
Abstract
Key message
QTLs for kernel characteristics and tolerance to N stress were identified, and the functions of ten known genes with regard to these traits were specified.
Abstract
Kernel size and quality characteristics in wheat (Triticum aestivum L.) ultimately determine the end use of the grain and affect its commodity price, both of which are influenced by the application of nitrogen (N) fertilizer. This study characterized quantitative trait loci (QTLs) for kernel size and quality and examined the responses of these traits to low-N stress using a recombinant inbred line population derived from Kenong 9204 × Jing 411. Phenotypic analyses were conducted in five trials that each included low- and high-N treatments. We identified 109 putative additive QTLs for 11 kernel size and quality characteristics and 49 QTLs for tolerance to N stress, 27 and 14 of which were stable across the tested environments, respectively. These QTLs were distributed across all wheat chromosomes except for chromosomes 3A, 4D, 6D, and 7B. Eleven QTL clusters that simultaneously affected kernel size- and quality-related traits were identified. At nine locations, 25 of the 49 QTLs for N deficiency tolerance coincided with the QTLs for kernel characteristics, indicating their genetic independence. The feasibility of indirect selection of a superior genotype for kernel size and quality under high-N conditions in breeding programs designed for a lower input management system are discussed. In addition, we specified the functions of Glu-A1, Glu-B1, Glu-A3, Glu-B3, TaCwi-A1, TaSus2, TaGS2-D1, PPO-D1, Rht-B1, and Ha with regard to kernel characteristics and the sensitivities of these characteristics to N stress. This study provides useful information for the genetic improvement of wheat kernel size, quality, and resistance to N stress.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wheat (Triticum aestivum L.) is one of the most important food crops worldwide, and it is used to make a variety of food products. Wheat, rice and corn provide more than 60 % of the calories and protein needed for human nutrition (Gill et al. 2004). Breeding new varieties of wheat with enhanced yield potential, high tolerance to biotic and abiotic stresses and improved processing and end-use quality are the three key aims of wheat breeding programs (Mann et al. 2009). Improvement of these traits, which are quantitatively inherited and significantly influenced by the environment, depends on a thorough understanding of their genetic basis.
Kernel size, including kernel weight and dimensions such as kernel length (KL) and kernel width (KW), has significant effects on both yield and quality (Campbell et al. 2001; Sun et al. 2009; Ramya et al. 2010; Tsilo et al. 2010; Cui et al. 2011, 2014b). A comparison of wheat grain yield and yield component selection has shown that selection for kernel weight could result in an increase in grain yield (Alexander et al. 1984). However, an increase in yield often results in a decrease in kernel quality parameters, such as a lower grain protein content (GPC), because of a dilution effect (Suprayogi et al. 2009). Notably, previous studies have shown that quantitative trait loci (QTLs) for yield are not always negatively correlated with quality (Sourdille et al. 2003; Huang et al. 2006; Kunert et al. 2007; Echeverry-Solarte et al. 2015). Therefore, knowledge of the genetic basis of kernel size and its correlation with quality is valuable for the genetic improvement of both yield and quality.
Characteristics that affect the processing and end-use of grain involve a complex group of traits collectively known as quality traits (Kunert et al. 2007). Of them, GPC, wet gluten content (WGC), dough tractility or extensibility (DT), test weight or hectoliter weight (TW), water absorption (ABS), the Zeleny sedimentation score (ZEL), and kernel hardness (KH) are all important grain quality traits in bread wheat. GPC, particularly glutenin and gliadin content is controlled by a complex genetic system and is strongly affected by the environment. It is an important factor for pasta-making and baking quality and is strongly correlated with WGC, DT, ABS, and ZEL (Zanetti et al. 2001; Nelson et al. 2006; Suprayogi et al. 2009; Li et al. 2011). KH, one of the most important determinants of milling yield, has profound effects on the milling, baking and end-use quality of bread wheat (Li et al. 2009). Numerous studies have identified an association between high- and low-molecular-weight (HMW/LMW) glutenin loci (Glu-A1, Glu-B1, and Glu-D1/Glu-A3, Glu-B3, and Glu-D3), and wheat quality, and these loci are located on the long/short arms of the homoeologous group 1 chromosomes (Perretant et al. 2000; Sourdille et al. 2003; Kuchel et al. 2006; Li et al. 2009; Mann et al. 2009; Suprayogi et al. 2009; Sun et al. 2010; Reif et al. 2011; Echeverry-Solarte et al. 2015). In addition, the Ha locus on chromosome 5DS and the Gpc-B1 locus on chromosome 6BS have also been reported to exhibit strong correlations with processing and end-use quality (Sourdille et al. 1996, 2003; Perretant et al. 2000; Campbell et al. 2001; Igrejas et al. 2002; Turner et al. 2004; Nelson et al. 2006; Kunert et al. 2007; Li et al. 2009; Sun et al. 2010). However, the majority of the reports described above have also shown that genetic factors influencing processing and end-use quality in both cultivars and in wild wheat are distributed over all 21 wheat chromosomes.
An adequate quantity of nitrogen (N) and an appropriate time of application can improve yield and quality; however, the use of N fertilizer leads to an increased cost as well as environmental consequences, such as groundwater pollution due to nitrate leaching (Laperche et al. 2007, 2008; Suprayogi et al. 2009; Cui et al. 2014a). Wheat varieties that maintain a high yield and quality under moderate or even severe N deficiency can be adapted for use in low-input management systems. An understanding of the genetic basis of N deficiency tolerance is essential for the breeding of such varieties. However, few QTLs for both yield/quality and their responses to N deficiency in wheat under field conditions have been documented to date (Habash et al. 2007; Laperche et al. 2007, 2008; Cui et al. 2014a). Favorable QTL alleles that decrease differences in yield and/or quality between high N (HN) and lower N (LN) conditions are of value in wheat breeding programs designed to increase N deficiency tolerance (Cui et al. 2014a).
Functional or gene-specific markers are derived from polymorphic sites within genes that are directly associated with phenotypic variations (Liu et al. 2012). In most cases, a functional marker of a gene co-localizes with the QTL peak position for the same trait (Sourdille et al. 2003). To date, numerous functional markers have been discovered that facilitate analyses of the co-localization of identified QTLs with known genes (Giroux and Morris 1997; Lei et al. 2006; He et al. 2009; Wang et al. 2010; Jiang et al. 2011; Ma et al. 2012). Moreover, QTL analysis based on genetic mapping, including analysis of functional markers, can reveal novel functions of important genes that are indirectly correlated with yield and quality traits. In this study, we performed QTL analysis of 11 kernel characteristics under LN and HN conditions using a genetic map enriched with functional markers. The aims of this study were to (1) identify the genetic factors affecting kernel size and quality and the tolerances of these characteristics to N deficiency, (2) characterize the genetic relationships between kernel size and quality, and (3) specify the functions of important genes with regard to kernel characteristics and their sensitivities to N stress.
Materials and methods
Plant materials, experimental design, and phenotypic evaluation
The plant materials included a set of 188 recombinant inbred lines (RILs) derived from Kenong 9204 (KN9204) × Jing 411 (J411) (denoted by KJ-RIL) (Cui et al. 2014a; Fan et al. 2015). The 188 kJ-RILs and their parents were grown from 2011 to 2012 in Shijiazhuang (L1); 2012–2013 in Shijiazhuang (L2), Beijing (L3) and Xinxiang (L4); and 2013–2014 in Shijiazhuang (L5). An LN treatment and an HN treatment were applied in each trial for a total of ten environments (year × location × treatment), designated as E1, E2, E3, E4, E5, E6, E7, E8, E9, and E10. The N treatments, field arrangements and experimental designs of L1 to L4 are detailed in our previous reports (Cui et al. 2014a; Fan et al. 2015). T5 followed the above trial design and was carried out from 2013 to 2014 in Shijiazhuang.
