
Abstract
Lactic acid bacteria (LAB) are a heterogeneous group of Gram-positive bacteria that comprise several species which have evolved in close association with humans (food and lifestyle). While their use to ferment food dates back to very ancient times, in the last decades, LAB have attracted much attention for their documented beneficial properties and for potential biomedical applications. Some LAB are commensal that colonize, stably or transiently, host mucosal surfaces, inlcuding the gut, where they may contribute to host health. In this review, we present and discuss the main factors enabling LAB adaptation to such lifestyle, including the gene reprogramming accompanying gut colonization, the specific bacterial components involved in adhesion and interaction with host, and how the gut niche has shaped the genome of intestine-adapted species. Moreover, the capacity of LAB to colonize abiotic surfaces by forming structured communities, i.e., biofilms, is briefly discussed, taking into account the main bacterial and environmental factors involved, particularly in relation to food-related environments. The vast spread of LAB surface-associated communities and the ability to control their occurrence hold great potentials for human health and food safety biotechnologies.
Similar content being viewed by others

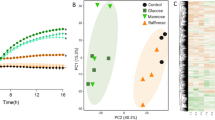
Avoid common mistakes on your manuscript.
Introduction
Within the phylum of Firmicutes, lactic acid bacteria (LAB) constitute a broad and heterogeneous group of Gram-positive microorganisms with low G+C content. Lactobacillus, Lactococcus, Leucocostoc, Pediococcus, Oenococcus, Enterococcus, and Streptococcus represent the main LAB genera (Douillard and de Vos 2014). LAB are asporigen, acid tolerant, rod or cocci shaped, catalase negative, microaerophilic, and share the metabolic feature to produce lactic acid as the major end-product of carbohydrate fermentation (Carr et al. 2002). Highly adapatble and versatile, LAB inhabit a variety of ecological niches, and many of them are in close interaction with humans. They populate diverse food-related habitats, including plants, dairy, milk, wine, and meat and are natural inhabitants of mammalian mucosal surfaces, such as those of the gastrointestinal (GI) tract, oral cavity, and vagina.
Such bacterial group boasts a remarkable association with human food, lifestyle, and health. LAB have a key role in food industry, as they have been traditionally used to drive fermentation processes, thereby contributing to quality and preservation of fermented food. Hence, the widespread consumption of fermented products has determined a regular ingestion of LAB, allowing them to colonize the human body. Besides their technological importance, in the last decades, LAB have attracted a growing interest also for their health-promoting potential and related biomedical applications. Among LAB, lactobacilli comprise several strains which are currently claimed as probiotics (Ouwehand et al. 2002), i.e., “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (Hill et al. 2014). Moreover, thanks to their generally recognized as safe (GRAS) or qualified presumption of safety (QPS) status, LAB can provide safe vectors for the delivery of therapeutic compounds to mucosal surfaces (Bermúdez-Humarán et al. 2013).
Here, we present an update overview of the main aspects concerning with the ability of LAB to colonize and interact with biotic and abiotic surfaces, with particular emphasis on GI mucosa and food-related niches, and focusing on the most investigated species, i.e., probiotics and strains of high technological and biomedical appeal.
Experimental approaches to study LAB colonization on host mucosae and on abiotic surfaces
Allochtonous LAB populate the gut mucosae possibly by forming stable, complex, multispecies biofilms (Schwab et al. 2014). Prompted by exciting medical prospects, a large body of research, mainly based on metagenomic analyses and culture-independent methods, has shed light on the composition and dynamics of the LAB communities associated to GI mucosae, and the details of the microbial structures involved in adhesion and interplay with host tissues are being disclosed more and more. However, the difficulties of sampling have impaired direct investigations on LAB biofilms associated to human GI mucosae in vivo.
Different in vitro and in vivo models have been adopted to study the colonization potential of LAB on host biotic surfaces. In vitro systems that mimick GI conditions represent a convenient approach to assess survival ability of potential probiotic strains (Fernández de Palencia et al. 2009; Bove et al. 2012; Van Bokhorst-van de Veen et al. 2012 ). Studies addressing the microbial adhesion abilities often use in vitro models, such as intestinal epithelial cell lines (e.g., Caco-2 and HT-29), immobilized mucus, or extracellular matrix molecules, including collagen and fibronectin (Le et al. 2013; von Ossowski et al. 2011). Dendritic cells (DC), pheripheral blood mononuclear cells (PBMC), and macrophages are widely adopted to study in vitro the immunomodulatory potential of colonizing LAB, while ex vivo approaches take adavantage of mucosal explants from different animals (Walter et al. 2007; Breshears et al. 2015; Dertli et al. 2015). Of course, the gold standard to investigate the different aspects of host-bacteria interaction relies on in vivo experiments on animals such as mice and other mammals, including human trials (Marco et al. 2010; Van Bokhorst-van de Veen et al. 2012). Mice provide convenient models of the human GI tract, although murine and human guts differ in anatomy, biochemical features, and microbial composition (Nguyen et al. 2015). In recent years, lower vertebrates such as zebrafish (Danio rerio), and even animals representative of other phyla, such as the nematode Cenorhabditis elegans, have emerged as amenable tools for investigating specific aspects of the host-bacteria interplay (Rieu et al. 2014; Russo et al. 2015; Park et al. 2014).
Biofilm formation on abiotic surfaces can be quantitatively analyzed by spectrophotometric methods and molecular approaches. Crystal violet is routinely used to stain LAB biofilms grown on plastic, glass, and other surface types (Arena et al. 2014). Quantitative PCR coupled to propidium monoazide (PMA) allows to quantify microbial cells involved in biofilm formation, discriminating between live and dead cells within the biofilm matrix (Àlvarez et al. 2013; Arena et al. 2015). Scanning electron microscopy, confocal microscopy, the use of fluorescent probes, and image analysis softwares permit to visualize the architecture of biofilms (Kniggendorf et al. 2016).
Colonization of the host gut and transcriptional reprogramming associated to the adaptation to the gut niche
The structure and composition of the gut microbiota varies considerably along the different regions. The size of the resident bacterial community is modest in the stomach and proximal small intestine (bacterial density <104 CFU/ml), where extreme conditions hamper the microbial colonization, while it expands in the distal small intestine, to reach its maximum in the colon (1011 to 1012 CFU/ml) (O’Hara and Shanahan 2006). LAB comprise species that exhibit different degree and mode of persistence in the gut. Some of them are genuinely autochthonous, i.e., they establish a long-lasting association and form stable populations that characterize a specific gut region, within a particular host. Yet, metagenomics and experimental evidences indicate that most LAB are allochthonous, i.e., transient colonizers that originate from food or the oral cavity and are thus introduced into the gut accidentally, or deliberately. LAB reach the maximal density within the large and small intestines, where they generally account for less than 1% of the total resident bacteria (Douillard and de Vos 2014). In the gut regions with relatively small endogenous microbial communities, the temporal abundance of dietary, passenger LAB, including probiotics, is thought to have a major impact on the host physiology (Kleerebezem and Vaughan 2009; Douillard and De Vos 2014).
A prerequisite to the intestinal colonization is the capacity to survive the harsh conditions of the GI tract, including the lytic action of digestive enzymes, the detergent-like activity of bile salts, the extremely low pH in the stomach, the oxidative and hyperosmotic stress in the colon, and the antimicrobial activity of the host immune effectors (Fig. 1). Gastric acidity usually represents the main hurdle to lactobacilli survival, whereas intestinal conditions are better tolerated (Kleerebezem and Vaughan 2009). Survival to GI transit and persistence in the host gut have been extensively demonstrated for several probiotic lactobacilli, both in vivo, including human feeding experiments (Marco et al. 2010; Van Bokhorst-van de Veen et al. 2012; David et al. 2014; Klijn et al. 1995; Sepp et al. 1993; Goldin et al. 1992; Kankainen et al. 2009; Pridmore et al. 2004), and in vitro, by using simplified systems that simulate GI conditions (Fernández de Palencia et al. 2008; Bove et al. 2012; Van Bokhorst-van de Veen et al. 2012).

Representation of biotic and abiotic surface features and bacterial molecules promoting adhesion and biofilm production. Biotic surfaces (on the left), such as gut mucosa, can be a hostile environment for bacteria, which are challenged by harsh conditions including the detergent-like activity of bile salts, low pH, host immune factors, variable nutrient conditions, the presence of other bacteria, and a mucus layer. LAB possess the abilities to survive unfavorable conditions, protect themselves from host immune response, compete for nutrients, inhibit other microorganisms, and, in general, interact with biotic surface components. The formation of biofilm on abiotic surfaces (on the right) is also determined by several aspects, such as roughness, temperature, stiffness, type of material, charge, hydrofobicity or hydrophilicity, topography, pH, and coating. Moreover, the composition of the bacterial cell surface and the molecules produced by LAB have a fundamental role in determining the attachment, the development, the maturation and, possibly, the disruption of the biofilm. BSH bile salt hydrolase, EPS exopolysaccharides, MUB mucus-binding proteins, MLP moonlighting proteins, AMP antimicrobial peptides, TLR Toll-like receptor, AIP auto-inducing peptides, AI-2 LuxS-derived autoinducer-2, MabA modulator of adhesion and biofilm A, eDNA extracellular DNA, TA teichoic acids
Passage along the GI tract requires LAB to reprogram their gene expression, thereby allowing adaptation to such peculiar environment (Marco et al. 2010). Interestingly, GI features and stress, such as bile and acid, act as environmental stimuli that induce those metabolic and structural changes allowing LAB to colonize the gut (Bron et al. 2004a; Koskenniemi et al. 2011; Bove et al. 2013). In probiotic lactobacilli, in vitro gastric stress simulation determined not only an increased expression of several stress-related genes but also an increment of factors potentially involved in mucosal adhesion and in competitive behavior towards other microbes, which might be necessary in the overcrowded intestinal niche (Bove et al. 2013; Weiss and Jespersen 2012). In oral and gut commensal LAB, mucins were found to promote microbial proteolytic activity by enhancing the expression of cell surface-associated proteases (Kindblom et al. 2012; Wickström et al. 2013). As mucins are degraded by such proteases, it is suggested that they could be exploited as a nutrient source by the colonizing bacteria. Therefore, their effect would prepare LAB to survival on mucosal surfaces. In probiotic lactobacilli, the genetic response to in vitro bile stress comprises the upregulation of chaperones and membrane-associated functions, including proteins potentially involved in bile detoxification, as well as the induction of redox enzymes, thus suggesting a link between bile exposure and oxidative stress (Bron et al. 2004b; Pfeiler et al. 2007; Whitehead et al. 2008). Moreover, a clone-based DNA microarray transcriptome analysis showed that bile exposure modulates several Lactobacillus plantarum genes encoding cell envelope-related functions, thus indicating major consequences on the integrity and functionality of the cell envelope (Bron et al. 2006). Following transcriptomic and proteomic analyses have confirmed that a reorganization of the cell envelope characterizes the bile stress response also in other lactobacilli (Koskenniemi et al. 2011; Alcántara and Zúñiga 2012). In L. plantarum, passage through mouse GI tract was found to activate the following: (i) stress genes, which reflect the hostile conditions of the gut habitat, (ii) genes encoding surface-anchored proteins, which could mediate interaction with host cells or with soluble components of the gut lumen, and (iii) genes involved in sugar metabolism and in acquisition and biosynthesis of non-sugar compounds (i.e., amino acids, nucleotides, cofactors, and vitamins), which may be poorly available in the gut (Bron et al. 2004a). Denou et al. (2007) showed that Lactobacillus johnsonii transcribes specific sets of genes in the different compartments of mice GI tract, including sugar transport systems (in the stomach, jejunum, and cecum) and multidrug transport systems (in the stomach). A combination of genotyping and expression microarrays was adopted to find L. johnsonii genes associated to a longer gut persistence: three genetic loci were identified, including (i) a gene involved in sugar transport (i.e., a phosphotransferase system (PTS) transporter), which confirms the importance of sugar catabolism for gut commensalism, (ii) an operon involved in exopolysaccharides (EPS) synthesis, and (iii) a gene annotated as putative IgA protease (Denou et al. 2008). A comparative study of the trascriptome of a commensal probiotic LAB (i.e., L. plantarum 299v), during passage along murine and human gut, revealed a substantial convergence in the adaptive response to these two niches (Marco et al. 2010), which involved the modulation of carbohydrate metabolism and cell surface properties. In the murine forestomach, the transcriptome of lactobacilli biofilms was characterized by abundant transcripts related to glucose and maltose utilization, peptide degradation, amino acid transport, and enzymes that would enable the utilization of mucus and cellulose as possible substrates (Schwab et al. 2014). Moreover, genes encoding pathways that enhance tolerance to oxidative and acid stress and extracellular proteins involved in adhesion and/or biofilm formation were significantly induced. More recently, using a rhesus macaque in vivo model, Golomb et al. (2016) observed that L. plantarum adapts to the small intestine by expressing genes required for tolerating oxidative stress, modifying cell surface composition, and consumption of host glycans.
