
Abstract
A comparison has been made for the first time between the cholinergic components of the nervous system of important human digeneans namely Schistosoma mansoni and Schistosoma haematobium from infected hamster (Cricentus auratus) in Egypt. In each parasite, the central nervous system consists of two cerebral ganglia and three pairs of nerve cords (ventral, lateral, and dorsal) linked together by some transverse connectives and numerous ring commissures. Peripheral cholinergic innervation was detected in oral and ventral suckers and in some parts of female reproductive system in both species, but there were some differences. The possible functions of some of these nervous components are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The nervous system occupies a position of biovital importance in flatworm biology. In addition to carrying sensory and neuromuscular signals, it may be responsible for the systemic transmission of developmental and hormonal cues, because, as acoelomates, these organisms lack the body cavity and circulatory system which would otherwise contribute to such functions (McVeigh et al. 2009). In the last few years, considerable attention has been paid to the nervous system of flatworms using enzyme and immunocytochemical techniques. Some studies have been carried out using the light microscope in combination with the confocal scanning laser microscope.
The cholinesterase activity was used as an indirect evidence of the presence of acetylcholine in the nervous system of flatworms by many authors, including digeneans, cestodes, and monogeneans (Halton and Gustafsson 1996). The indoxyl acetate enzymatic technique was used on whole mounts (Halton and Jennings 1964; Halton and Morris 1969; Cable et al. 1996). Zurawski et al. (2001) used chloro-indolyl acetate. Other authors used the acetylthiocholine iodide enzymatic technique (Rahemo and Gorgess 1987; Buchmann and Mellergaards 1988). According to Rahemo and Gorgess (1987), the acetylthiocholine iodide technique gave best results than the indoxyl acetate method.
Demonstration of the cholinesterase activity has been used as an indirect proof of the presence of acetylcholine in the nervous system of the monogeneans, such as Pseudodactylogyrus bini and Pseudodactylogyrus anguillae (Reda and Arafa 2002), Macrogyrodactylus clarii (El-Naggar et al. 2004), Mcongolensis (El-Naggar et al. 2007), and Gyrodactylus rysavyi (Arafa et al. 2007), and cestodes, such as Trilocularia acanthiaevulgaris (Fairweather et al. 1990) and Moniezia expansa (Maule et al. 1993), and Rahemo and Elkalake (1994) have studied six species of cestodes. Concerning the digeneans, the studies included larval and adult forms: larval forms such as larval stages of Schistosoma mansoni (Bruckner and Vage 1974), cercaria of S. mansoni and Schistosoma japonicum (Orido 1989), cercaria of Diplostomum pseudospathaceum (Niewiadomska & Moczon 1982), daughter and mother sporocyst of D. pseudospathaceum (Niewiadomska & Moczon 1990), and sporocysts of S. mansoni (Diconza & Basch 1975) and adult forms, such as S. mansoni (Fripp 1967), Fasciola hepatica and Fasciola gigantic (Prcbert and Durrani 1976), Gorgoderina vitelliloba (McKAY et al. 1991), G. vitelliloba and Gorgoderina euzeti (Rahemo 1993), and Haplorchoides cahirinus and Acanthostomum absconditum (Arafa et al. 2002). The aim of this study is to describe the nervous system of both S. mansoni and Schistosoma haematobium for the first time in detail, using acetylthiocholine iodide enzymatic technique. Such study may help to find an antihelminthic drug that works specifically on nervous system elements of the parasite in the future.
