
Abstract
This study demonstrates volatile organic compounds (VOCs) production as one of the defense mechanisms of the antagonistic endophyte Nodulisporium sp. GS4d2II1a, and the volatile changes in two times of the fungal growth; and, as result of its intra and interspecific interactions with the plant pathogen Pythium aphanidermatum. The antifungal activity of the volatile and diffusible metabolites was evaluated by means of three types of antagonism bioassays and by organic extract agar dilution. VOCs were obtained by gas chromatography coupled to mass spectrometry from 3- and 5-day Nodulisporium sp. cultures, as well as from its interspecific in vitro antagonistic interaction with the oomycete P. aphanidermatum, and its intraspecific Nodulisporium sp.–Nodulisporium sp. interaction. The GS4d2II1a strain completely inhibited the growth of two fungi and seven oomycetes by replacing their mycelia in simple antagonism bioassays and by producing in vitro volatile and diffusible metabolites that acted synergistically in multiple antagonism bioassays. Additionally, VOCs inhibited the growth of three oomycetes and one fungus in antagonism bioassays using divided plates. A total of 70 VOCs were detected, mainly including mono and sesquiterpenes, especially eucalyptol and limonene. Multiple correspondence analysis revealed four different volatile profiles, showing that volatiles changed with the fungus age and its intra and interspecific interactions. The metabolites produced by Nodulisporium sp. GS4d2II1a could be useful for biological control of fungal and oomycetes plant pathogens of economically important crops.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many microorganisms generate a variety of volatile organic compounds (VOCs) with biological activity. Among them, fungi produce volatile compounds with antifungal activity that have potential applications in medicine, industry, and in agriculture [1]. Endophytic fungi–microorganisms that live inside plant tissues without causing disease symptoms or displaying outward signs of habitation are capable to produce secondary metabolites with antibiotic activity, such as, nonvolatile and volatile compounds [2, 3]. Fungal volatiles mediate ecological relationships such as intra and interspecific communication and as defense compounds [4, 5]. In addition, endophytic fungi could protect the host plant by producing nonvolatile and volatile compounds with antifungal activity against plant pathogens, and by mycoparasitism and predation that limits the pathogen growth. In addition, these mechanisms could be responsible for their survival inside the plant [6, 7]. Recently, the study of endophytic fungi has acquired great relevance as a promising approach for the detection of volatile compounds, which can be used as biological control or as mycofumigant agents in pest management for crop protection against fungal plant pathogens [1, 8, 9].
In vitro studies with interacting fungi show that they produce primary and secondary antifungal metabolites such as enzymes, diffusible compounds, and volatile compounds that inhibit the fungal competitor at distance [10]. The interaction zone of the fungal antagonism also involves dead hyphae by mycoparasitism, pigments, and morphology changes of the mycelia [6, 11]. The fungus VOC profiles in these antagonistic interactions, may change qualitatively, quantitatively or both during inter- and intraspecific interactions [12]. VOCs profiles also change in antagonistic interactions between fungi and bacteria [5, 13].
There are some reports about changes in volatile production in interspecific interactions between saprobe and wood rotting basidiomycetes [12, 14], and between bacteria and fungi [5], when exploring the ecological role of volatile compounds produced by fungi and their possible application in agriculture. However, up to date, there are no reports on changes in the production of volatile compounds between an endophytic fungus and an oomycete plant pathogen in antagonistic interactions, since the metabolites produced are probably responsible for the biological activity observed in co-culture; generally, this bioactivity is attributed to the endophyte secondary metabolites produced when is cultivated under axenic monoculture conditions.
The fungal genus Hypoxylon and its anamorph, Nodulisporium, are members of the Xylariaceae family (Phylum Ascomycota), and are producers of antimicrobial VOCs. The compounds produced by Nodulisporium spp. include alcohols, carboxylic acids, esters, and monoterpenes which have antifungal activity against plant pathogens [15–20].
Herein, we describe an endophytic fungus, Nodulisporium sp. strain GS4d2II1a, isolated from Gliricidia sepium (Fabaceae), which produces antifungal VOCs that change in antagonistic interaction. With this study, we attempt to demonstrate that VOCs production is one of the defense mechanisms of the antagonistic endophyte Nodulisporium sp. GS4d2II1a, and how these VOCs change in intra and interspecific interactions. The metabolites produced by the GS4d2II1a strain could be useful in the biological control of plant pathogens and therefore emerge as a possible alternative to fungicides with lower environmental impact.
Materials and Methods
Fungal Isolation
The endophytic fungus GS4d2II1a was isolated from healthy leaves of G. sepium (Jacq.) Kunth ex Walp. (1842) (Fabaceae) following the protocol reported by Rodrigues [21]. Leaves were collected in the Reserva de la Biósfera Sierra de Huautla (REBIOSH), located in the Rio Balsas basin, Quilamula (8° 30′ 4.1″ N–98° 51′ 52″ W and 18° 32′ 12.2″ N–99° 02′ 05″ W 1080-1230 masl), Morelos, México, in September 2010. Leaves were washed with both running and distilled water, and were surface sterilized with the following solutions: 75 % ethanol for 60 s, 3.4 % sodium hypochlorite (65 % Clorox®) for 60 s, and washed with sterile distilled water between solutions and at the end. The surface-sterilized leaves were dried with sterile adsorbent paper; cut into 2 × 2 mm pieces at the central vein level of each leaf. Four pieces were placed onto potato–dextrose–agar (PDA) and distilled water-agar (WA) plates. Chloramphenicol (500 mg/l) (Sigma-Aldrich, St. Louis, MO, USA) was added to both media to inhibit bacterial growth. Plates were incubated at 25 °C in photoperiod of 12:12 h light–dark with a fluorescent light. Plates were observed daily and emerging colonies were re-inoculated in new PDA plates until pure cultures were obtained.
The purified strain GS4d2II1a is preserved in PDA slants at the Instituto de Química, UNAM. They are also maintained in WA (0.2 %) at 4 °C and in 30 % glycerol–potato–dextrose broth (PDB) at −80 °C at the Laboratorio de Micología C006, Instituto de Biología, UNAM. Dried PDA and oatmeal agar (OA) cultures are deposited in the Herbario Nacional de México (MEXU), UNAM, with the collection number MEXU 27-541.
Morphological and Molecular Identification
Morphological observations and sequencing of the ITS1-5.8-ITS2 region were used for taxonomic identification. For examination of macro- and micromorphological characteristics, the strain GS4d2II1a was grown onto OA and PDA plates at 27 °C in darkness for 2 weeks. Cultures were observed daily to measure growth rate and for colony description according to the protocols of Ellis [22] and Ju and Rogers [23]. Anamorphic structures were observed, measured, and photographed with a brightfield microscope; colony and pigment colors were determined with the Methuen Handbook of Colour [24]. The strain was identified as Nodulisporium sp. by its macro- and microscopic features and following the keys and diagnosis of Ellis [22] and Ju and Rogers [23].