Seven quality traits, including GPC, WGC, DT, TW, ABS, ZEL and KH, were measured by near-infrared reflectance spectroscopy (NIRS) with a Perten DA-7200 instrument (Perten Instruments, Huddinge, Sweden) and expressed on a 14 % moisture basis. The measurements were calibrated using calibration samples according to the manufacturer’s instructions. Previous studies have confirmed that NIRS is a powerful method for measuring the quality traits of wheat; that it is rapid, effective, and convenient compared with traditional methods; and that it is appropriate for use in wheat breeding programs designed to improve quality-related traits (Sourdille et al. 1996; Perretant et al. 2000; Turner et al. 2004; Kuchel et al. 2006; Nelson et al. 2006; Li et al. 2009, 2012; Mann et al. 2009; Suprayogi et al. 2009; Sun et al. 2010; Asif et al. 2015). The four kernel size-related traits were thousand-kernel weight (TKW), KL, KW, and kernel diameter ratio (KDR). These traits were evaluated after harvesting according to Cui et al. (2011). All lengths were reported in millimeters.
The differences in the values for the kernel-related traits (KRTs) between the HN and LN conditions in each trial were calculated as follows: KRTDV = KRT(HN)−KRT(LN), where KRTDV is the difference in the value for a KRT for each line in each trial between the HN and LN conditions, and KRT(HN) and KRT(LN) represent the KRTs under HN and LN, respectively.
Statistical and QTL analyses
Analysis of variance (ANOVA) was performed and phenotypic correlation coefficients were calculated between traits and between N treatments (Spearman’s rank correlation) using SPSS 19.0 (SPSS, Chicago, IL, USA; http://en.wikipedia.org/wiki/SPSS) and QGAStation 2.0 (http://ibi.zju.edu.cn/software/qga/v2.0/index_c.htm). To estimate the broad-sense heritability (h 2B ) of the corresponding traits, ANOVA was performed for each environment separately using QGAStation 2.0 according to Fan et al. (2015). The h 2 B values were calculated using the following formula h 2B = V G/V P; where V G and V P are the genetic variance and phenotypic variance, respectively. To estimate the genotype × location interaction (G × L) and the genotype × N treatment interaction effects (G × T), a combined ANOVA over L1 to L5 was performed using SPSS 19.0. The genotypes (188 RILs), N treatments (LN and HN), and locations (L1, L2, L3, L4 and L5) were considered as fixed effects, whereas the blocks (two replications) were considered as random effects. F tests were considered significant at p < 0.05. Significant differences between genotypes, N treatments, and trials and their interaction effects were assessed by analyzing F-protected least significant difference (LSD) values at a significant level of p < 0.05. Correlations between two traits and Spearman’s rank correlations between HN and LN were assessed by identifying significant Pearson’s correlation coefficients at p < 0.05 using SPSS 19.0. The trait means for the five LN treatments (E1, E3, E5, E7, and E9) and the five HN treatments (E2, E4, E6, E8, and E10) were used to calculate phenotypic correlation coefficients between the traits under the LN and HN conditions, respectively. The 188 KJ-RILs were ranked according to the desired direction of selection for each trait based on the trait means for the five LN and five HN treatments. The numerical order was used to calculate Spearman’s rank correlations cofficients between LN and HN.
A linkage map of the KJ-RIL population has been previously established (Cui et al. 2014a). In this study, 11 functional markers of kernel size and quality-related traits were used to saturate the KJ-RIL-derived genetic map, with the aim of estimating whether the QTLs were co-localized with known genes. The 11 functional markers are related to the genes Glu-B1, Glu-A3, cell wall-bound invertase (TaCwi-A1), sucrose synthase 2 (TaSus2), Hardness (Ha), and phytoene synthase 1 (Psy-B1). Relevant information about these functional markers is listed in Supplementary Table S1. In addition, six Rht-linked markers were used to saturate the previously constructed map. Relevant information about these markers, along with QTLs for plant height-related traits, will be detailed in another paper. The new version of the genetic linkage map amended with the abovementioned functional and Rht-linked markers was used for QTL analysis.
The observed phenotypic values obtained from each environment (E1–E10) were used for QTL mapping analysis. Concerning KRTDV, the phenotypic values of the 188 RILs in L1, L2, L3, L4, and L5 and the means for the above five trials (M) were individually used in QTL mapping analysis. Inclusive composite interval mapping was performed using IciMapping 4.0 based on stepwise regression, with simultaneous consideration of all marker information (http://www.isbreeding.net/). The walking speed chosen for all QTLs was 1.00 cM, with a p value inclusion threshold of 0.001. The threshold for the detection of QTLs was calculated using 1,000 permutations, with a type 1 error of 0.05. All of the QTLs were designated according to Fan et al. (2015). A QTL with an average LOD value >3.0 and an average phenotypic variance contribution >10 % was defined as a major QTL, and one showing significance in at least five/three environments/data sets was considered a stable QTL for the 11 KRTs/KRTDVs.
Results
Phenotypic variation, performance, and correlations
Components of variance obtained by combined ANOVA were all highly significant for all traits. All of these traits, except for KW and KDR, showed significant G × L and G × T interaction effects. For GPC, WGC, ZEL, and KDR, the largest variances were caused by the N treatment (LN and HN), affecting DT, TW, KH, KL, KW, and TKW, and were influenced mainly by location. The highest variance in ABS was caused by a G × L interaction effect. Compared with the HN condition, the LN stress significantly reduced GPC, WGC, DT, ZEL, and KH but increased TW. The LN stress had inconsistent effects on ABS, KL, KW, KDR, and TKW across L1 to L5 (Supplementary Tables S1 and S2).
KN9204 performed better than J411 under both LN and HN for GPC, WGC, DT, ABS, ZEL and KH, with the exception of DT in E6. Conversely, J411 had a higher TW, KL and KDR than KN9204 under both LN and HN. In seven of the ten environments, the KW of KN9204 was slightly higher than that of J411. Conversely, J411 had a slightly higher TKW than KN9204 in seven of the ten environments (Supplementary Table S3). In nearly all cases, all 11 KRTs were quantitatively characterized with h 2B values ranging from 43.8 % (TW, 24.29–69.46 %) to 70.36 % (ABS, 56.65–88.66 %) (Supplementary information, Section 1.1; Supplementary Table S3).
The phenotypic correlations among the 11 traits are listed in Table 1. Of the four kernel size-related traits, both KL and KW were significantly positively correlated with TKW under both LN and HN, whereas KDR was significantly negatively correlated with TKW. All kernel quality-related traits, except for TW, were significantly positively correlated with each other. A comparison of the kernel size-related and kernel quality-related traits revealed that KL and WGC, KW and WGC, KW and DT, KW and ABS, KW and KH, KDR and TW, TKW and DT, TKW and TW, TKW and ABS, and TKW and KH were positively correlated with each other under both LN and HN. The remaining traits were inconsistently correlated with each other across the environments (Supplementary information, Section 1.1).
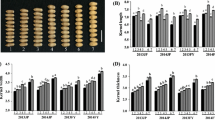
Spearman’s rank correlation coefficients between the two N treatments were high for KL (0.80), TKW (0.79), KDR (0.75), and ABS (0.70), moderate for KH (0.60), KW (0.56), DT (0.57), GPC (0.52), and WGC (0.52), and low for TW (0.36) and ZEL (0.39). Direct selection under HN and LN (up to a 15 % selection intensity) resulted in the selection of 50 % or fewer RILs with similar GPC, TW, ZEL, and KH traits. For example, if the top nine GPC (at a 5 % selection intensity), 19 (at a 10 % selection intensity), and 28 (at a 15 % selection intensity) RILs for GPC in the population were selected for each N treatment, then only fewer than half (2, 8, and 9, respectively) had this trait in common when the selection was based on both treatments. Direct selection under HN and LN showed relatively high consistency for the remaining seven traits investigated, especially for TKW and DT at selection intensities of 5, 10, and 15 % (Table 2; Fig. 1).