Material and methods
Adult flukes
Schistosoma species were obtained from infected hamster (Cricentus auratus) from Biological Production Unite (BPU) of Theodore Bilharz Research Institute (TBRI, Giza, Egypt). S. mansoni was perfused from the hepatic portal vessels and mesenteric veins, while S. haematobium was perfused from veins of urinary bladder using perfusion pump containing phosphate-buffered saline (PBS). All parasites were rapidly placed in culture media (RPMI-1640)
Acetylthiocholine iodide technique
To reveal the nervous system of Schistosoma species, about 250 living worms of S. mansoni and 30 of S. haematobium were flattened between microscope slides and coverslips then fixed in 10 % neutral formalin for about 30 min. They were washed in distilled water, incubated in working solution (acetylthiocholine iodide) according to Rahemo and Gorgess (1987), and examined using a steromicroscope at intervals. As soon as the details of the nervous system become clearly visible, the specimens were rapidly washed in distilled water and dehydrated in ascending series of ethanol. Finally, they were cleared in terpinol, mounted in DPX, and then examined with light microscope and oil immersion optical equipment.
Results
Nervous system
Although acetylthiocholine iodide technique (AChI) techniques applied on about 30 specimens of S. haematobium at the same time of S. mansoni, S. haematobium showed a strong positive reaction for cholinesterase enzyme after half an hour. There were differences in the positive reactions observed in the same parts of different specimens of S. mansoni. Concerning S. haematobium, the stain was stable and site-specific, while in S. mansoni, it was slightly faded and needed more time to be stable. Therefore, many specimens of S. mansoni about (250) through 2 years that were examined to set up the exact body organs have a positive reaction toward the stain (Table 1).
Cholinergic components of the nervous system of Schistosoma mansoni
Central nervous system
The central nervous system of S. mansoni is composed of two conspicuous triangular cerebral ganglia (cg) connected together by a single, thick ventral connection (vco) and located posterior to the oral sucker (Figs. 1, 4–14). Two anteroventral connectives (avc), arising from each cerebral ganglion, are extended anteriorly where they join the anterior terminal commissure (tc) lying at the distal extremity of the head region (Figs. 2 and 5a).

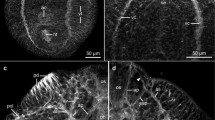
1 Diagram of whole mount showing the nervous system of S. mansoni as revealed by AChI staining. Avlc anteroventrolateral connectives, cg cerebral ganglia, dc1–dc4 dorsal connectives, dnc dorsal nerve cord, gk ganglion knots, lnc lateral nerve cord, m mouth, os oral sucker, pvlc posteroventrolateral connectives, rc ring commissures, vc venteral transverse connective, vn ventral sucker nerve, vnc ventral nerve cord, vs ventral sucker. 2 Diagram of oral sucker of S. mansoni showing the main nerves supplying it. adc anterodorsal connectives, avc anteroventral connectives, on oral sucker nerve, or oral sucker ring, rc1 and rc2 ring commissures, tc terminal commissure, vco ventral commissure, vlc ventrolateral connectives. Other abbreviations as in Fig. 1. 3 Diagram of ventral sucker of S. mansoni showing dorsal nerve cord (dnc), ventral sucker nerve (vn), and ventral nerve cord (vnc). Note fine nerves that anastomose as plexuses

Light micrographs of male S. mansoni treated with AChI. 4 Whole mount of the parasite; two ventral nerve cords (vnc) are separated and ended at the same level. Abbreviations as in Fig. 1. 5 a Ventral view of anterior region showing dorsal commissure (dco). Other abbreviations as in Fig. 2. b Dorsal view of anterior region. Note many radiating parallel nerves (small arrows). Abbreviations as in Fig. 2. 6 Anterior region. Note branches of fine nerves. Abbreviations as in Fig. 3. 7 Magnification of the ventral sucker (vs). Abbreviations as in Fig. 1. 8 Lateral view of middle region showing ganglion knots (gk). 9 Posterior region. Note the end of ventral nerve cord (white arrow head). Abbreviations as in Fig. 1. 10 Dorsal surface showing numerous nerve sensilla (ns). 11 End region showing excretory pore (ep) and excretory pore nerve (epn). 12 Anterior region showing esophagus (oe), oral sucker (os), and ventral (vs) sucker

Light micrographs of female S. mansoni treated with AChI. 13 Whole mount showing positive reaction of vitelline glands (vi). 14 Magnification of anterior region showing the positive reaction of (cg) cerebral ganglia and (vco) ventral commissure. 15 Ventral view of middle region showing dorsal nerve cord (dnc), lateral nerve cord (lnc), ring commissures (rc), and ventral nerve cord (vnc). 16 Dorsal view of middle region showing nerve cells (nc). 17 Whole mount of female. 18 Magnification of posterior region of reproductive system showing positive reaction of egg (e), nerve cells (nc), ootype (oӧ), and ovary (ov)
Two thick and prominent ventral nerve cords (vnc) originate one from each cerebral ganglion and run posteriorly, with the same thickness, ending separately at the same level (Figs. 1, 4–9). In the anterior part of the body and in posterior to the ventral sucker, only one ventral transverse connective (vc) is present and connect between the two ventral nerve cords (Fig. 1).
