
Abstract
Phalloidin fluorescence technique, enzyme cytochemistry and immunocytochemistry, in conjunction with confocal scanning laser microscopy, were used to describe the neuromusculature of the monogenean skin parasite Macrogyrodactylus congolensis from the Nile catfish Clarias gariepinus. The body wall muscles are composed of an outer layer of compactly arranged circular fibres, an intermediate layer of paired longitudinal fibres and an inner layer of well-spaced bands of diagonal fibres arranged in two crossed directions. The central nervous system consists of paired cerebral ganglia from which three pairs of longitudinal ventral, lateral and dorsal nerve cords arise. The nerve cords are connected at intervals by many transverse connectives. Both central and peripheral nervous systems are bilaterally symmetrical and better developed ventrally than laterally and dorsally. Structural and functional correlates of the neuromusculature of the pharynx, haptor and reproductive tracts were examined. Results implicate acetylcholine, FMRFamide-related peptides and serotonin in sensory and motor function. The results were compared with those of Macrogyrodactylus clarii, a gill parasite of the same host fish C. gariepinus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many authors have used the activity of cholinesterase (ChE) as indirect histochemical evidence of the presence of acetylcholine (ACh) in the nervous system of monogeneans (Halton and Jennings 1964; Halton and Morris 1969; Cable et al. 1996b; Zurawski et al. 2001) using an indoxyl acetate as substrate for carboxylic esterases. Other studies have employed acetylthiocholine iodide (AcThI) as substrate (Rahemo and Gorgees 1987; Buchmann and Mellergaard 1988; Buchmann and Prentø 1989; Reda and Arafa 2002) to provide a more specific demonstration of ChE itself.
In the past 10 years, many immunocytochemical studies using confocal imaging combined with the use of the phalloidin-fluorescence technique for F-actin were carried out to demonstrate aminergic and peptidergic elements in the nervous systems of oviparous monogeneans, such as Diclidophora merlangi (see Maule et al. 1990a,b), Discocotyle sagittata (see Cable et al. 1996b), Eudiplozoon nipponicum (see Zurawski et al. 2001) and Macrogyrodactylus clarii (see El-Naggar et al. 2004). These studies have considerably added to the understanding of nerve–muscle relationships in flatworm parasites (Halton et al. 1998; Halton 2004).
Like all gyrodactylids, monogeneans of the genus Macrogyrodactylus are viviparous; their uterus contains up to four embryos, one inside the other reminiscent of ‘Russian dolls’ (Cable et al. 1998). Consequently, the reproductive organs of gyrodactylids are modified by the twin pressures of viviparity (Llewellyn 1981; Tinsley 1983) and progenesis (Harris 1983). Macrogyrodactylus congolensis (Prudhoe 1957) Yamaguti 1963 is a skin parasite of the freshwater catfish, Clarias gariepinus (Syn: Clarias lazera) from the river Nile in Egypt (Arafa 1999; El-Naggar et al. 1999; Arafa et al. 2003). M. clarii Gussev 1961 infects the gills of the same host fish (El-Naggar and Serag 1987; El-Naggar 1993). El-Naggar et al. (2001a) studied the comparison between the chaetotaxy of these two parasites, mapping the argentophilic elements of their nervous systems. The neuromusculature of M. clarii was studied in detail using enzyme cytochemistry and immunocytochemistry in conjunction with confocal microscopy (El-Naggar et al. 2004). The present work involved a similar approach to explore the neuromusculature of M. congolensis. It is hoped that such a study will throw some light on the similarities and differences in the muscle and nervous systems of M. congolensis and M. clarii. It may also help researchers to find an anthelminthic drug that may specifically target neural elements of the parasite without substantial side effects on the host.
Materials and methods
Specimens of the Nile catfish C. gariepinus were caught from the Damietta branch of the River Nile near Mansoura City, Dakahlia Province, Egypt. Fish were kept alive in an aquarium containing aerated river water. Individual fish were transferred to a dissecting dish and quickly de-capitated with a sharp knife. The skin covering the head, body and fins was searched for M. congolensis. The parasites were collected with the aid of a fine needle and Pasteur pipette. Identification of M. congolensis was carried out according to Prudhoe (1957).
To reveal cholinergic components of the nervous system, specimens (n=20) of M. congolensis were flattened between a microscope slide and coverslip and fixed in 10% neutral (phosphate-buffered) formalin for 30 min. They were then washed in distilled water, incubated in a solution of AcThI according to Rahemo and Gorgees (1987) and examined using a stereomicroscope. As soon as details of the nervous system became visible, the specimens were washed in distilled water. Some of the treated specimens were counter-stained with aqueous solutions of toluidine blue (1%), alcian blue (1%), light green (0.2%), fast green (0.2%) or methyl green (2%). They were then de-hydrated through ethanol, cleared in terpiniol, mounted in Canada balsam and viewed using a Leitz Laborlux S light microscope under oil immersion. Control specimens (n=5) were treated as above but incubated without the AcThI substrate. Serotonin (5-hydroxytryptamine, 5-HT) and FaRP neuropeptide-immunoreactivities were visualised by the indirect immunofluorescence technique of Coons et al. (1955). Worms (n=20) were flat-fixed between slides in 4% (w/v) paraformaldehyde in 0.1 M phosphate-buffered saline (PBS: 0.145 M NaCl, 0.025 M NaH2PO4.2H2O and 0.075 M Na2HPO4 at pH 7.4) for 1 h then transferred to fresh fixative for further 3 h. Specimens were subsequently placed in 0.1 M antibody diluent [AbD: PBS containing 0.35% (v/v) Triton X-100, 0.1% (w/v) sodium azide and 0.1% (w/v) bovine serum albumin] and immediately air-mailed to Belfast. The specimens treated were then incubated with the following primary anti-sera for 72 h at 4°C: neuropeptide anti-serum raised in a guinea pig to the flatworm FaRP, GYIRFamide (working dilution of 1:400) or serotonin anti-serum raised in a rabbit and used at a working dilution of 1/300. After washing overnight in AbD, specimens were incubated for 48 h in rabbit anti-guinea pig IgG (1:100) for GYIRFamide or swine anti-rabbit IgG immunoserum for 5-HT. Secondary anti-sera were conjugated to either fluorescein iso-thiocyanate (FITC) or tetramethyl rhodamine iso-thiocyanate (TRITC) as fluorophores for visualisation of binding sites. After an overnight wash, FITC- or TRITC-labelled phalloidin was employed as a counter-stain for demonstration of filamentous F-actin of muscle. For this, immunostained specimens were incubated in 200 mg/ml phalloidin-FITC or -TRITC for 24 h, washed overnight in AbD and finally mounted in PBS/glycerol (1:9 v/v) and viewed using a Leica TCS-NT confocal scanning laser microscope (Leica Microsystems, Milton Keynes, UK). The controls (n=6) used were those with (1) omission of primary anti-serum, (2) replacement of primary anti-serum with non-immune serum from the donor species, and (3) pre-adsorption of primary anti-serum with 200 mg of appropriate antigen.