Observed rank changes in the top 10 % of the recombinant inbred lines (RILs; 19 lines), ranked under low nitrogen (LN) treatment and high-/normal-nitrogen (HN) conditions, for 11 different traits. The RILs were ranked according to the desired direction of selection [e.g., an RIL with a rank of 1 for grain protein content (GPC) was the highest-ranked RIL]. GPC grain protein content, WGC wet gluten content, DT dough tractility, TW test weight, ABS water absorption, ZEL Zeleny sedimentation value, KH kernel hardness, KL kernel length, KW kernel width, KDR kernel diameter ratio, TKW thousand-kernel weight
Genotype analysis of the ten genes related to kernel size and quality traits and a novel genetic linkage map
KN9204/J411 carried the null, 7 + 9, and 2 + 12/2*, 6 + 8, and 2 + 12 alleles at the three HMW glutenin loci. Concerning the three LMW glutenin loci, KN9204/J411 carried Glu-A3b or Glu-A3d/Glu-A3a or Glu-A3c and exhibited the loss of the Glu-B3/Glu-B3h alleles at Glu-A3 and Glu-B3, respectively. The two parents had the same Glu-D3 allele (Supplementary information, Section 1.2). Analysis of CWI21 and CWI22 revealed that KN9204 and J411 had the TaCwi-A1b and TaCwi-A1a alleles, which caused a decrease and increase in TKW, respectively. Analysis of Sus2-SNP-185/589H2 showed that KN9204 and J411 had the TaSus2 alleles, which resulted in an increase and decrease in TKW, respectively. Analysis of the functional markers of Ha revealed that KN9204 had the Pinb-D1b allele, whereas J411 had the Pinb-D1a allele. Further, analysis of Psy-B1, YP7B-1 and YP7B-2 revealed that KN9204 and J411 had the Psy-B1b and Psy-B1c alleles, respectively.
These functional markers were used to screen the 188 kJ-RILs and to perform linkage analysis to enrich the KJ-RIL map, and they were all assigned to the same chromosomes described in previous reports (Fig. S1). Due to the co-segregation of diagnostic markers for specific genes, the gene names [instead of its corresponding marker name (s)] were listed in the genetic map. In addition, six Rht-linked markers were added to the new version of the KJ-RIL map (data not shown). The novel genetic linkage map included 600 loci spanning 3947.6 cM with an average map density of one marker per 6.58 cM.
QTLs for the 11 KRTs and their responses to low N stress
A total of 109 and 49 QTLs were detected for the KRTs and KRTDVs, respectively. These QTLs were distributed across all wheat chromosomes except for 3A, 4D, 6D, and 7B. Of them, 74 QTLs were mapped to the B genome, 53 were mapped to the A genome, and 31 were mapped to the D genome (Supplementary Tables S4 and S5; Fig. 2). These QTLs individually explained an average of 2.85–35.57 % of the phenotypic variance, with average LOD values ranging from 2.07 to 18.23. Of the 109 QTLs for the 11 KRTs, 38 (34.86 %) were reproducibly detected in at least four of the ten environments (Table 3; Fig. 2). Of the 49 QTLs for the 11 KRTDVs, 14 (28.6 %) were reproducibly detected in at least three of the six data sets (L1, L2, L3, L4, L5, and M) (Table 4; Fig. 2).

Locations of QTLs for the 11 kernel size- and quality-related traits identified in ten individual environments based on a population of 188 kJ-RILs derived from a cross between Kenong 9204 and Jing 411. The short arms chromosomes are located at the top of the figure. The names of the marker loci and the QTLs are listed to the right of the corresponding chromosomes. The environments where the corresponding QTLs were detected are shown in parenthesis. The functional markers or gene names are marked by bold italic typeface. The positions of the marker loci are listed to the left of the corresponding chromosomes. The QTL intervals were associated with a LOD > 2.0, with LOD peak values of more than 2.5
QTLs for the 11 KRTs
Nine QTLs each for GPC, WGC, and DT; eight QTLs each for TW and ABS; and six QTLs each for ZEL and KH were identified. In addition, a total of 11, 13, 13, and 17 QTLs for KL, KW, KDR, and TKW were detected, respectively. These QTLs were distributed across 16 wheat chromosomes but were not found on 3A, 4D, 5A, 6D, or 7B. The favorable alleles for the ten QTLs for the KRTs (all but ZEL) were contributed by both KN9204 and J411. In addition, all six QTL alleles that increased ZEL were contributed by J411 (Table 3; Supplementary Table S4; Fig. 2; Supplementary information, Section 1.3).
QGpc-1A, QGpc-1B.2, QWgc-1A, QWgc-1B.1, QWgc-5D, QDt-1A.2, QDt-2A, QDt-4A, QDt-4B, QDt-5D, QTw-4B.2, QAbs-1B.1, QAbs-3B, QAbs-5D, QZel-2A, QZel-5D, QKh-5D, QKl-1B, QKl-2A.1, QKw-2D, QKdr-2A, QKdr-2D, QKdr-4B.2, QTkw-2D, QTkw-4A, QTkw-4B.2, and QTkw-5D were stable QTLs that were verified in at least five of the ten environments. Moreover, QDt-4B, QTw-4B.2, QAbs-5D, QZel-5D, QKh-5D, QKl-1B, and QTkw-4B.2 were major stable QTLs. Additionally, there were 11 QTLs that were detected reproducibly in four of the ten environments (Table 3; Supplementary Table S4; Fig. 2).
The 27 stable QTLs were distributed among eight chromosomal regions. QGpc-1A and QWgc-1A were within a confidence interval near Glu-A3 on chromosome 1A. QWgc-1B.1 and QAbs-1B.1 were within the confidence interval of wPt-5312–Xme16em12.1 on chromosome 1B. QGpc-1B.2 and QKl-1B co-localized on chromosome 1B in the vicinity of the known gene Glu-B1. QDt-2A, QZel-2A, QKl-2A.1, and QKdr-2A, were within a confidence interval near TaCwi-A1 on chromosome 2A. QKw-2D, QKdr-2D, and QTkw-2D co-localized on chromosome 2D in the vicinity of the known genes TaGS2-D1 and PPO-D1. QDt-4A and QTkw-4A were within the confidence interval of Xgpw2331–Xwmc760 on chromosome 4A. QDt-4B, QTw-4B.2, QKdr-4B.2, and QTkw-4B.2 were co-localized on chromosome 4B in the vicinity of the known gene Rht-B1. Finally, QWgc-5D, QDt-5D, QAbs-5D, QZel-5D, QKh-5D, and QTkw-5D were within a confidence interval near Ha on chromosome 5D (Table 3; Fig. 2).
QTLs for the 11 kernel-related traits with differences between HN and LN
A total of 49 QTLs were identified for the 11 KRTDVs that individually accounted for an average of 4.22–26.81 % of the phenotypic variation, with average LOD values of 2.07–8.57 (Supplementary Table S5; Fig. 2). These QTLs were mapped to chromosomes 1A (4 QTLs), 1B (10 QTLs), 2A (8 QTLs), 2B (3 QTLs), 2D (5 QTLs), 3B (1 QTL), 4B (6 QTLs), 5A (2 QTLs), 5D (4 QTLs), 6B (5 QTLs), and 7A (1 QTL). Twenty-two and 27 of these QTL alleles that reduced the KRTDVs were from KN9204 and J411, respectively.
