horning, Victoria Masquillier, Evelyn Merrill, Federico Riva, and Felix Sperling.
Author information
Authors and Affiliations
Contributions
ZGM, JHA, and SEN formulated the idea, all authors contributed to sampling methods and experimental design, ZGM and IDA conducted field work (island surveys) and acquired spatial data, ZGM completed all analyses presented, and all authors contributed to the writing of this manuscript.
Corresponding author
Ethics declarations
Funding
This study was funded by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (Grant number 2014-04842).
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable institutional and/or national guidelines for the care and use of animals were followed.
Additional information
Communicated by Konrad Fiedler.
Here we use butterfly assemblages on lake islands to test the recently proposed habitat amount hypothesis, which negates direct effects of habitat fragmentation on species diversity. By differentiating between potentially reproducing and transient butterfly species on individual islands based on larval food plant occurences, our study shows that inter-fragment movements of highly mobile species may inflate the observed diversity of small habitat fragments, obscuring important fragmentation effects on resident species diversity.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
MacDonald, Z.G., Anderson, I.D., Acorn, J.H. et al. Decoupling habitat fragmentation from habitat loss: butterfly species mobility obscures fragmentation effects in a naturally fragmented landscape of lake islands. Oecologia 186, 11–27 (2018). https://doi.org/10.1007/s00442-017-4005-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-017-4005-2