Results
Musculature
The body wall or sub-tegumental musculature of M. congolensis consists of three layers of muscle fibres: outer circular (cm), intermediate longitudinal (lm) and inner diagonal myofibres (dm), which form a well-organised, lattice-like arrangement encompassing the whole body (Figs. 1 and 7a,b). Additional dorsoventral muscle fibres extend between the dorsal and ventral borders of the body wall through the parasite. The circular muscle fibres form a thin layer of close and evenly spaced fibres running perpendicular to the longitudinal body axis, whilst the longitudinal fibres are more widely spaced than the circular fibres and run parallel to the longitudinal body axis. The longitudinal muscle fibres appear to be laterally concentrated. Some of the diagonal fibres are orientated singly but others are arranged in pairs (Fig. 7a,b). The diagonal fibres of the dorsal body wall (Fig. 7a) are more extensive and uniformly orientated than those found on the ventral surface (Fig. 7b). The two crossing sets of ventral diagonal somatic muscle fibres run at angles of approximately 30 and 120°, respectively, with respect to the longitudinal fibres, whilst those of the dorsal surface are aligned at approximately 60 and 150°, respectively. The anterior extremity of M. congolensis has two head lobes, each lobe bears an adhesive area (Figs. 1 and 7b,f). There are two pairs of ventral extrinsic muscle bands (vmb), each comprising a group of fibres lying beneath the sub-tegumental muscle (Figs. 1 and 7b). They cross each other at the median line of the head region before attaching to the adhesive sacs of the head lobes. A single orientated diagonal muscle fibre (dm), which is perceived ventrally, accompanies each of these ventral extrinsic muscle bands (Figs. 1 and 7b).

Diagram showing the musculature of the anterior region of M. congolensis. as Adhesive sac (area), aem anterior extrinsic muscle fibres, aph anterior region of the pharynx, cm circular muscle fibres, dm diagonal muscle fibres, hl head lobe, it intestine, lm longitudinal muscle fibres, lem lateral extrinsic muscle fibres, mdf meridional fibres, mo mouth opening, oes oesophagus, pl pharyngeal lumen, pp pharyngeal papillae, pph posterior region of the pharynx, rm radial muscle fibres, ui unbranched region of the intestine and vmb ventral muscle band
The mouth opens into a spacious buccal cavity accommodating the pharynx (Figs. 1, 7c and 8c). The musculature of the mouth opening is partly derived from the sub-tegumental musculature (Fig. 7c). Two bundles of retractor muscle (rt) were observed attaching the anterior extremity of the mouth to the sub-tegumental muscles. Each retractor muscle band consists of at least four prominent muscle fibres (Fig. 7c). The pharynx is divided into anterior and posterior portions by a superficial constriction (Figs. 1, 6a and 8c,e,f). There are six conical papillae projecting from the lips of the pharynx, each one of them is associated with prominent longitudinal muscle fibres (Figs. 1 and 8c,e,f). The anterior portion of the pharynx comprises an extremely dense arrangement of five bands of meridional fibres, the most posterior of which appears as considerably larger bands of muscle (Figs. 1 and 8f). In contrast, the posterior portion of the pharynx is in the form of a relatively large, globular and mainly glandular body. It possesses well-developed radial fibres that run from the tegumental lining of the pharyngeal lumen to the outer periphery of the pharynx (Figs. 1 and 8b,d,f). Moreover, there are four pairs of lateral extrinsic muscle (lem), two on each side, attached to the wall of the anterior region of the pharynx and run to the lateral margin of the body (Fig. 1). Another pair of anterior extrinsic muscle fibres (aem) appears to be attached to the anterior region of the pharynx and is directed to the anterior body margin (Fig. 1).
The pharyngeal lumen opens into the oesophagus through a sphincter composed of circular muscle fibres (Fig. 8d). Inner longitudinal muscle, wrapped with outer circular fibres, characterises the oesophagus. This pattern of muscle continues in a more loosely arranged fashion in the intestine (Fig. 7d), which divides into two blind caecal limbs that run posteriorly along the margins of the worm, ending a short distance from the haptor.
The single testis contains a diffuse array of muscle fibres and leads to a vas deferens, which dilates to form a highly muscular organ namely, vesicula seminalis (Fig. 7d). A short muscular ejaculatory duct arises from vesicula seminalis and opens into the copulatory organ. A highly muscular duct extends from the male accessory reservoir and joins the ejaculatory duct before opening into the male copulatory organ (Fig. 7d). In contrast, the two male accessory glands were not positively stained with phalloidin staining. The copulatory organ was seen to strongly fluoresce after staining with phalloidin, illustrating its highly muscular nature (Figs. 6a, 7d and 8b). The musculature of the female reproductive tract is less well defined in comparison with the male reproductive tract. The wall of egg cell-forming region (ECFR) is mainly composed of circular muscle fibres whilst the uterine wall contains both outer circular and inner longitudinal fibres (Fig. 7e). Embryos within the uterus displayed a well-developed musculature associated with tegument, pharynx, intestine, reproductive organs and haptor (Fig. 9d).
The haptor of M. congolensis is cup-shaped and delineated from the body proper by a prominent constriction. The present study is concentrating only on the most prominent haptoral muscles of M. congolensis using light microscopy and confocal scanning laser microscopy (Fig. 9a–c). The ventral side of the haptor accommodates two pairs of ventral extrinsic muscle fibres. The median pair (mem) is comparatively thicker than the lateral pair (lem, Fig. 9c). Both pairs of median and lateral ventral extrinsic muscles (mem and lem, respectively) extend from the anterior extremity of the haptor to the level of the testis.
Each lateral flap of the haptor is provided with a bundle of thick, transverse muscle (tm, Fig. 9c). The two hamuli are provided with a variety of muscle fibres. The hamulus roots are connected with each other by means of two transverse muscle bundles (rtm), one of them is thicker than the other. The shaft is provided with four groups of muscle, anterior (asm), median (msm), posterior (psm) and lateral (lsm). The anterior, median and posterior muscle groups are attached to the outer surface of the shaft whereas the lateral group attaches to the inner surface of the shaft (Fig. 9c).
There are numerous bundles of dorsal extrinsic muscles (dem) running in the pre-haptoral region and collected into two main thick groups as they pass from the anterior region into the posterior region of the haptor (Fig. 9a). At this site, i.e. at the level of the posterior flap of the haptor, each group of extrinsic muscle branches into numerous fine bundles of muscle fibres, which run nearly parallel to each other and enter the lobes bearing the marginal hooklets (Fig. 9b,c). Moreover, there two groups of median, transverse and crossing muscles (mtc) connecting between the lateral margins of the dorsal aspect of the haptor (Fig. 9b). The posterior flap of the haptor is also provided with at least five circular muscles (cm, Fig. 9a).