Fourteen stable QTLs are listed in Table 4. Of them, QGpcdv-4B, QWgcdv-4B, QTwdv-4B, QKwdv-4B, QKdrdv-4B, and QTkwdv-4B were within the confidence interval on chromosome 4B near Rht-B1. All of these QTLs were major stable QTLs, except for QTwdv-4B. QKwdv-5A and QTkwdv-5A were within the confidence interval on chromosome 5A. QAbsdv-5D and QKhdv-5D were co-localized on chromosome 5DS in the vicinity of the known gene Ha. The remaining four stable QTLs were located on chromosomes 1B, 2A, 6B, and 7A (Table 4; Fig. 2).
Coincident QTLs for the KRTs and their responses to low-Nstress
Twenty-five of the 49 QTLs for the KRTDVs coincided with the QTLs for the corresponding KRTs at nine locations on chromosomes 1B, 2A, 2B, 3B, 4B, 5D, 6B, and 7A (Table 5; Fig. 2). The Xbarc199–Xmag4087 interval (near Rht-B1) harbored six pairwise QTLs for KRTs and KRTDVs associated with the traits GPC, WGC, TW, KW, KDR, and TKW. At this chromosomal region, J411 contributed to a higher GPC, WGC, TW, KW, and TKW and to reduced responses of GPC, WGC, and KDR to LN stress, and the KN9204 alleles in this region increased KDR and enhanced the tolerances of TW, KW, and TKW to LN stress. On chromosome 2A, five pairwise QTLs for KRTs and KRTDVs associated with the traits ABS, ZEL, KH, KDR, and TKW were mapped to Xwmc598–Xbarc89. J411 alleles in this chromosomal region contributed to the higher values of all five of these traits and also promoted the enhanced tolerance of ABS, ZEL, and KH and to the reduced tolerance of KDR and TKW to LN stress. Four pairwise QTLs for KRTs including GPC, WGC, KL, and KDR as well as the associated KRTDVs co-segregated with the known gene Glu-B1 on chromosome 1B, with alleles from KN9204 increasing the tolerance of all four traits to LN stress. In this chromosomal region, KN9204 contributed to a higher GPC and WGC and J411 contributed to increased KL and KDR. Four pairwise QTLs for KRTs including ABS, ZEL, KH, and KW as well as the associated KRTDVs co-segregated with the known gene Ha on chromosome 5D, and KN9204 alleles in this region contributed to higher values for all four of these traits, in addition to the enhanced tolerance of ABS, ZEL, and KH to LN stress. QKwdv-2B, QKdrdv-2B, QKw-2B, and QKdr-2B.2 were within the confidence interval of wPt-9402–Xcfe212 on chromosome 2B. KN9204 and J411 alleles in this chromosomal region contributed to a greater KW and KDR, respectively; inversely, alleles that enhanced the tolerances of KW and KDR to LN stress were derived from J411 and KN9204, respectively. One QTL each for KL, KW, and TKW coincided with a QTL for KLDV, KWDV, and TKWDV, respectively, at three locations on chromosomes 3B, 6B, and 7A. J411 contributed to increased KL, KW, and TKW and to decreased KLDV, KWDV, and TKWDV in these chromosomal regions. On chromosome 1B, QZel-1B coincided with QZeldv-1B, and alleles from KN9204 enhanced the tolerance of ZEL to LN stress but reduced the ZEL.
Coincident QTLs for the kernel size- and quality-related traits
Eleven QTL clusters that simultaneously affected kernel size- and quality-related traits were identified on chromosomes 1B (two QTL clusters), 2A, 2B, 2D, 3B, 4A (two QTL clusters), 4B, 5D, and 6B, which harbored 28 QTLs for kernel size-related traits and 39 for kernel quality-related traits (Table 6; Fig. 2). All 11 QTL clusters, except for C4, C8, and C11, harbored major or stable or major stable QTLs for the KRTs. KN9204- or J411-derived alleles in C3, C5, C7, C9, and C11 that enhanced the kernel quality-related traits were also found to increase kernel size. In C1 and C10, KN9204 contributed to increased values for both kernel quality-related traits and to kernel size-related traits, with the exception of TW. In C2, alleles from KN9204 had additive effects on increases in the values for kernel quality-related traits and decreases in kernel size. In C4 and C8, alleles from J411 had additive effects on increases in the values of kernel quality-related traits and decreases in kernel size. In C6, KN9204 contributed to increased ABS and DT but to decreased ZEL, TW, and KL.
Discussion
Genetic relationships between kernel size and quality traits
In addition to productive spikes per unit area, kernels per spike, and kernel weight, kernel size is another major determinant of wheat yield (Cui et al. 2014b). The negative yield-quality correlation has long been a key obstacle in wheat breeding programs designed to simultaneously improve yield and quality (Bathia and Rabson 1987; Nelson et al. 2006; Kunert et al. 2007; Suprayogi et al. 2009). Consequently, QTLs involved in the control of kernel quality that are also independent of negative effects on grain yield should be identified to improve this characteristic.
In this study, TKW showed significant positive correlations with DT, TW, ABS, and KH under LN and HN. It also exhibited positive correlations with GPC, WGC, and ZEL, although these correlations were non-significant under LN (Table 1). These findings imply that common genetic factors that simultaneously cause increase in TKW and enhancement of quality traits exist in KN9204 and/or J411. Five such QTL clusters were identified in C3, C5, C7, C9, and C11 (Table 6; Fig. 2). These chromosomal regions should be valuable for the genetic improvement of both kernel size and quality. For example, in the QTL cluster of C3, J411-derived alleles should be preferably selected for the simultaneous genetic improvement of ABS, WGC, KH, GPC, DT, ZEL, KDR, KW, KL, and TKW (Table 6). The ten markers in the Xwmc598–Xbarc89 interval should be used in the foreground selection. Thus, these QTL clusters should be precisely mapped and cloned to identify markers for more efficient selection.
Five chromosomal regions on chromosomes 1A (two regions), 2D, 3B, and 7D harbored QTLs that only affected kernel quality (Supplementary Table S4; Fig. 2). Eight chromosomal regions harbored QTLs that only affected the kernel size and were distributed among chromosomes 1A, 3B, 3D, 4A, 6A, 7A (two regions) and 7D (Supplementary Table S4; Fig. 2). These findings indicate that kernel size and the quality traits are under independent genetic control in some cases. Thus, these chromosomal regions should be valuable for the genetic improvement of quality/kernel size without influencing kernel size/quality traits. Further, some of these QTLs were stable across the tested environments, for example, the QTLs QGpc-1A and QWgc-1A, which had overlapping confidence intervals on chromosome 1A and were significant in six and five environments, respectively. Such stable QTLs are of particular value for marker-assisted selection in the breeding of varieties adapted to various ecological environments.
Would indirect selection of superior genotypes under normal conditions maximize the genetic gain in breeding programs designed to improve tolerance to N-deficient conditions?
Indirect selection of superior genotypes in different systems will not always result in selection of the best possible genotypes for improving stress resistance in breeding programs. A recent study has shown that direct selection in a conventional and organic management system (selection intensity of up to a 15 %) resultes in the selection of 50 % or fewer RILs with common yield- and quality-related traits (Asif et al. 2015). Ceccarelli and Grando (1991) have reported that the best lines selected in low-yield environments outperform superior lines selected in high-yield environments. In addition, the indirect selection efficiency of superior genotypes is trait dependent, and indirect selection can lead to greater genetic gains than direct selection for highly heritable traits (Hill et al. 1998; Asif et al. 2015).
In this study, the Spearman’s rank correlation coefficient between the treatments for TKW was second to that for KL among the 11 traits investigated (Table 2; Fig. 1). Indirect selection for TKW can maximize genetic gain for the improvement of N stress tolerance. Similar inferences were also made for KL, KW, and KDR because they showed strong correlations with TKW. For DT and WGC, indirect selection under HN allowed for the genetic improvement of DT and WGC under LN in most cases; however, the negative effects of N stress on both DT and WGC were difficult to counteract. Indirect selection under HN allowed for the genetic improvements of ABS under LN to some extent, and it resulted in lower efficiencies for the genetic improvement of GPC, TW, ZEL, and KH under LN (Supplementary information, Section 2.1).
