
Abstract
Plant hormones regulate multiple physiological and metabolic systems through different signaling channels. The complex signaling network and metabolic processes play a major role in plant growth and responses to various environmental stresses. Extensive studies have unveiled most of the members of plant hormones and elucidate their principal effects on plant cell systems. Brassinosteroids (BRs) and ethylene are the two major biomolecules playing adorable roles in plant growth, physiological processes, and stress responses. Their collective interaction with each other and physiological parameters harmonize the important functions at different stages of plant growth and development. They also play a major role in biotic and abiotic stresses. This study examined the interrelation of ethylene and BRs during different developmental stages. It also highlights the two hormones’ role during fruit ripening, stomatal closure, reproduction, abiotic stresses, and biotic stresses. The BRs and ethylene possess an antagonistic influence on the expansin gene AtEXPA5 expression. That antagonistic interrelation is responsible for the hook formation during the gravitropic growth of hypocotyls. The ethylene and BR cross talk comprises a complex network of signaling pathways, e.g., the ACC synthase pathway. Phytotoxins positively interact with ethylene pushing the plant into more stressed conditions. In this study, we have accounted both the hormones together to understand the plant responses better. This will help in providing knowledge of different interacting processes involved in these hormones. The cross talks of important plant hormones, such as BRs and ethylene, will provide us remarkable proficiency to induce stress resistance and enhance plant productivity.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Plant hormones or phytohormones are small, naturally occurring organic molecules important in plant growth, development, cellular mechanisms, physiological processes, and specific molecular activities (Akhtar et al., 2020). Classic methodologies comprising biochemistry, genetics, and physiological studies have contributed to the progress of plant hormones. These studies have identified important functions of these plant hormones in the development, growth, and subsequent plant responses to numerous abiotic and biotic stresses (Jiang & Asami, 2018). Nine classes of the phytohormones have been discovered, including gibberellins, salicylic acid, abscisic acid, cytokinins, auxins, ethylene, jasmonic acid, brassinosteroids, and strigolactones (Wang & Irving, 2011).
Brassinosteroids (BRs) are organic steroidal polyhydroxylated plant hormones that play a very diverse role in different aspects of plant growth and developmental processes (Khan et al., 2019). BRs were initially defined depending on their growth-stimulating mechanisms. Recent cellular, molecular, and physiological studies have uncovered its role in senescence, pollen development, photosynthetic performance, stem elongation, plant development, seed germination, and responses to several stresses, including extreme temperatures (Nolan et al., 2020). The development of advanced approaches plays a crucial role in providing an in-depth understanding of molecular and physiological mechanisms important in BR degradation, signaling and biosynthesis cascades, and related pathways (Ahmad et al., 2014a). Recent studies have reported BRs to positively impact the plant response to specific abiotic stresses and environmental factors, including salinity, heavy metals, heat, drought, pesticides, temperature, and cold (Khan et al., 2017a). Conversely, the definite mechanisms involved in BR signaling, which stimulate stress tolerance, are still unclear (Vardhini & Anjum, 2015).
Recent studies have revealed the interaction of BRs with other hormones, including jasmonic acid, auxin, ethylene, abscisic acid, cytokinin, salicylic acid, and gibberellin (Bashir et al., 2016; Hossain et al., 2006; Wu et al., 2019). These interactions affect the developmental, cellular, and physiological mechanisms of plants. Deficiency in BR biosynthesis can result in abnormal developmental phenotypes, highlighting the prominence of various signaling pathways. It also highlights the importance of BR biosynthesis, concentrations, and activities in regulating the cellular mechanisms (Saini et al., 2015; Ahmad et al., 2021a).
Ethylene, the first known plant hormone, is an aging hormone involved in regulating different characteristics of a plant life cycle (Yasin et al., 2018a; Iqbal et al., 2017). The fruit ripening, germination of seed, organ longevity, root initiation, senescence, abscission, fruit ripening, root hair development, flower development, and responses toward external stressors mainly base upon ethylene biosynthesis (Schaller, 2012). It also controls various responses of the environment which are directly influencing reproduction in plants. Recently some significant advancement in understanding molecular and biochemical mechanisms involved in ethylene action and synthesis regulation has been reported (Lin et al., 2009). The ethylene hormone level changes due to environmental conditions are directly and indirectly involved in the plants’ regulating lifecycle, making ethylene cross talk a major subject of interest (Iqbal et al., 2017). This analysis provides an inclusive overview of the connection between the role of BRs and ethylene in plant growth and development and the impact of biotic and biotic stress.