This identity was confirmed by sequence analysis of the ITS1-5.8S-ITS2 region amplified by polymerase chain reaction (PCR) using the forward primer ITS1F (5′-CTTGGTCATTTAAGAGGAAGTAA-3′) [25], and the reverse primer ITS4-A (5′-CGCCGTTACTGGGGCAATCCCTG-3′) [26]. For this purpose, total genomic DNA was extracted from a 5 days culture in PDB, incubated at 29 °C at 180 rpm, following the modified protocol by Akins and Lambowitz [27]. The PCR reaction was carried out in 50 μl reaction volume containing 5 μl 1× PCR Buffer 10× (Invitrogen); 1.25 mM of each dNTP (Invitrogen); 2 mM MgCl2, 25 pM each primer, 300 ng (150 ng/ml) genomic DNA, and 1 U Taq polymerase (Invitrogen). DNA amplification was performed in a Thermal Cycler PCR (GeneAmp PCR System 2400, Applied Biosystems, CA, USA) consisting of an initial denaturing step at 94 °C for 1 min, followed by 30 cycles of 1 min at 94 °C, 45 s at 52 °C; 1 min at 68 °C, and a final extension step for 10 min at 68 °C. PCR amplification product was separated by electrophoresis in 0.8 % (w/v) agarose gel (Invitrogen), stained with ethidium bromide (Sigma-Aldrich, Steinheim, Germany) in 1× Tris-Acetate-EDTA (TAE) buffer, at 90 V for 40 min; and, visualized under a UV transilluminator (UVP Dual-Intensity Transilluminator, Upland, CA, USA). The fragment size was estimated by comparison with a TrackIt 100 pb ladder (Invitrogen) [28]. The sequence was obtained from Laragen Inc. (UT, USA). Alignment and editing were done with BioEditProgram v 7.0.5. The edited sequence was aligned in the GenBank database using BLASTN program [29], and deposited. The GenBank accession number of the Nodulisporium sp. GS4s2II1a sequence is depicted in the “Results” section.
Simple Direct Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi
The fungus Nodulisporium sp. GS4d2II1a and the plant pathogenic microorganisms were paired opposite to each other in 9 cm Petri dishes with PDA. The endophyte inoculum was placed on the medium 3 days before the antagonism bioassay. The plant pathogens utilized were: the oomycetes Pythium aphanidermatum, Pythium ultimum, Pythium polytylum, Phytophthora capsici, Phytophthora palmivora, Phytophthora cinnamomi, and Phytophthora parasitica, in addition to the fungi Fusarium oxysporum and Alternaria solani [30]. The microorganisms were incubated at 28 °C in 12:12 h light–dark photoperiod and the outcome of the interactions was determined after 22 days. The main considered outcomes were: (1) inhibition at distance, where the competitor’s mycelia did not meet; (2) partial replacement, where the Nodulisporium strain grew over its competitor but the mycelium never reach the opposite side of the plate; (3) replacement, where the Nodulisporium strain completely grew over its competitor [11]. Percent of inhibition was determined at 4, 8, 11, and 22 days of incubation by measuring the radius of each mycelium near the interaction zone (defined as a), and compared to their respective controls that are within the radius of the mycelium, far from the interaction zone (defined as b) with the following equation:│(a − b / b)│ × 100. In order to confirm whether or not Nodulisporium sp. GS4d2II1a could kill the plant pathogens, the recovery of these pathogens was evaluated from a 22-day antagonistic interaction assay by transferring an inoculum (5 mm) of the interaction zone with Nodulisporium sp. GS4d2II1a to a new Petri dish with PDA, and incubating it under the same conditions. The growth of the plant pathogens, of Nodulisporium sp. GS4d2II1a, or both, was observed after 8 days incubation.
Multiple Direct Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi
Nodulisporium sp. GS4d2II1a is a fungus of rapid growth; therefore, to determine if this fungus can inhibit the growth of the plant pathogens at distance, possibly due to VOCs production, we performed multiple antagonism bioassays using 15 cm PDA plates with four 5-mm diameter inocula of Nodulisporium sp. GS4d2II1a. Initially, the Nodulisporium sp. GS4d2II1a inocula was placed at the center of the plate and was grown at 28 °C for 3 (Nod3), 5 (Nod5), 7, and during 9 days. At each incubation time, the oomycetes and fungi were inoculated around the Nodulisporium sp. GS4d2II1a colonies and at the edge of the plate. After inoculation, these plates were re-incubated under similar conditions. The plant pathogens tested were the same used in the simple direct antagonism assays. The outcome of the interactions and percentage of inhibition were determined at 2, 4, 6, and 8 days of antagonism. The percent of inhibition was calculated by measuring the radius of each mycelium and comparing this with their respective controls. Control cultures consisted of PDA plates inoculated with only the plant pathogens, under the same conditions.
Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi in Divided Petri Dishes
Bioassays were performed in 9-cm divided Petri plates with the two compartments containing 10 mL PDA. The plate dividing wall prevented the diffusion of the soluble compounds produced by Nodulisporium sp. GS4d2II1a, but did allow the free exchange of VOCs [31]. Antifungal activity of VOCs was tested against the plant pathogenic microorganisms P. aphanidermatum, P. capsici, P. cinnamomi, P. parasitica, F. oxysporum, and A. solani at three incubation times of Nodulisporium sp. GS4d2II1a growth. An inoculum of Nodulisporium sp. GS4d2II1a was placed in one of the compartments of the plate and incubated at 28 °C in 12:12 h light–dark photoperiod, for 3, 5, and 7 days before the bioassay. The inoculum from the plant pathogenic fungi and oomycetes was then placed in the other dish compartment. The plate was sealed with Parafilm® and incubated as above. The bioassays were made by triplicate. Control cultures were obtained by growing each plant pathogen alone, under the same conditions. Results of the bioassays were observed for 7 days. Percentages of growth inhibition were calculated after 2, 3, 4, and 7 days of antagonistic interaction by measuring the colony diameter of the each plant pathogenic fungi and oomycete and comparing with the respective controls [31].
Cultures of Nodulisporium sp. GS4d2II1a and Obtaining of the Secondary Metabolites
Cultures of Nodulisporium sp. GS4d2II1a were performed on 15-cm PDA plates containing 50 ml of culture medium. The inoculum consisted of a 5-mm diameter agar plug of the growing periphery of an 8-day culture of Nodulisporium sp. GS4d2II1a growing in PDA and then placed on the center of 10 PDA plates. Plates were incubated for three weeks at 28 °C in a 12:12 h light–dark photoperiod under fluorescent light. At the end of the incubation time, the mycelia were fragmented into small pieces and extracted with dichloromethane CH2Cl2 (500 ml × 3). The organic phase was filtered over anhydrous Na2SO4 and concentrated in vacuo. The extract obtained was a brown solid with a yield of 51.22 mg/50 ml of PDA.
Determination of Antifungal Activity of the Secondary Metabolites Against Plant Pathogenic Fungi and Fungoids
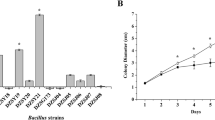
The antifungal bioassay with the dichloromethane extract was carried out with the plant pathogens P. ultimum, P. capsici, A. solani, and F. oxysporum. The extract was evaluated at 250 μg/ml on the radial growth of the microorganisms by dissolving the extract in 300 μl of methanol, and diluting this in PDA plates by quadruplicate. Plates were inoculated with a 5-mm diameter agar plug of each microorganism and incubated at 28 °C in 12:12 h light–dark photoperiod. The commercial fungicides Ridomil 4E® (Metalaxyl: 2-[(2,6-dimethyl-phenyl)-(2-methoxy-acetyl)-amino]-propionic acid methyl ester), and Prozicar® (carbendazim: methyl benzimidazol-2-yl-carbamate), for fungoids and fungi, respectively, were used as positive controls. Plates with PDA and PDA with 300 μl of methanol were used as negative controls. Percent of inhibition were calculated after 4 days incubation of the plant pathogens by measuring the mycelium diameter of each microorganism and comparing with the controls [32].