Neuroanatomy and neurochemistry
The central nervous system (CNS) of M. congolensis consists of paired cerebral ganglia (cg) from which anterior and posterior neuronal pathways arise and inter-linked by cross-connectives and commissures. The peripheral nervous system (PNS) includes innervation of the alimentary tract, reproductive organs, attachment organs (anterior adhesive areas and haptor) and sub-tegumental muscles. Both CNS and PNS are bi-laterally symmetrical and better developed ventrally than laterally and dorsally. They display strong staining for ChE activity and extensive immunoreactivity for FaRP. No staining was observed in the absence of ChE substrate or primary anti-serum or after substitution of primary anti-serum with non-immune serum; the addition of antigen in liquid phase before immunostaining abolished any reactivity.
Cholinergic enzyme histochemical staining
The CNS of M. congolensis (Figs. 2 and 6e) possesses a curved thick mass of paired cerebral ganglia (cg) located ventrally just anterior to the pharynx (Figs. 2, 3 and 6c–e). A single anterior ventral commissure (avc) originates from the posterolateral region of the cerebral ganglia and runs in a semi-circular manner in the region of the head just posterior to the head lobes (Figs. 3 and 6c–e). Two considerably thick projections extend from the anterior median region of the cerebral ganglia in a butterfly-like appearance (Figs. 3 and 6c,d). Each projection gives rise to three cerebral nerves (cn) that extend anteriorly where they innervate the anterior adhesive areas (Figs. 3 and 6c).

Diagram showing the nervous system of M. congolensis as revealed by AcThI staining. ah Anterolateral marginal hooklet, ap anterior projection, avc anterior ventral commissure, cg cerebral ganglia, db dorsal bar, dc1–dc3 dorsal connectives, dhn dorsolateral haptoral nerve, dlc1–dlc3 dorsolateral connectives, dnc dorsal nerve cord, dt dorsal testicular nerve, dvc1 and dvc2 dorsoventral connectives, gc ganglionic connectives, h hamulus, ha haptor, hl head lobe, ihn inner haptoral nerve, lnc lateral nerve cord, ohn outer haptoral nerve, ph pharynx, phg1–phg2 pre-haptoral ganglia, phn posterior hook nerves, pmh posterior marginal hooklets, pn pharyngeal nerve, t testis, ut uterus, vb ventral bars, vc1–vc5 ventral connective, vlc1–vlc4 ventrolateral connectives and vnc ventral nerve cord

Diagram showing the anterior region of the body of M. congolensis as revealed by AcThI staining technique. cn Cerebral nerves and pr pharyngeal ring. Other abbreviations as in Fig. 2
Two thick and prominent ventral nerve cords (vnc) originate from the cerebral ganglia and run posteriorly, one on each side of the body where each one joins a pre-haptoral ganglion (phg1, Figs. 2 and 6e). Another pair of pre-haptoral ganglia (phg2) lies one on each side of the body, a short distance posterior to the phg1 (Figs. 2, 5d and 6e). Both phg1 and phg2 on each side of the body are connected via two ganglionic connectives (Figs. 2, 4 and 5d). In the region posterior to the pharynx, two considerably thin branches arise, one from each ventral nerve cord, and re-connect to it at the level of the anterior part of the uterus (Fig. 2). Another thin branch originates from the posterior region of each ventral nerve cord and re-connects the pre-haptoral ganglion phg1 (Fig. 2). Five ventral transverse connectives were detected, three between the two ventral nerve cords and two between the pre-haptoral ganglia phg1 and phg2 (Figs. 2 and 6d,e).

Diagram showing cholinergic innervation of the haptor of M. congolensis. al Antero-lateral lobe and hpa haptoral papillae. Other abbreviations as in Fig. 2

Light micrographs of M. congolensis treated with AcThI. a Middle region of the body showing innervation of the testis (t) with dorsal testicular nerve (dtn). Other abbreviations as in Fig. 2. b Middle region of the body showing innervation of the funnel-shaped birth duct (fs), temporary opening of birth (to), copulatory organ (c), vas deferens (vd), uterus (ut), mouth opening (mo), lateral nerve cord (lnc) and ventral nerve cord (vnc). c Anterior body proper of M. congolensis showing the nervous system of the embryo (e), which resembles that of the adult. ecg Cerebral ganglia of the embryo. d, e Posterior region of the body showing innervation of the haptor. Abbreviations as in Figs. 2 and 4

a, b Confocal scanning laser micrograph of M. congolensis showing immunoreactivity to GYIRFamide (red); muscles stained with FITC-phalloidin (green). c–e Light micrographs of M. congolensis treated with AcThI showing cholinergic components of CNS and PNS. In subpanel e, toluidine blue was used as counter stain. a Anterior region of the worm showing the anterior region of the pharynx (aph), posterior region of the pharynx (pph), copulatory organ (c) and cerebral ganglia (cg). b Inset shows GYIRFamide immunoreactive in the cerebral ganglia of the embryo (ecg) and the pharynx of the embryo (eph). c, d Anterior head region of the body. Abbreviations as in Fig. 3. e Whole worm. ECFR Egg cell-forming region, fs funnel-shaped region, and to temporary birth opening. Other abbreviations as in Fig. 2
Two considerably thin lateral nerve cords (lnc) arise from the posterolateral region of the cerebral ganglia and run posteriorly, one adjacent to each lateral margin of the body where they join the pre-haptoral ganglia phg1 (Figs. 2 and 6e). At regular intervals, the ventral nerve cords communicate with the lateral nerve cords by means of four pairs of ventrolateral connectives (vlc1–vlc4, Figs. 2 and 6e). Two thin dorsal nerve cords (dnc) arise from the posterior median region of the cerebral ganglia and extend posteriorly between the ventral nerve cords where they connect with the anterior pre-haptoral ganglia (phg1). The dorsal nerve cords are cross-linked by three dorsal transverse connectives (dc1–dc3, Figs. 2 and 6e). The dorsal nerve cords are also connected to the lateral nerve cords via three dorsolateral connectives (dlc1–dlc3) whilst they are linked to the ventral nerve cords via two dorsoventral connectives (dvc1 and dvc2, Figs. 2 and 6e).
The haptor of M. congolensis is extensively innervated by two outer (ohn) and four inner (ihn) relatively thick haptoral nerves originating from the pre-haptoral ganglia (phg2) and ventral connective (vc5), respectively. Another two dorsolateral haptoral nerve (dhn), arising one from each pre-haptoral ganglia (phg2), run posteriorly where they eventually branch into seven pairs of posterior hook nerves (phn) and one pair of anterior hook nerve (ahn). The hook nerves innervate the muscles associated with the marginal hooklets (Figs. 4 and 5d,e).