Root Growth
Roots are the essential plant organs responsible for structural anchorage; absorption of water and nutrients for the survival of plant by controlling its growth and development. They are also involved in the interaction with soil-living biota and serve as a symbiotic interaction site for soil-living microorganisms (Grierson et al., 2014). Root hairs and their root epidermal cells tubular extensions assist or increase subsequent functions by significantly increasing the absorptive surface. The development of root hair is persistently adjusted to alteration in the surrounding of roots, ultimately allowing root function optimizations in the soil environment (Ibrahim et al., 2017; Li et al., 2021). The interaction of plant hormones with other hormones contributes to various growth mechanisms in the plant roots. Furthermore, some important signal molecules, including reactive oxygen species (ROS), are also involved in root development (Swanson & Gilroy, 2010). The interconnection between hormone signaling and root hair signaling mechanisms with different biotic and abiotic alterations subsequently in the rhizosphere facilitates vibrant hormone-stimulated alterations in root hair growth, density, length, and morphology (Vissenberg et al., 2020).
The investigation of the connection between ethylene, BRs, and ROS has been reported. In this study, the screening of EMS mutant was carried out to identify Arabidopsis mutant (det2-9) with deficiency of BR synthesis, which subsequently depended on the short root phenotype. Meanwhile, the ethylene and ROS signaling cascade were increased in the Arabidopsis det2-9 mutant. It was proposed that the short root phenotype was the ultimate result of ethylene and superoxide anion (O2−) accumulation. The exogenous BR application indicated that the ethylene biosynthesis regulation was carried out depending on its given concentration. The ethylene production was significantly decreased in the seedlings, which were treated with a low concentration (10–100 nM) of 24-epibrassinolide (EBL). On the other hand, the one treated with higher EBL concentrations ≥500 nM displayed a sharp increase (Lv et al., 2018; Shah et al., 2020).
Consequently, low concentrations of BRs result in the inhibition of ethylene response factor (ERF) expression. In contrast, when the concentration is high, it increases ERF expression, which is consistent with ethylene results after BR treatment. The study was carried out to evaluate the connection between 1-aminocyclopropane-1-carboxylic acid synthases (ACSs) enzymes by certain brassinosteroid-regulated transcription factors (BES1 and BZR1), and their role in ethylene biosynthesis was further confirmed by qRT-PCR. This study revealed that ethylene biosynthesis repression was carried out by certain transcription factors and was ultimately controlled by BR regulation. It was concluded that a high level of BRs resulted in increased production of ethylene by stimulating ACS enzymes (Lv et al., 2018). The directional growth regulation is necessary for the proper growth and development of roots and longitudinal growth (Tariq et al., 2020; Ahmad & Ashraf, 2016). Studies also proposed numerous environmental signals and factors that can further stimulate that plant root elongation and gravitropism. Previous studies showed that induced glucose stimulates root growth of the seedling, and when BRs are applied, it further increases this modulation type. Thus, the results proved that glucose increased the BR signaling by modulating BRI1 endocytosis from cell membrane to early endosomes (Singh et al., 2014a).
Singh and coworkers also evaluated the interaction of plant hormones and glucose in controlling root growth. The results suggested that the presence of cytokinins and ethylene could eradicate root growth when glucose or BRs were regulated. In this case, ethylene and cytokinins act antagonistically with BRs for subsequent growth regulation. Cytokinin pathway follows the BR signaling, which ultimately antagonizes the roots’ directional growth by using ethylene-stimulated machinery (Singh et al., 2014b).
Shoot Growth and Apical Hook Development
The interaction between different plant hormones results in cell elongation, which is involved in shoot growth in plants. BRs are considered important hormone which promotes the activity of cell elongation. The experiments conducted observed that when the AtRALF1 gene is partially silenced, the AtEXPA5 expansin gene involved in cell expansion expression was increased. The exogenous application of BRs results in an induced AtEXPA5 level. It illuminates an antagonistic effect between BR and AtRALF1 for expansin genes. Ethylene reduces the expression of AtEXPA5 and regulates hypocotyl growth. The results from different experiments also suggested that the interaction of ethylene and AtRALF1 could achieve the same effect (Bergonci et al., 2014).