Qualitative Analysis of VOCs
The analysis of VOCs was carried out in solid phase micro extraction (SPME) vials with a polypropylene hole-cap and PTFE/coated silicone septa, containing 15 ml PDA. In order to tentatively identify the VOCs responsible of the biological activity, and to determine their changes when Nodulisporium sp. GS4d2II1a is interacting with a plant pathogen, we performed VOCs identification by triplicate under the following conditions: (1) A Nodulisporium sp. GS4d2II1a culture, grown for 3 days (Nod3); (2), another culture grown for 5 days (Nod5); (3) a P. aphanidermatum culture grown for 2 days. For intra- and interspecific interactions: (4) a 5-day antagonistic interaction of two Nodulisporium sp. GS4d2II1a inocula, placed 2 cm apart (Nod-Nod); and (5) a Nodulisporium sp. GS4d2II1a inoculum placed in a SPME vial and, after 3 days incubation, a P. aphanidermatum inoculum placed 2 cm apart (Nod-Pyt). The Nod-Pyt VOCs analysis was performed after 2 days of antagonistic interaction. The inoculum size was 5 mm diameter from 3 days culture of Nodulisporium sp. GS4d2II1a (Nod3), and 1-day culture of P. aphanidermatum.
VOCs in the headspace (HS) of the treatments were obtained by SPME. The SPME fiber (2 cm, 50/30 μm DVB/Carboxen/PDMS fiber, Supelco Bellefonte, PA, USA) was conditioned for 40 min at 250 °C, placed in the HS of the vials and exposed to the vapor phase during 60 min. The fiber was directly inserted into the Agilent 6890N series GC inlet (Agilent Technology, Palo Alto, CA, USA), at 300 °C, splitless mode. The desorption time was 2 min, and the desorbed compounds were separated on a 10 m × 0.18 mm × 0.18 μm, DB-5MS capillary column, using the following GC oven temperature program: 3 min at 40 °C up to 300 °C during 5 min, at 20 °C/min. Helium was used as the carrier gas at a flow rate of 1 ml/min. The GC was coupled to a LECO time of flight mass spectrometer (MS-TOF, Leco Corporation, St. Joseph, MI, USA) operating at unit resolution. The electronic ionization energy was 70 eV, and the mass range scanned was 33–500 uma. Scan rate was 20 spec/s. Transfer line and ionization chamber temperatures were 250 °C and 200 °C respectively. Data acquisition and processing were performed with LECO ChromaTOFTM software system. The volatile compounds were tentatively identified based on their MS fragmentation patterns and Kovats Index, calculated in relation to the retention times of a series of alkanes (C8–C20). Both were compared with chemical compounds data gathered by Adams [33] and the spectral database of the National Institute of Standards and Technology (NIST). Conclusive identification was made by comparing the MS and Kovats Index of compounds with available authentic standards purchased from Sigma-Aldrich (≥98 % purity). The authentic standards were: 3-methyl-1-butanol, 2-methyl-1-butanol, β-pinene, α-terpinene, eucalyptol, limonene, o-cymene, γ-terpinene and β-caryophyllene, as shown in Table 3. Since the objective was not the determination of absolute quantities of each compound, we determined the relative amounts of individual components of the treatments. The relative quantities were expressed as percentages of the peak area within the total peak area and as an average of the three replicates.
In order to evaluate the metabolites present in the culture medium, comparable analyses were performed with vials containing only PDA, and the identified compounds were removed from the data analyses.
Statistical Analysis
Experimental results regarding plant pathogens inhibition in the antagonism bioassays were analyzed by t-tests utilizing GraphPad Prism ver. 6.0 statistical computer software (GraphPad software, La Joya, CA, USA). Experimental results considering the inhibitory effect of the extract were analyzed by analysis of variance (ANOVA, α = 0.05), and Tukey statistical tests using the GraphPad Prism ver. 6.0 statistical software [31]. Data are represented as mean ± standard deviation (SD). A P value of <0.05 (*) was used to indicate statistical significance.
We visualized in a Venn diagram the distribution of the VOCs produced by the treatments [5, 34]. However, due to the complexity in the VOCs profiles induced by the experimental conditions, the results provided by the Venn diagram were difficult to describe. Therefore, we performed a multiple correspondence analysis (MCA) [35, 36] to separate and compare the volatile profiles produced between treatments. MCA was used to determine the treatment-specific profile of VOCs production according to their diversity of chemical families (i.e., monoterpenes, alkanes, ketones, etc.), and relative abundance (low, medium, or high). Additionally, we assessed the VOCs profile of Nodulisporium sp. GS4d2II1a and its possible changes associated to time and the kind of interaction. For the model, we included the treatment (Nod3, Nod5, Nod-Nod, and Nod-Pyt); the VOCs shared between two, three, or four treatments, altogether with the chemical family and the relative abundance of those compounds found exclusively on one treatment. For the relative content of VOCs, we used percentile distribution for each experimental condition, and separated the compounds into low (<33.3 %), medium (33.3–66.7 %), and high (>66.7 %) relative area percentage. MCA was performed using Statistical Analysis Software (SAS 9.0, SAS Institute, Cary, NC, USA), and graphs were constructed with Prism 6.0 (GraphPad software, La Joya, CA, USA).
Results
Fungal Identification
According to its macro- and micromorphological features, the strain GS4d2ll1a was identified as Nodulisporium sp. [22, 23].
Nodulisporium strain GS4d2ll1a presented the following cultural and morphological characteristics. The colony grown on PDA at 28 °C showed a fast growth and covered a 8.5 cm Petri dish in 1 week; with a growth rate of 10.42 cm2/day; it was at first whitish (M5A1), becoming buff brown (M6E3) with sporulation, velvety, reverse dark brown (M7F3). Sporulation was restricted to the center of the colony (Fig. 1a). Conidiophores were macronematous, momonematous, penicilliate branched toward the apex, hyaline to light brown, smooth to finely roughened, 1.5–2 mm; conidiogenous cells were hyaline to light brown (7.8–21 × 2.5–3.8 μm), and bore smooth, ellipsoidal or ovoid, light brown to brown one-celled conidia (5–7.5 × 2.7–2.7 μm) (Fig. 1b–e).

Nodulisporium sp. GS4d2II1a anamorph of H. anthochroum. a Colony on potato-dextrose–agar (PDA) after 7 days of growth at 28 °C. b, c Penicilliate branched conidiophores, smooth or slightly roughened; d conidiogenous cells bearing smooth, ellipsoidal or ovoid, light brown to brown conidia, stained with lactofuchsin, ×750; e conidiogenous cells with ovoid conidia, stained with lactofuchsin, ×950
The colony on OA incubated at 28 °C in darkness grew slower than in PDA, covering the Petri dish in 2 weeks. Mycelium was white, becoming brown (M6E5) with sporulation, reverse uncolored. Sporulation was present over the entire surface of the colony. Conidiogenous structure Nodulisporium-like had a central axis and additional branches, resulting in a complex structure; it was from hyaline to yellowish and from smooth to finely roughen. Conidiogenous cells and conidia were similar in shape and size to those described in PDA.
While morphological features depicted the anamorphic stage of the isolate as Nodulisporium sp., comparison of the ITS1-5.8-ITS2 sequence into the GenBank database gave the highest similarity (99 %) with the species Hypoxylon anthochroum (accession no. KF192825 and JN660819, with 95 % coverage), and Hypoxylon griseobrunneum (accession no. KC968928, with 96 % coverage), whose anamorphic state is Nodulisporium. GenBank accession number of the ITS1-5.8-ITS2 sequence of the GS4d2II1a strain is KM516709.
However, the macro and microscopic morphology, with its Nodulisporium-like branch pattern, supports the identification of the GS4d2II1a strain as Nodulisporium sp., anamorph of H. anthochroum, and not of H. griseobrunneum that presents a Vigariella-like conidogenous structure [23, 37].