The AcThI method revealed peripheral innervations of the mouth opening (mo, Figs. 5b and 6e), pharynx (Fig. 6c,d), ECFR (Fig. 6e), funnel-shaped birth duct (fs) and temporary birth opening (to, Figs. 5b and 6e). The pharynx is innervated by two pairs of pharyngeal nerves (pn) originating from the ventral connective (vc1). They direct anteriorly and unite to form one pair before connecting with the pharyngeal rings (Figs. 3 and 6d). The testis is innervated by two dorsal testicular nerves (dtn) arising from the dorsal nerve cord (Figs. 2, 5a and 6e). Application of the AcThI method has revealed that the CNS and PNS of the first embryo are well developed and displayed a pattern similar to that obtained for adults (Fig. 5c).
FaRP immunostaining
FaRPergic components were observed throughout the CNS and PNS of M. congolensis. They are distributed in the same manner as the cholinergic components but they are comparatively less extensive. GYIRFamide immunoreactivity was evident in the head region including the cerebral ganglia (cg), anterior thick projections (ap), anterior ventral commissure (avc) and nerves supplying head lobes (hl) and the terminal adhesive areas (as; Figs. 6a, 7f and 8a–c,e). Of the three pairs of posteriorly directed nerve cords, the ventral nerve cords (vnc) were those developed best (Fig. 7f). The pharynx is innervated by two pharyngeal nerves (pn) originating from the first ventral connective (vc1) and direct anteriorly amongst the radial muscle fibres in the posterior region (Fig. 8d,f). FaRPergic cell bodies were observed in both sides of the anterior and posterior regions of the pharynx (Fig. 8a,d). The musculature of each pharyngeal papilla is innervated with FaRPergic nerve fibres (Fig. 8c). Extensive immunoreactivity was observed in the pre-haptoral ganglia (Fig. 9b). Fine nerves extend from these ganglia to innervate the dorsal and ventral extrinsic muscles that enter the haptor and the crossing over of muscles inside the haptor (Fig. 9b,c). Moreover, the immunoreactivity was observed in close association with muscle fibres operating the marginal hooklets (Fig. 9c).

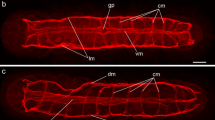
Confocal scanning laser micrographs of whole-mount preparations of M. congolensis stained by indirect immunofluorescence using anti-serum to the FaRP neuropeptide GYIRFamide and TRITC-coupled secondary antibody (red) muscles stained with FITC-phalloidin (green). a Body wall muscle. Note the outer thin layer of circular muscle (cm), an intermediate thicker layer of longitudinal fibres (lm) and a well-developed inner group of diagonal fibres (dm). b Anterior region showing crossing over of the ventral muscle bands (vmb) running into the head lobes. as Adhesive sac, cm circular muscle fibres, dm diagonal muscle fibres and lm longitudinal muscle fibres. c Retractor muscle (rt) attaches to the mouth (mo), which opens into the buccal cavity (bc). Note the longitudinal muscles (lm) and the circular muscles (cm) surrounding the mouth opening. d Branches from the ventral nerve cord (vnc) innervate the male copulatory organ (c), male accessory reservoir (mr), vesicula seminalis (vs) and ejaculatory duct (ej). it Intestine. e Weak immunostaining (unlabelled arrows) in association with the proximal region of the uterus (ut), egg cell-forming region (ECFR), vas deferens (vd) and testis (t). it, Intestine. f The anterior region of the body showing the anterior ventral commissure (avc) and anterior projection (ap), which extend anteriorly from the cerebral ganglia (cg) into the head lobes (hl) to innervate the adhesive sacs (as). cb Cell bodies, ph pharynx and vnc ventral nerve cord

a–f Confocal scanning laser micrographs of the anterior region of M. congolensis showing immunoreactivity to GYIRFamide (green fluorescence of FITC in subpanel a and red fluorescence of TRITC in subpanels b–d). a Forebody showing immunostaining in the cerebral ganglia (cg). cb Cell bodies and ph pharynx. b Forebody showing anterior region of the pharynx (aph), posterior region of the pharynx (pph), copulatory organ (c), cell bodies (cb) and cerebral ganglia (cg). c High powered confocal micrograph of the anterior region. Note the innervation of the pharyngeal papillae (pp) with FaRPergic nerve fibre (np). cb Cell body and mo mouth opening. d High-powered part of the anterior region of the pharynx (aph) and the posterior region (pph) innervated with FaRPergic pharyngeal nerve (pn), which extend from the ventral commissure (vc1). cb Cell body, rm radial muscles and oes oesophagus. e Forebody showing immunostaining for FaRP (red fluorescence of TRITC). Note the protrusible pharyngeal papillae (pp) (green fluorescence of FITC). cg Cerebral ganglia. f Immunoreactivity in the pharyngeal nerve (pn) in close association with the pharynx. aph Anterior region of the pharynx, pp pharyngeal papillae and rm radial muscles. Other abbreviations as in Fig. 7a

Confocal scanning laser micrographs of whole-mount preparations of M. congolensis stained by indirect immunofluorescence using anti-serum to the FaRP neuropeptide NPF and TRITC-coupled secondary antibody (red) muscles stained with FITC-phalloidin (green). a Posterior region showing dorsal extrinsic muscle fibres (dem) running into the haptor (ha) and the circular muscle (cm) in the posterior flap. b Posterior region showing immunoreactivity to NPF (unlabelled arrows) anterior to the haptor and in association with the marginal hooklets. Note the muscle fibres entering the haptor and the crossing over of median transverse muscle fibres (mtc) in the haptor. c A deeper plane of focus of the posterior region. Note that NPF immunoreactivity (unlabelled arrow) closely associated with the muscle fibres entering the haptor (ha). Note that the shaft of the hamulus (h) is provided with anterior (asm), median (msm), posterior (psm) and lateral (lsm) muscle fibres. lem lateral ventral extrinsic muscles, mem median ventral extrinsic muscles, mh marginal hooklet and rtm root transverse muscles. d Middle region of the worm showing the developing embryo in the uterus. Immunoreactivity to NPF can be seen in the cerebral ganglia of the embryo (ecg) and pharynx (eph) of the embryo
FaRPergic nerve tracts (nt) branch from the two ventral nerve cords and direct transversely to innervate the male accessory reservoir and vesicula seminalis (Fig. 7d). FaRPergic immunostaining of the uterine muscles was more developed than that of the ECFR, vas deferens and testis (Fig. 7e).
A strong immunoreactivity for GYIRFamide was revealed in the cerebral ganglia of the first embryo (Figs. 6b and 9d). No FaRP immunostaining was observed in the second embryo.