Many studies concluded that BRs and ethylene affect hypocotyl development in plants (Hoque et al., 2016; Shafique et al., 2014). The research included mutant Arabidopsis plant screening and identification with an improved response to acsinone7303. The acsinone7303 performs as an inhibitor for ACS enzymes. Numerous mutants of ret. with decreased sensitivity to acsinone7303 were also investigated. Furthermore, ret41 and ret8 were characterized. The Map-based cloning results concluded that ret8 depicts a mutation in CESA6/cellulose synthase six, while ret4 represents a mutation in de-etiolated-2 (DET2). The enzyme DET2 catalyzed the campesterol to campesterol reduction process within the BR biosynthesis pathway. Whereas, CESA6 was a major part of the primary wall CESA complex (Verma et al., 2007).
Another study suggested that the mutant seedlings had short roots and hypocotyls when the mutation of eto1 was removed. That showed that the increased ethylene level did not completely affect the hypocotyl phenotype. Moreover, it was observed that the inhibitors of ethylene biosynthesis did not completely decrease the response of cesa6ret8 and det2ret41 mutants. This further suggested that mutations in DET2 and CESA6 cause short hypocotyls in mutants of cesa6ret8 and det2ret41, respectively. They play a very important role in the growth and development of seedlings in plants. The ethylene-induced level in eto1 stimulated the plant short hypocotyl phenotype in det2 and cesa6. Numerous experimental studies with subsequent EBL eto1, det2ret41, and det2-1 treatment indicated that ethylene and BR level balance is significantly important in hypocotyl growth accurate regulation (Chen et al., 2013).
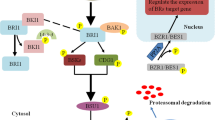
The growth and development of the apical hook are very important for plant growth. This growth and development are followed by seed germination in plants (Bashir et al., 2013; Ahmad et al., 2021b). The early stages of Arabidopsis hypocotyl development include apical hook development, which plays an important part in protecting the apical meristem cotyledons of the shoot as the seedling growth takes place in the soil. The hook development stages include hook maintenance, the hook formation, and most importantly, the hook opening. Previous studies have reported the role of ethylene in the apical hook development phase, where BRs activate the maintenance phase, which further delays the hook opening phase. These stages of hook development are strictly controlled by a complex network of different hormones (Mazzella et al., 2014). Various experiments validated the results from these studies to investigate the BR biosynthesis role and specific signaling mutants for ethylene (Fig. 7.1).

Ethylene interaction with brassinosteroids (BRs) under light and dark conditions
Furthermore, the interactions have been demonstrated on the growth and development of the plants. The arrows show positive interaction, while blunt head arrows show a negative interaction
Flowering
Flower formation and development are the most important phases in plant development, directly impacting plant reproduction and production. One of the plant families, called Cucurbitaceae, is known for its sex expression phenotypic variety (Abbas et al., 2020; Shafaghat, 2011). The development of plants in this family includes early male flower production followed by female flower production. Another study evaluated the role of BRs in cucurbit sex expression regulation. The experimental plant models included three different species, such as zucchini, cucumber, and melon. The cucumber plants were treated with BRs, and female buds and female flowers were observed. The ethylene level was induced simultaneously, which further concluded that the BR effect was ethylene mediated. The melon and zucchini plants also displayed the same induced level of ethylene production shown by the cucumber. Still, the induced level of femaleness was not present in melon and zucchini plants after BR treatment. This study concluded that BR-treated cucumber plants produced more ethylene than control cucumber plants. Hence, the effect of BRs on sexual expression in the cucumber plant is usually facilitated by the ethylene hormone. Different plant species have different sensitivity to ethylene. Therefore, the mechanism of interaction between ethylene and BRs in flower development BRs was found to act indirectly through ethylene increase production and increasing femaleness rate depending on specific species sensitivity to hormone ethylene (Papadopoulou & Grumet, 2005).