The environments in which a QTL is expressed determine the manner by which breeders use the corresponding QTL in molecular breeding programs, especially for stable QTLs. All the 27 stable QTLs for the 11 traits were expressed in both N environments (Supplementary Table S4; Fig. 2). This finding indicates that marker-assisted selection using the flanking markers of these QTLs is efficient under both HN and LN conditions. However, three of these stable QTLs—QWgc-1B.1, QTkw-4B.2, and QTkw-5D—were mainly expressed under HN condition; therefore, the flanking markers of these QTLs are useful for breeding wheat lines that can be grown under optimal N conditions in most cases. Of the 82 environment-specific QTLs, 17 and 23 were expressed only under LN and HN conditions, respectively, and the remaining 42 environment-sensitive QTLs were expressed under both N conditions (Supplementary Table S4; Fig. 2). QGpc-2B, QWgc-2B, QWgc-7D, QAbs-2D.2, QKw-4B.2, and QKdr-1B were significant in three of the five HN environments, and these six QTLs are useful for breeding wheat for growth under optimal N conditions. QGpc-4B, QWgc-1B.2, QWgc-2A, QAbs-2A, QKw-6B, and QTkw-6A were significant in three of the five LN environments, and these six QTLs are useful for breeding wheat for growth under N-stress environments.
Genomic regions conferring tolerance to low-N stress and their value in wheat breeding programs
Wheat varieties that maintain yield and quality under N stress can adapt to low-input management systems (Cui et al. 2014a). The examination of genetic variation in adaptive traits for N deficiency is essential for the breeding of varieties with N stress tolerate. However, few QTLs for yield and quality and their responses to N deficiency in wheat under field conditions have been documented (Habash et al. 2007; Laperche et al. 2007, 2008; Cui et al. 2014a).
QTL alleles enhance both KRTs and their tolerances to N stress should be valuable in wheat molecular breeding programs. In this study, 25 QTLs for KRTDVs were within the confidence intervals of the QTLs for the corresponding KRTs (Table 5; Fig. 2). Of them, 13 pairwise QTLs for KRTDVs and the corresponding KRTs showed opposite additive effects, indicating that these regions harbor genes that simultaneously enhance KRTs and their N stress tolerance. Interestingly, QAbsdv-5D, QAbs-5D, QKhdv-5D, and QKh-5D all mapped to locations near the Ha locus, and they were all stable across the tested environments. These findings indicated that the Ha locus tended to have stable pleiotropic effects on both the kernel quality-related traits (especially KH and ABS) and their responses to N stress. QGpcdv-4B, QGpc-4B, QWgcdv-4B, and QWgc-4B all mapped to locations near Rht-B1, and each was identified in at least four environments, except for QWgc-4B, which was significant in two environments. These findings indicated that the Rht-B1 affected not only plant height but also grain quality and its adaptability to N-deficient environments. QTwdv-7A and QTw-7A both mapped to Xbarc219–Xbarc174 on chromosome 7A and were verified in four of the six data sets and in two of the ten environments, respectively. Thus, closely linked markers of these QTLs are of value for the genetic improvement of LN stress tolerance.
Stable QTLs for KRTDVs that have no direct effects on the corresponding KRTs can also be used for the genetic improvement of LN stress tolerance. In this study, such stable QTLs were distributed among chromosomes 2A (QAbsdv-2A.2), 6B (QKhdv-6B), and 5A (Kwdv-5A and Tkwdv-5A) and were verified in three, four, three, and three data sets, respectively (Table 4). Of them, Kwdv-5A and Tkwdv-5A both mapped to Xmag1241–Xme12em13.1 on chromosome 5A, and Tkwdv-5A was a major stable QTL individually accounting for an average of 11.77 % of the phenotypic variance, with an average LOD value of 4.67. Thus, markers that are closely linked with these QTLs could be used for the genetic improvement of these traits for LN stress tolerance.
Novel, stable QTLs and QTL co-segregation with known genes
Numerous genes associated with simply inherited traits have already been mapped, and corresponding functional makers have been developed (Liu et al. 2012). In most cases, the functional marker of a gene is located in very close proximity to the QTL for the same trait (Sourdille et al. 2003). The previous KJ-RIL-derived genetic map included seven functional markers, namely Ax2* (Glu-A1), FM1 (UREG), Glu-b3h (Glu-B1), In10 (Gs2-D1), PPO33 (PPO-A1), STS01 (PPO-D1), and FM2 (PIP) (Cui et al. 2014a). In this study, 11 functional markers of kernel size- and quality-related traits, along with six Rht-linked markers, were added to the new version of the KJ-RIL genetic map (Supplementary Fig. S1). These data can facilitate the determination of whether the QTLs for KRTs are co-localized with known genes.
Numerous studies have identified associations between HMW and LMW glutenin subunits and wheat quality (Payne 1987; Perretant et al. 2000; Sourdille et al. 2003; McCartney et al. 2006; Nelson et al. 2006; Zhang et al. 2008; Li et al. 2009; Mann et al. 2009; Suprayogi et al. 2009; Reif et al. 2011; Li et al. 2012; Echeverry-Solarte et al. 2015). In this study, KN9204 and J411 differed at two HMW glutenin loci (Glu-A1 and Glu-B1) and two LMW glutenin loci (Glu-A3 and Glu-B3). QTL mapping analysis showed that the 2*-HMW subunit encoded by the Glu-1Ab allele from J411 had a positive effect on DT; that the Bx7 + By9 allele from KN9204 could increase GPC and WGC, reduce kernel size, and increase the tolerance of GPC, WGC, KL, and KDR to LN stress; and that the Glu-A3b or Glu-A3d alleles from KN9204 could simultaneously increase GPC, WGC, and DT (Table 5). No adverse effects of the 1BL.1RS translocation on the quality trait were identified for Glu-B3, consistent with Johnson et al. (1999) (Supplementary information, Section 2.2).
The cell wall invertase (CWI) gene is associated with TKW (Ma et al. 2012; Liu et al. 2012; Rasheed et al. 2014). In this study, QTL mapping analysis showed that the TaCwi-A1a allele may be associated with not only an increased kernel size but also improved kernel milling and baking quality and that it could enhance the tolerance of both quality and yield to N stress. The sucrose synthase 2 (Sus2) gene is associated with wheat yield (Jiang et al. 2011; Liu et al. 2012; Rasheed et al. 2014). In this study, QTL mapping analysis showed that TaSus2 had opposite effects on the yield and quality traits. In addition, TaSus2 could influence the responses of KL and KW to N stress, but it had no effect on the response of per-plant yield (Cui et al. 2014a). Glutamine synthetase (GS2) is a key enzyme involved in plant N metabolism that is responsible for the first step of ammonium assimilation and transformation into glutamine; thus, it plays a key roles in the growth, N use and yield potential of cereal crops, including wheat (Gadaleta et al. 2011; Li et al. 2011). Polyphenol oxidase (PPO) activity is highly correlated with the undesirable browning of wheat-based end products, especially Asian noodles (He et al. 2007; Liu et al. 2012). TaGS2-D1 and PPO-D1 have been mapped to 210.30 and 214.80 cM on chromosome 2DL in the KJ-RIL map, respectively (Cui et al. 2014a). In this study, QTL mapping analysis showed that TaGS2-D1a from KN9204 had positive effects on KW, TKW, and KDR, and that PPO-D1 from KN9204 had positive effects on DT and ABS. QKh-2D.1 and QTw-2D might have been identified due to the presence of other linked genes. The Rht-B1 gene is associated with physiological processes that affect not only plant height but also yield, grain quality, seedling vigor and adaptability to adverse environments (Fischer and Stockman 1986; Gooding et al. 1999, 2012; McCartney et al. 2006; Zheng et al. 2010; Bai et al. 2013; Zhang et al. 2013; Asif et al. 2015). This study indicated that Rht-B1b had negative effects on TKW, KW, KDR, TW, DT, and ZEL. The associations with QGpc-4B and QWgc-4B might have been due to the presence of other linked genes (Supplementary Fig. S2). In addition, Rht-B1b enhanced the tolerance of TW, KW, and TKW to N-deficiency (Table 5). KH, which has a profound effect on milling and end-use quality, is largely determined by the Pina-D1 and Pinb-D1 genes that encode the puroindoline a and puroindoline b proteins, respectively (Li et al. 2009; Liu et al. 2012). This study indicated that Pinb-D1b had pleiotropic effects that involved increasing KH and thus increasing WGC, DT, ABS, ZEL, KW, and TKW while decreasing TW, consistent with the additive effects listed in Table 6 (Supplementary Fig. S3). Moreover, the Pinb-D1b alleles from KN9204 enhanced the tolerance of KH, ABS, and ZEL to N deficiency but reduced the tolerance of KW (Table 5) (Supplementary information, Section 2.2).
