
Abstract
Knowing how species’ traits relate to processes that underlie occupancy patterns such as colonisation and population persistence, is important for our understanding of how species survive in fragmented and changing landscapes. We used automatic UV light-traps to sample noctuid moths on two remote islands, and compared traits of island occupants with those of a species pool from mainland southeast Sweden. Widely distributed species, generalist species, species with a long adult activity period and species active late in the summer had higher probability of occupancy on the remote islands. The results were consistent between islands. The traits of host plant specificity and species with an adult activity period during late summer remained robust and were statistically significant after controlling for any possible phylogenetic bias. This indicates that species exhibiting those traits survive better when habitat and climate changes. It is crucial to include our results in; (1) conservation planning, e.g. when devising conservation measures in fragmented landscapes; (2) for predictions of future occupancy patterns; and (3) ecosystem impact assessments, e.g. the importance of moths as pollinators, herbivores and being the functional link between parasitoids, plants, consumers and predators.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Species survival in fragmented landscapes is influenced by dispersal, colonisation success, and population persistence (Fahrig and Merriam 1994). A general hypothesis is that dispersal ability and width of environmental tolerance, which is related to habitat availability, will largely influence where a species occurs (Hanski 1999; Hill et al. 2001). Thus, with the increasingly fragmented distribution of many key habitats, species communities might be expected to become dominated by widely distributed habitat generalists, which are usually strong dispersers (Warren et al. 2001; Hill et al. 2002; Mattila et al. 2006, 2009). Whereas several studies have addressed the relationship between life-history traits and population trends in butterflies (Kotiaho et al. 2005; Öckinger et al. 2006; Wenzel et al. 2006; Pöyry et al. 2009) and carabid beetles (Desender et al. 2010), the relations between species traits and occupancy patterns have yet to be explored for many species groups.
Data on occupancy patterns from isolated true islands can be important for improving our understanding of the relationship between species’ traits and those processes that determine occupancy patterns. It can be advantageous to study island communities because they are easier to define and they are surrounded by a homogenous matrix of water, whereas mainland communities are embedded within a more heterogeneous matrix, which may potentially bias the results (Bergman et al. 2004). A number of recent studies have explored butterfly populations on islands with respect to climate, rarity, range and endemism (Dapporto and Dennis 2008; Dennis et al. 2008, 2010). Several underlying factors are already known to affect population dynamics on islands, e.g. area, latitude, altitude, isolation, geology, biotope availability, land use, management and age (MacArthur and Wilson 1967), and such background knowledge lends further support to the use of true islands as a suitable study system for investigating occupancy patterns in relation to species traits.
Moths constitute a large proportion of the Lepidoptera (Scoble 1988), and are an important food resource for many birds and bats. However, the patterns, rates and causes of movements, colonisation ability and population persistence in moths are very poorly known (Woiwood and Stewart 1990; New 2004). Because moths have a high ecological, economic and conservation significance, it is important that we understand their occupancy patterns (Shreeve 1995). In the present study we investigate occupancy patterns, and record moth species on two remote islands, using automatic light-traps. We explore if the traits of body size, larval host plant specificity, habitat preferences, distribution area, length of flight period, and adult activity period, can be used to predict island occupancy in noctuid moths. We compare the traits of moths recorded on the two islands with those of a mainland species pool of assumed potential colonisers, in order to explore the patterns that lay behind island occupancy.
Recent studies have shown that large species (Nieminen et al. 1999; Gathmann and Tscharntke 2002; Öckinger et al. 2010), and species with large distribution areas (Mace and Kershaw 1997), are strong dispersers and are potentially more able to reach isolated islands. Species with a long flight period should have a greater chance of colonising such islands, as well as species that are active in late summer when high temperatures enhance movements (Sparks et al. 2005, Betzholtz and Franzén in press). Finally, several studies have shown a recent decline among specialist species (Kotiaho et al. 2005; Nilsson et al. 2008; Pöyry et al. 2009), suggesting that such species are unlikely to reach, or become successfully established, on isolated islands. Therefore, we expect large species, species with a large distribution area, species with a long flight period, species active in late summer, and habitat generalists as well as host plant generalists, to be over-represented on the studied islands.