5-HT immunostaining
In general, staining for 5-HT was considerably less extensive in the brain and CNS compared with to FaRP, although it followed a similar distribution pattern and its description will not be repeated. Moreover, immunostaining for 5-HT revealed the presence of many cell bodies in the cerebral ganglia from which tracts of nerves proceeded in pairs, both anteriorly and posteriorly. Those running anteriorly sub-divided and anastomosed throughout the anterior lobes where staining was expressed as a carpet of immunoreactivity along the entire anteroventral margin of the worm. Circular patterns of staining were also observed in the anterior lobes, corresponding to the region at the base of each sensory organ. A grid-like pattern of serotoninergic fibres, derived from projections originating in the cerebral ganglia, innervate the anterior portion of the pharynx. Peripheral 5-HT-immunoreactive (IR) fibres surround the male copulatory organ and seminal vesicle and immunostaining was also evident around the testis and close to the posterolateral regions of the ECFR. Posteriorly, a peripheral nerve ring of 5-HT-IR fibres runs around the lateral margins of the haptor forming a neural circuit from which fine fibres extend to innervate the terminal region of the haptor and musculature of the marginal hooklets. Staining was also observed in the tegumental papillae that are distributed on the three margins of the ventral surface of the haptor. First-born, and to a lesser extent, second-born daughter embryos within the uterus exhibited immunostaining in both CNS and PNS, mirroring that displayed by the adult.
Discussion
The use of phalloidin staining in conjunction with confocal microscopy revealed the body wall of M. congolensis to be largely composed of an outer layer of compactly arranged circular fibres, an intermediate layer of paired longitudinal fibres and an inner layer of well-spaced bands of diagonal fibres arranged in two crossed directions. This arrangement resembles that described in M. clarii by El-Naggar et al. (2004) for free-living (Rieger et al. 1994) and parasitic flatworms (Halton et al. 1998; Mair et al. 1998; Stewart et al. 2002). El-Naggar et al. (2001b) studied the patterns of movement for three gyrodactylid monogeneans M. clarii, M. congolensis and Gyrodactylus rysavyi. The authors recorded elongation and shortening movement, rotation movement, self-cleaning movement, pendulum-like movement, ordinary upside down leech-like locomotion and swimming movement. They reported that each of the examined gyrodactylid species was found to have a characteristic movement. M. clarii performs scanning (searching) movement, M. congolensis exercises pendulum-like movement and the swimming behaviour was restricted to G. rysavyi. It seems likely that elongation and shortening are brought about by effective contraction of the circular and longitudinal muscles, respectively. Also, it seems most likely that rotation and self-cleaning movement, during which the body is twisted, could be attributed to contraction and relaxation of the diagonal muscle fibres, which are located mainly in the anterior region of the body. The pendulum-like movement in M. congolensis might be achieved by contraction of longitudinal muscle fibres in one side and relaxation of the corresponding muscles in the other side.
Two pairs of ventral muscle bands were revealed beneath the sub-tegumental muscle in the region anterior to the pharynx. Another pronounced, anterior extrinsic muscle bands extend from the pharynx to the anterior body margin. Characteristic features of the ventral muscle bands of M. congolensis, which differentiate them from their corresponding M. clarii (see El-Naggar et al. 2004), are that these muscle bands cross each other at the median line of the head region and that a single orientated diagonal muscle fibre accompanies each of these muscle bands. Moreover, the two anterior extrinsic muscle bands extending from the anterior region of the pharynx attach the anterior body margin in M. congolensis, but they are directed to the adhesive areas in M. clarii. In the leech-like locomotion, the adhesive sacs evert during attachment and retract during detachment. El-Naggar et al. (2004) suggested that the retraction of the adhesive sacs into the head lobes of M. clarii is brought about by contraction of the ventral muscle bands extending from the anterior region of the pharynx. In M. congolensis, retraction of the adhesive sacs into the head lobes seems to be carried out by contraction of the two crossed ventral extrinsic muscle bands. A similar contribution of the ventral muscle bands to the retraction of adhesive sacs was suggested by El-Naggar and Kearn (1980) in the monogenean gill parasites, Dactylogyrus amphibothrium and Dactylogyrus hemiamphibothrium.
Application of the phalloidin staining technique in the present study has revealed that the pharynx of M. congolensis consists of an anterior highly muscular region and posterior mainly glandular region, which accommodates radial muscle fibres. Five pairs of extrinsic muscles were observed to be attached to the anterior region of the pharynx. In M. clarii, six pairs of these muscles were observed (El-Naggar et al. 2004). The anterior portion of the pharynx of the gyrodactylid monogeneans was reported to protrude during feeding (El-Naggar and Serag 1987; Arafa 1999; El-Naggar and El-Abbassy 2003) as well as other monogeneans, namely, Entobdella soleae and Acanthocotyle sp. (Kearn 1963), Macrogyrodactylus polypteri (see Khalil 1970), D. hemiamphibothrium, D. amphibothrium (see El-Naggar and Kearn 1980) and Quadriacanthus aegypticus (see El-Naggar and Serag 1986). Protrusion of the anterior region of the pharynx of M. congolensis may be controlled by the extrinsic muscles attached to this region. The anterior pharyngeal region accommodates six pharyngeal papillae in both M. congolensis and M. clarii and eight pharyngeal papillae in G. rysavyi. The tips of the pharyngeal papillae protrude during feeding in gyrodactylids (Khalil 1970; El-Naggar and Serag 1987; El-Naggar et al. 1999; El-Naggar and El-Abbassy 2003). This protrusion may be initiated by contraction of the muscle bands in the anterior part of the pharynx, whilst contraction of the radial muscle of the posterior portion may drive secretion into the pharyngeal papillae.
The present study has revealed that the copulatory organ of M. congolensis fluoresces strongly with phalloidin staining, illustrating its highly muscular nature. A similar observation was recorded in M. clarii by El-Naggar et al. (2004). The muscle fibres in the copulatory organ could have an important role in the operation of the copulatory bulb and associated large and small spines. El-Naggar et al. (1999) suggested that a plug of co-copulant tissue may be drawn into the copulatory bulb of M. congolensis through suction generated by contraction of the radial muscle in this organ. This may then be secured by the combined action of contraction of equatorial muscle around the rim of the bulb and insertion of the small copulatory spines into the neck of the plug.
The haptor of M. Congolensis was revealed by using confocal scanning laser microscopy to be supplied with dense bands of well-developed ventral and dorsal extrinsic muscles. Although it was difficult to trace the way of the ventral extrinsic muscles in the haptor, it is possible that they attach to the roots of the hamuli. Another two groups of muscles, namely, transverse muscle bands (rtm) are found connecting between the roots of the hamuli. It seems possible that contraction of these transverse muscle bands helps in the protrusion of the hooked regions of the hamuli from their openings. This is followed by contraction of the ventral extrinsic muscles (lem and mem), which pull the hamuli roots in an anterior direction leading to a firm insertion of the hooked regions of the hamuli into the tissue of the host. Moreover, the hamuli of M. congolensis are provided with other groups of muscles, namely, anterior (asm), median (msm), posterior (psm) and lateral (lsm) muscles. It seems probable that these muscles serve in preventing the lateral rotation of the hamuli during attachment, thereby creating a firm insertion of the hamuli. Withdrawal of the pointed hooked region of the hamuli may be achieved by relaxation of both ventral extrinsic muscles and transverse bundles. For a complete understanding of the operating mechanism of the hamuli and marginal hooklets, more investigations are needed using phase-contrast microscopy, different stains for the haptoral muscles and more observations using phalloidin stain.