No QTLs for kernel characteristics were mapped to areas near the remaining functional markers on the KJ-RIL genetic map, which might be attributed to the absence of associated genes affecting these traits in these regions. In addition, functional markers of several important major genes for grain shape and size including GS3, GW2 and GASR7, did not show polymorphisms between the two parental lines (data not shown). We detected QTLs for kernel size on the corresponding chromosomes, including 6A, 7A, and 7D, which should have no associations with these genes. To infer whether common interacting QTLs or regions were presented across genetic backgrounds, we compared the results of our study with those of similar previous studies on wheat (Table 3). Of the 38 QTLs that were reproducibly detected in at least four of the ten environments, 26 had been reported in previous studies. All of the QTLs that were found to be located near the abovementioned functional markers were QTLs or regions that commonly interacted across genetic backgrounds and have been reported in numerous similar studies (Table 3). In addition, QWgc-1A, QWgc-1B.1, QAbs-1B.1, QKl-1B, QDt-2A, QKdr-2A, QKl-2A.1, QWgc-5D, QDt-4A, QDt-4B, QKw-6B, and QDt-7D were novel QTLs that have not been reported previously, but some of them coincided with QTLs that interacted with other traits, indicating pleiotropic effects or the presence of linked genes. Interestingly, all of these novel QTLs except for QKw-6B and QDt-7D were stable across the environments, and both QKl-1B and QDt-4B were major stable QTLs. To our knowledge, this is the first study to report contributions of all of the QTLs for these 11 KRTs to N stress tolerance, and 14 of these QTLs were found to be stable across environments (Table 4). These findings should be valuable for the genetic improvement of both kernel characteristics and N-deficiency tolerance via marker-assisted selection.
In summary, this study has revealed the genetic factors affecting kernel size and quality and has provided the first report of QTLs associated with N-deficiency tolerance. A total of 109 putative additive QTLs for the 11 KRTs and 49 QTLs for the 11 KRTDVs were identified, 27 and 14 of which were stable across environments, respectively. Genetic relationships between kernel size and quality were well characterized at the QTL level. The feasibility of indirect selection of a superior genotype for kernel size and quality under normal N conditions in breeding programs designed for adaptation to a lower-input management system was discussed in depth. In addition, we specified the functions of genes important to kernel characteristics and the sensitivities of these characteristics to N stress. This study provides useful information for the genetic improvement of wheat kernel-related traits and the tolerance of wheat to N stress.
Author contribution statement
Fa Cui, Xiaoli Fan, and Junming Li designed the study; Fa Cui, Xiaoli Fan, Chunhua Zhao, Na Zhang and Mei Chen conducted genotyping of the KJ-RIL population; Fa Cui, Xiaoli Fan, Mei Chen, Chunhua Zhao, Wei Zhang, Ji Jun, Jie Han, Xueqiang Zhao, Lijuan Yang, Yiping Tong, Zongwu Zhao, and Junming Li conducted phenotyping of the KJ-RIL population; Fa Cui analyzed the data and wrote the paper; and Junming Li and Tao Wang shared primary responsibility for the final content. All authors read and approved the final manuscript.
Abbreviations
- GPC:
-
Grain protein content
- WGC:
-
Wet gluten content
- DT:
-
Dough tractility
- TW:
-
Test weight
- ABS:
-
Water absorption
- ZEL:
-
Zeleny sedimentation value
- KH:
-
Kernel hardness
- KL:
-
Kernel length
- KW:
-
Kernel width
- KDR:
-
Kernel diameter ratio
- TKW:
-
Thousand-kernel weight
- KRT:
-
Kernel-related trait
- KRTDV:
-
Differences in the value for a kernel-related trait between the high-nitrogen and low-nitrogen treatments
References
Alexander WL, Smith EL, Dhanasobhan C (1984) A comparison of yield and yield component selection in winter wheat. Euphytica 33:953–961. doi:10.1007/BF00021926
Ammiraju JSS, Dholakia BB, Santra DK, Singh H, Lagu MD, Tam-hankar SA, Dhaliwal HS, Rao VS, Gupta VS, Ranjekar PK (2001) Identification of inter simple sequence repeat (ISSR) markers associated with seed size in wheat. Theor Appl Genet 102:726–732. doi:10.1007/s001220051703
Asif M, Yang RC, Navabi A, Iqbal M, Kamran A, Lara EP, Randhawa H, Pozniak C, Spaner D (2015) Mapping QTL, selection differentials, and the effect of Rht-B1 under organic and conventionally managed systems in the Attila × CDC Go spring wheat mapping population. Crop Sci 55:1129–1142. doi:10.2135/cropsci2014.01.0080
Bai C, Liang YL, Hawkesford MJ (2013) Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat. J Exp Botany 64:1745–1753. doi:10.1093/jxb/ert041
Bathia CR, Rabson R (1987) Relationship of grain yield and nutritional quality. In: Nutritional quality of cereal grains: genetic and agronomic improvement. Agronomy Monograph No. 28, ASA-CSSA-SSSA, Madison, pp 11–43
Breseghello F, Finey PL, Gaines C, Andrews L, Tanaka J, Penner G, Sorrells ME (2005) Genetic loci related to kernel quality differences between a soft and a hard wheat cultivar. Crop Sci 45:1685–1695. doi:10.2135/cropsci2004.0310
Campbell KG, Finney PL, Bergman CJ, Gualberto DG, Anderson JA, Giroux MJ, Siritunga D, Zhu J, Gendre F, Roué C, Vérel A, Sorrells ME (2001) Quantitative trait loci associated with milling and baking quality in a soft × hard wheat cross. Crop Sci 41:1275–1285. doi:10.2135/cropsci2004.0310
Ceccarelli S, Grando S (1991) Environment of selection and type of germplasm in barley breeding for low-yielding conditions. Euphytica 57:207–219. doi:10.1007/BF00039667
Cui F, Ding A, Li J, Zhao C, Li X, Feng D, Wang X, Wang L, Gao J, Wang H (2011) Wheat kernel dimensions: how do they contribute to kernel weight at an individual QTL? J Genet 90:409–425. doi:10.1007/s12041-011-0103-9
Cui F, Fan X, Zhao C, Zhang W, Chen M, Ji J, Li J (2014a) A novel genetic map of wheat: utility for mapping QTL for yield under different nitrogen treatments. BMC Genet 15:57. doi:10.1186/1471-2156-15-57
Cui F, Zhao CH, Ding A, Li J, Wang L, Li X, Bao Y, Li J, Wang H (2014b) Construction of an integrative linkage map and QTL mapping of grain yield–related traits using three related wheat RIL populations. Theor Appl Genet 127:659–675. doi:10.1007/s00122-013-2249-8
Cuthbert JL, Somers DJ, Brũlé-Babel AL, Brown PD, Crow GH (2008) Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor Appl Genet 117:595–608. doi:10.1007/s00122-008-0804-5