Genetic traits that promote the ecological fitness to the gut
Comparative genomics has revealed that the adapation of LAB to diverse ecological niches has proceeded by the gain of new genes and the degeneracy or loss of no longer necessary functions (Cai et al. 2009; Douillard et al. 2013; Ceapa et al. 2016). For instance, LAB adapted to food matrices such as dairy habitats have lost metabolic activities that are non-essential in such nutrient-rich, relatively constant, and poorly competitive environments. By contrast, LAB colonizing the intestine, a niche characterized by variable nutrient avalaibility, peculiar biochemical-physical stress, and complex microbial communities, exhibit broader metabolic flexibility and specific lifestyle functions essential for survival in the gut.
As indicated by functional genomic studies, the main genes that confer LAB adequate fitness to the GI tract are those that promote survival, metabolic activities, and interactions with host and endogenous microbiota. More in detail, they include genes involved in bile resistance, sugar transport and utilization, mucus-binding capacity, EPS/biofilm production, and defense systems (Klaenhammer et al. 2008; Denou et al. 2008; Pridmore et al. 2004; Altermann et al. 2005; Douillard et al. 2013; Azcarate-Peril et al. 2008; Cai et al. 2009; Douillard and de Vos 2014).
Bile tolerance
Bile salt hydrolases (BSH) activity is prevalent in commensal bacteria of the GI tract, including probiotic LAB. Multiple BSH homologs characterize the genome of some intestine-adapted strains, such as L. plantarum WCFS1 and Lactobacillus acidopihlus NCMF (Begley et al. 2006). BSHs were reported to enhance bacterial resistance to bile acid and in vivo survival in the GI tract, furthermore, metagenomic analyses confirmed that such enzymes represent a conserved microbial adaptation to the human gut environment (Jones et al. 2008). BSHs would benefit commensal LAB by contrasting the effects of bile acids and by inducing cell surface modifications that protect against host-defense systems, including lytic enzymes and antimicrobial peptides (Begley et al. 2006). Putative bile trasporters genes were identified in Lactobacillus gasseri (Azcarate-Peril et al. 2008) and L. johnsonii (Elkins et al. 2001; Pridmore et al. 2004); moreover, they were transcriptionally upregulated by artificial GI environment in L. plantarum (Bron et al. 2004a). Multidrug resistance (MDR) transporters were demostrated also to contribute to bile detoxification in L. acidophilus NCMF (Pfeiler and Klaenhammer 2009) and in Lactobacillus reuteri (Whitehead et al. 2008).
Metabolic features
The intestinal environment is dynamic and nutritionally variable: it may be even a nutrient and carbon source-limited habitat, depending on the gut section and on the physiological state of the host. Moreover, the high bacterial density results in a strong competition for nutrients. On the other hand, thanks to the complex and rich microbial community, LAB can even rely on metabolic cooperations for their nutritional and biosynthetic needs. The utilization of carbohydrate resources typically found in the GI niche characterizes not only the major members of the intestinal microbiota, such as Bacteroides or Bifidobacterium, but also intestine-adapted lactobacilli. Intestinal LAB share the ability to use diverse fermentable carbohydrates, which can derive either from dietary components, such as complex sugars that escape host digestion, or from host extracellular components. Accordingly, their genome exhibit broad gene repertoirs related to carbohydrate utilization, including diversified sugar hydrolases and phosphotransferase system (PTS) sugar transporters (Pridmore et al. 2004; Altermann et al. 2005; Denou et al. 2008; Azcarate-Peril et al. 2008; Claesson et al. 2006). Clearly, the ability to ferment host-derived glycans constitutes a competitive advantage for persistence in the gut. An example of such capacity is provided by l-fucose, which occurs in human secretions and is a constituent of the intestinal mucin glycans, therefore it is abundant in the GI niche and represents a potential carbon source for members of the commensal microbiota. Recently, Becerra et al. (2015) demostrated that Lactobacillus rhamnosus LGG possesses a functional l-fuc operon that enables the fermentation of such sugar, thus improving its fitness to the host intestine.
In silico analyses of lactobacilli commonly found in the GI tract also predict a considerable degree of auxotrophy for amino acids, vitamins, and cofactors (Pridmore et al. 2004; Altermann et al. 2005; Claesson et al. 2006; Azcarate-Peril et al. 2008; Frese et al. 2011). Such limited biosynthetic abilities are often compensated by the presence of a large number of genes encoding peptidases and related transport systems, which allow the uptake and utilization of exogenous metabolites and nutrients, when available in the gut (Schwab et al. 2014).
The relevance of secretome and exoproteome
The extracellular features of LAB, including the non-proteinaceous components, are pivotal for their interaction with host factors during colonization of the GI tract. Zhou et al. (2010) compared the secretome of several LAB, identifying clusters of secreted proteins specific for intestine-adapted species. In this regards, the role of sortases (i.e., transpeptidases that anchor surface proteins to the peptidoglycan layer) and LPxTG-anchored proteins has been emphasized (Call and Klaenhammer 2013). However, contrasting findings describe the influence of such proteins on mucosal adhesion and gut persistence of LAB. For instance, in Lactobacillus salivarius, the absence of sortases significantly reduced adherence to Caco-2 cells (van Pijkeren et al. 2006), whereas Remus et al. (2013) showed that sortase deficiency did not affect the persistence or survival of L. plantarum in murine GI tract, though sortase-dependent proteins could be ascribed an active role in host immunomodulation. Accordingly, the removal of sortase was shown to affect immune modulation of DC by both L. acidophilus and L. gasseri, while sortase-deficient L. acidophilus had an impaired ability to persist in the GI tract of germ-free mice (Call et al. 2015). In order to decipher the adaptation process allowing probiotic persistence in the GI tract, van Bokhorst-van de Veen et al. (2013) repeatedly exposed L. plantarum WCFS1 to mouse GI tract by repeated rounds of feeding and re-feeding. Such approach resulted in the isolation of intestine-adapted derivative strains that exhibited enhanced GI tract robustness and were characterized by genomic modifications (i.e., SNPs and single nucleotide insertions) concentrated in regions encoding functions related to cell envelope and energy metabolism. Apart from demonstrating that it is possible to enhance and extend the probiotic persistence in the GI tract, such finding once more corroborates the importance of cell-envelope remodeling to promote LAB ecological fitness to the gut.
Adhesive capacity
The relevance of mucin-binding capacity in LAB adaptation to the intestinal niche is confirmed by the remarkable number of adhesin-encoding genes in the genomes of gut isolates of L. johnsonii, L. acidophilus, L. gasseri, and L. reuteri (Pridmore et al. 2004; Altermann et al. 2005; Azcarate-Peril et al. 2008; Frese et al. 2011). The long intestinal persistence of L. rhamnosus GG, a well-known probiotc LAB, reflects its excellent mucus-adhesive capacity, which seems mainly ascribable to strain-specific fimbriae-like structures, i.e., pili (Kankainen et al. 2009; Lebeer et al. 2012). Interestingly, comparative genomics and phenotypical analysis of several L. rhamnosus strains from different ecological niches revealed that the functional SpaCBA pili gene cluster is significantly more prevalent in human isolates than in dairy isolates (Douillard et al. 2013). Moreover, among human isolates, mucus-binding pili are produced by intestinal strains, but not by strains from mouth and vagina, which highlights a key role for pili in the GI habitat and reflects a mechanism of niche specialization.
Competition and defense systems
Production of bacteriocins may give a relevant competitive advantage to LAB inhabiting densely populated niches, such as the gut. Comparative analysis highlighted the presence of megaplasmid-encoded bacteriocin genes in several L. salivarius human intestinal isolates (Raftis et al. 2011). Accordingly, most L. rhamnosus strains isolated from the human body, including gut and vagina, were found to produce antimicrobial compounds against common human pathogens, while dairy isolates appeared to have lost such trait (Douillard et al. 2013). A consistent number of putative genes implicated in the production and processing of bacteriocins was predicted also in L. acidophilus NCMF (Altermann et al. 2005) and bacteriocin genes characterized a L. johnsonii strain with high gut persistence (Denou et al. 2008). Moreover, the induction of the plantaricin immunity protein PlnI, during murine Gl transit of L. plantarum (Marco et al. 2007, 2009), reinforces the idea that bacteriocins enhance the in vivo performance of LAB. Intriguingly, some studies even suggested that LAB bacteriocins, like host antimicrobial peptides, might modulate immune response of host cells, including DC and PBMC (Meijerink et al. 2010; van Hemert et al. 2010).
Protection from the host environnment
The prevalence of EPS biosynthesis operons in many intestine-adapted LAB strains (Azcarate-Peril et al. 2008; Denou et al. 2008; Lebeer et al. 2009; Raftis et al. 2011; Claesson et al. 2006) strongly supports a role for such surface components in interaction with host and protection from gut environment, as was plainly demonstrated for L. rhamnosus GG (Lebeer et al. 2011). Intriguingly, in L. salivarius, which includes strains indigenous to the gut and oral cavity, two gene clusters for EPS biosynthesis corresponded to regions of significant, intraspecies, genomic diversity, hence emphasizing their contribution to niche adapatation (Raftis et al. 2011).
Mechanims and molecules promoting adhesion to mucosal surfaces and host-microbe interplay
A mucus layer protects the epithelium of hollow organs (including airways, mouth, gut, and vagina) and serves as a niche for commensal microbes. The mucus consists mainly of large, glycosylated proteins, i.e., mucins, and contains also other proteins, lipids, and glycolipids (Juge 2012). Bacterial adhesion to the mucosal surface is the fisrt step for the successful colonization of the host surfaces, and LAB have evolved multiple factors to realize it (Fig. 1). With few exceptions (Mukai et al. 2002; Nishiyama et al. 2013, 2016; Tytgat et al. 2016a), the biochemical details of such interactions (e.g., the nature of the host glycan receptor) are still elusive and difficult to determine, because of the heterogeneity of mucus components, the multitude of factors involved, and the overall complexity of the adhesion process (Juge 2012). Yet, it is clear that the architecture of the microbial cell envelope is crucially involved in the mechanisms underlying colonization, persistence, and interaction with host.
Structure and composition of the cell wall determines surface properties, which, in turn, influence adhesiveness. The cell membrane of LAB is wrapped by a multilayered peptidoglycan (PG) shell, which is decorated with teichoic acids (TA; including lipotheicoic acid (LTA) and wall teichoic acid (WTA)), pili, proteins, and EPS. In some species, the PG shell may be futher surrounded by a paracristallyne envelope of S-layer proteins (Chapot-Chartier and Kulakauskas 2014). All of these components have been shown to be involved, to different extent, in the adhesive properties of LAB.
TA
TA are anionic, amphiphilic polymers, which contribute to the hydrophobic character and the electrostatic charge of the microbial cell surface, thereby influencing its adhesiveness. d-Alanylation (d-Ala) of TA is widespread among Gram-positive bacteria, suggesting its biological relevance in a variety of habitats. d-Ala seems also important for the host-commensal relationship in the GI tract. LTA were important to mediate the attachment of L. johnsonii La1 to human intestinal epithelial cells (Granato et al. 1999). More recently, the importance of TA in the colonization of murine gut was confirmed in vivo for L. reuteri, besides suggesting that their d-Ala modification might be specifically protective against the unfavorable conditions of murine forestomach, including high acidity and host, cationic antimicrobial peptides (Walter et al. 2007). Likewise, the incorporation of d-Ala residues into LTAs was shown to enhance virulence, resistance to antimicrobial peptides, and adherence capacity of several pathogenic strains (Abachin et al. 2002; Kristian et al. 2005; Poyart et al. 2003; Collins et al. 2002), which emphasizes how commensal lactobacilli and bacterial pathogens may adopt similar strategies to colonize the mammalian host. Interestingly, LTA and d-Ala-substituted LTA are recognized as microbe-associated molecular patterns (MAMP) by host cells and, given their structural diversity, can induce species- and strain-specific immune responses, as was extensively demonstrated, both in vitro and in vivo, for diverse probiotic lactobacilli (Matsuguchi et al. 2003; Grangette et al. 2005; Claes et al. 2010, 2012; Perea Velez et al. 2007; Mohamadzadeh et al. 2011; Smelt et al. 2013).