Direct Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi
In the direct confrontation bioassay, the Nodulisporium GS42d2II1a strain first inhibited the growth of the plant pathogens with or without direct contact of their mycelia, followed by growth of its mycelia over the pathogens colony. After 4 days of antagonistic interaction, Nodulisporium sp. GS4d2II1a totally inhibited growth of P. capsici, P. cinnamomi, and P. parasitica, but did not inhibit P. aphanidermatum. Then, after 8 days interaction, Nodulisporium sp. GS4d2II1a grew over the pathogens colonies (partial replacement), and totally inhibited to P. ultimum, P. capsici, P. cinnamomi, P. parasitica, F. oxysporum and A. solani, while partially affecting the growth of P. aphanidermatum (15 %). Finally, after 11 days incubation, and up to 22 days of interaction, Nodulisporium sp. GS4d2II1a completely replaced six of the nine pathogens tested: P. aphanidermatum, P. capsici, P. palmivora, P. cinnamomi, P. parasitica, and A. solani (Table 1). Figure 2 shows Nodulisporium sp. GS4d2II1a colonies grown for 4 (Fig. 2a) and 8 (Fig. 2b) days; P. capsici colonies grown for 4 (Fig. 2c) and 10 (Fig. 2d) days; and P. aphanidermatum colony at 4 days (Fig. 2e) of growth. Figure 2 also shows the antagonistic interaction between Nodulisporium sp. GS4d2II1a with P. capsici at 4 (Fig. 2f) and 10 (Fig. 2g) days, and with P. aphanidermatum at 4 (Fig. 2h) and 21 days (Fig. 2i), where it is evident how Nodulisporium sp. GS4d2II1a overgrows on both pathogens.

Colonies of Nodulisporium sp. GS4d2II1a, P. capsici and P. aphanidermatum at different days of growth, and replacement of P. capsici and P. aphanidermatum by Nodulisporium sp. GS4d2II1a. Nodulisporium sp. GS4d2II1a at 4 (a) and 8 days (b) of growth. P. capsici at 4 (c) and 10 days (d) of growth. P. aphanidermatum at 4 days of growth (e). Nodulisporium sp. GS4d2II1a growing over P. capsici at 4 (f) and 10 days (g) of antagonistic interaction. Nodulisporium sp. GS4d2II1a growing over P. aphanidermatum at 4 (h) and 21 days (i) of antagonistic interaction
Additionally, Nodulisporium sp. GS4d2II1a killed the oomycetes P. aphanidermatum, P. capsici, P. cinnamomi, and P. parasitica, since they were not recovered; and, in the direct antagonism bioassays, the mycelium of those pathogens was replaced by the endophyte. In addition, although Nodulisporium sp. GS42d2ll1a partially (P. ultimum, P. polytylum, and F. oxysporum) or totally (P. palmivora and A. solani) replaced the mycelia of the pathogens, it did not kill them but inhibited their growth (Table 1).
Multiple Direct Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi
The antifungal activity of Nodulisporium sp. GS42d2ll1a was present after 3 and 5 days of growth, and reached its maximum inhibitory effect after 5 days of incubation; therefore, only the corresponding percent of inhibition at these two culture times is presented.
From 2 to 6 days of incubation in the antagonistic interaction, between Nod3 or Nod5 and the plant pathogens, the endophyte inhibited growth of the pathogens at distance. This was probably due to the VOCs with antifungal properties production. After 2 days of antagonistic interaction, between the Nod3 culture and the pathogens, Nod3 mainly reduced the growth of A. solani (25.4 %). After 4 days, it mainly inhibited to P. aphanidermatum (49.2 %) and F. oxysporum (32.0 %). After 6 days of antagonistic interaction, the highest suppression effect of Nod3 was observed on F. oxysporum (73.8 %) and on P. aphanidermatum (62.6 %); and, after 8 days A. solani (75.4 %), F. oxysporum (72.5 %), and P. aphanidermatum (72.6 %) were the most affected. However, at this time of interaction there were mycelial contact between these colonies and Nodulisporium sp. GS42d2ll1a (Table 2). Finally, Nodulisporium sp. GS42d2ll1a kept growing in all samples until the colony met the nine plant pathogen colonies and began to grow over them.
On the other hand, Nod5 inhibited at distance the competitors, but also limited its own growth. The reduction of the plant pathogens growth at distance, after 2 days of antagonistic interaction, was significant, mainly on P. aphanidermatum (58.8 %) (Table 2). This reduction continued during several days of antagonistic interaction. At day four, a growth reduction of 45.8, 41.0, and 78.1 % on A. solani, P. cinnamomi and P. aphanidermatum, respectively, was observed. After 6 days a growth inhibition of 67.0 % on F. oxysporum, and of 51.0 % on A. solani, was detected, and was conserved on P. aphanidermatum (78.5 %) (Fig. 3). And, after 8 days of antagonistic interaction, the inhibition was mainly due to contact of mycelia. Figure 3 shows the effect of Nod5 VOCs production on growth of the plant pathogens after 6 days of antagonistic interaction.

Multiple direct antagonism bioassay between Nodulisporium sp. GS4d2II1a and plant pathogens after 6 days of interaction. (1) P. cinnamomi, (2) P. polytylum, (3) P. capsici, (4) P. ultimum, (5) F. oxysporum, (6) P. palmivora, (7) A. solani, (8) P. aphanidermatum, (9) P. parasitica
Finally, when Nodulisporium sp. GS4d2II1a was grown for 7 and 9 days, its mycelium almost covered the plate surface and contacted the plant pathogens colonies and rapidly suppressed their growth (data not shown); therefore, these times did not allow observation of the distance inhibition.
Antagonism Bioassays with Plant Pathogenic Oomycetes and Fungi in Divided Petri Dishes
Bioactivity of VOCs was statistically significant (P < 0.05) after 3 days of Nodulisporium sp. GS42d2II1a growth (Nod3). It only suppressed P. parasitica (29.7 ± 3.7 %), P. cinnamomi (18.1 ± 4.3 %), P. capsici (25.7 ± 1.2 %), and F. oxysporum (21.6 ± 1.0 %) after 3 days of antagonism. VOCs were not active after 5 and 7 days of the fungal culture.
Determination of Antifungal Activity of the Secondary Metabolites from Nodulisporium sp. GS42d2II1a
The dichloromethane extract did not significantly inhibit the growth of the tested microorganisms.
Analysis of VOCs: Similarities and Diferences Among Treatments
Nodulisporium sp. GS4d2II1a produced a mixture of volatile secondary metabolites when cultured in vitro for 3 (Nod3), and 5 (Nod5) days, in interspecific interaction with P. aphanidermatum (Nod-Pyt), and in intraspecific interaction with other inoculum of Nodulisporium sp. GS4d2II1a (Nod-Nod). In total, 70 metabolites were detected in four out of the five treatments (P. aphanidermatum did not produce VOCs). The VOCs produced were mainly terpenes (mono and sesquiterpenes) that represent at least 63 % of the total. In particular, the monoterpenes were more abundant in Nod-Pyt, and the sesquiterpenes in Nod-Nod interactions. The VOCs produced in less proportion were alkenes, benzene derivatives, esters, alkanes, alkynes, amines, ketones, alcohols, and carboxylic acids (Fig. 4). Table 3 shows the total number of metabolites detected and listed by retention index (RI). Figure 5 lists the ten VOCs with highest fiber affinity, in each treatment, and their relative content. These metabolites represent 79.7 % in Nod3; 59.4 % in Nod5, 60.9 % in Nod-Nod; and, 72.5 % in Nod-Pyt of the total area, where eucalyptol was the compound with the highest affinity in Nod3 and Nod-Pyt, and limonene in Nod5 and Nod-Nod. Additionally, differences among the treatments were evident in the four representative chromatograms, where the peaks number and their areas changed (Fig. 6).