Echeverry-Solarte M, Kumar A, Kianian S, Simsek S, Alamri MS, Mantovani EE, McClean PE, Deckard EL, Elias E, Schatz B, Xu SS, Mergoum M (2015) New QTL alleles for quality-related traits in spring wheat revealed by RIL population derived from supernumerary × non-supernumerary spikelet genotypes. Theor Appl Genet 128:893–912. doi:10.1007/s00122-015-2478-0
Fan X, Cui F, Zhao C, Zhang W, Yang L, Zhao X, Han J, Su Q, Ji J, Zhao Z, Tong Y, Li J (2015) QTLs for flag leaf size and their influence on yield-related traits in wheat (Triticum aestivum L.). Mol Breeding. doi:10.1007/s11032-015-0205-9
Fischer R, Stockman Y (1986) Increased kernel number in Norin 10-derived dwarf wheat: evaluation of the cause. Aust J Plant Physio 13:767–784. doi:10.1071/PP9860767
Gadaleta A, Nigro D, Giancaspro A, Blanco A (2011) The glutamine synthetase (GS2) genes in relation to grain protein content of durum wheat. Funct Integr Genomics 11:665–670. doi:10.1007/s10142-011-0235-2
Gegas VC, Nazari A, Griffiths S, Simmonds J, Fish L, Orford S, Sayers L, Doonan JH, Snape JW (2010) A genetic framework for grain size and shape variation in wheat. Plant Cell 22:1046–1050. doi:10.1105/tpc.110.074153
Gill BS, Appels R, Botha-Oberholster A-M, Buell CR, Bennetzen JL, Chalhoub B, Chumley F, Dvořák J, Iwanaga M, Keller B, Li W, McCombie R, Ogihara Y, Quetier F, Sasaki T (2004) A workshop report on wheat genome sequencing: international genome research on wheat consortium. Genetics 168:1087–1096. doi:10.1534/genetics.104.034769
Giroux MJ, Morris CF (1997) A glycine to serine change in puroindoline b is associated with wheat grain hardness and low levels of starch-surface friabilin. Theor Appl Genet 95:857–864. doi:10.1007/s001220050636
Gooding MJ, Canon ND, Thompson AJ, Davies WP (1999) Quality and value of organic grain from contrasting bread making wheat varieties and near isogenic lines differing in dwarfing genes. Biol Agri Hortic 16:335–350. doi:10.1080/01448765.1999.9755237
Gooding MJ, Uppal RK, Addisu M, Harris KD, Uauy C, Simmonds JR, Murdoch AJ (2012) Reduced height alleles (Rht) and Hagberg falling number of wheat. J Cereal Sci 55:305–311. doi:10.1016/j.jcs.2012.01.003
Habash DZ, Bernard S, Schondelmaier J, Weyen J, Quarrie SA (2007) The genetics of nitrogen use in hexaploid wheat: N utilisation, development and yield. Theor Appl Genet 114:403–419. doi:10.1007/s00122-006-0429-5
He XY, He ZH, Zhang LP, Sun DJ, Morris CF, Fuerst EP, Xia XC (2007) Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat. Theor Appl Genet 115:47–58. doi:10.1007/s00122-007-0539-8
He XY, He ZH, Ma W, Appels R, Xia XC (2009) Allelic variants of phytoene synthase 1 (Psy1) genes in Chinese and CIMMYT wheat cultivars and development of functional markers for flour colour. Mol Breed 23:553–563. doi:10.1007/s11032-009-9255-1
Hill J, Becker HC, Tigerstedt PMA (1998) Quantitative and ecological aspects of plant breeding. Chapman and Hall, London
Huang XQ, Cöster H, Ganal MW, Röder MS (2003) Advanced backcross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat (Triticum aestivum L.). Theor Appl Genet 106:1379–1389. doi:10.1007/s00122-002-1179-7
Huang XQ, Cloutier S, Lycar L, Radovanovic N, Humphreys DG, Noll JS, Somers DJ, Brown PD (2006) Molecular detection of QTL for agronomic and quality traits in a doubled haploid population derived from two Canadian wheats (Triticum aestivum L.). Theor Appl Genet 113:753–766. doi:10.1007/s00122-006-0346-7
Igrejas G, Leroy P, Charmet G, Gaborit T, Marion D, Branlard G (2002) Mapping QTLs for grain hardness and puroindoline content in wheat (Triticum aestivum L.). Theor Appl Genet 106:19–27. doi:10.1007/s00122-002-0971-8
Jiang Q, Hou J, Hao C, Wang L, Ge H, Dong Y, Zhang X (2011) The wheat (T. aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits. Funct Integr Genom 11:49–61. doi:10.1007/s10142-010-0188-x
Johnson JM, Griffey CA, Harris CH (1999) Comparative effects of 1BL/1RS translocation in relation to protein composition and milling and baking quality of soft red winter wheat. Cereal Chem 76:467–472. doi:10.1094/CCHEM.1999.76.4.467
Kuchel H, Landridge P, Mosinek L, Williams K, Jefferies SP (2006) The genetic control of milling yield, dough rheology and baking quality of wheat. Theor Appl Genet 112:1487–1495. doi:10.1007/s00122-006-0252-z
Kunert A, Naz AA, Dedeck O, Pillen K, Léon J (2007) AB-QTL analysis in winter wheat: I. Synthetic hexaploid wheat (T. turgidum ssp. dicoccoides × T. tauschii) as a source of favourable alleles for milling and baking quality traits. Theor Appl Genet 115:683–695. doi:10.1007/s00122-007-0600-7
Laperche A, Brancourt-Hulmel M, Heumez E, Gardet O, Hanocq E, Devienne-Barret FD, Gouis JL (2007) Using genotype × nitrogen interaction variables to evaluate the QTL involved in wheat tolerance to nitrogen constraints. Theor Appl Genet 115:399–415. doi:10.1007/s00122-007-0575-4
Laperche A, Gouis JL, Hanocq E, Brancourt-Hulmel M (2008) Modelling nitrogen stress with probe genotypes to assess genetic parameters and genetic determinism of winter wheat tolerance to nitrogen constraint. Ephytica 161:259–271. doi:10.1007/s10681-007-9433-3
Lei ZS, Gale KR, He ZH, Gianibelli C, Larroque O, Xia XC, Butow BJ, Ma W (2006) Y-type gene specific markers for enhanced discrimination of high molecular weight glutenin alleles at the Glu-B1 locus in hexaploid wheat. J Cereal Sci 43:94–101. doi:10.1016/j.jcs.2005.08.003
Li Y, Song Y, Zhou R, Branland Jia J (2009) Detection of QTLs for bread-making quality in wheat using a recombinant inbred line population. Plant Breed 128:235–243. doi:10.1007/s00122-012-1829-3
Li X, Zhao X, He X, Zhao G, Li B, Liu D, Zhang A, Zhang X, Tong Y, Li Z (2011) Haplotype analysis of the genes encoding glutamine synthetase plastic isoforms and their association with nitrogen-use- and yield-related traits in bread wheat. New Phytol 189(2):449–458. doi:10.1111/j.1469-8137.2010.03490.x
Li J, Cui F, Ding A, Zhao C, Wang X, Wang L, Bao Y, Qi X, Li X, Gao J, Feng D, Wang H (2012) QTL detection of seven quality traits in wheat using two related recombinant inbred line populations. Euphytica 183:207–226. doi:10.1007/s10681-011-0448-4
Liu Y, He Z, Appels R, Xia X (2012) Functional markers in wheat: current status and future prospects. Theor Appl Genet 125:1–10. doi:10.1007/s00122-012-1829-3
Ma D, Yan J, He Z, Wu L, Xia X (2012) Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers. Mol Breed 1:43–52. doi:10.1007/s11032-010-9524-z