EPS
In LAB, EPS participate to several functions, including stress tolerance, biofilm formation, communication with other microbes, and interaction with host cells (Caggianiello et al. 2016). Being exposed on the bacterial surface, EPS contribute its physicochemical characteristics and influence the adhesion properties. Several studies have addressed the effects of EPS on the ability of LAB, especially lactobacilli, to colonize the host surfaces. For LAB indigenous to the oral cavity, EPS form a matrix that promotes cell aggregation and biofilm formation on the dental surfaces (Banas and Vickerman 2003; Parisotto et al. 2010). The EPS layer can mask adhesion factors on the microbial cell surface and/or compete for adhesion on host-binding sites. Accordingly, EPS were usually found to hinder the recognition mechanisms required for a stable adherence of LAB on host mucosae (Ruas-Madiedo et al. 2006; Lebeer et al. 2009; Denou et al. 2008; Dertli et al. 2015; Lee et al. 2016). However, the contribution of EPS to the overall colonization process is somehow controversial. In L. plantarum, EPS removal affected adhesion on Caco-2 cells in a strain-dependent fashion (Lee et al. 2016). In L. rhamnosus GG, knockout of the gene responsible for EPS biosynthesis deprived the derivative mutant strain of long, galactose-rich EPS molecules, resulting in enhanced adherence to human intestinal epithelial cells and increased biofilm formation (Lebeer et al. 2009). However, the mutant exhibited lower survival in vivo, within the murine GI tract, probably because EPS protect bacterial cells from host innate immune factors, such as complement-mediated lysis and cationic antimicrobial peptides (Lebeer et al. 2011). Likewise, in L. johnsonii, reduction of EPS promoted autoaggregation, biofilm formation, and adhesion to chicken gut explants, yet it diminished its resistance to some stress, which could mine its ability to reach and survive in a highly competitive and stressful niche, as is the gut (Dertli et al. 2015). In L. reuteri TMW1.106, a sourdough isolate, the removal of EPS by gene deletion, impaired cell aggregation, biofilm formation, and in vivo colonization of mouse GI tract (Walter et al. 2008). In another L. reuteri strain, common inhabitant of the murine proximal gut, the loss of EPS by genetic knockout (KO) did not affect biofilm formation and allowed mouse gut colonization, even though this was significantly impaired under competition with the wild-type, parental strain (Sims et al. 2011). In this regard, the authors hypothesize that, after adopting a commensal relationship with vertebrate hosts, EPS might have assumed novel additional roles, relative to their original function. More in detail, L. reuteri EPS could enhance its colonization potential, for instance by exerting immune modulatory effects that would generate immunological tolerance towards the commensal (Sims et al. 2011). In Pediococcus strains isolated from wine and cider, EPS positively influenced probiotic properties and adhesion on Caco-2 cells (Fernández de Palencia et al. 2009; Garai-Ibabe et al. 2010; García-Ruiz et al. 2014). Moreover, exogenous addition of Pediococcus EPS potentiated in vitro adhesion of L. plantarum WCFS1 to human intestinal epithelial cells (Russo et al. 2012) and a recombinant strain of L. paracasei NFBC 338, expressing the glucosyltransferase gene of P. parvulus 2.6, produced β-glucan EPS and exhibited increased tolerance to GI stress (Stack et al. 2010).
EPS are known to participate to host-microbe interaction also by their immunoregulatory properties, including anti-inflammatory effects, as recently demonstrated for some LAB (Notararigo et al. 2014; Tang et al. 2015; Murofushi et al. 2015). Intriguingly, the consequences of EPS removal/production on characteristics related to the interaction with the host, such as adhesion, gut survival, and immune modulation, may not be easily predictable as they seem species and strain dependent (Walter et al. 2008; Sims et al. 2011; Lee et al. 2016). To sum up, given their documented protective role and their prevalence in lactobacilli isolated from gut ecosystems, EPS may afford LAB a greater ecological performance during gut colonization.
PILI
Adhesion of LAB to host surfaces is mainly protein mediated. Among the proteinaceous components of the cell envelope, pili have been shown to be pivotal for adhesion to host tissue and are thus considered to favor colonization and persistence of Gram-positive bacteria (Danne and Dramsi 2012). Pili were demostrated to promote adhesion to host cells by commensal opportunistic LAB, including Streptococcus agalactiae, a commensal bacterium colonizing the gastrointestinal and urogenital tract of women (Dramsi et al. 2006) and Streptococcus pneumoniae, a common colonizer of the human upper respiratory tract (Barocchi et al. 2006). Pili are also essential virulence factors as they mediate host tissue colonization by potentially pathogenic enterococcal species (Nallapareddy et al. 2011a, b).
The SpaCBA pilus of L. rhamnosus GG was immunodetected on the bacterial cell surface (Kankainen et al. 2009) and shown to be critical for efficient adherence to human intestinal epithelial cells and for biofilm formation (von Ossowski et al. 2010; Lebeer et al. 2012). Oral assumption of piliated and non-piliated mutant strains also suggested a key role for pili in promoting residence in the human colon of healthy volunteers (Kankainen et al. 2009). Recently, another type of GG pilus was phenotypically characterized by heterologous expression of the spaFED operon in Lactococcus lactis, demonstrating cell wall surface localization and mucus-binding capacity (Rintahaka et al. 2014). Surface proteome analysis of a natural vegetal isolate of L. lactis allowed the detection of pilins, which are involved in adhesion to Caco-2 cells (Meyrand et al. 2013; Le et al. 2013). Notably, the gene cluster involved in L. lactis pili biogenesis is located on a plasmid, thus indicating a recent gain by horizontal gene tranfer and suggesting a mechanism underlying the spread of this function among plant lactococci.
Apart from their well-documented relevance in adhesion, some recent studies shed new light on the role of pili in immune interaction with the host. L. rhamnosus GG SpaCBA pili were shown to mediate, in vitro, the adhesion to macrophages and suggested to promote anti-inflammatory effects (Vargas García et al. 2015). Tytgat et al. (2016a) showed that SpaCBA pili are post-translationally glycosylated and their glycans are recognized by human DC through the DC-SIGN lectin receptor, thereby modulating cytokines production. A recent paper demontrates that SpaCBA pili prevent mucus adhesion by potential pathogens, i.e., Enterococcus faecium, thus providing a molecular basis for the succesfull clinical use of GG in the prevention and treatment of vancomycin-resistant enterococci infections (Tytgat et al. 2016b).
MUB proteins
Mucus-binding proteins (MUB) constitute a family of peptidoglycan-anchored proteins that play a relevant role in LAB adhesion to the mucus layer (Juge 2012). MUBs own multiple Mub repeats, i.e., the domains involved in binding to mucus, and a C-terminal LPxTG anchoring motif, for their covalent attachment to the bacterial cell wall. The first identified and functionally characterized MUB was that from L. reuteri (Roos and Jonsson 2002; MacKenzie et al. 2009, 2010). A MUB-related protein with mannose-specific adhesion (Msa) capacity was then identified in different L. plantarum strains (Pretzer et al. 2005; Gross et al. 2010). In L plantarum 299v, Msa was ascribed a role in bacterial adherence and in the induction of host responses in the pig intestine (Gross et al. 2008). Furthermore, variations in mannose-adhering capacity of different L. plantarum strains were related to the genetic variability in the msa gene locus, e.g., number of MUB domain repeats (Gross et al. 2010). A high genetic heterogeneity of MUB and MUB-like proteins was also observed among L. reuteri isolates, and the proteins could be immunodetected on the cell surface only in a few strains (Mackenzie et al. 2010). Mucus-binding adhesins have been identified even in other probiotic lactobacilli, and their contribution to the interaction with intestinal cells and/or mucus was demonstrated (Buck et al. 2005a, b; Von Ossowski et al. 2011; Jensen et al. 2014). Notably, a genome and protein database search for MUB domain-containing proteins revealed that potential mucus adhesins occur only in LAB and are particularly abundant in lactobacilli of the GI tract, thus pointing to the Mub repeat as a conserved and relevant functional unit for host-microbe interactions, which could be the outcome of a long-term co-evolution (Boekhorst et al. 2006).
Moonlighting proteins
The so-called moonlighting proteins constitute a remarkable family of multifunctional proteins that fulfill multiple, biologically unrelated roles, often localized to separate cellular compartments. Some moonlighting proteins have been demonstrated to be cell surface associated, thereby acting as adhesins, in addition to their primary, usually intracellular, function. In diverse lactobacilli, the elongation factor Tu (EF-Tu) (Granato et al. 2004; Dhanani and Bagchi 2013; Nishiyama et al. 2013), heat shock proteins (Bergonzelli et al. 2006; Katakura et al. 2010), and glycolytic enzymes (Kinoshita et al. 2008; Ramiah et al. 2008; Patel et al. 2016; Glenting et al. 2013; Spurbeck and Arvidson, 2010; Castaldo et al. 2009; Kainulainen et al. 2012; Katakura et al. 2010) have been recognized as multitasking proteins with the special ability to bind proteinaceous components of the extracellular matrix. Interestingly, such proteins are anchorless, i.e., not covalently bound to the cell wall; moreover, they lack signal sequences or hydrophobic membrane-spanning domains that could target them to secretory pathways. Indeed, the mechanisms allowing their translocation and envelope anchoring are likely to be species specific and are, as yet, poorly understood. Intriguingly, it was suggested that environmental challenges and biochemical features typical of the host gut, including starvation, bile acids, pH stress, and antimicrobial peptides, could modulate availability, adhesiveness, cell surface association, and extracellular release of such multifunctional proteins (Antikainen et al. 2007; Saad et al. 2009; Candela et al. 2010; Kainulainen et al. 2012; Bove et al. 2013). This phenomenon highlights how the host signals, including its innate immune effectors, can impact the cell surface architecture of commensal lactobacilli, thereby promoting their adhesiveness. Multitasking proteins also comprise putative subunits of ATP-binding cassette (ABC) transport systems that were characterized as potential adhesins in L. reuteri NCIB11951 (Roos et al. 1996), L. fermentum (Macías-Rodríguez et al. 2009), and Lactobacillus mucosae ME340 (Watanabe et al. 2010).
S-layer
The bacterial cell surface commonly features paracrystalline protein arrays referred to as S-layers (Sára and Sleytr 2000). In LAB, the functions of S-layer proteins (Slp) are still elusive, although acumulating evidence underpins a role in host-microbe interactions. In Lactobacillus crispatus, Lactobacillus brevis, and L. acidophilus, Slp were shown to mediate adhesion to proteins of the host extracellular matrix (Sillanpää et al. 2000; Hynönen et al. 2002; Buck et al. 2005a, b). Accordingly, Avall-Jääskeläinen et al. (2003) elegantly demonstrated that the ability to adhere to gut epithelial cells could be transferred to a poorly adhesive LAB, i.e., L. lactis, by heterologous expression of the N-terminal region of the Slp from L. brevis. Moreover, in L. acidophilus NCFM, the S-layer protein A (SlpA) was found to regulate dendritic and T cell functions by specific binding on their surface lectin receptor DC-SIGN (Konstantinov et al. 2008).
Colonization of abiotic surfaces by LAB
Several LAB are able to colonize surfaces by forming biofilms, i.e., sessile bacterial communities, strongly associated to a surface and embedded in a self-produced, extracellular, polymeric matrix (Piard and Briandet 2015). LAB biofilms occur on both biotic and abiotic surfaces, in different environments, including plant material, food and food-related niches, animal mucosae, medical instrumentations, and domestic settings. Biofilm bacteria exhibit distinctive phenotypic traits. Moreover, the biofilm structure provides protection against environmental stresses, such as unfavorable pH and oxygen values, biocides, antibiotics, and other hostile factors (Watnick and Kolter 2000; Kubota et al. 2008). For instance, the resistance to disinfectants is significantly higher in biofilms than in planktonic cells (Bridier et al. 2011). Likewise, the organization of LAB in communities anchored to a surface gives greater resilience compared with the planktonic counterparts (Somers et al. 2001; Kubota et al. 2009). In probiotic lactobacilli, the biofilm mode of life enhances resistance to OGI conditions, moreover, it modulates probiotic functions, such as anti-inflammatory effects and antagonism against pathogens (Rieu et al. 2014; Aoudia et al. 2016).