Ethylene and BRs mediate the sexual expression of different species of plants. Ethylene plays a vital role in regulating sex expression in plants, especially in the Cucurbitaceae family (Naemi et al., 2014; Hashemi et al., 2019). Its level in the buds of flowers initiates female flower development in plants. When plants were treated with ethylene biosynthesis inhibitors, such as aminoethoxyvinylglycine (AVG) or silver thiosulphate (STS), an increase in male flower development was observed. Another study compared the sensitivity of different genotypes to hormones BRs and ethylene by comprehensively working on different BRs and ethylene treatments exclusively on flower development and sex expression of different Cucurbita pepo genotypes, including Vegetable Spaghetti (Veg) and Bolognese (Bog). The experimental results from this study indicated that the effect of ethylene is greater as compared to BRs on flower development and sex expression in C. pepo. The ethylene stimulates the female flower development and decreases the formation of male flower development. The use of ethylene inhibitors like AVG and STS decreases female flower development and, on the other hand, increases male phase development. The Bog genotype produces more ethylene, and they were more responsive to ethylene inhibitors like AVG and STS, resulting in a decreasing number of female flowers. The other Veg genotype showed lower ethylene production, reduced the male flower development, and increased female flower production by responding better to ethephon. Results showed that the development of male or female flowers was not altered or affected by the treatment of brassinazole in C. pepo. This showed that BRs play a significant role in ethylene production regulation. It also partially affects sexual expression and flower development in C. pepo and is directly involved in male and female flower development (Manzano et al., 2011).
Ripening and Postharvest Development of Fruit
Ripening of the fruits is a complex event, in which multiple phytohormones coordinate together, for normal growth, fertilization, and morphogenesis. The final fruit development involves four different stages: fruit set, fruit development, fruit maturation, or ripening phase. The last phase in plant development is fruit ripening, which plays a crucial role in making fruit attractive, edible, nutritional, and valuable agricultural commodities. This ripening process also includes physiological, cellular, and biochemical alterations such as cell wall structure modification, increased flavors and aroma, starch to sugar conversion, and changes in pigment biosynthesis (Kumar et al., 2014; McAtee et al., 2013).
The fruit ripening is classified in climacteric and non-climacteric fruit ripening depending upon its respiration and ethylene biosynthesis levels. The climacteric fruits are also called as ethylene-dependent fruits. These types of fruits can ripen once they are harvested with the ethylene production. The climacteric fruits include avocado, tomatoes, bananas, cucurbits, and apples. They are accompanied by a dramatic increase in ethylene production and respiration during their ripening process (Kumar et al., 2014; Cherian et al., 2014; Azzi et al., 2015). The non-climacteric fruits cannot ripen once they are removed from the parent plant, and ethylene is not required for their ripening. These fruits include citrus, strawberry, raspberry, and grapes (Kumar et al., 2014; Cherian et al., 2014).
BRs, a new class of plant hormones, are involved in plant growth and development and regulate ethylene production. In plant vegetative tissues, the BR exogenous application is involved in induced ethylene production and thus stimulates ethylene-mediated growth response. BRs and ethylene act together and collectively control plant metabolism (Zaheer et al., 2017; Yasin et al., 2018b). Moreover, studies have reported that BRs and ethylene hormones have antagonistic effects in the Arabidopsis (Deslauriers & Larsen, 2010). In fruits like strawberry and mango, the application of endogenous BRs is present in small amounts and may not be important for fruit ripening.
In some cases, ethylene production takes place without variations of low BR levels in ripened fruit. While in other cases, the applied BRs stimulate ethylene production, signifying that ethylene production can be independent of BR (Zaharah et al., 2012; Greco et al., 2012). The role of ethylene in regulating climacteric fruit ripening, such as mango, is well-known, and numerous studies have been carried out to reveal further the mechanisms involved in it (Müller & Stummann, 2003). To further understand the role of plant hormone in regulating fruit ripening, the endogenous levels of ethylene and BRs were investigated in mango fruit ripening. The study also evaluated the effect of exogenous application of BRs and ethylene on fruit ripening. The results from this study suggested that BR endogenous level may not display an important role in the ripening of climacteric mango fruit (Zaharah et al., 2012). Recent studies have also highlighted the role of BRs in non-climacteric fruit grapes ripening (Symons et al., 2006). The exogenous BR application in climacteric fruit like tomato is involved in promoting tomato pericarp disc ripening and increased ethylene production. The endogenous BR high concentration in tomato fruit was also reported during early developmental stages. The BR-induced fruit ripening was interconnected with ethylene-increased production. The results also suggested the ability of BRs to stimulate fruit ripening and fruit senescence (Montoya et al., 2005).