Mann G, Diffey S, Cullis B, Azanza F, Martin D, Kelly A, McIntyre L, Schmidt A, Ma W, Nath Z, Kutty I, Leyne PE, Rampling L, Quail KJ, Morell MK (2009) Genetic control of wheat quality: interactions between chromosomal regions determining protein content and composition, dough rheology, and sponge and dough baking properties. Theor Appl Genet 118:1519–1537. doi:10.1007/s00122-009-1000-y
McCartney CA, Somers DJ, Humphreys DG, Lukow O, Ames N, Noll J, Cloutier S, McCallum BD (2005) Mapping quantitative trait loci controlling agronomic traits in the spring wheat cross RL4452 × ‘AC Domain’. Genome 48:870–883. doi:10.1139/g05-055
McCartney CA, Somers DJ, Lukow O, Ames N, Noll J, Cloutier S, Humphreys DG, McCallum BD (2006) QTL analysis of quality traits in the spring wheat cross RL4452 × ‘AC Domain’. Plant Breed 125:565–575. doi:10.1111/j.1439-0523.2006.01256.x
Nelson JC, Andreescu C, Breseghello F, Finney PL, Gualberto DG, Bergman CJ, Peña RJ, Perretant MR, Leroy P, Qualset CO, Sorrells ME (2006) Quantitative trait locus analysis of wheat quality traits. Euphytica 149:145–159. doi:10.1007/s10681-005-9062-7
Payne PI (1987) Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu Rev Plant Physiol 38:141–153. doi:10.1146/annurev.pp.38.060187.001041
Perretant MR, Cadalen T, Charmet G, Sourdlle P, Nicolas P, Boeuf C, Tixier MH, Branlard G, Bernard S, Bernard M (2000) QTL analysis of bread-making quality in wheat using a doubled haploid population. Theor Appl Genet 100:1167–1175. doi:10.1007/s001220051420
Ramya P, Chaubal A, Kulkarni K, Gupta L, Kadoo N, Dhaliwal HS, Chhuneja P, Lagu M, Gupt P (2010) QTL mapping of 1000-kernel weight, kernel length, and kernel width in bread wheat (Triticum aestivum L.). J Appl Genet 51(4):421–429. doi:10.1007/BF03208872
Rasheed A, Xia X, Ogbonnaya F, Mahmood T, Zhang Z, Mujeeb-Kazi A, He ZH (2014) Genome-wide association for grain morphology in synthetic hexaploid wheats using digital imaging analysis. BMC Plant Biol 14:128. doi:10.1186/1471-2229-14-128
Reif JC, Gowda M, Maurer HP, Longin CFH, Korzun V, Ebmeyer E, Bothe R, Pietsch C, Würschum T (2011) Association mapping for quality traits in soft winter wheat. Theor Appl Genet 122:961–970. doi:10.1007/s00122-010-1502-7
Sourdille P, Perretant MR, Charmet G, Leroy P, Gautier MF, Joudrier P, Nelson JC, Sorrells ME, Bernard M (1996) Linkage between RFLP markers and gene affecting kernel hardness in wheat. Theor Appl Genet 93:580–586. doi:10.1007/BF00417951
Sourdille P, Cadalen T, Guyomarc’h H, Snape JW, Perretant MR, Charmet G, Boeuf C, Bernard S, Bernard M (2003) An update of the Courtot × Chinese Spring intervarietal molecular marker linkage map for the QTL detection of agronomic traits in wheat. Theor Appl Genet 106:530–538. doi:10.1007/s00122-002-1044-8
Sun X, Wu K, Zhao Y, Kong F, Han G, Jiang HM, Huang X, Li R, Wang H, Li S (2009) QTL analysis of kernel shape and weight using recombinant inbred lines in wheat. Euphytica 165:615–624. doi:10.1007/s10681-008-9794-2
Sun X, Marza F, Ma H, Carver BF, Bai G (2010) Mapping quantitative trait loci for quality factors in an inter-class cross of US and Chinese wheat. Theor Appl Genet 120:1041–1051. doi:10.1007/s00122-009-1232-x
Suprayogi Y, Pozniak CJ, Clarke FR, Clarke JM, Knox RE, Singh AK (2009) Identification and validation of quantitative trait loci for grain protein concentration in adapted Canadian durum wheat populations. Theor Appl Genet 119:437–448. doi:10.1007/s00122-009-1050-1
Tsilo TJ, Hareland GA, Simsek S, Chao S, Anderson JA (2010) Genome mapping of kernel characteristics in hard red spring wheat breeding lines. Theor Appl Genet 121:717–730. doi:10.1007/s00122-010-1343-4
Turner AS, Bradburne RP, Fish L, Snape JW (2004) New quantitative trait loci influencing grain texture and protein content in bread wheat. J Cereal Sci 40:51–60. doi:10.1016/j.jcs.2004.03.001
Wang RX, Hai L, Zhang XY, You GX, Yan CS, Xiao SH (2009) QTL mapping for grain filling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai × Yu8679. Theor Appl Genet 118:313–325. doi:10.1007/s00122-008-0901-5
Wang L, Li G, Peña RJ, Xia X, He Z (2010) Identification of novel allelic variants at Glu-A3 locus and development of STS markers in common wheat (Triticum aestivum L.). J Cereal Sci 51:305–312. doi:10.1016/j.jcs.2010.01.005
Wang J, Lin W, Wang H, Li L, Wu J, Yan X, Li X, Gao A (2011) QTL mapping of yield-related traits in the wheat germplasm 3228. Euphytica 177:277–292. doi:10.1007/s10681-010-0267-z
Zanetti S, Winzeler M, Feuillet C, Keller B, Messmer M (2001) Genetic analysis of bread-making quality in wheat and spelt. Plant Breed 120:13–19. doi:10.1046/j.1439-0523.2001.00552.x
Zhang W, Chao S, Manthey F, Chicaiza O, Brevis JC, Echenique V, Dubcovsky J (2008) QTL analysis of pasta quality using a composite microsatellite and SNP map of durum wheat. Theor Appl Genet 117:1361–1377. doi:10.1007/s00122-008-0869-1
Zhang J, Dell B, Biddulph B, Drake-Brockman F, Walker E, Khan N, Wong D, Hayden M, Appels R (2013) Wild-type alleles of Rht-B1 and Rht-D1 as independent determinants of thousand-grain weight and kernel number per spike in wheat. Mol Breed 32:771–783. doi:10.1007/s11032-013-9905-1
Zhang G, Wang Y, Guo Y, Zhao Y, Kong F, Li S (2015) Characterization and mapping of QTLs on chromosome 2D for grain size and yield traits using a mutant line induced by EMS in wheat. Crop J 3:135–144. doi:10.1016/j.cj.2014.11.002
Zheng BS, Le Gouis J, Leflon M, Rong WY, Laperche A, Brancourt-Hulmel M (2010) Using probe genotypes to dissect QTL × environment interactions for grain yield components in winter wheat. Theor Appl Genet 121(8):1501–1517. doi:10.1007/s00122-010-1406-6
Acknowledgments
This research was supported by grants from the National Natural Science Foundation of China (No. 31471573), the National Basic Research Program of China (2014CB138100), the Chinese Academy of Sciences (No. XDA08030107) and the Ministry of Agriculture of China (No. CARS-03-03B).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical standards
All of the authors have read and have abided by the statement of ethical standards for manuscripts submitted to Theoretical and Applied Genetics.
Additional information
Communicated by I. Mackay.
F. Cui, X. Fan, and M. Chen contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Cui, F., Fan, X., Chen, M. et al. QTL detection for wheat kernel size and quality and the responses of these traits to low nitrogen stress. Theor Appl Genet 129, 469–484 (2016). https://doi.org/10.1007/s00122-015-2641-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-015-2641-7