The development and endurance of biofilms are influenced by several aspects, including the type of biofilm-producing strains, the symbiotic relationship between different species and/or strains taking part in the biofilm construction, the nature of the surface and other unpredictable factors, such as moisture and nutrient availability (Van Acker et al. 2014). The food industry, especially during the manufacturing process, offers a number of dynamics conducive to the biofilm formation (Gunduz and Tuncel 2006). LAB, both autochthonous to the raw material and as inoculated starters, are widely used in food preparation, hence, biofilms formed by LAB have been reported in different products (Piard and Briandet 2015). In several cases, LAB biofilms may alter the quality of food, as they produce molecules which modify the original aroma profile and texture (Suzuki et al. 2008; Fernández Ramírez et al. 2015). The contamination of manufacturing plants of very common food products can be caused by biofilm-producing LAB such as Lactobacillus fructivorans (responsible for mayonnaise and miso spoilage), Lactobacillus acetotolerans and L. brevis (vinegar spoilage), L. plantarum subsp. plantarum (pickled cabbage spoilage), and Lactobacillus curvatus and Lactobacillus fermentum (found on stainless steel utensils and ripening vats of Cheddar cheese). The biofilms produced by several other LAB species have been reported to affect the quality of meat, cheese, sake, beer, and salad (Somers et al. 2001; Kubota et al. 2009) and well known is the undesirable alteration provoked by biofilms of Pediococcus and Lactobacillus species in wine (Lonvaud-Funel 2016).
Although the biofilms formed on food and food processing plants usually spoil the products and damage both equipments and working surfaces (Flemming and Wingender 2010), yet, in some manufacture, biofilms are advantageous for the food technology. For example, in the production of the traditional cheeses Italian Ragusano and French Salers, both made from raw milk, the wooden vats, used for fermentation and ripening, host a microbial biofilm (formed mainly by Streptococcus thermophilus, Lactobacillus, Lactococcus, Enterococcus, and Leuconostoc species) which is desired and, more precisely, essential for achieving the uniqueness of the product (Licitra et al. 2007; Didienne et al. 2012). Additionally, the biofilm on vat surfaces not only governs the fermentation processes without the use of any starter, it also inhibits spoilage and potentially pathogenic microorganisms (Mariani et al. 2007). Strains of L. plantarum isolated from biofilms on the floor of poultry processing plants were found to contrast the development of Listeria monocytogenes (Zhao et al. 2013). Thus, the biofilm may contribute to food safety. Moreover, LAB biofilms can improve the properties of the final product and extend its shelf life, as is the case of fermented olives, whose surface and fermentation equipment host Latobacillus pentosus communities, and sausages, which are produced in vessels colonized by Lactobacillus sakei biofilms (Arroyo-Lopez et al. 2012; Landeta et al. 2013). Similarly, Leuconostoc biofilms can reduce the sugar crystallization during sucrose refining, due to the presence of dextran in the biofilm matrix (Leathers and Bischoff 2011).
Microbial biofilms occur also on numerous indwelling medical devices, including contact lenses, catheters, connectors, tubes, and valves, thus representing a threat for public health. The colonization of the abiotic surfaces of medical instruments is facilitated by the presence of body fluids and liquid medications (Donlan 2001). Examples of LAB, which can contaminate medical instrumentations, are Enterococcus faecalis and Streptococcus viridans, which typically derive form the skin of patients or healthcare workers.
Mechanisms associated to biofilm formation on abiotic surfaces
Bacterial factors
When bacterial cells, commonly from diverse species, organize themselves in a biofilm, the communication between the different members of the community plays a crucial role. The fundamental mechanism of coordination allowing the structural organization of the biofilm is known as “quorum sensing” (Di Cagno et al. 2011). Bacterial cells perceive changes from the environment and exchange information with other bacteria, by producing small, diffusible molecules that modulate the adaptation and development of the biofilm consortium. In the formation of LAB biofilm, a major role in the cell-to-cell communication system has been ascribed to two types of signal molecules called auto-inducing peptides (AIPs) and LuxS-derived autoinducer-2 (AI-2) (Choudhary and Schmidt-Dannert 2010; Lebeer et al. 2007a) (Fig. 1). Such compounds are secreted through specific export systems. When the concentration of AIPs and AI-2 remains below a threshold value, their specific receptors, located either on the cell membrane or intracellularly, respectively, are degraded; when the signal molecules accumulate in the extracellular microenvironment, their receptors are activated and trigger a trascriptional cascade that leads to the modulation of several genes involved in the biofilm production (Stock et al. 2000; Bassler and Losick 2006). Growth phase-dependent AIP systems have been identified in L. plantarum WCFS1 (Sturme et al. 2005) and E. faecalis (Hancock and Perego 2004), while an AI-2 complex has been identified in L. rhamnosus GG (Lebeer et al. 2007b). Both systems have been associated to the modulation of adherence to abiotic surfaces, and the regulation of gene involved in production of polysaccharides and cell membrane protein.
Quorum sensing is not the only mechanism through which the biofilm formation is initiated and preserved. Different cell surface molecules and structures are implicated in adhesion and aggregation of planktonic bacteria (Fig. 1). Notably, the same microbial factors often drive the colonization of both abiotic and biotic supports, thus emphasizing similarities in the mechanisms leading to biofilm formation in the different niches where LAB may live (Piard and Briandet, 2015). Pili and fimbriae contribute to the initial stage of biofilm formation by mediating autoaggregation and adhesion of LAB (Mandlik et al. 2008; Oxaran et al. 2012; Adlerberth et al. 1996). Moreover, dedicated enzymes can be committed to the first phase of biofilm formation, such as sortase A, which was found to promote cell-to-cell and cell-to-surface interactions in E. faecalis and L. plantarum (Guiton et al. 2009; Malik et al. 2013). MabA was characterized as a cell wall protein that modulates adhesion and biofilm formation in L. rhamnosus GG (Vélez et al. 2010), and its higher expression level was associated to enhanced biofilm formation capacity (Savijoki et al. 2011).
Although it is not yet clear how and in which step, EPS are crucially involved in the biofilm construction. EPS physically support the biofilm structure and contribute to make up a sticky matrix that encloses a protective microenvironment and promotes bacteria-bacteria and bacteria-surface contacts (Caggianiello et al. 2016). In LAB, the ability to produce EPS, their chemical features and extracellular localization are strain-specific traits (Monsan et al. 2001; Lee et al. 2016). An assortment of glucosyltransferase makes some LAB strains prone to the production of biofilm by forming polysaccharide matrices constituted by different monosaccharides and specific glycosidic bonds (Theilacker et al. 2011).
Extracellular DNA (eDNA) is essential for determining the 3D structure of biofilm. Indeed, LAB strains lacking autolysin, the enzyme involved in the extracellular release of DNA, loose the ability to form structured biofilms (Mercier et al. 2002; Guiton et al. 2009). Additionally, teichoic and lipoteichoic acids play a crucial role in the adhesion to surfaces by changing the net negative charge of bacterial membrane and, consequently, its cohesive capability (Fabretti et al. 2006).
Environmental factors
The induction of the biofilm is also influenced by specific environmental parameters (Donian, 2002). Generally, microorganisms take advantage by the biofilm growth mode when some environmental conditions become unfavorable to their survival, e.g., starvation or nutrient-rich conditions, low pH, and temperatures outside the optimal range (Myszka and Czaczyk 2009; Kubota et al. 2009). While, on the one hand, the composition of the bacterial cell membrane and the microbial molecules have a fundamental role in determining the attachment, development, maturation and, eventually, the disruption of the biofilm, on the other hand, an equally important role is played by the intrinsic properties of the abiotic surface, especially in the initial step of the adhesion process (Shi and Zhu 2009). Thus, the material type, coating, roughness, free energy, charge, topography, and stiffness may all modulate the bacterial colonization (Hahnel et al. 2015) (Fig. 1).
The topographical features of the surface, particularly its roughness, influence the bacterial adhesion, as a greater extent of irregularities promotes the biofilm formation, by making a larger surface area available for bacterial interactions and providing concave spaces where the microorganisms can find favorable and protective microenvironments (Anselme et al. 2010; Ionescu et al. 2012). Moreover, the presence of irregularities hampers the cleaning of the surface, hence favoring the accumulation of nutrient substrates and microbes (Teughels et al. 2006). The distribution of peaks and valleys, especially on a scale comparable with the microbial size, also affects the biofilm formation. Remarkably, bacterial cells seem able to discriminate the spatial scattering of microscopic depressions or elevations of a surface area (Perera-Costa et al. 2014). However, the roughness influences the biofilm-forming ability in a strain-dependent fashion (Mitik-Dineva et al. 2008). The bacterial attachment was shown to be more efficient on soft than on hard surfaces (Saha et al. 2013), thus highlighting the importance of surface stiffness both for the initial step and for maintaining the adhesive properties of the biofilm (Guegan et al. 2014).
Moreover, the type of material of which the surface is constituted (e.g., stainless steel, Teflon, plastic, ceramic, polystyrene, and metal) and the possible coating by organic molecules, which is frequent in food manufactory, define the chemical affinity to create biofilm (Cazzaniga et al. 2015; Renner and Weibel 2011). The surface charge determines the binding force through which the bacteria anchor to the material. Generally, positively charged surfaces promote interaction with bacterial membranes and formation of biofilm, although quaternary ammonium and polyethylenimines have the property to be antibiofilm functional groups (Campoccia et al. 2013). Bacteria tend to adhere to glass-forming monolayers, while easily form clumps during the adhesion on nylon and tin (Chmielewski and Frank 2003). Furthermore, the surface energy, which is directly correlated to the surface reactivity (Cazzaniga et al. 2015), and the hydrophobicity or hydrophilicity influence the bacteria-to-surface interaction, depending on bacterial strains (Zhang et al. 2013). Metallic materials provide high-energy, negatively charged, hydrophilic surfaces, whereas Teflon provides low-energy, poor negatively charged hydrophobic surfaces (Faille et al. 2002). Thus, stainless steel and glass which offer high free surface energy are relatively hydrophilic and generally facilitate the biofilm formation of a great number of bacteria, with respect to other hydrophobic surfaces such as Teflon, nylon, buna-N rubber, and fluorinated polymers (Chmielewski and Frank, 2003).
Conclusion and prospectives
LAB communities are widespread on both biotic and abiotic surfaces, and their occurrence may be either beneficial or detrimental to humans. Colonization of GIT and vaginal mucosa by probiotic LAB can serve as immune barrier against potential pathogens and provide other health benefits related to their probiotic activities. On the other hand, streptococci and lactobacilli included in the microbial biofilms of the oral cavity are associated with dental caries lesions. Likewise, contamination of food matrices by LAB biofilm communities is often functional to the manufacture process and to achieve the distinctive features of the product. Yet, colonization by undesirable LAB can also deteriorate food and food equipments. A deep knowledge of the genetic basis, the environmental factors, and the cellular mechanisms allowing LAB colonization of biotic and abiotic surfaces is essential for any strategy aiming to either prevent or favor the formation of such structured communities.
Controlling the production of LAB biofilms holds fascinating applications in both health and food biotechnologies. Such potentials are mainly related to their protective effects and competitive behavior towards other microbes. For instance, LAB colonization of food-related surfaces can be exploited to counteract pathogens and spoilage microorganisms, notably reducing food poisoning, product deterioration, and the use of chemical agents. EPS and/or biofilm-producing strains have been proposed also for food packaging as biodegradable polymers or food coating, thereby reducing food browning and dehydration. Moreover, the piloted immobilization of LAB on specific surfaces could be advantageous for the in situ production of several bioactive compounds, including antimicrobials, food-preserving agents, and therapeutics. Similarly, through the colonization of the host mucosae, LAB represent biotic alternatives or additive treatments to the use of antibiotics and drugs in human and animal medical practice.