Molecule classification of the volatile metabolites produced by Nodulisporium sp. GS4d2II1a and relative content by treatment (%). Nodulisporium sp. GS4d2II1a at 3 days (Nod3), and 5 days (Nod5), in the intraspecific (Nod-Nod) and interspecific interactions with P. aphanidermatum (Nod-Pyt)

The ten VOCs with the highest fiber affinity (% Area) in the different treatments: Nodulisporium sp. GS4d2II1a at 3 days (Nod3), and 5 days (Nod5), in the intraspecific (Nod-Nod) and interspecific interactions with P. aphanidermatum (Nod-Pyt)

Representative chromatograms of the four treatments. a Nodulisporium sp. GS4d2II1a grown for 3 days in PDA at 28 °C (Nod3). b Nodulisporium sp. GS4d2II1a grown for 5 days in PDA at 28 °C (Nod5). c Interspecific interaction between Nodulisporium sp. GS4d2II1a and P. aphanidermatum (Nod-Pyt). d Intraspecific interaction (Nod-Nod)
Multiple Correspondence Analysis of VOCs Profiles
The Venn diagram (Fig. 7) revealed a complex pattern of volatile metabolites production between treatments (days of growth and type of interaction), since metabolites were produced exclusively in one treatment or in a mixture of treatments.

Venn diagram showing the volatile metabolites shared between treatments. Nodulisporium sp. GS4d2II1a at 3 days (Nod3), Nodulisporium sp. GS4d2II1a at 5 days (Nod5), intraspecific interaction (Nod-Nod), and interspecific interaction with P. aphanidermatum (Nod-Pyt)
Figure 8 shows the VOCs distribution by chemical family, and by relative content (% area: low, medium or high) that uncover the qualitative and quantitative chemical differences among treatments to provide a clear picture of the VOCs production, based on the volatile profile of each treatment. We observed a profile of VOCs production that varied according to the treatment. In particular, monoterpenes were produced in high proportion by Nod-Nod and Nod-Pyt treatments; on the other hand, the sesquiterpenes relative abundance was similar in all other treatments. Thus, the difference between treatments was in accordance to the larger or lesser production of the diverse chemical family groups, aside from mono and sesquiterpenes, found in different relative abundance (% area). For example, alkynes production by Nod5 that were absent in Nod3. Nod3 is characterized by high production of esters and other compounds families, and Nod5 by high production of esters and unknown compounds. Amines, monoterpenes, and sesquiterpenes production was constant between Nod3 and Nod5. The difference is given by the relative abundance of alkanes, alkenes, alcohols, benzene derivatives, alkynes (absent in Nod3); unknown metabolites, and other compounds. On the other hand, monoterpenes and alkenes production was constant in intra and interspecific interactions. The difference between these two treatments was the chemical families produced in different proportion: alkanes, alkynes, amines, alcohols and benzene derivatives, besides the absent ketones in Nod-Pyt.

Multiple correspondence analysis (MCA) of the association between diversity of the VOCs belonging to the principal chemical families and the relative content of these compounds for each experimental condition. aRelative content was based on the 33.3 and 66.7 % percentile distribution of the compounds for each treatment: low, < 33.3 %; medium, 33.3–66.7 %; and high, >66.7 % according to the total area. Nod3 (<0.38, 0.38–1.69 and >1.69), Nod5 (<0.75, 0.75–2.27, and >2.27), Nod-Pyt (<0.49, 0.49–1.87, and >1.87), and Nod-Nod (<0.82, 0.82–2.68 and >2.68). Arbitrary ellipses have been drawn to assist interpretation on the association of each relative content and the chemical families associated to these categories. Note the association between high relative content and monoterpenes in the antagonistic interactions Nod-Nod and Nod-Pyt and the constant production of sesquiterpenes in all treatments. Percentages (%) next to the chemical family symbol represent the relative amount of each chemical family
Figure 9 shows the MCA analysis based on the total volatile composition detailed in Table 3. It is evident the clear separation or metabolic trends between the four treatments, and the different chemical composition in each time and kind of interaction. Each ellipse encloses the metabolites that are exclusively produced by each treatment at the expense of the metabolites on the opposite extreme of the plot. Nod3 individual profile is closer to Nod-Pyt profile; thus, indicating they share common metabolic trends, and they are distant from Nod5 and Nod-Nod meaning that there are differences in the volatile production. Metabolites projected near the origin are produced by all treatments (18 compounds) and they do not characterize any volatile profile, they are constitutively produced. Metabolites outside ellipses are produced by two or three of the near treatments on the plot in different amounts, and are not representative of any treatment.

Multiple correspondence analysis (MCA) of the VOCs profile obtained during each experimental condition. Each number corresponds to the compounds presented in Table 3. aRelative content was based on the 33.3 and 66.7 % percentile distribution of the compounds for each treatment: low, < 33.3 %; medium, 33.3–66.7 %; and high, >66.7 % according to the total area. Arbitrary ellipses have been drawn to assist interpretation of the differential volatile profile of each treatment. Note the clear separation between time of growth and kind of antagonistic interaction
VOCs in Nod3 and Nod5 Treatments
Thirty four VOCs were detected in Nod3 samples, and these compounds were mainly terpenes—mono and sesquiterpenes—(Figs. 4 and 8). The compound with the highest fiber affinity was eucalyptol (33.22 %), followed by a caryophyllene-like compound (ID 55, 10.98 %), and m-cymene (9.77 %) (Fig. 5). In addition, 44 VOCs were detected in the Nod5 treatment (Table 3), and these VOCs were also mainly mono and sesquiterpenes (Figs. 4 and 8). The compounds with the highest fiber affinity were limonene (9.85 %), eucalyptol (6.92 %), and o-cymene (6.86 %) (Fig. 5). From the total VOCs produced, the two treatments shared 23; 5 were exclusively produced by Nodulisporium sp. GS4d2II1a grown for 3 days and 14 compounds, when grown during 5 days (Figs. 7 and 9).
VOCs in Intraspecific Antagonistic Interaction Nod-Nod
Nodulisporium sp. GS4d2II1a VOCs also changed in the antagonistic interaction with another inoculum of Nodulisporium sp. GS4d2II1a. Under this condition, 38 VOCs (mainly monoterpenes and sesquiterpenes) were produced, but 9 compounds were specific to this interaction (Figs. 4, 7 and 9). The compounds with the highest fiber affinity were limonene (13.16 %), eucalyptol (12.83 %), and γ-terpinene (7.70 %) (Fig. 5). On the other hand, from the total identified VOCs, 24 were also found in the 3-day Nodulisporium sp. GS4d2II1a growth; and, 24 compounds in the 5-day growth (Figs. 7 and 9).
VOCs in Interspecific Antagonistic Interaction Nod-Pyt
We analyzed the VOCs production in the interaction between Nodulisporium sp. GS4d2II1a and P. aphanidermatum, because P. aphanidermatum was the most affected microorganism in the multiple antagonism bioassays.
VOCs profile differed in the interspecific interaction and exhibited antifungal properties because they inhibited P. aphanidermatum growth completely in the SPME vial. In this antagonistic interaction, 37 compounds were identified mainly as mono and sesquiterpenes (Figs. 4 and 8). The compounds with the highest fiber affinity were eucalyptol (22.58 %), limonene (13.66 %), and o-cymene (10.77 %) (Fig. 5). From these VOCs, five, compounds were identified only in the antagonistic interspecific interaction (Figs. 7 and 9), and belonged to Nodulisporium sp. GS4d2II1a, since P. aphanidermatum did not produce volatile compounds and this has not been reported previously. The compounds produced exclusively from the interspecific interaction were 2-methyl-1-butanol, 4-carene, two unidentified monoterpenes (ID 37 and 42) and 3,5-dimethylanisole. In addition, from the total compounds produced, 25 were also found in VOCs identified during 3 days Nodulisporium sp. GS4d2II1a vial cultures, and 27 during 5 days vials cultures (Figs. 7 and 9). On the other hand, 24 out of the 37 detected compounds were also found in the antagonistic interaction between two Nodulisporium sp. inocula. From the total VOCs detected, only 18 were present in all treatments, but in different proportion. These metabolites were: thujene, 2-pentylfuran, α-phellandrene, α-terpinene, eucalyptol, limonene, γ-terpinene, α-terpinolen, an unidentified sesquiterpene (ID 47), β-caryophyllene, β-elemene, isocaryophyllene, α-humullene, 4-methylene-2,8,8-trimethyl-2-vinyl-bicyclo[5.2.0]nonane; 2-methylene-5-(1-methylvinyl)-8-methyl-bicyclo[5.3.0]decane, a caryophyllene like compound (ID 55), 2,4a,5,6,9a-hexahydro-3,5,5,9-tetramethyl(1H)benzocycloheptene and β-selinene (Figs. 7 and 9).