Two thin lateral nerve cords (lnc) run one on each side of the body in parenchymal tissue and parallel to the body margins (Figs. 1 and 4). Each lateral nerve cord connects anteriorly to the anterior terminal commissure and joins the other in the posterior extremity of the body (Fig. 1). The ventral nerve cords communicate with the lateral nerve cords by means of one pair of ventrolateral connectives (vlc) (Fig. 2), six pairs of ventrolateral connectives (avlc) anteriorly, and four pairs posteriorly (pvlc) (Fig. 1).
Two thinner dorsal nerve cords (dnc) arise one from each cerebral ganglion and extend posteriorly and dorsally in the middle region of the body where they join each other anteriorly to the union of the lateral nerve cords. There are two anterodorsal connectives (adc), arising one from each cerebral ganglion and run in an anterodorsal direction where they join the anterior terminal commissure (Figs. 1 and 5b). Three dorsal connectives (dc1–dc3) connect the dorsal nerve cords in the end of the body.
Two ring commissures (rc1–rc2) are situated in the area between the oral sucker and cerebral ganglia. Each ring commissure connects the dorsal nerve cords with the lateral nerve cords where four small ganglion knots (gk) are formed (Fig. 2). There are numerous ring commissures extending from the cerebral ganglia to the end of the body (Figs. 7 and 8) and (Fig. 15) in female.
Peripheral nervous system
The acetylthiocholine iodide method has revealed the presence of extensive peripheral innervations of the oral, ventral suckers, esophagus, uterus, vitelline glands, ovary, oӧtype, nerve sensilla, and excretory pore. The oral sucker is innervated by two short oral sucker nerves (on) arising one from each anterioventral connective. These nerves surround the oral sucker forming oral sucker ring (or). The oral ring gives rise to many radiating parallel nerves (Figs. 2 and 5a). Also, oral sucker is innervated by two very thin nerves arising one from each anterodorsal connective. These nerves are running just anterior to oral ring and branch to many fine nerves that anastomose as plexuses (Figs. 2 and 5b). The venteral sucker is innervated by two ventral sucker nerve (vn) which is deeply stained, arising one from each ventral nerve cord, and each ventral nerve give rise to three nerve rings that branch to many fine nerves (Figs. 3 and 6). Numerous nerve sensilla are observed and extending through the whole body. It is more numerous in dorsal surface than ventral one and randomly distributed, not in linear arrangement, so making of distribution pattern of surface sensilla is difficult. Each nerve sensillum appears as a minute volcanic opening with a relatively dark center surrounded by a black ring (Fig. 10). Esophagus is moderately nerve stained as well as the excretory pore (Figs. 11 and 12, respectively). The cholinergic innervations of the female (Figs. 13–17) are observed in vitelline glands which show moderate nerve activity (Fig. 13). There is some multipolar nerve cells (nc) that are detected near dorsal surface of the body. They are located in the region posterior to the ventral sucker and adjacent to the dorsal nerve cords, and each nerve cell has a conspicuous nucleus and two or more extensions (Fig. 16). The wall of uterus showed deep activity, and each egg in ootype and ovary showed marked brown granular appearance (Fig. 18). No evidence of any cholinergic innervations of the male reproductive system is observed.