References
Abachin E, Poyart C, Pellegrini E, Milohanic E, Fiedler F, Berche P, Trieu-Cuot P (2002) Formation of D-alanyl-lipoteichoic acid is required for adhesion and virulence of Listeria monocytogenes. Mol Microbiol 43:1–14
Adlerberth I, Ahrne S, Johansson ML, Molin G, Hanson LA, Wold AE (1996) A mannose-specific adherence mechanism in Lactobacillus plantarum conferring binding to the human colonic cell line HT-29. Appl Environ Microbiol 62:2244–2251
Alcántara C, Zúñiga M (2012) Proteomic and transcriptomic analysis of the response to bile stress of Lactobacillus casei BL23. Microbiol 158:1206–1218
Altermann E, Russell WM, Azcarate-Peril MA, Barrangou R, Buck BL, McAuliffe O, Souther N, Dobson A, Duong T, Callanan M, Lick S, Hamrick A, Cano R, Klaenhammer TR (2005) Complete genome sequence of the probiotic lactic acid bacterium Lactobacillus acidophilus NCFM. PNAS 102:3906–3912
Àlvarez G, González M, Isabal S, Blanc V, León R (2013) Method to quantify live and dead cells in multi-species oral biofilm by real-time PCR with propidium monoazide. AMB Express 3:1
Anselme K, Davidson P, Popa A, Giazzon M, Liley M, Ploux L (2010) The interaction of cells and bacteria with surfaces structured at the nanometer scale. Acta Biomater 6:3824–3846
Antikainen J, Kuparinen V, Lähteenmäki K, Korhonen TK (2007) pH dependent association of enolase and glyceraldehyde-3-phosphate dehydrogenase of Lactobacillus crispatus with the cell wall and lipoteichoic acids. J Bacteriol 189:4539–4543
Aoudia N, Rieu A, Briandet R, Deschamps J, Chluba J, Jego G, Garrido C, Guzzo C (2016) Biofilms of Lactobacillus plantarum and Lactobacillus fermentum: effect on stress responses, antagonistic effects on pathogen growth and immunomodulatory properties. Food Microbiol 53:51–59
Arena MP, Russo P, Capozzi V, López P, Fiocco D, Spano G (2014) Probiotic abilities of riboflavin-overproducing Lactobacillus strains: a novel promising application of probiotics. Appl Microbiol Biotechnol 98:7569–7581
Arena MP, Caggianiello G, Russo P, Albenzio M, Massa S, Fiocco D, Capozzi V, Spano G (2015) Functional starters for functional yogurt. Foods 4:15–33
Arroyo-Lopez FN, Bautista-Gallego J, Dominguez-Manzano J, Romero-Gil V, Rodriguez-Gómez F, García-García P, Garrido-Fernández A, Jiménez-Díaz R (2012) Formation of lactic acid bacteria-yeasts communities on the olive surface during Spanish-style Manzanilla fermentations. Food Microbiol 32:295–301
Avall-Jääskeläinen S, Lindholm A, Palva A (2003) Surface display of the receptor-binding region of the Lactobacillus brevis S-layer protein in Lactococcus lactis provides nonadhesive lactococci with the ability to adhere to intestinal epithelial cells. Appl Environ Microbiol 69:2230–2236
Azcarate-Peril MA, Altermann E, Goh YJ, Tallon R, Sanozky-Dawes RB, Pfeiler EA, O’Flaherty S, Logan Buck B, Dobson A, Duong T, Miller MJ, Barrangou R, Klaenhammer TR (2008) Analysis of the genome sequence of Lactobacillus gasseri ATCC 33323 reveals the molecular basis of an autochthonous intestinal organism. Appl Environ Microbiol 74:4610–46257
Banas JA, Vickerman MM (2003) Glucan-binding proteins of the oral streptococci. Crit Rev Oral Biol Med 14:89–99
Barocchi MA, Ries J, Zogaj X, Hemsley C, Albiger B, Kanth A, Dahlberg S, Fernebro J, Moschioni M, Masignani V, Hultenby K, Taddei AR, Beiter K, Wartha F, von Euler A, Covacci A, Holden DW, Normark S, Rappuoli R, Henriques-Normark B (2006) A pneumococcal pilus influences virulence and host inflammatory responses. Proc Natl Acad Sci U S A 103:2857–2862
Bassler BL, Losick R (2006) Bacterially speaking. Cell 125:237–246
Becerra JE, Yebra MJ, Monedero V (2015) An l-fucose operon in the probiotic Lactobacillus rhamnosus GG is involved in adaptation to gastrointestinal conditions. Appl Environ Microbiol 81:3880–3888
Begley M, Hill C, Gahan CGM (2006) Bile salt hydrolase activity in probiotics. Appl Environment Microbiol 72:1729–1738
Bergonzelli GE, Granato D, Pridmore RD, Marvin-Guy LF, Donnicola D, Corthésy-Theulaz IE (2006) GroEL of Lactobacillus johnsonii La1 (NCC 533) is cell surface associated: potential role in interactions with the host and the gastric pathogen Helicobacter pylori. Infect Immun 74:425–434
Bermúdez-Humarán LG, Aubry C, Motta JP, Deraison C, Steidler L, Vergnolle N, Chatel JM, Langella P (2013) Engineering lactococci and lactobacilli for human health. Curr Opin Microbiol 16:278–283
Boekhorst J, Helmer Q, Kleerebezem M, Siezen RJ (2006) Comparative analysis of proteins with a mucus-binding domain found exclusively in lactic acid bacteria. Microbiol 152:273–280
van Bokhorst-van de Veen H, Smelt MJ, Wels M, van Hijum SA, de Vos P, Kleerebezem M, Bron PA (2013) Genotypic adaptations associated with prolonged persistence of Lactobacillus plantarum in the murine digestive tract. Biotechnol J 8:895–904
Bove P, Gallone A, Russo P, Capozzi V, Albenzio M, Spano G, Fiocco D (2012) Probiotic features of Lactobacillus plantarum mutant strains. Appl Microbiol Biotechno l96:431–441
Bove P, Russo P, Capozzi V, Gallone A, Spano G, Fiocco D (2013) Lactobacillus plantarum passage through an oro-gastro-intestinal tract simulator: carrier matrix effect and transcriptional analysis of genes associated to stress and probiosis. Microbiol Res 168:351–359
Breshears LM, Edwards VL, Ravel J, Peterson ML (2015) Lactobacillus crispatus inhibits growth of Gardnerella vaginalis and Neisseria gonorrhoeae on a porcine vaginal mucosa model. BMC Microbiol 15:276–288
Bridier A, Briandet R, Thomas V, Dubois-Brissonnet F (2011) Resistance of bacterial biofilms to disinfectants: a review. Biofouling 27:1017–1032
Bron PA, Grangette C, Mercenier A, De Vos WM, Kleerebezem M (2004a) Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal tract of mice. J Bacteriol 186:5721–5729
Bron PA, Marco M, Hoffer SM, Van Mullekom E, de Vos WM, Kleerebezem M (2004b) Genetic characterization of the bile salt response in Lactobacillus plantarum and analysis of responsive promoters in vitro and in situ in the gastrointestinal tract. J Bacteriol 186:7829–7835
Bron PA, Molenaar D, de Vos WM, Kleerebezem M (2006) DNA micro-array-based identification of bile-responsive genes in Lactobacillus plantarum. J Appl Microbiol 100:728–738
Buck BL, Altermann E, Svingerud T, Klaenhammer TR (2005a) Functional analysis of putative adhesion factors in Lactobacillus acidophilus NCFM. Appl Environ Microbiol 71:8344–8351
Caggianiello G, Kleerebezem M, Spano G (2016) Exopolysaccharides produced by lactic acid bacteria: from health-promoting benefits to stress tolerance mechanisms. Appl Microbiol Biotechnol 100:3877–3886
Cai H, Thompson R, Budinich MF, Broadbent JR, Steele JL (2009) Genome sequence and comparative genome analysis of Lactobacillus casei: insights into their niche-associated evolution. Genome Biol Evol 1:239–257
Call EK, Klaenhammer TR (2013) Relevance and application of sortase and sortase-dependent proteins in lactic acid bacteria. Front Microbiol 4:73–83
Call EK, Goh YJ, Selle K, Klaenhammer TR, O’Flaherty S (2015) Sortase-deficient lactobacilli: effect on immunomodulation and gut retention. Microbiol 161:311–321
Campoccia D, Montanaro L, Arciola CR (2013) A review of the biomaterials technologies for infection-resistant surfaces. Biomaterials 34:8533–8554
Candela M, Centanni M, Fiori J, Biagi E, Turroni S, Orrico C, Bergmann S, Hammerschmidt S, Brigidi P (2010) DnaK from Bifidobacterium animalis subsp. lactis is a surface-exposed human plasminogen receptor upregulated in response to bile salts. Microbiol 156:1609–1618
Carr FJ, Chill D, Maida N (2002) The lactic acid bacteria: a literature survey. Crit Rev Microbiol 28:281–370
Castaldo C, Vastano V, Siciliano RA, Candela M, Vici M, Muscariello L, Marasco R, Sacco M (2009) Surface displaced alfa-enolase of Lactobacillus plantarum is a fibronectin binding protein. Microb Cell Factories 8:14–24
Cazzaniga G, Ottobelli M, Ionescu A, Garcia-Godoy F, Brambilla E (2015) Surface properties of resin-based composite materials and biofilm formation: A review of the current literature. Am J Dent 28
Ceapa C, Davids M, Ritari J, Lambert J, Wels M, Douillard FP, Smokvina T, de Vos WM, Knol J, Kleerebezem M (2016) The variable regions of Lactobacillus rhamnosus genomes reveal the dynamic evolution of metabolic and host-adaptation repertoires. Genome Biol Evol 8:1889–1905
Chapot-Chartier MP, Kulakauskas S (2014) Cell wall structure and function in lactic acid bacteria. Microb Cell Factories 13:S9
Chmielewski RAN, Frank JF (2003) Biofilm formation and control in food processing facilities. Compr Rev Food Sci Food Saf 2:22–32
Choudhary S, Schmidt-Dannert C (2010) Applications of quorum sensing in biotechnology. Appl Microbiol Biotechnol 86:1267–1279
Claes IJ, Lebeer S, Shen C, Verhoeven TL, Dilissen E, De Hertogh G, Bullens DM, Ceuppens JL, Van Assche G, Vermeire S, Rutgeerts P, Vanderleyden J, De Keersmaecker SCJ (2010) Impact of lipoteichoic acid modification on the performance of the probiotic Lactobacillus rhamnosus GG in experimental colitis. Clin Exp Immunol 162:306–314
Claes IJ, Segers ME, Verhoeven TL, Dusselier M, Sels BF, De Keersmaecker SC, Vanderleyden J, Lebeer S (2012) Lipoteichoic acid is an important microbe-associated molecular pattern of Lactobacillus rhamnosus GG. Microb Cell Factories 11:161
Claesson MJ, Li Y, Leahy S, Canchaya C, van Pijkeren JP, CerdeñoTárraga AM, Parkhill J, Flynn S, O’Sullivan GC, Collins JK, Higgins D, Shanahan F, Fitzgerald GF, van Sinderen D, O’Toole PW (2006) Multireplicon genome architecture of Lactobacillus salivarius. Proc Natl Acad Sci U S A 2006(103):6718–6723
Collins LV, Kristian SA, Weidenmaier C, Faigle M, van Kessel KPM, van Strijp JAG, Götz F, Neumeister B, Peschel A (2002) Staphylococcus aureus strains lacking D-alanine modifications of teichoic acids are highly susceptible to human neutrophil killing and are virulence attenuated in mice. J Infect Dis 186:214–219
Danne C, Dramsi S (2012) Pili of gram-positive bacteria: roles in host colonization. Res Microbiol 163:645–658
David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y, Fischbach MA, Biddinger SB, Dutton RJ, Turnbaugh PJ (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505:559–556
Denou E, Berger B, Barretto C, Panoff JM, Arigoni F, Brussow H (2007) Gene expression of commensal Lactobacillus johnsonii strain NCC533 during in vitro growth and in the murine gut. J Bacteriol 189:8109–8119
Denou E, Pridmore RD, Berger B, Panoff J-M, Arigoni F, Brüssow H (2008) Identification of genes associated with the long-gut persistence phenotype of the probiotic Lactobacillus johnsonii strain NCC533 using a combination of genomics and transcriptome analysis. J Bacteriol 190:3161–3168
Dertli E, Mayer MJ, Narbad A (2015) Impact of the exopolysaccharide layer on biofilms, adhesion and resistance to stress in Lactobacillus johnsonii FI9785. BMC Microbiol 15:8–12
Dhanani AS, Bagchi T (2013) The expression of adhesin EF-Tu in response to mucin and its role in Lactobacillus adhesion and competitive inhibition of enteropathogens to mucin. J Appl Microbiol 115:546–554