Regarding the dorsal aspect of the haptor of M. congolensis, there are numerous bundles of dorsal extrinsic muscles (dem) running in the pre-haptoral region and collected into two main thick groups as they pass from the anterior region into the posterior region of the haptor. The precise function of these muscles is not clearly known. However, because these muscles are branched into numerous fibres in the posterior haptoral flap bearing the marginal hooklets, it seems possible that these muscles play a role in operating the posterior flap.
The present study has revealed that CNS of M. congolensis consists mainly of a mass of cerebral ganglia and three pairs of ventral, lateral and dorsal longitudinal nerve cords connected by transverse connectives and commissures; it is better developed ventrally than dorsally and laterally. The CNS of M. congolensis resembles in basic structure that of all previously studied flatworms, including monogeneans, digeneans and cestodes (Reuter 1987; Halton and Gustafsson 1996; Reuter et al. 1998; Arafa and Reda 2002; Reda and Arafa 2002; Arafa et al. 2002; El-Naggar et al. 2004). Typically, the PNS innervates the alimentary system, reproductive organs, attachment organs and sub-tegumental muscles. These neural pathways and their organization were previously described in 11 other monogeneans: Diplozoon paradoxum (see Halton and Jennings 1964), Polystoma integerrimum (see Rahemo and Gorgees 1987), Gyrodactylus salaris (see Reuter 1987), Pseudodactylogyrus anguillae (see Buchmann and Mellergaard 1988; Reda and Arafa 2002), E. nipponicum (see Lyukshina and Shishov 1988; Zurawski et al. 2001), D. merlangi (see Maule et al. 1990a,b), Pseudodactylogyrus bini, P. anguillae (see Reda and Arafa 2002), E. soleae (see Marks et al. 1994), D. sagittata (see Cable et al. 1996a), Protopolystoma xenopodis (see McKay et al. 1991) and M. clarii (see El-Naggar et al. 2004).
Application of silver nitrate staining technique by El-Naggar et al. (2001a) has revealed the presence of eight pairs of large, probably nerve cells, in M. clarii and ten pairs in and M. congolensis. These cells were found distributed bi-laterally on the dorsal and ventral side of the body proper of the warms. The AcThI method (El-Naggar et al. 2004) has also revealed the presence of six pairs of large cholinergic neurons distributed bi-laterally on the dorsal and ventral side of the body proper of M. clarii. However, application of the AcThI method on many specimens of M. congolensis in the present study did not reveal these cells. The nature of these neuronal cell bodies may be understood using transmission electron microscopy. At TEM level, Shaw (1982) described two types of neuronal cell body within the brain of Gastrocotyle trachuri; Rohde (1972) revealed large, ovoid and membrane-bound secretory granules in the large nerves and axons of Polystomoides asiaticus and concluded these to be neurosecretory grana; and Bahatanger et al. (1980) reported two types of nerve cells in Ceylonocotyle scoliocoelium, distinguishing them on the basis of their fine structure as neurosecretory (A-type) and non-neurosecretory (B-type). FaRPergic and serotoninergic staining of M. congolensis revealed numerous pairs of neuronal cell bodies associated with the cerebral ganglia, pharynx and pre-haptoral nerve. In a related monogenean, G. salaris, FaRPergic-IR cell bodies were also detected in the brain, nerve cords, pre-haptoral plexuses and in the middle region of the body (Reuter 1987). In E. soleae, serotoninergic-IR cell bodies and associated innervation dominate the cerebral ganglia, pharynx and ootype and ovovitelline duct (Marks et al. 1994).
The cholinergic and peptidergic components of the nervous system of the studied M. congolensis are partially overlapped. The same phenomenon was recorded in the nervous system of M. clarii by El-Naggar et al. (2004), of D. merlangi by Maule et al. (1990a) and of D. sagittata by Cable et al. (1996b). Halton and Gustafsson (1996) reported that the peptidergic pathways in platyhelminths more follow closely those of the cholinergic system, whilst those of the serotoninergic system are often quite separate and distinctive in construction with the staining localised to a different sub-set of neurons. The present study has revealed that cholinergic and peptidergic components of the nervous system of M. congolensis are more extensive than the aminergic components. ACh is considered as an inhibitory neurotransmitter in a number of parasitic flatworms (Sukhdeo et al. 1984; Thompson and Mettrick 1989; Ward et al. 1986), whereas the function of FaRP neuropeptides in flatworms is yet unclear, although their ubiquitous occurrence throughout the flatworm nervous system implies a fundamental role in nerve muscle physiology (Halton and Gustafsson 1996). When experimentally examined, FaRPs were found to be potently myoactive (Day and Maule 1999).
The nerves supplying the head lobes of M. congolensis may play a role in temporary attachment of the adhesive sacs during locomotion and feeding and/or coordinating muscular movements involved in the release of secretory bodies from the gland cells of the anterior adhesive apparatus. Extensive innervation of the head lobes, particularly the adhesive sacs with FaRPergic-IR nerves, resembles that of M. clarii and is consistent with the presence in these areas of numerous sensilla (El-Naggar et al. 2001a, 2004; Arafa et al. 2003), which may be sensitive to contact stimulation. Contact perception (and possibly contact chemoreception) by this region is likely to be important during temporary leech-like locomotion. Cholinergic and FaRPergic fibres in the posterior region of the pharynx may interact to modulate contraction of the radial muscles, which drive the secretion into the pharyngeal papillae, whilst those fibres supplying the anterior portion of the pharynx may regulate the protrusion of the pharyngeal papillae themselves during feeding. A ring commissure was detected around the mouth of M. congolensis in the present study. A similar commissure was not observed by El-Naggar et al. (2004) in M. clarii. However, in all monogeneans studied hitherto, an oral ring commissure was found (for example, see Rohde 1968; Reuter 1987). Rohde (1968) suggested that the oral commissure is a character, which distinguishes the nervous system of the Monogenea from that of Digenea.
In M. congolensis, FaRPergic staining was evident in nerve fibres that branch from the ventral nerve cord to innervate muscle around the ejaculatory duct, the vesicula seminalis and accessory reservoir, whilst 5-HT-IR traced the muscular ducts of this complex. These findings imply that whilst FaRPs may modulate release of sperm/male accessory secretions, 5-HT may function in their motility through control of the circular muscle fibres of these ducts. The copulatory bulb itself was surrounded by fibres immunoreactive for both 5-HT and FaRPs, suggesting that these neuroactive substances may be involved in bringing about the muscle action responsible for insertion of the small and large spines into the co-copulant.