Materials and methods
Study area and design
Utklippan is a small island (9 ha) of primary rocks, situated at the southeast corner of the Karlskrona archipelago in southeast Sweden (Fig. 1). The island is situated 8 km from the archipelago and 16 km from the province of Blekinge on the Swedish mainland. The vegetation on Utklippan is sparse, restricted to crevices in the rocks, with only a few isolated bushes and trees (Mattsson 1976). Gotska Sandön is an island of 3600 ha, situated 40 km north of the Baltic island Gotland and about 100 km east of the Swedish mainland (Fig. 1). The major part of the island is covered with pine forest. Calluna vulgaris dominates the lower vegetation. Areas with high flower richness are rare, and sand-dunes form the perimeter of the island. Only small patches with deciduous forests, shrubs and grassland habitats occur on the island (Lindgren 1968; Jonasson 2002). On both islands, the local climate is harsh compared to the mainland of Sweden, being characterised by lower temperatures, stronger winds and salt spray.

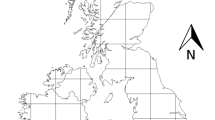
Map of the study area. The two remote islands, Gotska Sandön and Utklippan, are denoted by crosses. Species recorded in any of the provinces Scania, Blekinge, Småland, Öland, Gotland, Östergötland and Södermanland constitute the potential species pool
The species pool of assumed potential colonisers (n = 360) was defined as those species occurring in any of the surrounding provinces Scania, Blekinge, Småland, Öland, Gotland, Östergötland and Södermanland in southeast Sweden (Fig. 1). Besides being the most probable origin of the species recorded on the islands, the moth fauna of this area, and their distribution, is well documented (Svensson et al. 1994). We therefore consider the demarcation of the species pool to southeast Sweden as a conservative measure in our analyses. Furthermore, the selected species pool is strongly correlated to that of other areas surrounding the Baltic Sea. All of the selected noctuid moths are similarly attracted to UV light-sources (Scoble 1988).
We used automatic Ryrholm light-traps (Söderman 1994) with a mercury vapour lamp (125 W Hg-bulb). On Utklippan, one light-trap was operated from May to October during 2004–2006; on Gotska Sandön, two light-traps were operated from May to October 2008. The Ryrholm light-trap is highly efficient at capturing noctuid moths, and it has been shown to be more effective in detecting species than other kind of light-traps used in Finland and Great Britain (Leinonen et al. 1998). Data from another locality in southern Sweden where light-traps were used has shown that at least 85% of the total numbers of recorded species were found in 1 year (M. Franzén, unpublished data). Moreover, because light-traps are operated during all the hours of darkness, regardless of weather, they are particularly effective tools in scientific studies and surveys. Surprisingly, even though the Macrolepidopteran fauna of Gotska Sandön was considered to be relatively well known before this study (Jonasson 2002; Lindeborg 2009), 59 Noctuidae species from the light-trap catches in 2008 were recorded as being new to the fauna of the island.
The phylogeny of the Noctuidae is not resolved (Fibiger 1993) and any correction for phylogenetic independence (Harvey and Pagel 1991; Harvey and Purvis 1991) using an unresolved phylogeny may result in an unreliable analysis. Because the Noctuidae taxa are very diverse at the generic level compared to other families of Macrolepidoptera, we expect the phylogenetic dependence to be low. However, we have explored the possibility of phylogenetic dependence in the traits by analysing traits at both the species and generic levels. We argue that our results are robust and non-biased if the results are similar both at species and generic levels. Sibling species were treated according to our current knowledge of their ecology: Diachrysia tutti/D. chrysitis and Hadena bicruruis/H. capsincula were treated as separate species; Mesapamea secalis/M. didyma and Euxoa tritici/E. nigrofusca/E. crypta were pooled. The systematics follow Karsholt and Razowski (1996).
Analysed variables
Presence/absence data
From the defined species pool, we used the presence or absence of each species and genus, on each island, as the dependent variable.
Body size
We used the male wingspan (mm) as presented in Skou (1991) and Emmet (1991) as a proxy for body size. We arbitrarily decided to use the size of males, but since the size of males and females is strongly correlated (e.g. Mattila et al. 2008) this is unlikely to affect our results. At the generic level, the mean wingspan of member species was used.