Di Cagno R, De Angelis M, Calasso M, Gobbetti M (2011) Proteomics of the bacterial cross-talk by quorum sensing. J Proteome 74:19–34
Didienne R, Defargues C, Callon C, Meylheuc T, Hulin S, Montel MC (2012) Characteristics of microbial biofilm on wooden vats (‘gerles’) in PDO Salers cheese. Int J Food Microbiol 156:91–101
Donian RM (2002) Biofilms: microbial life on surfaces. Emerg Infect Dis 8:881–890
Donlan RM (2001) Biofilms and device-associated infections. Emerg Infect Dis 7:277
Douillard FP, de Vos WM (2014) Functional genomics of lactic acid bacteria: from food to health. Microb Cell Factories 13:S8
Douillard FP, Ribbera A, Kant R, Pietilä TE, Järvinen HM, Messing M, Randazzo CL, Paulin L, Laine P, Ritari J, Caggia C, Lähteinen T, Brouns SJ, Satokari R, von Ossowski I, Reunanen J, Palva A, de Vos WM (2013) Comparative genomic and functional analysis of 100 Lactobacillus rhamnosus strains and their comparison with strain GG. PLoS Genet 9:e1003683
Dramsi S, Caliot E, Bonne I, Guadagnini S, Prevost MC, Kojadinovic M, Lalioui L, Poyart C, Trieu-Cuot P (2006) Assembly and role of pili in group B streptococci. Mol Microbiol 60:1401–1413
Elkins CA, Moser SA, Savage DC (2001) Genes encoding bile salt hydrolases and conjugated bile salt transporters in Lactobacillus johnsonii 100-100 and other Lactobacillus species. Microbiol 147:3403–3412
Fabretti F, Theilacker C, Baldassarri L, Kaczynski Z, Kropec A, Holst O, Huebner J (2006) Alanine esters of enterococcal lipoteichoic acid play a role in biofilm formation and resistance to antimicrobial peptides. Infect Immun 74:4164–4171
Faille C, Jullien C, Fontaine F, Bellon-Fontaine MN, Slomianny C, Benezech T (2002) Adhesion of Bacillus spores and Escherichia coli cells to inert surface: role of surface hydrophobicity. Can J Microbiol 48:728–738
Fernández de Palencia P, Werning ML, Sierra-Filardi E, Dueñas MT, Irastorza A, Corbí AL, López P (2009) Probiotic properties of the2-substituted (1,3)-beta-D-glucan-producing bacterium Pediococcus parvulus 2.6. Appl Environ Microbiol 75:4887–4891
Fernández Ramírez MD, Smid EJ, Abee T, Nierop Groot MN (2015) Characterisation of biofilms formed by Lactobacillus plantarum WCFS1 and food spoilage isolates. Int J Food Microbiol 207:23–29
Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8:623–633
Frese SA, Benson SK, Tannock GW, Loach DM, Kim J, Zhang M, Oh PL, Heng NCK, Patil PB, Juge N, MacKenzie DA, Pearson BM, Lapidus A, Dalin E, Tice H, Goltsman E, Land M, Hauser L, Ivanova N, Kyrpides NC, Walter J (2011) The evolution of host specialization of the vertebrate gut symbiont Lactobacillus reuteri. PLoS Genet 7:e1001314
Garai-Ibabe G, Dueñas MT, Irastorza A, Sierra-Filardi E, Werning ML, López P, Corbí AL, Fernández de Palencia P (2010) Naturally occurring 2-substituted (1,3)-beta-D-glucan producing Lactobacillus suebicus and Pediococcus parvulus strains with potential utility in the production of functional foods. Bioresour Technol 101:9254–9263
García-Ruiz A, González de Llano D, Esteban-Fernández A, Requena T, Bartolomé B, Moreno-Arribas MV (2014) Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol 44:220–225
Glenting J, Beck HC, Vrang A, Riemann H, Ravn P, Hansen AM, Antonsson M, Ahrné S, Israelsen H, Madsen S (2013) Anchorless surface associated glycolytic enzymes from Lactobacillus plantarum 299v bind to epithelial cells and extracellular matrix proteins. Microbiol Res 168:245–253
Goldin BR, Gorbach SL, Saxelin M, Barakat S, Gualtieri L, Salminen S (1992) Survival of Lactobacillus species (strain GG) in human gastrointestinal tract. Dig Dis Sci 37:121–128
Golomb BL, Hirao LA, Dandekar S, Marco ML (2016) Gene expression of Lactobacillus plantarum and the commensal microbiota in the ileum of healthy and early SIV-infected rhesus macaques. Scientific Reports 6:24–23
Granato D, Perotti F, Masserey I, Rouvet M, Golliard M, Servin A, Brassart D (1999) Cell surface-associated lipoteichoic acid acts as an adhesion factor for attachment of Lactobacillus johnsonii La1 to human enterocyte-like Caco-2 cells. Appl Environ Microbiol 65:1071–1077
Granato D, Bergonzelli GE, Pridmore RD, Marvin L, Rouvet M, Corthésy-Theulaz IE (2004) Cell surface-associated elongation factor tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect Immun 72:2160–2169
Grangette C, Nutten S, Palumbo E, Morath S, Hermann C, Dewulf J, Pot B, Hartung T, Hols P, Mercenier A (2005) Enhanced antiinflammatory capacity of a Lactobacillus plantarum mutant synthesizing modified teichoic acids. Proc Natl Acad Sci U S A 102:10321–10326
Gross G, van der Meulen J, Snel J, van der Meer R, Kleerebezem M, Niewold TA, Hulst MM, Smits MA (2008) Mannose-specific interaction of Lactobacillus plantarum with porcine jejunal epithelium. FEMS Immunol Med Microbiol 54:215–223
Gross G, Snel J, Boekhorst J, Smits MA, Kleerebezem M (2010) Biodiversity of mannose-specific adhesion in Lactobacillus plantarum revisited: strain-specific domain composition of the mannose-adhesin. Benef Microbes 1:61–66
Guegan C, Garderes J, Le Pennec G, Gaillard F, Fay F, Linossier I, Herry JM, Bellon Fontaine MN, Vallee Rehel K (2014) Alteration of bacterial adhesion induced by the substrate stiffness. Colloids Surf B Biointerfaces 114:193–200
Guiton PS, Hung CS, Kline KA, Roth R, Kau AL, Hayes E, Heuser J, Dodson KW, Caparon MJ, Hultgren SJ (2009) Contribution of autolysin and sortase a during Enterococcus faecalis DNA-dependent biofilm development. Infect Immun 77:3626–3638
Gunduz GT, Tuncel G (2006) Biofilm formation in an ice cream plant. Antonie Van Leeuwenhoek 89:329–336
Hahnel S, Wieser A, Lang R, Rosentritt M (2015) Biofilm formation on the surface of modern implant abutment materials. Clin Oral Implants Res 26:1297–1301
Hancock LE, Perego M (2004) The Enterococcus faecalis fsr two-component system controls biofilm development through production of gelatinase. J Bacteriol 186:5629–5639
Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, Morelli L, Berni Canani R, Flint HJ, Salminen S, Calder FC, Sanders ME (2014) Expert consensus document. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol 11:506–514
Hynönen U, Westerlund-Wikström B, Palva A, Korhonen TK (2002) Fibronectin-binding function in the SlpA surface protein of Lactobacillus brevis. J Bacteriol 184:3360–3367
Ionescu A, Wutscher E, Brambilla E, Schneider-Feyrer S, Giessibl FJ, Hahnel S (2012) Influence of surface properties of resin-based composites on in vitro Streptococcus mutans biofilm development. Eur J Oral Sci 120:458–465
Jensen H, Roos S, Jonsson H, Rud I, Grimmer S, van Pijkeren JP, Britton RA, Axelsson L (2014) Role of Lactobacillus reuteri cell and mucus-binding protein A (CmbA) in adhesion to intestinal epithelial cells and mucus in vitro. Microbiol 160:671–681
Jones BV, Begley M, Hill C, Gahan CGM, Marchesi JR (2008) Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc Natl Acad Sci U S A 105:13580–13585
Juge N (2012) Microbial adhesins to gastrointestinal mucus. Trends Microbiol 20:30–39
Kainulainen V, Loimaranta V, Pekkala A, Edelman S, Antikainen J, Kylväjä R, Laaksonen M, Laakkonen L, Finne J, Korhonen TK (2012) Glutamine synthetase and glucose-6-phosphate isomerase are adhesive moonlighting proteins of Lactobacillus crispatus released by epithelial cathelicidin LL-37. J Bacteriol 194:2509–2519
Kankainen M, Paulin L, Tynkkynen S, von Ossowski I, Reunanen J, Partanen P, Satokari R, Vesterlund S, Hendrickx APA, Lebeer S, De Keersmaecker SCJ, Vanderleyden J, Hamalainen T, Laukkanen S, Salovuori N, Ritari J, Alatalo E, Korpela R, Mattila-Sandholm T, Lassig A, Hatakka K, Kinnunen KT, Karjalainen H, Saxelin M, Laakso K, Surakka A, Palva A, Salusjarvi T, Auvinen P, de Vos WM (2009) Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc Natl Acad Sci U S A 106:17193–17198
Katakura Y, Sano R, Hashimoto T, Ninomiya K, Shioya S (2010) Lactic acid bacteria display on the cell surface cytosolic proteins that recognize yeast mannan. Appl Microbiol Biotechnol 86:319–326
Kindblom C, Davies JR, Herzberg MC, Svensäter G, Wickström C (2012) Salivary proteins promote proteolytic activity in Streptococcus mitis biovar 2 and Streptococcus mutans. Mol Oral Microbiol 27:362–372
Kinoshita H, Uchida H, Kawai Y, Kawasaki T, Wakahara N, Matsuo H, Watanabe M, Kitazawa H, Ohnuma S, Miura K, Horii A, Saito T (2008) Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J Appl Microbiol 104:1667–1674
Klaenhammer TR, Altermann E, Pfeiler E, Buck BL, Goh YJ, O'Flaherty S, Barrangou R, Duong T (2008) Functional genomics of probiotic lactobacilli. J Clin Gastroenterol 42:S160–S162
Kleerebezem M, Vaughan EE (2009) Probiotic and gut lactobacilli and bifidobacteria: molecular approaches to study diversity and activity. Annu Rev Microbiol 63:269–290
Klijn N, Weerkamp AH, de Vos WM (1995) Genetic marking of Lactococcus lactis shows its survival in the human gastrointestinal tract. Appl Environ Microbiol 61:2771–2774
Kniggendorf AK, Nogueira R, Kelb C, Schadzek P, Meinhardt-Wollweber M, Ngezahayo A, Roth B (2016) Confocal Raman microscopy and fluorescent in situ hybridization–a complementary approach for biofilm analysis. Chemosphere 161:112–118
Konstantinov SR, Smidt H, de Vos WM, Bruijns SC, Singh SK, Valence F, Molle D, Lortal S, Altermann E, Klaenhammer TR, van Kooyk Y (2008) S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc Natl Acad Sci U S A 105:19474–19479
Koskenniemi K, Laakso K, Koponen J, Kankainen M, Greco D, Auvinen P, Savijoki K, Nyman TA, Surakka A, Salusjärvi T, de Vos WM, Tynkkynen S, Kalkkinen N, Varmanen P (2011) Proteomics and transcriptomics characterization of bile stress response in probiotic Lactobacillus rhamnosus GG. Mol Cell Prot 10:M110–002741
Kristian SA, Datta V, Weidenmaier C, Kansal R, Fedtke I, Peschel A, Gallo RL, Nizetl V (2005) D-alanylation of teichoic acids promotes group a Streptococcus antimicrobial peptide resistance, neutrophil survival, and epithelial cell invasion. J Bacteriol 187:6719–6725
Kubota H, Senda S, Nomura N, Tokuda H, Uchiyama H (2008) Biofilm formation by lactic acid bacteria and resistance to environmental stress. J Biosci Bioeng 106:381–386
Kubota H, Senda S, Tokuda H, Uchiyama H, Nomura N (2009) Stress resistance of biofilm and planktonic Lactobacillus plantarum subsp. plantarum JCM 1149. Food Microbiol 26:592–597
Landeta G, Curiel JA, Carrascosa AV, Muñoz R, De Las RB (2013) Technological and safety properties of lactic acid bacteria isolated from Spanish dry-cured sausages. Meat Sci 95:272–280
Le DT, Tran TL, Duviau MP, Meyrand M, Guérardel Y, Castelain M, Loubière P, Chapot-Chartier MP, Dague E, Mercier-Bonin M (2013) Unraveling the role of surface mucus-binding protein and pili in muco-adhesion of Lactococcus lactis. PLoS One 8:e79850
Leathers TD, Bischoff KM (2011) Biofilm formation by strains of Leuconostoc citreum and L. mesenteroides. Biotechnol Lett 33:517–523