Discussion
Nodulisporium sp. GS4d2II1a inhibited the growth of oomycetes and fungi by replacing their mycelia, partially or completely, and killed four oomycetes. The mechanisms that probably mediate these antagonistic interactions are mycoparasitism, predation or hyphal interference [6, 7, 10]. Further studies should attempt to demonstrate which is the principal mechanism when these fungi and oomycetes mycelia interact.
On the other hand, after 2 to 6 days of antagonistic interaction between Nod3 or Nod5 and the plant pathogens in the multiple antagonism bioassay, Nodulisporium sp. GS4d2II1a suppressed the pathogens at distance, which indicated the production of VOCs with antifungal properties as an antagonistic mechanism. Therefore, we made antagonism bioassays in divided plates to verify if this was the only mechanism involved, since the production of antifungal diffusible metabolites could be present. Additionally, Nod5 also was self-inhibited probably due to the high VOCs concentration in the plates; or, also due to production of defense metabolites by the plant pathogens (dual antagonism) [10].
Bioactive VOCs production was demonstrated in divided Petri dish bioassays; however, this inhibition was lower and on fewer microorganisms than that observed in the simple and multiple direct antagonism bioassays. Moreover, the dichloromethane extract from Nodulisporium sp. GS42d2II1a culture did not inhibit plant pathogens growth; so, secondary metabolites have no significant inhibitory activity in these culture and extraction conditions. These results indicated that the inhibitory effect at distance, observed in multiple antagonisms, is likely due to a synergistic effect between metabolites diffusible through the culture medium, and VOCs produced by Nodulisporium sp. GS42d2II1a. Also, the antifungal activity could result from the volatiles mixture activity and not just from the activity of one compound. It is known that VOCs act synergistically to produce the biological activity and lose it when they act alone [9, 38, 39]. On the other hand, the VOCs production could increase or change in multiple direct antagonistic interactions (interspecific interaction); thus, we detected the VOCs produced in four treatments (Nod3, Nod5, Nod-Nod, and Nod-Pyt) to verify this hypothesis.
Like other species of Hypoxylon and Nodulisporium, Nodulisporium sp. GS4d2II1a produced a mixture of volatile secondary metabolites when cultured in vitro. They were detected in the four treatments. MCA showed that volatile metabolites changed depending on the fungus age, which is in agreement with the biological activity observed in the multiple antagonism bioassays. They also changed in the antagonistic interaction. This was more evident in Fig. 9, where four different profiles were formed. In addition, Nodulisporium sp. GS4d2II1a produced 18 compounds constitutively and its production did not depend on the fungus age, inoculum size or whether it was interacting or not with P. aphanidermatum.
The volatiles specific to the intraspecific antagonistic interaction Nod-Nod could serve as a signal for recognition of another Nodulisporium sp. colony. Also, the inoculum size could affect the outcome of the interaction by changing the VOCs produced. The VOCs production only during interspecific antagonistic interaction Nod-Pyt—showed that the pathogen presence induced production of other metabolites that were not synthesized in other treatments (de novo biosynthesis), while other were suppressed. De novo biosynthesis of these metabolites could be achieved through activation of genes that were suppressed in cultures of Nodulisporium sp. GS4d2II1a and activated by biotic stress; that is, the presence of P. aphanidermatum [40]. Induction of terpenes synthesis has been reported through terpenes synthases activation in the mixed fermentations of Fusarium spp. with fungi or bacteria [13, 40, 41]. In this study, three out of the five compounds, exclusively produced in the interspecific interaction, were monoterpenes, and these five compounds could have antifungal properties that inhibit competitors, as demonstrated in studies about co-cultivation of microorganisms for the induction of antimicrobial secondary metabolites [40]. In addition, the change in VOCs profile should be determined when Nodulisporium sp. GS4d2II1a is interacting with other plant pathogenic fungi or oomycetes, as in the multiple antagonism bioassays, because the profile might change depending on the competitors; and, therefore, their biological activity. In addition, the VOCs produced in the interaction could act against other plant pathogens. Thus, Nodulisporium sp. GS4d2II1a likely produce VOCs as a defense mechanism against fungal competitors inside the host plant, but also as host defensive responses against plant pathogens and competitors [31].
The qualitative and quantitative changes in the VOCs profile, during the time of the fungal growth and in antagonistic interaction, were reported previously in wood rotting basidiomycetes [12, 14]. In the study conducted by Hynes et al. [14], it was observed that, although there was no long distance inhibition between the basidiomycetes Hypholoma fasciculare and Resinicium bicolor, VOCs production was detected in both fungi, which changed during the interaction time. In the case of Nodulisporium sp. GS4d2II1a, we observed that VOCs were a distance defense mechanism in the antagonistic interactions. However, they also could be present in antagonistic interactions where no long distance inhibition was observed.
According to these results, further studies should be carried out in order to better understand the ecological functions of the secondary metabolites produced by Nodulisporium sp. GS4d2II1a by making in vivo test in the host plant G. sepium to test if Nodulisporium sp. GS4d2II1a can protect the plants of pathogen attacks by direct inhibition, or by VOCs production. In addition, further studies in crop protection might be needed, since the products described in this work can be useful for biological control against fungal and oomycetes plant pathogens (mycofumigation); either by using an inoculum or more of Nodulisporium sp. GS4d2II1a; or through the interspecific interaction (Nod-Pyt) for the inhibition; or growth control of plant pathogens in postharvest treatments; or directly in crops. Likewise, the chemical composition of the extracts should be studied.
Ecological Function and Biological Activity of Identified VOCs
Terpenes (myrcene, eucalyptol, caryophyllene, terpinene, among others) produced by Nodulisporium sp. GS4d2II1a are essential oils components of medicinal plants and frequently produced by fungi [1, 3, 38].
Terpenes are also ecologically significant. They play important roles in communication among plants, plant–insects, and plant–pathogen interactions. Mono and sesquiterpenes have in some cases antifungal activity, which could constitute an antagonistic mechanism during fungal interactions [12, 42]. Sesquiterpenes are useful for invertebrates to communicate among them, and to localize plant or animal hosts; also, by plants and fungi to attract insects for propagule dispersal, and for defense against pathogens. In addition, they are used for plant growth promotion and as antifungal agents [3, 4, 14, 42].
Some of the compounds produced by Nodulisporium sp. GS4d2II1a were previously identified from other sources. Compounds like eucalyptol, β-caryophyllene, β-elemene, γ-gurjunene, and 3-methyl-1-butanol were also produced by the endophytic Nodulisporium sp., isolated from Myroxylon balsamum; and, its VOCs had antifungal activity against plant pathogens [17]. The compounds limonene, eucalyptol, γ-terpinene, β-ocimene, 3-methyl-1-butanol, 4-carene, δ-guaiene and 1,2,4-tris(methylene)-cyclohexane were identified in the endophyte Nodulisporium sp. CMU-UPE34, isolated from Lagerstroemia loudoni. This fungus has potential as biocontrol agent for postharvest diseases of citrus fruits caused by Penicillium expansum and Penicillium digitatum, and against plant pathogenic fungi [19]. Other endophytic Nodulisporium spp., isolated from Thelypteris angustifolia [18], and Persea indica [16], produce 1,2,4-tris(methylene)-cyclohexane and eucalyptol; and, the endophyte isolated from Cassia fistula [20], also produces eucalyptol. Their VOCs inhibit plant pathogenic fungi and oomycetes. On the other hand, the remaining detected compounds have not been reported previously for the Nodulisporium species.