In the monogenean parasite M. clarii, El-Naggar et al. (2004) suggested that cholinergic and FaRPergic nerves supplying the anterior region of the uterus help coordinate events involved in the delivery of fully formed embryo, whilst the FaRPergic nerves innervating the junction between the ECFR and uterus may play a role in controlling movement of the mature oocyte by their directive effect on the muscles found in this region. Cholinergic and FaRPergic innervation of the reproductive organs of M. congolensis may perform a similar role. However, Cable and Harris (2002) suggested that the large oocyte proceeds into the uterus through the physical pressure of the oocyte on the adjacent empty uterus. Cable et al. (1996a) suggested that the neuropeptide secretions may influence movement of ova and vitelline cells, release of spermatozoa and Mehlis’ gland secretions and peristaltic contractions of the ootype, which shape the egg, and regulate the release of egg from the ECFR into the uterus.
The neuromusculature of the haptor of M. congolensis is richly innervated with cholinergic, FaRPergic and serotoninergic elements, suggesting a major role in motor function for the haptoral nerves. Similarly, in D. merlangi, all three neuroactive substances were detected in motor elements innervating the buccal suckers, pharynx and haptoral clamps (Halton and Morris 1969; Maule et al. 1990a), and their action was investigated on the motility of worms in vitro (Maule et al. 1989). Some evidence for a sensory role for those nerves connected to the tegumental papillae was recorded by Rahemo and Gorgees (1987) on P. integerrimum, but this is yet to be experimentally confirmed.
El-Naggar et al. (2001a) recorded many differences between the surface sensilla pattern (number and distribution) of M. clarii and M. congolensis. The authors suggested that these differences in number and distribution of surface sensilla could be used in addition to anatomical and morphological differences (El-Naggar and Serag 1987; El-Naggar 1993; El-Naggar et al. 1999; Arafa et al. 2003) to discriminate between M. clarii and M. congolensis. In the present study, some differences were recorded between the muscular systems of M. clarii and M. congolensis, mainly in the anterior ventral extrinsic muscle band, the ventral muscle bands extending anteriorly form the anterior region of the pharynx and the musculature of the haptor. These differences could be attributed to the differences in the mode of feeding and attachment due to the difference in the micro-habitat (gills for M. clarii and skin for M. congolensis). These differences could also be used to discriminate between these two parasites. However, the minor differences recorded between the organization of the nervous system of M. congolensis and M. clarii are not considered to be sufficiently reliable for use in discriminating between these two species.
References
Arafa SZ (1999) Biological studies on some monogenean parasites of the teleost Clarias gariepinus in Dakahlia province. Ph.D. thesis, Faculty of Sciences, Mansoura University, Egypt
Arafa SZ, Reda ES (2002) Cholinergic components of the nervous system of the digeneans Astiotrema reniferum, Orientocreadium batrachoides and Eumasenia aegypticus from the catfish Clarias gariepinus in Egypt. J Egypt Ger Soc Zool 38:75–91
Arafa SZ, Reda ES, El-Naggar MM (2002) Cholinergic components of the nervous system of the digenean parasites, Haplorchoides cahirinus and Acanthostomum absconditum from catfish Bagrus bayad in Egypt. Acta Parasitol 47:272–279
Arafa SZ, El-Naggar MM, Kearn GC (2003) Scanning electron microscope observations on the monogenean skin parasite Macrogyrodactylus congolensis (Prudhoe, 1957) Yamaguti, 1963. Acta Parasitol 48:272–279
Bahatanger A, Gupta A, Srivastava R (1980) Histological and histochemistry of the neuroendocrine components of Ceylonocotyle scoliocoelium (Digenea: Trematoda). Z Parasitenkd 64:77–84
Buchmann K, Mellergaard S (1988) Histochemical demonstration of the inhibitory effect of Nuvan and Neguvon on cholinesterase activity in Pseudodactylogyrus anguillae (Monogenea). Acta Vet Scand 29:51–55
Buchmann K, Prentø P (1989) Cholinergic and aminergic elements in the nervous system of Pseudodactylogyrus bini (Monogenea). Dis Aquat Org 6:89–92
Cable J, Harris PD (2002) Gyrodactylid developmental biology: historical review, current status and future trends. Int J Parasitol 32:255–280
Cable J, Harris PD, Tinsley RC (1996a) Ultrastructural adaptations for viviparity in the female reproductive system of gyrodactylid monogeneans. Tissue Cell 28:515–526
Cable J, Marks NJ, Halton DW, Shaw C, Johnston CF, Tinsley RC, Gannicott AM (1996b) Cholinergic, serotoninergic and peptidergic components of the nervous system of Discocotyle sagittata (Monogenea: Polyopisthocotylea). Int J Parasitol 26:1357–1367
Cable J, Harris PD, Tinsley RC (1998) Life history specializations of monogenean flatworms: a review of experimental and microscopical studies. Microsc Res Tech 42:186–199
Coons AH, Leduc EH, Connelly JM (1955) Studies on antibody production. I. A method for histochemical demonstration of specific antibody and its application to a study of the hyper-immune rabbit. J Exp Med 102:42–60
Day TA, Maule AG (1999) Parasitic peptides! The structure and function of neuropeptides in parasitic worms. Peptides 20:999–1019
El-Naggar MM (1993) Scanning electron microscope observations on the head lobes and haptor of the monogenean Macrogyrodactylus clarii Gussev, 1961. J Egypt Ger Soc Zool 10:143–155
El-Naggar MM, El-Abbassy SA (2003) Anatomical observations on the viviparous monogenean Gyrodactylus rysavyi Ergens, 1973 from the Nile catfish Clarias gariepinus in Egypt. Egypt J Zool 40:225–249
El-Naggar MM, Kearn GC (1980) Ultrastructural observations on the anterior adhesive apparatus in the monogeneans Dactylogyrus amphibothrium Wagener, 1857 and D. hemiamphibothrium Ergens 1956. Z Parasitenkd 61:223–241
El-Naggar MM, Serag HM (1986) Quadriacanthus aegypticus n sp., a monogenean gill parasite from the Egypt teleost Clarias lazera. Syst Parasitol 8:129–140
El-Naggar MM, Serag HM (1987) Redescription of Macrogyrodactylus clarii Gussev 1961, a monogenean gill parasite of Clarias lazera in Egypt. Arab Gulf J Sci Res Agric Biol Sci 5:257–271
El-Naggar MM, Kearn, GC, Hagras AE, Arafa SZ (1999) On some anatomical features of Macrogyrodactylus congolensis, a viviparous monogenean ectoparasite of the catfish Clarias gariepinus from Nile water. J Egypt Ger Soc Zool 29:1–24