Host plant specificity
We classified the larval host-plant specificity into three classes: monophagous species that feed mainly on a single plant species; oligophagous species that feed on a few plant species (less than or equal to five), or which are restricted to a particular genus or family of plants; and polyphagous species that feed on several different plant species (six or more) or genera. Information about food plants was extracted from Emmet (1991), Svensson (1993) and Huldén et al. (2000). When such information was not consistent in the literature, the information stated in Huldén et al. (2000) was used because this reference contains the most reliable information about life-history traits for the present study, being based on extensive studies of the Finnish fauna, adjacent to Sweden. The number of host plants used by each genus was summarized and classified as stated above.
Habitat preference
Each species was classified according to its preferred habitat in a broad sense. The following habitat classes were used: habitat generalists (including species occurring in all habitats); those of open habitats (grasslands and other open areas); and those of deciduous forest and coniferous forest habitats. The information of habitat preferences was extracted from Emmet (1991), Skou (1991) and Huldén et al. (2000) with some complementary data from Svensson (1993). When different habitat preferences were stated for the same species in different sources the information in Huldén et al. (2000) was used. For each genus, the dominant habitat of the species was selected. In the classification for the analysis at the generic level, a genus was classified as a generalist if it contained at least one generalist species.
Distribution area
For each species, we extracted the number of European countries from which it had been recorded, according to Karsholt and Razowski (1996). We used the number of European countries because this is the most homogenous data set available concerning species distributions. For each genus we extracted the number of European countries from which the genus has been recorded.
Length of flight period
We used the average length of the flight period in weeks in southern Sweden according to Svensson (1993). For species with two generations, we summed the flight periods because the longer the flight period, the greater the chance for a movement to an island to occur. No multivoltine species occur in the study area. For each genus the mean length of flight period was used.
Adult activity period
We categorised species according to adult activity period (following Svensson 1993) into late summer species and others. Late summer species were restricted to the period from August to September; others to the period from May to July. For each genus, we classified a genus as being active during late summer as stated above.
Island
To control for possible island effects we included island (Gotska Sandön and Utklippan) as a factor in the statistical analysis.
Number of species in the genus
The number of species in each genus was used as a covariate to control for possible bias. Species-rich genera might influence the trait classification because the presence of one generalist species in a genus indicates it as being a generalist genus, or the presence of one species with a large distribution as being a genus with a large distribution area. The variable ‘number of species in each genus’ was only used in the analysis at the generic level.
Statistical analyses
We compared species traits of noctuid moths recorded on two remote islands in the Baltic Sea, with the traits of the species pool of assumed potential colonisers in southeast Sweden. First, we explored possible collinearity between the analysed variables using Pearson correlations (two tailed significance). According to Agresti and Finlay (1986) the independent variables were not strongly correlated (Supplementary material 1). Thus we judged our analysed traits to be independent. The relationship between occurrence data and traits was analysed in a multiple logistic regression model including all two-way interactions with presence or absence of the moth species on the two islands as the binary response variable. We included island as a factor in the analyses in order to determine if differences between the two islands affected our results. We used a backward elimination process (log likelihood ratio statistics) and excluded parameters with the highest P-value for likelihood ratios tests until the final most parsimonious model contained the maximum number of significant variables at α = 0.05. All variables that were included in a significant interaction were also included as main effects. The final model was compared with competing models by using Akaikes information criteria. To disentangle interactions in the final model, we split the data with the categorical variable in the interaction and performed a second multiple logistic regression. Significant categorical variables in the final model were explored by a multinomial logistic regression. Using the same procedure as described above, we also used presence/absence of each genus as a dependent variable to control for possible phylogenetic bias. In this analysis, the number of species in each genus was included as a covariate to control for possible bias from differences in the number of species in the genus. All calculations were performed with SPSS (version 17.0 for Windows; SPSS, Chicago, Illinois).
Results
148 species (87 genera) were recorded on Gotska Sandön, and 98 species (58 genera) on Utklippan. 72 species and 19 genera occurred on both islands, giving a total of 174 (99 genera) separate species recorded on the two islands from the potential mainland pool of 360 species (160 genera). The traits, host plant specificity, habitat preference, distribution area, length of flight period, adult activity period and island, all predicted occupancy by noctuid moths (Table 1). There was also an effect of the interaction between length of flight period and habitat preference.