In addition, the biological functions of several of the individual VOCs identified from Nodulisporium sp. GS4d2II1a were previously reported. For example, the monoterpene eucalyptol or 1,8-cineol is a constituent of essential oils of many medicinal and aromatic plants, with antimicrobial [43] and antifungal activity against Aspergillus flavus, Aspergillus parasiticus [44], Aspergillus niger [45], and Sclerotinia sclerotiorum. Furthermore, a fuel potential has been ascribed to this compound [20]. In addition, β-cariophyllene is produced by F. oxysporum MSA 35 [42] and Talaromyces wortmannii [3, 46]. This sesquiterpene could enhance plant growth and induce tolerance or resistance to the fungal pathogens Colletotrichum higginsianum and Botrytis cinerea. Streptomyces globisporus JK-1 produces β-caryophyllene that inhibits the mycelial growth, conidial germination, and sporulation of Penicillium italicum [47]. Also, endophytes such as Muscodor yucatanensis [31], Phoma sp. [48], and the ectomycorrhizal Paxillus involutus strains MAJ and NAU [49] produce β-cariophyllene. Additionally, the monoterpene limonene is an antifungal compound that inhibits mycelial growth and aflatoxin B1 production of A. flavus [50]. Limonene also suppresses mycelial growth, conidial germination, and sporulation of P. italicum [47]. Thus, eucalyptol, β-cariophyllene, and limonene, some of the compounds found with high fiber affinity, are compounds with antifungal activity that might give the bioactivity to the VOCs mixture produced by Nodulisporium sp. GS4d2II1a. Finally, 2-methyl-1-butanol and 4-carene, volatile compounds produced exclusively in the interspecific interaction, were also detected in endophytes producers of antifungal VOCs [19, 31, 47].
In summary, Nodulisporium sp. GS4d2II1a inhibited growth, or killed plant pathogens, by emitting VOCs, producing diffusible metabolites; and, by hyphal interactions that act in conjunction. These VOCs change qualitatively and quantitatively after 3 and 5 days of endophyte culture, and in the intraspecific and interspecific interaction with P. aphanidermatum. Finally, volatile compounds could be used as biological control of plant pathogenic fungi and oomycetes.
References
Morath SU, Hung R, Bennett JW (2012) Fungal volatile organic compounds: a review with emphasis on their biotechnological potential. Fungal Biol Rev 26:73–83. doi:10.1016/j.fbr.2012.07.001
Yu H, Zhang L, Li L et al (2010) Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol Res 165:437–49. doi:10.1016/j.micres.2009.11.009
Zhi-Lin Y, Yi-Cun C, Bai-Ge X, Chu-Long Z (2012) Current perspectives on the volatile-producing fungal endophytes. Crit Rev Biotechnol 32:363–73. doi:10.3109/07388551.2011.651429
Effmert U, Kalderás J, Warnke R, Piechulla B (2012) Volatile mediated interactions between bacteria and fungi in the soil. J Chem Ecol 38:665–703. doi:10.1007/s10886-012-0135-5
Spraker JE, Jewell K, Roze LV et al (2014) A volatile relationship: profiling an inter-kingdom dialogue between two plant pathogens, Ralstonia solanacearum and Aspergillus flavus. J Chem Ecol 40:502–513. doi:10.1007/s10886-014-0432-2
Gams W, Diederich P, Põldmaa K (2004) Fungicolous Fungi. In: Mueller GM, Bills GF, Foster MS (eds) Biodiversity of fungi: inventory and monitoring methods. Elsevier, Burlington, MA, pp 343–392
Gao F, Dai C, Liu X (2010) Mechanisms of fungal endophytes in plant protection against pathogens. African J Microbiol Res 4:1346–1351
Stinson M, Ezra D, Hess WM et al (2003) An endophytic Gliocladium sp. of Eucryphia cordifolia producing selective volatile antimicrobial compounds. Plant Sci 165:913–922. doi:10.1016/S0168-9452(03)00299-1
Strobel G (2011) Muscodor species—endophytes with biological promise. Phytochem Rev 10:165–172. doi:10.1007/s11101-010-9163-3
Boddy L (2000) Interspecific combative interactions between wood-decaying basidiomycetes. FEMS Microbiol Ecol 31:185–194. doi:10.1111/j.1574-6941.2000.tb00683.x
Rotheray TD, Chancellor M, Jones TH, Boddy L (2011) Grazing by collembola affects the outcome of interspecific mycelial interactions of cord-forming basidiomycetes. Fungal Ecol 4:42–55. doi:10.1016/j.funeco.2010.09.001
Evans J, Eyre C, Rogers HJ et al (2008) Changes in volatile production during interspecific interactions between four wood rotting fungi growing in artificial media. Fungal Ecol 1:57–68. doi:10.1016/j.funeco.2008.06.001
Minerdi D, Bossi S, Gullino ML, Garibaldi A (2009) Volatile organic compounds: a potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA 35. Environ Microbiol 11:844–854. doi:10.1111/j.1462-2920.2008.01805.x
Hynes J, Müller CT, Jones TH, Boddy L (2007) Changes in volatile production during the course of fungal mycelial interactions between Hypholoma fasciculare and Resinicium bicolor. J Chem Ecol 33:43–57. doi:10.1007/s10886-006-9209-6
Park M-S, Ahn J-Y, Choi G-J et al (2010) Potential of the volatile-producing fungus Nodulisporium sp. CF016 for the control of postharvest diseases of apple. Plant Pathol J 26:253–259. doi:10.5423/PPJ.2010.26.3.253
Tomsheck AR, Strobel G, Booth E et al (2010) Hypoxylon sp., an endophyte of Persea indica, producing 1,8-cineole and other bioactive volatiles with fuel potential. Microb Ecol 60:903–14. doi:10.1007/s00248-010-9759-6
Mends MT, Yu E, Strobel GA et al (2012) An endophytic Nodulisporium sp. producing volatile organic compounds having bioactivity and fuel potential. J Pet Environ Biotechnol 03:117. doi:10.4172/2157-7463.1000117
Riyaz-Ul-Hassan S, Strobel G, Geary B, Sears J (2013) An endophytic Nodulisporium sp. from Central America producing volatile organic compounds with both biological and fuel potential. J Microbiol Biotechnol 23:29–35
Suwannarach N, Kumla J, Bussaban B et al (2013) Biofumigation with the endophytic fungus Nodulisporium spp. CMU-UPE34 to control postharvest decay of citrus fruit. Crop Prot 45:63–70. doi:10.1016/j.cropro.2012.11.015
Nigg J, Strobel G, Knighton WB et al (2014) Functionalized para-substituted benzenes as 1,8-cineole production modulators in an endophytic Nodulisporium species. Microbiology 160:1772–1782. doi:10.1099/mic.0.079756-0
Rodrigues KF (1994) The foliar fungal endophytes of the Amazonian palm Euterpe oleracea. Mycologia 86:376. doi:10.2307/3760568
Ellis MB (1971) Dematiaceous Hyphomycetes. Commonwealth Mycological Institute, Kew, Surrey, England
Ju Y-M, Rogers JD (1996) A revision of the genus Hypoxylon. APS, St. Paul, MI
Kornerup A, Wanscher JH (1967) Methuen handbook of colour, 3dth edn. Methuen E, London
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol Ecol 2:113–118. doi:10.1111/J.1365-294x.1993.Tb00005.X