El-Naggar MM, Arafa SZ, El-Abbassy SA, Kearn GC (2001a) Chaetotaxy of the monogeneans Macrogyrodactylus clarii and M. congolensis from the gills and skin of the catfish Clarias gariepinus in Egypt, with a note on argentophilic elements in the nervous system. Folia Parasitol 48:201–208
El-Naggar, MM, EL-Naggar AM, El-Abbassy SA (2001b) Microhabitat and movement of the viviparous monogeneans Gyrodactylus alberti, Macrogyrodactylus clarii and M. congolensis from the Nile catfish Clarias gariepinus. J Egypt Ger Soc Zool 35:169–187
El-Naggar MM, Arafa SZ, Stewart MT, El-Abbassy SA, Halton DW (2004) Neuromusculature of Macrogyrodactylus clarii, a monogenean gill parasite of the Nile catfish Clarias gariepinus in Egypt. Parasitol Res 94(3):163–175
Gussev AV (1961) A viviparous monogenetic trematode from freshwater basins of Africa. Doklady Akademii Nauk Armãnskoj SSR 136:490–493
Halton DW (2004) Microscopy and the helminth parasite. Micron 35:361–390
Halton DW, Gustafsson MKS (1996) Functional morphology of the platyhelminth nervous system. Parasitology 113:S57–S72
Halton DW, Jennings JB (1964) Demonstration of the nervous system of the monogenetic trematode Diplozoon paradoxum Nordmann by the indoxyl acetate method for esterases. Nature, London 202:510–511
Halton DW, Morris GP (1969) Occurrence of cholinesterase and ciliated sensory structures in a fish gill fluke, Diclidophora merlangi (Trematoda: Monogenea). Z Parasitenkd 33:21–30
Halton DW, Maule AG, Mair GR, Shaw C (1998) Monogenean neuromusculature: some structural and functional correlates. Int J Parasitol 28:1609–1623
Harris PD (1983) The morphology and life-cycle of the oviparous Gyrodactylus farlowellae gen. et sp. nov. (Monogenea Gyrodactyidae). Parasitology 87:405–420
Kearn GC (1963) Feeding in some monogenean skin parasites: Entobdella solea on Solae solae and Acanthocotyle sp. on Raia clavata. J Biol Assoc UK 43:749–766
Khalil LF (1970) Further studies on Macrogyrodactylus polypteri, a monogenean on the African freshwater fish Polypterus senegalus. J Helminthol 44:329–348
Llewellyn J (1981) Evolution of viviparity and invasion by adults. Parasitology 82:64–66
Lyukshina LM, Shishov BH (1988) Biogenic amines in the nervous system of Eudiplozoon nipponicum (Monogenea). In: Sakharov DA (ed) Simple nervous systems (in Russian). Nauka, Moscow, pp 173–176
Mair GR, Maule AG, Shaw C, Johnston CF, Halton DW (1998) Gross anatomy of the muscle systems of Fasciola hepatica as visualized by phalloidin-fluorescence and confocal microscopy. Parasitology 117:75–82
Marks NJ, Halton DW, Kearn GC, Shaw C, Johnston CF (1994) 5-Hydroxytryptamine-immunoreactivity in the monogenean parasite Entobdella soleae. Int J Parasitol 24:1011–1018
Maule AG, Halton DW, Allen JM, Fairweather I (1989) Studies on motility in vitro of the ectoparasitic monogenean, Diclidophora merlangi. Parasitology 98:85–93
Maule AG, Halton DW, Johnston CF, Shaw C, Fairweather I (1990a) The serotoninergic, cholinergic and peptidergic components of the nervous system in the monogenean parasite Diclidophora merlangi. Parasitology 100:255–274
Maule AG, Halton DW, Johnston CF, Shaw C, Fairweather I (1990b) A cytochemical study of the serotoninergic, cholinergic and peptidergic components of the reproductive system of the monogenean parasite Diclidophora merlangi. Parasitol Res 76:409–419
McKay DM, Halton DW, Maule AG, Johnston CF, Shaw C, Fairweather I (1991) Putative neurotransmitters in two monogeneans. Helminthologia 28:75–81
Prudhoe S (1957) Trematoda. In Exploration du Prac National de l’Upemba. I. Mission G. F. et R. Witte en collaboration avec W. Adam, A. Janssens, L. Van Meet et R. Verheyen (1946–1949), vol. 48, pp 1–28
Rahemo ZIF, Gorgees NS (1987) Studies on the nervous system of Polystoma integerrimum as revealed by acetylthiocholine activity. Parasitol Res 73:234–239
Reda ES, Arafa SZ (2002) Cholinergic components of the nervous system of the monogenean gill parasites, Pseudodactylogyrus bini and P. anguillae from the eel Anguilla anguilla in Nile Delta waters. Egypt J Zool 38:41–54
Reuter M (1987) Immunocytochemical demonstration of serotonin and neuropeptides in the nervous system of Gyrodactylus salaris (Monogenea). Acta Zool 68:187–193
Reuter M, Mäntylä K, Gustafsson MKS (1998) Organization of the orthogon—main and minor cords. Hydrobiologia 383:175–182
Rieger RM, Salvenmoser W, Legneti A, Tyler S (1994) Phalloidin-rhodamine preparations of Macrostomum hystricinum marinum (Platyhelminthes): morphology and postembryonic development of the musculature. Zoomorphology 114:133–147
Rohde K (1968) Das Nervensystem der Gattung Polystomoides Ward, 1977 (Monogenea). Z Morphol Tiere 62:58–76
Rohde K (1972) Fine structure of the Monogenea, especially Polystomoides Ward. Adv Parasitol 10:1–33
Shaw MK (1982) The fine structure of the brain of Gastrocotyle trachuri (Monogenea: Platyhelminthes). Cell Tissue Res 226:449–460
Stewart MT, Mousley A, Koubková B, Šebelová Š, Marks NJ, Halton DW (2002) Gross anatomy of the muscle systems and associated innervation of Apatemon cobitidis proterorhini metacercaria (Trematoda: Strigeidea), as visualised by confocal microscopy. Parasitology 126:273–282
Sukhdeo MVK, HSU SC, Thompson CS, Mettrick DF (1984) Hymenolepis diminuta: behavioural effects of 5-hydroxytryptamine, acetylcholine, histamine and somatostatin. J Parasitol 70:682–688
Thompson CS, Mettrick DF (1989) The effects of 5-hydrxytryptamine and glutamate on muscle contraction in Hymenolepis diminuta (Cestoda). Can J Zool 67:1257–1262
Tinsley RC (1983) Ovoviviparity in platyhelminth life-cycles. Parasitology 86:161–196
Ward SM, Allen JM, McKerr G (1986) Neuromusculature physiology of Grillotia erinaceus metacestodes (Cestoda: Trypanorhyncha) in vitro. Parasitology 93:587–597
Yamaguti S (1963) Systema helminthum, vol. IV: Monogenea and Aspidocotylea. Interscience, New York
Zurawski TH, Mousley A, Mair GR, Brennan GP, Maule AG, Gelnar, M, Halton DW (2001) Immunomicroscopical observations on the nervous system of adult Eudiplozoon nipponicum (Monogenea: Diplozoidae). Int J Parasitol 31:779–783
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
El-Naggar, M.M., Arafa, S.Z., El-Abbassy, S.A. et al. Neuromusculature of Macrogyrodactylus congolensis, a monogenean skin parasite of the Nile catfish Clarias gariepinus . Parasitol Res 100, 265–279 (2007). https://doi.org/10.1007/s00436-006-0235-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-006-0235-7