Lebeer S, De Keersmaecker SC, Verhoeven TL, Fadda AA, Marchal K, Vanderleyden J (2007a) Functional analysis of luxS in the probiotic strain Lactobacillus rhamnosus GG reveals a central metabolic role important for growth and biofilm formation. J Bacteriol 189:860–871
Lebeer S, Verhoeven TLA, Perea Velez M, Vanderleyden J, De Keersmaecker SCJ (2007b) Impact of enviromental and genetic factor son biofilms formation by the probiotic strain Lactobacillus rhamnosus GG. Appl Environ Microbiol 73:6768–6775
Lebeer S, Verhoeven TL, Francius G, Schoofs G, Lambrichts I, Dufrêne Y, Vanderleyden J, De Keersmaecker SC (2009) Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional analysis of the priming glycosyltransferase. Appl Environ Microbiol 75:3554–3563
Lebeer S, Claes IJ, Verhoeven TL, Vanderleyden J, De Keersmaecker SC (2011) Exopolysaccharides of Lactobacillus rhamnosus GG form a protective shield against innate immune factors in the intestine. Microb Biotechnol 4:368–374
Lebeer S, Claes IJ, Tytgat HLP, Verhoeven TLA, Marien E, von Ossowski I, Reunanen J, Palva A, de Vos WM, De Keersmaecker SCI, Vanderleyden J (2012) Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl Environ Microbiol 78:185–193
Lee IC, Caggianiello G, van Swam II, Taverne N, Meijerink M, Bron PA, Spano G, Kleerebezem M (2016) Strain-specific features of extracellular polysaccharides and their impact on host interactions of Lactobacillus plantarum. Appl Environ Microbiol AEM-00306
Licitra G, Ogier JC, Parayre S, Pediliggieri C, Carnemolla TM, Falentin H, Madec MN, Carpino S, Lortal S (2007) Variability of bacterial biofilms of the “tina” wood vats used in the ragusano cheese-making process. Appl Environ Microbiol 73:6980–6987
Lonvaud-Funel A (2016) Undesirable compounds and spoilage microorganisms in wine. In: Wine Safety, Consumer Preference, and Human Health Springer International Publishing, pp. 3–26
Macías-Rodríguez ME, Zagorec M, Ascencio F, Vázquez-Juárez R, Rojas M (2009) Lactobacillus fermentum BCS87 expresses mucus- and mucin-binding proteins on the cell surface. J Appl Microbiol 107:1866–1874
MacKenzie DA, Tailford LE, Hemmings AM, Juge N (2009) Crystal structure of a mucus-binding protein repeat reveals an unexpected functional immunoglobulin binding activity. J Biol Chem 284:32444–32453
Mackenzie DA, Jeffers F, Parker ML, Vibert-Vallet A, Bongaerts RJ, Roos S, Walter J, Juge N (2010) Strain-specific diversity of mucus-binding proteins in the adhesion and aggregation properties of Lactobacillus reuteri. Microbiol 156:3368–3378
Malik S, Petrova MI, Claes IJ, Verhoeven TLA, Busschaert P, Vaneechoutte M, Lievens B, Lambrichts I, Siezen RJ, Balzarini J, Vanderleyden J, Lebeer S (2013) The high auto-aggregative and adhesive phenotype of the vaginal Lactobacillus plantarum strain CMPG5300 is sortase-dependent. Appl Environ Microbiol 79:4576–4585
Mandlik A, Swierczynski A, Das A, Ton-That H (2008) Pili in gram-positive bacteria: assembly, involvement in colonization and biofilm development. Trends Microbiol 16:33–40
Marco ML, Bongers RS, de Vos WM, Kleerebezem M (2007) Spatial and temporal expression of Lactobacillus plantarum genes in the gastrointestinal tracts of mice. Appl Environ Microb 73:24–132
Marco ML, Peters TH, Bongers RS, Molenaar D, van Hemert S, Sonnenburg JL, Gordon JI, Kleerebezem M (2009) Lifestyle of Lactobacillus plantarum in the mouse caecum. Environ Microbiol 11:2747–2757
Marco ML, de Vries MC, Wels M, Molenaar D, Mangell P, Ahrne S, de Vos WM, Vaughan EE, Kleerebezem M (2010) Convergence in probiotic Lactobacillus gut-adaptive responses in humans and mice. ISME J 4:1481–1484
Mariani C, Briandet R, Chamba JF, Notz E, Carnet-Pantiez A, Eyoug RN, Oulahal N (2007) Biofilm ecology of wooden shelves used in ripening the French raw milk smear cheese reblochon de Savoie. J Dairy Sci 90:1653–1661
Matsuguchi T, Takagi A, Matsuzaki T, Nagaoka M, Ishikawa K, Yokokura T, Yoshikai Y (2003) Lipoteichoic acids from Lactobacillus strains elicit strong tumor necrosis factor alpha-inducing activities in macrophages through toll-like receptor 2. Clin Diagn Lab Immunol 10:259–266
Meijerink M, van Hemert S, Taverne N, Wels M, de Vos P, Bron PA, Savelkoul HF, van Bilsen J, Kleerebezem M, Wells JM (2010) Identification of genetic loci in Lactobacillus plantarum that modulate the immune response of dendritic cells using comparative genome hybridization. PLoS One 5:e10632
Mercier C, Durrieu C, Briandet R, Domakova E, Tremblay J, Buist G, Kulakauskas S (2002) Positive role of peptidoglycan breaks in lactococcal biofilm formation. Mol Microbiol 46:235–243
Meyrand M, Guillot A, Goin M, Furlan S, Armalyte J, Kulakauskas S, Cortes-Perez NG, Thomas G, Chat S, Péchoux C, Dupres V, Hols P, Dufrêne YF, Trugnan G, Chapot-Chartier MP (2013) Surface proteome analysis of a natural isolate of Lactococcus lactis reveals the presence of pili able to bind human intestinal epithelial cells. Mol Cell Proteomics 12:3935–3947
Mitik-Dineva N, Wang J, Mocanascu CR, Stoddart PR, Crawford RJ, Ivanova EP (2008) Impact of nano-topography on bacterial attachment. Biotechnol J 3:536–544
Mohamadzadeh M, Pfeiler EA, Brown JB, Zadeh M, Gramarossa M, Managlia E, Bere P, Sarraj B, Khan MW, Pakanati KC, Ansaria MJ, O'Flahertyc S, Barretta T, Klaenhammer TR (2011) Regulation of induced colonic inflammation by Lactobacillus acidophilus deficient in lipoteichoic acid. Proc Natl Acad Sci U S A 108:4623–4630
Monsan P, Bozonnet S, Albenne C, Joucla G, Willemot RM, Remaud-Simeon M (2001) Homopolysaccharides from lactic acid bacteria. Int Dairy J 11:675–685
Mukai T, Asasaka T, Sato E, Mori K, Matsumoto M, Ohori H (2002) Inhibition of binding of Helicobacter pylori to the glycolipid receptors by probiotic Lactobacillus reuteri. FEMS Immunol Med Microbiol 32:105–110
Murofushi Y, Villena J, Morie K, Kanmani P, Tohno M, Shimazu T, Aso H, Suda Y, Hashiguchi K, Saito T, Kitazawa H (2015) The toll-like receptor family protein RP105/MD1 complex is involved in the immunoregulatory effect of exopolysaccharides from Lactobacillus plantarum N14. Mol Immunol 64:63–75
Myszka K, Czaczyk K (2009) Characterization of adhesive exopolysaccharide (EPS) produced by Pseudomonas aeruginosa under starvation conditions. Curr Microbiol 58:541–546
Nallapareddy SR, Sillanpaa J, Mitchell J, Singh KV, Chowdhury SA, Weinstock GM, Sullam PM, Murray BE (2011a) Conservation of Ebp-type pilus genes among enterococci and demonstration of their role in adherence of Enterococcus faecalis to human platelets. Infect Immun 79:2911–2920
Nallapareddy SR, Singh KV, Sillanpaa J, Zhao M, Murray BE (2011b) Relative contributions of Ebp pili and the collagen adhesin ace to host extracellular matrix protein adherence and experimental urinary tract infection by Enterococcus faecalis OG1RF. Infect Immun 79:2901–2910
Nguyen TL, Vieira-Silva S, Liston A, Raes J (2015) How informative is the mouse for human gut microbiota research? Dis Model Mech 8:1–16
Nishiyama K, Ochiai A, Tsubokawa D, Ishihara K, Yamamoto Y, Mukai T (2013) Identification and characterization of sulfated carbohydrate-binding protein from Lactobacillus reuteri. PLoS One 8:e83703
Nishiyama K, Ueno S, Sugiyama M, Yamamoto Y, Mukai T (2016) Lactobacillus rhamnosus GG SpaC pilin subunit binds to the carbohydrate moieties of intestinal glycoconjugates. Anim Sci J 87:809–815
Notararigo S, de las Casas-Engel M, de Palencia PF, Corbí AL, López P (2014) Immunomodulation of human macrophages and myeloid cells by 2-substituted (1–3)-β-d-glucan from P. parvulus 2.6. Carbohydr Polym 112:109–113
O’Hara AM, Shanahan F (2006) The gut flora as a forgotten organ. EMBO Rep 7:688–693
Ouwehand AC, Salminen S, Isolauri E (2002) Probiotics an overview of beneficial effects. Antonie Van Leeuwenhoek 83:279–289
Oxaran V, Ledue-Clier F, Dieye Y, Herry JM, Péchoux C, Meylheuc T, Briandet R, Juillard V, Piard JC (2012) Pilus biogenesis in Lactococcus lactis: molecular characterization and role in aggregation and biofilm formation. PLoS One 7:e50989
Parisotto TM, Steiner-Oliveira C, Duque C, Peres RC, Rodrigues LK, Nobre-dos-Santos M (2010) Relationship among microbiological composition and presence of dental plaque, sugar exposure, social factors and different stages of early childhood caries. Arch Oral Biol 55:365–373
Park MR, Yun HS, Son SJ, Oh S, Kim Y (2014) Short communication: development of a direct in vivo screening model to identify potential probiotic bacteria using Caenorhabditis elegans. J Dairy Sci 97:6828–6834
Patel DK, Shah KR, Pappachan A, Gupta S, Singh DD (2016) Cloning, expression and characterization of a mucin-binding GAPDH from Lactobacillus acidophilus. Int J Biol Macromol 91:338–346
Perea Velez M, Verhoeven TL, Draing C, Von Aulock S, Pfitzenmaier M, Geyer A, Lambrichts I, Grangette C, Pot B, Vanderleyden J, De Keersmaecker SC (2007) Functional analysis of D-alanylation of lipoteichoic acid in the probiotic strain Lactobacillus rhamnosus GG. Appl Environ Microbiol 73:3595–3604
Perera-Costa D, Bruque JM, González-Martín ML, Gómez-García AC, Vadillo-Rodríguez V (2014) Studying the influence of surface topography on bacterial adhesion using spatially organized microtopographic surface patterns. Langmuir 30:4633–4641
Pfeiler EA, Klaenhammer TR (2009) Role of transporter proteins in bile tolerance of Lactobacillus acidophilus. Appl Environ Microbiol 75:6013–6016
Pfeiler EA, Azcarate-Peril MA, Klaenhammer TR (2007) Characterization of a novel bile-inducible operon encoding a two-component regulatory system in Lactobacillus acidophilus. J Bacteriol 189:4624–4634
Piard JC, Briandet R (2015) Lactic acid bacteria biofilms: from their formation to their health and biotechnological potential. In: Mozzi F, Raya RR, Vignolo GM (eds) Biotechnology of lactic acid bacteria: novel applications. Wiley-Blackwell, Hoboken, pp 341–361
Poyart C, Pellegrini E, Marceau M, Baptista M, Jaubert F, Lamy M-C, Trieu-Cuot P (2003) Attenuated virulence of Streptococcus agalactiae deficient in D-alanyl lipoteichoic acids is due to an increased susceptibility to defensins and phagocytic cells. Mol Microbiol 49:1615–1625
Pretzer G, Snel J, Molenaar D, Wiersma A, Bron PA, Lambert J, de Vos WM, van der Meer R, Smits MA, Kleerebezem M (2005) Biodiversity based identification and functional characterization of the mannose-specific adhesin of Lactobacillus plantarum. J Bacteriol 187:6128–6136
Pridmore RD, Berger B, Desiere F, Vilanova D, Barretto C, Pittet AC, Zwahlen MC, Rouvet M, Altermann E, Barrangou R, Mollet B, Mercenier A, Klaenhammer T, Arigoni F, Schell MA (2004) The genome sequence of the probiotic intestinal bacterium Lactobacillus johnsonii NCC 533. Proc Natl Acad Sci U S A 101:2512–2517
Raftis EJ, Salvetti E, Torriani S, Felis GE, O’Toole PW (2011) Genomic diversity of Lactobacillus salivarius. Appl Environ Microbiol 77:954–965
Ramiah K, van Reenen CA, Dicks LM (2008) Surface-bound proteins of Lactobacillus plantarum 423 that contribute to adhesion of Caco-2 cells and their role in competitive exclusion and displacement of Clostridium sporogenes and Enterococcus faecalis. Res Microbiol 159:470–475