Species with a longer flight period (7.51 ± 0.19 weeks) occurred to a significantly greater extent on the remote islands compared to species with a shorter flight period (6.33 ± 0.12) (Table 1). This result was not consistent among the four habitat categories. Species with a longer flight period, that were habitat generalists or associated with coniferous forests, had a significantly greater island occupancy compared to species associated with open habitats or deciduous trees, which explains the interaction (Fig. 2; Table 2). Polyphagous species had a significantly greater occurrence on the islands than oligo-or monophagous species (Fig. 3; Table 3). Species with a large European distribution (28.4 ± 0.33 countries) occurred to a significantly greater extent than species with a more restricted distribution (25.3 ± 0.29) (Table 1). Furthermore, species with an adult activity period from August to September had a significantly greater occurrence compared to species with an adult activity period from May to July (Table 3). In the analysis controlling for any possible phylogenetic bias, the traits of host plant specificity, and species with an adult activity period during late summer, remained statistically significant (Table 1). There was no significant effect of body size.

The interaction between the length of the flight period (in weeks) and habitat preference. Habitat generalists or species associated with coniferous forest had a significantly greater island occupancy compared to species associated with open habitats or deciduous woods. Filled circles denote species present; open circles denote species absent. Error bars indicate the 95% confidence interval

Probability of island occupancy in noctuid moths in relation to host plant specificity. Mono Monophagous, Oligo Oligophagous, Poly Polyphagous. Error bars indicate the 95% confidence interval
Discussion
A number of other studies have investigated butterflies on islands in relation to traits, but little attention has been given to moths (Lindroth et al. 1973; Dennis et al. 2000, 2010; Dapporto and Dennis 2009, 2010). We found that host plant specificity, distribution area, adult activity period, and the interaction between length of flight period and habitat preference, all predicted island occupancy in noctuid moths (Table 1). This indicates that these traits influence the processes that lay behind occupancy patterns, i.e. dispersal, colonisation and population persistence. The factor ‘island’ also affected the occupancy pattern, but since ‘island’ was not included in any interaction it only reflects the fact that the recorded species differed between islands, which is consistent with the concept of island biogeography (MacArthur and Wilson 1967). Even though the two islands differ in area, distance from closest potential species pool, and suitability for breeding, the significant traits were consistent between islands, indicating a more general occupancy pattern in noctuid moths. However, our results contradict previous studies showing that the probability of occurrence on islands may vary greatly in closely related organisms with superficially similar traits (Dapporto and Dennis 2008).
We found greater island occupancy in polyphagous moths (Fig. 3). One possible explanation is that polyphagous moths can utilise more resources (host plants) than oligo- and monophagous species (Woiwood and Stewart 1990; Nieminen et al. 1999; Dennis et al. 2004), and that they are more abundant and widespread than specialist species (Inkinen 1994). Monophagous species often occupy relatively small proportions of the ranges of the host they consume (New et al. 1995; León-Cortés et al. 2003; Hardy et al. 2007). No relationship between host plant specialisation and occupancy has been found among butterflies on British islands (Dennis and Shreeve 1997). Instead, it is the distribution of the host plant that is important, and butterflies occupying islands often had widespread host plants. Indeed, it might not be possible to generalise from conclusions drawn from butterfly studies, because the proportion of widespread species specialising on widespread host plant species, such as Urtica dioica, is much larger in butterflies (>5% of the community in southern Sweden; modified from Svensson 1993) than in noctuid moths (<1% of the community in southern Sweden; modified from Svensson 1993). One theory is that the population variability is larger and more synchronised among mono- and oligophagous species, which might reduce population persistence (Dennis et al. 2004). Several host plants can provide insurance for population survival by providing a broader range of microsites for the pre-imaginal stages and hence, can buffer species against a variety of climatic extremes (Benton et al. 2003; Dennis et al. 2004; Piha et al. 2007; Dover and Settele 2009), as well as against mortality caused by parasites, viruses and fungi (Wilson and Roy 2009).