Cholinergic components of the nervous system of Schistosoma haematobium
Central nervous system
The central nervous system of S. haematobium is composed of two conspicuous spherical cg connected together by a single, thick vco and located just posterior to the oral sucker (Figs. 19, 22, and 23). Two avc, arising from each cerebral, are extended anteriorly where they join the anterior tc lying at the distal extremity of the head region (Fig. 19).

19 Diagram of whole mount showing the nervous system of S. haematobium as revealed by AChI staining. adc anterodorsal connectives, avc anteroventral connectives, avlc anteroventrolateral connectives, cg cerebral ganglia, dc1–dc11 dorsal connectives, dlc dorsolateral connectives, dnc dorsal nerve cord, gk ganglion knots, lnc lateral nerve cord, m mouth, on oral sucker nerve, or oral sucker ring, os oral sucker, pvlc posteroventrolateral connectives, rc ring commissures, rc1–rc4 ring commissures of anterior region, tc terminal commissure, vc venteral transverse connective, vco ventral connection, ve vesicle, vlc ventrolateral connectives, vn ventral sucker nerve, vnc ventral nerve cord, vs ventral sucker. 20 Diagram of ventral sucker of S. haematobium showing nerve branches (nb), ventral sucker nerve (vn), and ventral sucker ring (vr). 21 Diagram of the posterior region of S. haematobium showing dorsal connectives (dc7–dc11), excretory pore (ep), excretory pore nerve (epn), inner dorsal nerve cords (idnc), and vesicle nerve (ven). Other abbreviation as in Fig. 19

Light micrographs of male S. haematobium treated with AChI. 22 Whole mount of the parasite; two ventral nerve cords (vnc) are separated and ended at different level. Abbreviations as in Fig. 19. 23 Anterior region showing positive reaction of parenchima (p) and other regions as in draw. Other abbreviations as in Fig. 19. 24 Oral sucker (os). 25 Magnification of oral sucker. Abbreviations as in Fig. 19. 26 and 27 Ventral sucker (vs) region. Abbreviations as in Fig. 19. 28 The region below the ventral sucker. Abbreviations as in Fig. 19. 29 Middle region. Abbreviations as in Fig. 19
Two thick and prominent vnc originate one from each cerebral ganglion and run posteriorly, with the same thickness, ending separately at different levels (Figs. 19, 22, 29–34). In the anterior part of the body and in posterior to the ventral sucker, only one vc is present and connect between the two ventral nerve cords (Figs. 19 and 28).

Light micrographs of male S. haematobium treated with AChI. 30 Top view of middle region. Abbreviations as in Fig. 19. 31 Lateral view showing lateral nerve cord (lnc) and parenchima (p). 32 Dorsal surface of ventral sucker region showing nerve cells (nc). 33 Lateroventral view of posterior region showing excretory pore (ep). 34 Lateroventral view of posterior region. Abbreviations as in Fig. 19. 36 and 37 Light micrographs of female S. mansoni treated with AChI. 35 Anterior half of the body. 36 Magnification of reproductive system showing egg (e), ootype (oӧ), ovary (ov), and uterus (u)
Two thin lnc run one on each side of the body in parenchymal tissue and parallel to the body margins (Figs. 19 and 29–30). Each lateral nerve cord connects anteriorly to the anterior terminal commissure and joins the other in the posterior extremity of the body (Fig. 19). The ventral nerve cords communicate with the lateral nerve cords by means of one pair of vlc, seven pairs of avlc (Figs. 19 and 27), and five pairs pvlc (Fig. 19).