Remus DM, Bongers RS, Meijerink M, Fusetti F, Poolman B, de Vos P, Wells JM, Kleerebezem M, Bron PA (2013) Impact of Lactobacillus plantarum sortase on target protein sorting, gastrointestinal persistence, and host immune response modulation. J Bacteriol 195:502–509
Renner LD, Weibel DB (2011) Physicochemical regulation of biofilm formation. MRS Bull 36:347–355
Rieu A, Aoudia N, Jego G, Chluba J, Yousfi N, Briandet R, Deschamps J, Gasquet B, Monedero V, Garrido C, Guzzo J (2014) The biofilm mode of life boosts the anti-inflammatory properties of lactobacillus. Cell Microbiol 16:1836–1853
Rintahaka J, Yu X, Kant R, Palva A, von Ossowski I (2014) Phenotypical analysis of the Lactobacillus rhamnosus GG fimbrial spaFED operon: surface expression and functional characterization of recombinant SpaFED pili in Lactococcus lactis. PLoS One 9:e113922
Roos S, Jonsson H (2002) A high-molecular-mass cell-surface protein from Lactobacillus reuteri 1063 adheres to mucus components. Microbiol 148:433–442
Roos S, Aleljung P, Robert N, Lee B, Wadström T, Lindberg M, Jonsson H (1996) A collagen binding protein from Lactobacillus reuteri is part of an ABC transporter system? FEMS Microbiol Lett 144:33–38
Ruas-Madiedo P, Gueimonde M, Margolles A, de los Losreyes-Gavilán CG, Salminen S (2006) Exopolysaccharides produced by probiotic strains modify the adhesion of probiotics and enteropathogens to human intestinal mucus. J Food Prot 69:2011–2015
Russo P, López P, Capozzi V, Fernández de Palencia P, Dueñas MT, Spano G, Fiocco D (2012) Beta-glucans improve growth, viability and colonization of probiotic microorganisms. Int J Mol Sci 13:6026–6039
Russo P, Iturria I, Mohedano ML, Caggianiello G, Rainieri S, Fiocco D, Angel Pardo M, López P, Spano G (2015) Zebrafish gut colonization by mCherry-labelled lactic acid bacteria. Appl Microbiol Biotechnol 99:3479–3490
Saad N, Urdaci M, Vignoles C, Chaignepain S, Tallon R, Schmitter JM, Bressollier P (2009) Lactobacillus plantarum 299v surface-bound GAPDH: a new insight into enzyme cell walls location. J Microbiol Biotechnol 19:1635–1643
Saha N, Monge C, Dulong V, Picart C, Glinel K (2013) Influence of polyelectrolyte film stiffness on bacterial growth. Biomacromol 14:520–528
Sára M, Sleytr UB (2000) S-layer proteins. J Bacteriol 182:859–868
Savijoki K, Lietzen N, Kankainen M, Alatossava T, Koskenniemi K, Varmanen P, Nyman TA (2011) Comparative proteome cataloging of Lactobacillus rhamnosus strains GG and Lc705. J Proteome Res 10:3460–3473
Schwab C, Tveit AT, Schleper C, Urich T (2014) Gene expression of lactobacilli in murine forestomach biofilms. Microbial Biotechnol 7:347–359
Sepp E, Mikelsaar M, Salminen S (1993) Effect of administration of Lactobacillus casei strain GG on the gastrointestinal microbiota of newborns. Microb Ecol Health Dis 6:309–314
Shi X, Zhu X (2009) Biofilm formation and food safety in food industries. Trends Food Sci Technol 20:407–413
Sillanpää J, Martínez B, Antikainen J, Toba T, Kalkkinen N, Tankka S, Lounatmaa K, Keränen J, Höök M, Westerlund-Wikström B, Pouwels PH, Korhonen TK (2000) Characterization of the collagen-binding S-layer protein CbsA of Lactobacillus crispatus. J Bacteriol 182:6440–6450
Sims IM, Frese SA, Walter J, Loach D, Wilson M, Appleyard K, Eason J, Livingston M, Baird M, Cook G, Tannock GW (2011) Structure and functions of exopolysaccharide produced by gut commensal Lactobacillus reuteri 100-23. ISME J 5:1115–1124
Smelt MJ, de Haan BJ, Bron PA, van Swam I, Meijerink M, Wells JM, Kleerebezem M, Faas MM, de Vos P (2013) The impact of Lactobacillus plantarum WCFS1 teichoic acid D-alanylation on the generation of effector and regulatory T-cells in healthy mice. PLoS One 30:e63099
Somers EB, Johnson ME, Wong AC (2001) Biofilm formation and contamination of cheese by nonstarter lactic acid bacteria in the dairy environment. J Dairy Sci 84:1926–1936
Spurbeck RR, Arvidson CG (2010) Lactobacillus jensenii surface-associated proteins inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect Immun 78:3103–3111
Stack HM, Kearney N, Stanton C, Fitzgerald GF, Ross RP (2010) Association of beta-glucan endogenous production with increased stress tolerance of intestinal lactobacilli. Appl Environ Microbiol 76:500–507
Stock AM, Robinson VL, Goudreau PN (2000) Two-component signal transduction. Annu Rev Biochem 69:183–215
Sturme MH, Nakayama J, Molenaar D, Murakami Y, Kunugi R, Fujii T, Vaughan EE, Kleerebezem M, de Vos WM (2005) An agr-like two-component regulatory system in Lactobacillus plantarum is involved in production of a novel cyclic peptide and regulation of adherence. J Bacteriol 187:5224–5235
Suzuki K, Asano S, Iijima K, Kitamoto K (2008) Sake and beer spoilage lactic acid bacteria—a review. J Inst Brew 114:209–223
Tang Y, Dong W, Wan K, Zhang L, Li C, Zhang L, Liu N (2015) Exopolysaccharide produced by Lactobacillus plantarum induces maturation of dendritic cells in BALB/c mice. PLoS One 10:e0143743
Teughels W, Van Assche N, Sliepen I, Quirynen M (2006) Effect of material characteristics and/or surface topography on biofilm development. Clin Oral Implants Res 17:68–81
Theilacker C, Kaczyński Z, Kropec A, Sava I, Ye L, Bychowska A, Holst O, Huebner J (2011) Serodiversity of opsonic antibodies against Enterococcus faecalis-glycans of the cell wall revisited. PLoS One 6:e17839
Tytgat HL, van Teijlingen NH, Sullan RM, Douillard FP, Rasinkangas P, Messing M, Reunanen J, Satokari R, Vanderleyden J, Dufrêne YF, Geijtenbeek TB, de Vos WM, Lebeer S (2016a) Probiotic gut microbiota isolate interacts with dendritic cells via glycosylated heterotrimeric pili. PLoS One 11:e0151824
Tytgat HL, Douillard FP, Reunanen J, Rasinkangas P, Hendrickx AP, Laine PK, Paulin L, Satokari R, de Vos WM (2016b) Lactobacillus rhamnosus GG outcompetes Enterococcus faecium via mucus-binding pili: evidence for a novel and heterospecific probiotic mechanism. Appl Environ Microbiol 82:5756–5762
Van Acker H, Van Dijck P, Coenye T (2014) Molecular mechanisms of antimicrobial tolerance and resistance in bacterial and fungal biofilms. Trends Microbiol 22:326–333
Van Bokhorst-van de Veen H, van Swam I, Wels M, Bron PA, Kleerebezem M (2012) Congruent strain specific intestinal persistence of Lactobacillus plantarum in an intestine-mimicking in vitro system and in human volunteers. PLoS One 7:e44588
van Hemert S, Meijerink M, Molenaar D, Bron PA, de Vos P, Kleerebezem M, Wells JM, Marco ML (2010) Identification of Lactobacillus plantarum genes modulating the cytokine response of human peripheral blood mononuclear cells. BMC Microbiol 10:293
van Pijkeren JP, Canchaya C, Ryan KA, Li Y, Claesson MJ, Sheil B, Steidler L, O'Mahony L, Fitzgerald GF, van Sinderen D, O'Toole PW (2006) Comparative and functional analysis of sortase-dependent proteins in the predicted secretome of Lactobacillus salivarius UCC118. Appl Environ Microbiol 76:4143–4153
Vargas García CE, Petrova M, Claes IJJ, De Boeck I, Verhoeven TLA, Dilissen E, von Ossowski I, Palva A, Bullens DM, Vanderleyden J, Lebeer S (2015) Piliation of Lactobacillus rhamnosus GG promotes adhesion, phagocytosis, and cytokine modulation in macrophages. Appl Environ Microbiol 81:2050–2062
Vélez MP, Petrova MI, Lebeer S, Verhoeven TL, Claes I, Lambrichts I, Tynkkynen S, Vanderleyden J, De Keersmaecker SC (2010) Characterization of MabA, a modulator of Lactobacillus rhamnosus GG adhesion and biofilm formation. FEMS Immunol Med Microbiol 59:386–398
von Ossowski I, Reunanen J, Satokari R, Vesterlund S, KankainenM M, Huhtinen H, Tynkkynen S, Salminen S, de Vos WM, Palva A (2010) Mucosal adhesion properties of the probiotic Lactobacillus rhamnosus GG SpaCBA and SpaFED pilin subunits. Appl Environ Microbiol 76:2049–2057
Von Ossowski I, Satokari R, Reunanen J, Lebeer S, de Keersmaecker SCJ, Vanderleyden J, de Vos WM, Palva A (2011) Functional characterization of a mucus-specific LPXTG surface adhesin from probiotic Lactobacillus rhamnosus GG. Appl Environ Microbiol 77:4465–4472
Walter J, Loach DM, Alqumber M, Rockel C, Hermann C, Pfitzenmaier M, Tannock GW (2007) D-alanyl ester depletion of teichoic acids in Lactobacillus reuteri 100-23 results in impaired colonization of the mouse gastrointestinal tract. Environ Microbiol 9:1750–1760
Walter J, Schwab C, Loach DM, Ganzle MG, Tannock GW (2008) Glucosyltransferase A (GtfA) and inulosucrase (Inu) of Lactobacillus reuteri TMW1.106 contribute to cell aggregation, in vitro biofilm formation, and colonization of the mouse gastrointestinal tract. Microbiol 154:72–80
Watanabe M, Kinoshita H, Nitta M, Yukishita R, Kawai Y, Kimura K, Taketomo N, Yamazaki Y, Tateno Y, Miura K, Horii A, Kitazawa H, Saito T (2010) Identification of a new adhesin-like protein from Lactobacillus mucosae ME-340 with specific affinity to the human blood group a and B antigens. J Appl Microbiol 109:927–935
Watnick P, Kolter R (2000) Biofilm, city of microbes. J Bacteriol 182:267–2679
Weiss G, Jespersen L (2012) Transcriptional analysis of genes associated with stress and adhesion in Lactobacillus acidophilus NCFM during the passage through an in vitro gastrointestinal tract model. J Mol Microbiol Biotechnol 18:206–214
Whitehead K, Versalovic J, Roos S, Britton RA (2008) Genomic and genetic characterization of the bile stress response of probiotic Lactobacillus reuteri ATCC 55730. Appl Environ Microbiol 74:1812–1819
Wickström C, Chávez de Paz L, Davies JR, Svensäter G (2013) Surface-associated MUC5B mucins promote protease activity in Lactobacillus fermentum biofilms. BMC Oral Health 13:43
Zhang X, Wang L, Levanen E (2013) Superhydrophobic surfaces for the reduction of bacterial adhesion. RSC Adv 3:12003–12020
Zhao T, Podtburg TC, Zhao P, Chen D, Baker DA, Cords B, Doyle MP (2013) Reduction by competitive bacteria of Listeria monocytogenes in biofilms and Listeria bacteria in floor drains in a ready-to-eat poultry processing plant. J Food Prot 76:601–607
Zhou M, Theunissen D, Wels M, Siezen RJ (2010) LAB-secretome: a genome-scale comparative analysis of the predicted extracellular and surface-associated proteins of lactic acid bacteria. BMC Genomics 11:651
Acknowledgements
This research was partialy supported by the Ministry of Education, University and Research (PON02_00186_2937475) in the framework of the project named Protocolli innovativi per lo sviluppo di alimenti funzionali^ (Pro. Ali. Fun.). Vittorio Capozzi was supported by a grant of the Apulian Region in the framework of “FutureInResearch” programme (practice code 9OJ4W81).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
This article does not contain any studies with human or animal subjects.
Conflict of interest
The authors declare that they have no competing interests.
Rights and permissions
About this article

Cite this article
Arena, M.P., Capozzi, V., Spano, G. et al. The potential of lactic acid bacteria to colonize biotic and abiotic surfaces and the investigation of their interactions and mechanisms. Appl Microbiol Biotechnol 101, 2641–2657 (2017). https://doi.org/10.1007/s00253-017-8182-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-017-8182-z