Dennis and Shreeve (1997) showed that the incidence of butterflies on British offshore islands is related to the width of their geographical ranges. It can be expected that wide-ranging species occur to a larger extent on the studied islands compared with species having a more restricted distribution (Gaston et al. 2000). We found that moths with a large distribution area had a greater island occupancy compared to moths with a more restricted distribution area. Further, the probability of occupancy was greater for moths active during late summer compared to species active in other periods (Table 3). Higher temperatures during late summer might enhance mobility (Betzholtz and Franzén in press) and explain the greater occupancy by these species (cf. Sparks et al. 2005, 2007). Species with a long adult activity period had greater island occupancy than species with a short activity period. The probability of coinciding movement with suitable conditions increases with a longer adult life span.
In our study the length of the flight period affected island occupancy and differed among habitat categories, explaining the significant interaction (Fig. 2; Table 1). Habitat generalists or species associated with coniferous forests had a significantly greater occupancy of islands compared to species associated with open habitats or deciduous woods. Habitat generalists may use a larger proportion of resources and may therefore move around more than habitat specialists (Brown 1984). Species associated with coniferous forests may also benefit from the increased areas of coniferous forest plantations in southern Sweden, as well as from enhanced possibilities for movements from there being more conifer woodland available, and hence a greater pool of potential colonisers (cf. Conrad et al. 2004). The low proportion of recorded species associated with open land suggests that the loss of open land that has occurred during the last 50 years has had a severe impact on the numbers of populations of species associated with this habitat. We are aware of the problems caused by categorising species according to certain habitats even though they may occupy and use resources in different habitats (e.g. nectar and host plants) (cf. Dennis et al. 2003). In the present study we use habitat in its broad sense as being synonymous with biotope. An investigation of the different resources utilized by the different species goes beyond the scope of the present study, but based on their larval development, noctuid species are relatively easy to classify as having an affinity to one of the four habitat types used in this study. Further steps could be taken to analyse the fauna using a finer habitat resolution, and so take into account the behavioral component of how species utilize different resources (cf. Shreeve et al. 2001; Dennis 2010; Shreeve and Dennis 2011).
We found no relation between body size and island occupancy. Nieminen et al. (1999) did not find any relationship between body size and island occupancy in noctuid moths either, although they did show a positive correlation with dispersal. This result is consistent with a recent study on mobility in noctuid moths, which also found no relation between mobility and body size (Betzholtz and Franzen in press). This indicates that processes other than dispersal are likely to influence island occupancy, e.g. a successful colonisation, and persistence on the island (c.f. Dennis and Shreeve 1997; Cook et al. 2001). The difference between the results of the analyses at the species and generic levels was small; but habitat preference, distribution area, and length of flight time were not significant in the generic analysis. These three variables are strongly influenced by the number of species in the genus, hence this finding is not only dependent on phylogeny. Because of the high number of genera in the Noctuidae, and the variable number of species in each genus, the phylogenetic dependence in traits can be expected to be of minor importance here. Moreover, phylogenetic dependence is mainly found at the taxonomic levels of order and family (Stearns 1983), and rarely within families (Mattila et al. 2008).
Implications for conservation
Species exhibiting the traits we found to be related to island occupancy will have greater possibilities of colonizing, and persisting, on islands and habitat fragments. Therefore generalist species, species with large distributions, and species active during late summer will have higher survival probabilities in fragmented and changing landscapes compared with specialist species, species with small distribution areas, and species not active late in the summer. There is a widespread and ongoing decline of moths in northwestern Europe (Conrad et al. 2006; Franzén and Johannesson 2007; Groenendijk and Ellis 2010). In Sweden 70 noctuid species (16% of all noctuids) are red-listed (Gärdenfors 2010), but on the studied island only four red-listed species were found (1.6% of the fauna), and clearly the red-listed species are under-represented on the studied islands. Hence, the findings of this study, that certain traits can be used to predict which species do or do not have the capacity to occupy remote islands, could be very useful when planning conservation measures.