Two slightly thin dnc arise one from each cerebral ganglion and extend posteriodorsally in the middle region of the body where they join each other anteriorly to the union of the lateral nerve cords. The most remarkable interesting feature in this system is the presence of two very thin inner dorsal nerve cords (idnc) running posteriorly from seventh dorsal connective (dc7) to join the dorsal nerve cords near excretory pore (Fig. 21). Also, these cords were detected by silver impregnation technique. There are two adc, arising one from each cerebral ganglion and run in an anterodorsal direction where they join the anterior terminal commissure (Fig. 19). Three dorsal connectives (dc1–dc3) connect the dorsal nerve cords in the anterior half of the body. Also, eight dorsal connectives (dc4–dc11) linking the dorsal nerve cords are observed in the posterior part of the body. The dorsal nerve cords are also connected to the lateral nerve cords via one dorsolateral connective (dlc) (Fig. 19).
Four ring commissures (rc1–rc4) are situated at the area between the oral sucker and cerebral ganglia. Each ring commissure connects the dorsal nerve cords with both lateral nerve cords where eight small gk are formed (Fig. 19). There are numerous ring commissures extending just from posterior to the cerebral ganglia to the level of excretory pore (Figs. 19 and 30).
Peripheral nervous system
The acetylthiocholine iodide method has revealed the presence of extensive peripheral innervations of the oral, ventral suckers, uterus, oӧtype, nerve cells, excretory vesicle, and excretory pore. The oral sucker (Fig. 24) is innervated by two short on arising one from each anteroventral connective (Fig. 25). These nerves surround the oral sucker forming or (Fig. 19). The ventral sucker is innervated by two vn which are deeply stained (Fig. 26), arising one from each ventral nerve cord, and each ventral sucker nerve give rise to three nerve branches (nb) (Fig. 20). These nerve branches on each side give rise to ventral sucker ring (vr) that branch to many fine nerves and anastomose as plexuses (Fig. 20). The wall of uterus showed slight nerve activity, and each egg of oӧtype showed marked brown granular appearance (Figs. 35 and 36). The vitelline glands also show slight nerve activity. No evidence of any cholinergic innervations of the male reproductive system was observed. The excretory vesicle is innervated by four fine vesicle nerves (ven) arising from dorsal connective (dc11) while excretory pore innervating by two excretory pore nerves (epn) arising from dorsal nerve cords (Figs. 21 and 33). Some bipolar and multipolar nc are detected near dorsal surface of the body. They are located at the region posterior to the ventral sucker, adjacent to the dorsal nerve cords, and each cell has a conspicuous nucleus and two or more extensions (Fig. 32).
Discussion
The present study describes for the first time the nervous systems of the digeneans, S. mansoni and S. haematobium. The enzyme cytochemical reaction applied here showed extensive staining for cholinesterase, as indirect evidence for the presence of acetylcholine in the nervous system of the two parasites. This extensive staining of cholinergic components in the nervous system is consistent with all previous studies on flatworms (see Halton and Gustafsson 1996).
The central nervous system of the present digeneans is composed of pair cerebral ganglia connected together by a thick ventral connective and gives rise to three main longitudinal nerve cords (ventral, lateral, and dorsal) which are linked by many ring commissures and some transverse connectives. The number of transverse connectives which link the main nerve cords (dorsal connectives) in S. haematobium is greater than that in S. mansoni.
The gross structure of the central nervous system in S. mansoni and S. haematobium has the same pattern in other flatworms. In the present study, the ventral nerve cords run backward separately (see Halton and Gustafsson 1996 and Reuter and Gustafsson 1995). Also, the same pattern was recorded by Arafa and Reda (2002) in digeneans Orientocreadium batrachoides, Astiotrema reniferum, and Eumasenia aegyptiacus.