Larena I, Salazar O, González V et al (1999) Design of a primer for ribosomal DNA internal transcribed spacer with enhanced specificity for ascomycetes. J Biotechnol 75:187–194. doi:10.1016/S0168-1656(99)00154-6
Akins RA, Lambowitz AM (1985) General method for cloning Neurospora crassa nuclear genes by complementation of mutants. Mol Cell Biol 5:2272–2278. doi:10.1128/MCB.5.9.2272
Martin KJ, Rygiewicz PT (2005) Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. doi:10.1186/1471-2180-5-28
Altschul SF, Gish W, Miller W et al (1990) Basic local alignment search tool. J Mol Biol 215:403–10. doi:10.1016/S0022-2836(05)80360-2
Macías-Rubalcava ML, Hernández-Bautista BE, Jiménez-Estrada M et al (2008) Naphthoquinone spiroketal with allelochemical activity from the newly discovered endophytic fungus Edenia gomezpompae. Phytochemistry 69:1185–1196. doi:10.1016/j.phytochem.2007.12.006
Macías-Rubalcava ML, Hernández-bautista BE, Oropeza F et al (2010) Allelochemical effects of volatile compounds and organic extracts from Muscodor yucatanensis, a tropical endophytic fungus from Bursera simaruba. J Chem Ecol 36:1122–1131. doi:10.1007/s10886-010-9848-5
Meléndez-González C, Murià-González MJ, Anaya AL et al (2015) Acremoxanthone E, a novel member of heterodimeric polyketides with a bicyclo[3.2.2]nonene ring, produced by Acremonium camptosporum W. GAMS (Clavicipitaceae) endophytic fungus. Chem Biodivers 12:133–147
Adams RP (2007) Identification of essential oil components by gas chromatography/mass spectroscopy, 4th edition, 4th ed. Biochem Syst Ecol. doi:10.1016/0305-1978(96)83708-2
Oliveros JC (2007) VENNY. An interactive tool for comparing lists with Venn diagrams. In: http://bioinfogp.cnb.csic.es/tools/venny/index.html
Costa PS, Santos NC, Cunha P, et al. (2013) The use of Multiple Correspondence Analysis to explore associations between categories of qualitative variables in healthy ageing. J Aging Res 1–12. doi: 10.1155/2013/302163
Semmar N (2013) Two computational simplex approaches to graphical highlighting metabolic phenotypes and their functional origins: Correspondence Analysis and Weighted Metabolic Profiles Analysis. Metabolomics coming age with its Technol. Divers., 1st ed. Elsevier, pp 441–492
Kuhnert E, Fournier J, Peršoh D et al (2014) New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers 64:181–203. doi:10.1007/s13225-013-0264-3
Mulyaningsih S, Sporer F, Zimmermann S et al (2010) Phytomedicine synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Eur J Integr Med 17:1061–1066. doi:10.1016/j.phymed.2010.06.018
Strobel GA, Dirkse E, Sears J, Markworth C (2001) Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 147:2943–2950. doi:10.1099/00221287-147-11-2943
Bertrand S, Bohni N, Schnee S et al (2014) Metabolite induction via microorganism co-culture: a potential way to enhance chemical diversity for drug discovery. Biotechnol Adv 32:1180–1204. doi:10.1016/j.biotechadv.2014.03.001
Müller MEH, Steier I, Köppen R et al (2012) Cocultivation of phytopathogenic Fusarium and Alternaria strains affects fungal growth and mycotoxin production. J Appl Microbiol 113:874–887. doi:10.1111/j.1365-2672.2012.05388.x
Minerdi D, Bossi S, Maffei ME et al (2011) Fusarium oxysporum and its bacterial consortium promote lettuce growth and expansin A5 gene expression through microbial volatile organic compound (MVOC) emission. FEMS Microbiol Ecol 76:342–351. doi:10.1111/j.1574-6941.2011.01051.x
Nikolić B, Mitić-Ćulafić D, Vuković-Gačić B, Knežević-Vukčević J (2011) Modulation of genotoxicity and DNA repair by plant monoterpenes camphor, eucalyptol and thujone in Escherichia coli and mammalian cells. Food Chem Toxicol 49:2035–45. doi:10.1016/j.fct.2011.05.015
Vilela GR, de Almeida GS, D’Arce MABR et al (2009) Activity of essential oil and its major compound, 1,8-cineole, from Eucalyptus globulus Labill., against the storage fungi Aspergillus flavus Link and Aspergillus parasiticus Speare. J Stored Prod Res 45:108–111. doi:10.1016/j.jspr.2008.10.006
Stojković D, Soković M, Glamočlija J et al (2011) Chemical composition and antimicrobial activity of Vitex agnus-castus L. fruits and leaves essential oils. Food Chem 128:1017–1022. doi:10.1016/j.foodchem.2011.04.007
Yamagiwa Y, Inagaki Y, Ichinose Y et al (2011) Talaromyces wortmannii FS2 emits β-caryophyllene, which promotes plant growth and induces resistance. J Gen Plant Pathol 77:336–341. doi:10.1007/s10327-011-0340-z
Li Q, Ning P, Zheng L et al (2010) Fumigant activity of volatiles of Streptomyces globisporus JK-1 against Penicillium italicum on Citrus microcarpa. Postharvest Biol Technol 58:157–165. doi:10.1016/j.postharvbio.2010.06.003
Strobel G, Singh SK, Riyaz-Ul-Hassan S et al (2011) An endophytic/pathogenic Phoma sp. from creosote bush producing biologically active volatile compounds having fuel potential. FEMS Microbiol Lett 320:87–94. doi:10.1111/j.1574-6968.2011.02297.x
Müller A, Faubert P, Hagen M et al (2013) Volatile profiles of fungi-chemotyping of species and ecological functions. Fungal Genet Biol 54:25–33. doi:10.1016/j.fgb.2013.02.005
Singh P, Shukla R, Prakash B et al (2010) Chemical profile, antifungal, antiaflatoxigenic and antioxidant activity of Citrus maxima Burm. and Citrus sinensis (L.) Osbeck essential oils and their cyclic monoterpene, DL-limonene. Food Chem Toxicol 48:1734–1740. doi:10.1016/j.fct.2010.04.001
Acknowledgments
This work was supported by the CONACyT grant 179194. Rosa E. Sánchez-Fernández is a doctoral student from the Posgrado en Ciencias Biomédicas, Instituto de Química, UNAM. She was recipient of a doctoral fellowship from CONACyT, Mexico. We wish to thank to M. Sc. Allan Espinosa Gómez from Instituto de Investigaciones Biomédicas, UNAM, for his help with the molecular identification; to PhD. Bertha Tlapal Bolaños and PhD. Olga Gómez from Instituto de Fitosanidad, Colegio de Postgraduados, Montecillo, Estado de Mexico, for the plant pathogens donation used in the bioassays; to Gonzalo Roque Flores, Instituto de Química, UNAM, for his support with the images edition; to M. Sc. Rebeca Martínez from Instituto de Biología, UNAM, for her support with the brightfield photomicrographs; to M. Sc. Ernesto L. Guevara form Facultad de Medicina, UNAM, for his support in the article revision and especially to M. Sc. Rafael Ibarra Contreras from Facultad de Química, UNAM, for language revision.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Sánchez-Fernández, R.E., Diaz, D., Duarte, G. et al. Antifungal Volatile Organic Compounds from the Endophyte Nodulisporium sp. Strain GS4d2II1a: a Qualitative Change in the Intraspecific and Interspecific Interactions with Pythium aphanidermatum . Microb Ecol 71, 347–364 (2016). https://doi.org/10.1007/s00248-015-0679-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-015-0679-3