References
Agresti A, Finlay B (1986) Statistical methods for the social sciences. Prentice Hall, New York
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Bergman KO, Askling J, Ekberg O, Ignell H, Wahlman H, Milberg P (2004) Landscape effects on butterfly assemblages in an agricultural region. Ecography 27:619–628
Betzholtz PE, Franzen M (201X) Mobility is related to species traits in noctuid moths. Ecol Entomol. doi:10.1111/j.1365-2311.2011.01281.x
Brown JH (1984) On the relationship between abundance and distribution of species. Am Nat 124:255–279
Conrad KF, Woiwood IP, Parson MS, Fox R, Warren MS (2004) Long-term population trends in widespread British moths. J Insect Conserv 8:119–136
Conrad KF, Warren MS, Fox R, Parsons MS, Woiwood IP (2006) Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol Conserv 132:279–291
Cook LM, Dennis RLH, Hardy PB (2001) Butterfly-hostplant fidelity, vagrancy and measuring mobility from distribution maps. Ecography 24:497–504
Dapporto L, Dennis RLH (2008) Species richness, rarity and endemicity on Italian offshore islands: complementary signals from island-focused and species-focused analyses. J Biogeogr 35:664–674
Dapporto L, Dennis RLH (2009) Conservation biogeography of large Mediterranean islands. Butterfly impoverishment, conservation priorities and inferences for an ecologica “island paradigm”. Ecography 32:169–179
Dapporto L, Dennis RLH (2010) Skipper butterfly impoverishment on large Mediterranean islands (Lepidoptera Hesperiidae): deterministic factors and stochastic events. Biodivers Conserv 19:2637–2649
Dennis RLH (2010) A resource-based habitat view for conservation. Butterflies in the British landscape. Wiley-Blackwell, Oxford
Dennis RLH, Shreeve TG (1997) Diversity of butterflies on British islands: ecological influences underlying the roles of area, isolation and the size of the faunal source. Biol J Linn Soc 60:257–275
Dennis RLH, Donato B, Sparks TH, Pollard E (2000) Ecological correlates of island incidence and geographical range among British butterflies. Biodivers Conserv 9:343–359
Dennis RLH, Shreeve TG, van Dyck H (2003) Towards a functional resource-based concept for habitat: a butterfly biology viewpoint. Oikos 102:417–426
Dennis RLH, Hodgson JG, Grenyer R, Shreeve TG, Roy DB (2004) Host plants and butterfly biology. Do host plant strategies drive butterfly status? Ecol Entomol 29:12–26
Dennis RLH, Dapporto L, Shreeve TG, John E, Coutsis JG, Kudrna O, Saarinen K, Ryrholm N, Williams WR (2008) Butterflies of European islands: the implications of the geography and ecology of rarity and endemicity for conservation. J Insect Conserv 12:205–236
Dennis RLH, Dapporto L, Sparks TH, Williams SR, Greatorex-Davies JN, Asher J, Roy DB (2010) Turnover and trends in butterfly communities on two British tidal islands: stochastic influences and deterministic factors. J Biogeogr 37:2291–2304
Desender K, Dekoninck W, Dufrêne M, Maes D (2010) Changes in the distribution of carabid beetles in Belgium revisited: have we halted the diversity loss? Biol Conserv 143:1549–1557
Dover J, Settele J (2009) The influences of landscape structure on butterfly distribution and movement: a review. J Insect Conserv 13:3–27
Emmet AM (1991) Life history and habits of the British Lepidoptera. In: Emmet AM, Heath J (eds) The moths and butterflies of Great Britain and Ireland. Harley Books, Colchester, pp 61–203
Fahrig L, Merriam G (1994) Conservation of fragmented populations. Conserv Biol 8:50–59
Fibiger M (1993) Noctuidae Europaeae. Noctuidae I. Entomological press, Sorö
Franzén M, Johannesson M (2007) Predicting extinction risk of butterflies and moths (Macrolepidoptera) from distribution patterns and species characteristics. J Insect Conserv 11:367–390
Gärdenfors U (2010) The 2010 red list of Swedish species. Swedish species information centre, SLU, Uppsala