In S. mansoni and S. haematobium, ganglionic knots were detected at the crossing points between ring commissures and lateral cords. Ganglionic knots were previously described in parasitic flatworms, for example, Amphilina foliacea by Biserova et al. (2000), H. cahirinus and A. absconditum by Arafa et al. (2002) as well as in free-living flatworms Dugesia tigrina by Reuter and Gustafsson (1995) and Planaria torva by Mantyla et al. (1998).
In the peripheral nervous system of S. haematobium, cholinesterase activity was detected in association with the oral and ventral suckers, uterus, ootype, nerve cells, excretory vesicle as well as excretory pore. Similarly, cholinergic innervation was revealed associated with the oral, ventral suckers, esophagus, uterus, vitelline glands, ovary, oӧtype, nerve sensilla, and excretory pore of S. mansoni. As the esophagus, oral and ventral suckers are highly muscular organs; they are likely to be extensively innervated. Their cholinergic innervation probably plays an important role in controlling the feeding mechanism of both esophagus and oral sucker in addition to the attachment mechanism of the ventral sucker. Also, innervation of the excretory vesicle and excretory opening may control the excretory mechanism.
Cholinergic innervations of the oral and ventral suckers were demonstrated in other digeneans like S. mansoni and F. hepatica (Fripp 1967; Halton 1967, respectively). Moreover, the oral and ventral sucker of the digeneans, F. hepatica, Haplometra cylindracea, S. mansoni, Corriga vitta, and G. vitelliloba, were reported to be provided with a multiplicity of serotoninergic and peptidergic nervous arising from the ventral nerve cords and anastomosing as plexuses of fine nerves among the muscle bands (Magee et al. 1989, 1993; McKay et al. 1990, 1991; Skuce et al. 1990; Marks et al. 1995).
The cholinergic components of the reproductive system were detected in uterus and oӧtype of S. haematobium and uterus, oӧtype, ovary, and vitelline glands of S. mansoni. Cholinergic innervation of the above organs may play a role in copulation and egg formation. Cholinergic innervation of the reproductive system has been previously recorded in few monogeneans as P. bini and P. anguillae (Reda and Arafa 2002) and digeneans H. cahirinus and A. absconditum (Arafa et al. 2002). In some flatworms such as Eudiplozoon nipponicum, cholinesterase activity was totally absent from the reproductive system (Zurawski et al. 2001). However, aminergic and peptidergic innervation were reported in the gonoducts including vas deferens, seminal vesicle, cirrus sac, oviduct, vitelline duct, oӧtype, uterus, and common genital opening of all monogeneans and digeneans thus for examined (Halton and Gustafsson 1996).
The present work has revealed that some cholinergic bipolar and multipolar nerve cells are located very close to the dorsal surface and associated with the dorsal nerve cords. Cholinergic bipolar and multipolar nerve cells were demonstrated in digenean A. absconditum (Arafa et al. 2002). Application of immunocytochemical techniques on S. mansoni and S. haematobium is needed to demonstrate serotoninergic and peptidergic components of the nervous system. Also, further studies of the most powerful effect against Schistosoma species such as Dizygotheca kerchovana and Azadirachta indica extracts as antischistosomal drug are requiring in the future (Abdel Ghaffar et al. 2013).
References
Abdel Ghaffar F, Bakry FA, Taha SA (2013) Impact of plant extracts as molluscicides agent against Biomphalaria alexandrina snails and Schistosoma mansoni. Int J Sci Eng 4:420–426
Arafa SA, El-Naggar MM, El-Abbassy SA, Stewart MT, Halton DW (2007) Neuromusculature of Gyrodactylus rysavyi, a monogenean gill and skin parasite of the Nile catfish Clarias gariepinus. Parasitol Int 56:297–307
Arafa SA, Reda ES (2002) Cholinergic components of the nervous system of the digenean parasites, Orientocreadium batrachoides, Astiotrema reniferum and Eumasenia aegyptiacus from the catfish Clarias gariepinus in Egypt. Egypt J Zool 38D:75–91
Arafa SA, Reda ES, El-Naggar MM (2002) Cholinergic components of the nervous system of the digenean parasites, Haplorchoides cahirinus and Acanthostomum absconditum from the catfish Bagrus bayad in Egypt. Acta Parasitol 47(4):272–279
Biserova NM, Dudicheva VA, Terenina NB, Reuter M, Halton DW, Maule AG, Gustafsson MK (2000) The nervous system of Amphilina foliacea (Platyhelminthes, Amphilinidea). An immunocytochemical, ultrastructural and spectrofluorometrical study. Parasitol 4:441–453
Bruckner DA, Vage M (1974) The nervous system of larval Schistosoma mansoni as revealed by acetylcholinesterase staining. J Parasitol 60(3):437–346
Buchmann K, Mellergaards (1988) Studies on the nervous system of Pseudodactylogyrus anguillae (Monogenea) as revealed by acetylthiocholine activity. Dis Aquat Organ 6:89–92
Cable J, Marks NJ, Halton DW, Shaw C, Johnston CF, Tinsley RC, Gannicott AM (1996) Cholinergic, serotoninergic and peptidergic components of the nervous system of Discocotyle sagittata (Monogenea: Polyopisthocotylea). Int J Parasitol 26:1357–1367
Diconza JJ, Basch PF (1975) Histochemical demonstration of acetylcholin esterase in sporocysts of Schistosoma mansoni (Trematoda). Parasitol 71:305–310
El-Naggar MM, Arafa SZ, El-Abbassy SA, Stewart MT, Halton DW (2004) Neuromusculature of Macrogyrodactylus clarii, a monogenean gill parasite of the Nile catfish Clarias gariepinus in Egypt. Parasitol Res 94(3):163–175
El-Naggar MM, Arafa SA, El-Abbassy SA, Stewart MT, Halton DW (2007) Neuromusculature of Macrogyrodactylus congolensis, a monogenean skin parasite of the Nile catfish Clarias gariepinus. Parasitol Res 100:265–279
Fairweather I, Mahendrasingam S, Johnston CF, Halton DW, McCullough JS, Shaw C (1990) An ontogenetic study of the cholinergic and serotoninergic nervous systems in Trilocularia acanthiaevulgaris (Cestoda, Tetraphyllidea). Parasitol Res 76(6):487–496
Fripp PJ (1967) Histochemical localization of esterase activity in Schistosomes. Exp Parasitol 21:380–390
Halton DW (1967) Studies on phosphatase activity in trematoda. J Parasitol 53(1):46–54
Halton DW, Gustafsson MKS (1996) Functional morphology of the platyhelminth nervous system. Parasitol Res 113:S47–S72
Halton DW, Jennings JB (1964) The demonstration of the nervous system in the monogenetic trematode Diplozoon paradoxum Nordmann by the indoxyl acetate method for esterases. Nature London 202:510–511
Halton DW, Morris GP (1969) Occurrence of cholinesterase and ciliated sensory structures in a fish gill-fluke, Diclidophora merlangi (Trematoda: Monogenea). Parasitol Res 33:21–30
Mantyla K, Halton DW, Reuter M, Gustafsson MK (1998) IV.The nervous system of Tricladida. Neuroanatomy of Planaria torva (Paludicola, Planaridae): an immunocytochemical study. Hydrobiologia 383:167–173
Magee RM, Fairweather I, Johnston CF, Halton DW, Shaw C (1989) Immunocytochemical demonstration of neuropeptides in the nervous system of the liver fluke, Fasciola hepatica (Trematoda, Digenea). Parasitol 98(02):227–238