Gaston KJ, Blackburn TM, Greenwood JJD, Gregory RD, Quinn RM, Lawton JH (2000) Abundance-occupancy relationships. J Appl Ecol 37:39–59
Gathmann A, Tscharntke T (2002) Foraging ranges of solitary bees. J Anim Ecol 71:757–764
Groenendijk D, Ellis W (2010) The state of the Dutch larger moth fauna. J Insect Conserv 15:95–101
Hanski I (1999) Metapopulation ecology. Oxford University Press, New York
Hardy PB, Sparks TH, Isaac NJB, Dennis RLH (2007) Specialism for larval and adult consumer resources among British butterflies: implications for conservation. Biol Conserv 138:440–452
Harvey PH, Pagel MD (1991) The comparative method in evolutionary biology. Oxford University Press, Oxford
Harvey PH, Purvis A (1991) Comparative methods for explaining adaptations. Nature 351:619–625
Hill JK, Collingham YC, Thomas CD, Blakeley DS, Fox R, Moss D, Huntley B (2001) Impacts of landscape structure on butterfly range expansion. Ecol Lett 4:313–321
Hill JK, Thomas CD, Fox R, Telfer MG, Willis SG, Asher J, Huntley B (2002) Responses of butterflies to twentieth century climate warming: implications for future ranges. Proc R Soc B 269:2163–2171
Huldén L, Albrecht A, Itämies J, Malinen P, Wettenhovi J (2000) Atlas of finnish macrolepidoptera. Lepidopterologiska sällskapet i Finland, Helsingfors
Inkinen P (1994) Distribution and abundance in British noctuid moths revisited. Ann Zool Fenn 31:235–243
Jonasson JÅ (2002) Småfjärilsfaunan på Gotska Sandön. Rapport 5164. Naturvårdsverket, Stockholm
Karsholt O, Razowski J (1996) The Lepidoptera of Europe—a distributional checklist. Apollo books, Stenstrup
Kotiaho J, Kaitala V, Komonen A, Päivinen J (2005) Predicting the risk of extinction from shared ecological characteristics. Proc Nat Acad Sci 102:1963–1967
Leinonen R, Söderman G, Itämies J, Rytkonen S, Rutanen I (1998) Intercalibration of different light-traps and bulbs used in moth monitoring in northern Europe. Entomol Fenn 9:37–51
León-Cortés JL, Lennon JJ, Thomas CD (2003) Ecological dynamics of extinct species in empty habitat networks. 2. The role of host plant dynamics. Oikos 102:465–477
Lindeborg M (2009) Intressanta fynd av storfjärilar (Macrolepidoptera) i Sverige 2008. Entomol Tidskr 130:11–20
Lindgren S (1968) Lövskogsvegetation på Gotska Sandön. Stockholms universitet, Botaniska institutionen, Stockholm
Lindroth CH, Andersson H, Bodvarsson H, Richter SH (1973) Surtsey, Iceland. The development of a new fauna, 1963–1970. Terrestrial invertebrates. Entomol Scand, Suppl 5:280, Copenhagen
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Mace GM, Kershaw M (1997) Extinction risk and rarity on an ecological timescale. In: Kunin WE, Gaston KJ (eds) The Biology of rarity: the causes and consequences of rare-common differences. Chapman & Hall, London, pp 131–149
Mattila N, Kaitala V, Komonen A, Kotiaho JS, Paivinen J (2006) Ecological determinants of distribution decline and risk of extinction in moths. Conserv Biol 20:1161–1168
Mattila N, Kotiaho JS, Kaitila V, Komonen A (2008) The use of ecological traits in extinction risk assessments: a case study on geometrid moths. Biol Conserv 141:2322–2328
Mattila N, Kotiaho JS, Kaitala V, Komonen A (2009) Interactions between ecological traits and host plant type explain distribution change in Noctuid moths. Conserv Biol 23:703–709
Mattsson L (1976) Utklippan—en utpost i havet. Fåglar i Blekinge 12:216–225
New TR (2004) Moths (Insecta: Lepidoptera) and conservation: background and perspective. J Insect Conserv 8:79–94
New TR, Pyle RM, Thomas JA, Thomas CD, Hammond PC (1995) Butterfly conservation management. Annu Rev Entomol 40:57–83
Nieminen M, Rita H, Uuvana P (1999) Body size and migration rate in moths. Ecography 22:697–707
Nilsson SG, Franzén M, Jönsson E (2008) Long-term land-use changes and extinction of specialised butterflies. Insect Conserv Divers 1:197–207