Marks NJ, Halton DW, Maule AG, Brennan GP, Shaw C, Southgate VR, Johnston CF (1995) Comparative analyses of the neuropeptide F (NPF)-and FMRFamide-related peptide (FaRP)-immunoreactivities in Fasciola hepatica and Schistosoma spp. Parasitol 110(04):371–381
Maule AG, Halton DW, Shaw C, Johnston CF (1993) The cholinergic, serotoninergic and peptidergic components of the nervous system of Moniezia expansa (Cestoda, Cyclophyllidea). Parasitol 106(Pt4):429–440
McKay DM, Halton DW, Johneston CF, Fairweather I, Shaw C (1990) Occurrence and distribution of putative neurotransmitters in the frog-lung parasite Haplometra cylindracea (Trematoda: Digenea). Parasitol Res 76(6):509–517
McKay DM, Halton DW, Johneston CF, Fairweather I, Shaw C (1991) Cytochemical demonstration of cholinergic, serotoninergic and peptidergic nerve elements in Gorgoderina vitelliloba (Trematoda: Digenea). Int J Parasitol 21:71–80
McVeigh P, Mair GR, Atkinson L, Ladurner P, Zamanian M, Novozhilova E, Marks NJ, Day TA, Maule AG (2009) Discovery of multiple neuropeptide families in the phylum Platyhelminthes. Intl J Parasitol 11:1243–1252
Niewiadomska K, Moczon T (1982) The nervous system of Diplostomum pseudospathaceum Niewiadomska, (Digenea, Diplostomatidae). I. Nervous system and chaetotaxy in the cercaria. Parasitol Res 68:304–295
Niewiadomska K, Moczon T (1990) The nervous system of Diplostomum pseudospathaceum Niewiadomska, 1984 (Digenea, Diplostomatidae). IV. Nervous system and distribution of sensilla in the daughter sporocyst. Parasitol Res 76:635–637
Orido Y (1989) Histochemical evidence of the catecholamine-associated nervous system in certain Schistosome cercariae. Parasitol Res 76(2):146–149
Prcbert AJ, Durrani MS (1976) Fasciola hepatica and Fasciola gigantica: total cholinesterase, characteristics and effects of specific inhibitors. Exp Parasitol 42:203–210
Rahemo ZIF, Gorgess NS (1987) Studies on the nervous system of Polystoma integerrimum as revealed by acetylthiocholine activity. Parasitol Res 73:234–239
Rahemo ZIF, Salah al-Din M (1993) The nervoussystem of two frog parasitic trematodes. MU’TAH Lil-Buhooth Wa Al-Dirasat 8:9–20
Rahemo ZIF, Alclak S (1994) Study of the nervous system for six species of cestodes histochemically. MU’TAH Lil-Buhooth Wa Al-Dirasat 21:67–83
Reda ES, Arafa SZ (2002) Cholinergic components of the nervous system of the monogenean gill parasites Pseudodactylogyrus bini and P. anguillae from the Eel Anguilla anguilla in Nile delta waters. Egypt J Zool 38:41–54
Reuter M, Gustafsson MK (1995) The flatworm nervous system: pattern and phylogeny. In: Breidbach O, Kutsch W (eds). The Nervous System of Invertebrates: an Evolutionary and Comparative Approach. Birkhäuser; Basel, 25–59
Skuce PJ, Johnston CF, Fairweather I, Halton DW, Shaw C, Buchanan KD (1990) Immunoreactivity to the pancreatic polypeptide family in the nervous system of the adult human blood fluke, Schistosoma mansoni. Cell Tiss Res 261(3):573–581
Zurawski TH, Mousley A, Mair GR, Brennan GP, Maule AG, Gelnar M, Halton DW (2001) Immunomicroscopical observations on the nervous system of adult Eudiplozoon nipponicum (Monogenea: Diplozoidae). Int J Parasitol 31:783–792
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Rights and permissions
About this article

Cite this article
Reda, E.S., El-Shabasy, E.A., Said, A.E. et al. Cholinergic components of nervous system of Schistosoma mansoni and S. haematobium (Digenea: Schistosomatidae). Parasitol Res 115, 3127–3137 (2016). https://doi.org/10.1007/s00436-016-5070-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-016-5070-x