Öckinger E, Hammarstedt O, Nilsson SG, Smith HG (2006) The relationship between local extinctions of grassland butterflies and increased soil nitrogen levels. Biol Conserv 128:564–573
Öckinger E, Schweiger O, Crist TO, Debinski DM, Krauss J, Kuussaari M, Petersen JD, Pöyry J, Settele J, Summerville KS, Bommarco R (2010) Life-history traits predict species responses to habitat area and isolation: a cross-continental synthesis. Ecol Lett 13:969–979
Piha M, Linden A, Pakkala T, Tiainen J (2007) Linking weather and habitat to population dynamics of a migratory farmland songbird. Ann Zool Fenn 44:20–34
Pöyry J, Luoto M, Heikkinen RK, Kuussaari M, Saarinen K (2009) Species traits explain recent range shifts of Finnish butterflies. Glob Change Biol 15:732–743
Scoble MJ (1988) The lepidoptera: form, function and diversity. Oxford University Press, Oxford
Shreeve TG (1995) Butterfly mobility. In: Pullin AS (ed) Ecology and conservation of butterflies. Chapman and Hall, London
Shreeve TG, Dennis RLH (2011) Landscape scale conservation: resources, behaviour, the matrix and opportunities. J Insect Conserv. doi:10.1007/s10841-010-9336-9
Shreeve TG, Dennis RLH, Roy DB, Moss D (2001) An ecological classification of British butterflies: ecological attributes and biotope occupancy. J Insect Conserv 5:145–161
Skou P (1991) Nordens ugler. Apollo Books, Stenstrup
Söderman G (1994) Moth monitoring scheme—a handbook for field work and data reporting. Environment Data Centre (EDC). National Board of Waters and the Environment, Helsinki
Sparks TH, Roy DB, Dennis RLH (2005) The influence of temperature on migration of lepidoptera into Britain. Glob Change Biol 11:507–514
Sparks TH, Dennis RL, Croxton PJ, Cade M (2007) Increased migration of lepidoptera linked to climate change. Eur J Entomol 104:139–143
Stearns SC (1983) The influence of size and phylogeny on patterns of covariation among life-history traits in the mammals. Oikos 41:173–187
Svensson I (1993) Lepidoptera-calender. Hans Hellberg, Stockholm
Svensson I, Elmquist H, Gustafsson B, Hellberg H, Imby L, Palmqvist G (1994) Catalogus Lepidopterorum Sueciae. Entomologiska Föreningen, Stockholm
Warren MS, Hill JK, Thomas JA, Asher J, Fox R, Huntley B, Roy DB, Telfer MG, Jeffcoate S, Harding P, Jeffcoate G, Willis SG, Greatorex-Davies JN, Moss D, Thomas CD (2001) Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414:65–69
Wenzel M, Schmitt T, Weitzel M, Seitz A (2006) The severe decline of butterflies on western German calcareous grasslands during the last 30 years: a conservation problem. Biol Conserv 128:542–552
Wilson RJ, Roy D (2009) Butterfly population structure and dynamics. In: Settele J, Shreeve TG, Konvicka M, van Dyck H (eds) Ecology of butterflies. Cambridge University Press, Cambridge
Woiwood IP, Stewart AJA (1990) Butterflies and moths—migration in the agricultural environment. In: Bunce RGH, Howard DC (eds) Species dispersal in agricultural habitats. Belhaven, London
Acknowledgments
We thank Anders Forsman, Patrik Dinnétz and Erik Öckinger for valuable statistical advice and constructive comments on the manuscript. Thanks also to Tomas Nilsson, Kjell Arvidsson, Lennart Rovin and Sjöräddningssällskapet Hasslö for organising transport and making it possible to perform the study on Utklippan; Dag Pedersen and the other park wardens on Gotska Sandön. Permission for using light-traps in protected areas was issued by the County Administration Board of Blekinge (Utklippan) and Gotland (Gotska Sandön). Financial support was provided by “Stiftelsen Längmanska Kulturfonden”.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Franzén, M., Betzholtz, PE. Species traits predict island occupancy in noctuid moths. J Insect Conserv 16, 155–163 (2012). https://doi.org/10.1007/s10841-011-9401-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-011-9401-z