Numerous studies evaluated the effects of BRs on postharvest development and fruit ripening. A recent study investigated the effects of BRs and ethylene on non-climacteric fruit ripening. In this study, strawberries were used as a study model for non-climacteric fruits. The treatment of exogenous spray of ethylene and EBL was done on this study model. The experimental results of the study showed that ethylene and BRs influence the levels of phenolic compounds in plants. The treatment of ethylene increases the level of phenolic compounds while the BR application results in reducing the level of phenolic compounds. The ethylene treatment results in high levels of phenolic compounds that result in senescence. When BR application reduces the level of the phenolic compound, the induced antioxidant activity helps in the stimulation of fruit conservation (Ayub et al., 2018).
Another study highlighted the role of ethylene and BRs in fruit ripening and postharvest development. In this study, BRs were dynamically produced during fruit ripening in the tomato plant. The transgenic lines overexpressing or silencing SlCYP90B3 were further generated. The accumulation of carotenoids and ethylene production were strongly linked with SlCYP90B3 level by the alteration in gene expression of carotenoid biosynthetic and ethylene pathway genes. The results suggested that the SlCYP90B3 gene is involved in BR biosynthesis and fruit ripening in tomato plants, making it a gene of interest for the improvement of nutritional, visual, texture, and flavor qualities of tomato fruits (Hu et al., 2020).
BRs affect ethylene biosynthesis primarily by regulating ACC-synthase enzyme (ACS) and ACC-oxidase component activities (Ul Haq et al., 2020; Hafeez et al., 2019). The high BR levels induce ethylene biosynthesis by increasing the ACS protein stability, while the low levels of BRs decrease ethylene biosynthesis by the activity of high expression of BZR1/BES1. These transcription factors play a major role in the BR signaling pathway and inhibit the transcription of ACS genes (Lv et al., 2018). Studies suggested that exogenous application of BRs can stimulate fruit ripening in bananas due to increased MaACO14, MaACS1, and MaACO13 expression. The exogenous application of BRs can stimulate and induce postharvest ripening, increasing the development of quality characteristics and subsequently increasing ethylene production in tomato by increasing ACS2 and ACS4 gene transcriptional levels.
Stress Response
The BRs and ethylene plant hormones are involved in plant growth and development and play a diverse role in plant responses to biotic and abiotic stress responses (Yasin et al., 2018a; Fariduddin et al., 2014; Ahmad et al., 2020a).
Abiotic Stresses
BRs, the natural steroid plant hormones, play a diverse role in plant growth and developmental mechanisms such as cell division, reproductive development, cell elongation, vascular differentiation, and response to abiotic stresses or tolerance. BRs play a significant role in decreasing abiotic and biotic stresses at different levels (Khan et al., 2017a; Ahmad et al., 2021b; Yasin et al., 2017). Abiotic stress factors adversely affect the plant growth, fruit yield, and agricultural productivity in plants. They interrupt the physiology and morphology of plants by different metabolic changes. This results in reducing plant growth by causing cell injury (Parvin et al., 2015). Salt affects more than 20% of cultivated land worldwide, increasing day by day, hampering crop productivity (Flowers, 2004). Plants have well-developed defense systems, including biochemical and physiological processes for protection against abiotic stress-induced injuries, including osmoregulation, antioxidant responses, and homeostasis. The plant responds to stress by stimulating antioxidant systems. These antioxidant systems can be enzymatic or non-enzymatic. The enzymatic antioxidant system includes catalase, peroxidases, superoxide dismutase, and glutathione reductase. Whereas, the non-enzymatic antioxidant system comprises carotenoids, vitamins C, vitamin E, flavonoids, and phenolic compounds. Among these, the phenolic compounds play an important role as the most dominant antioxidants (Yousaf et al., 2015; Ahmad et al., 2014b, 2020b). The study evaluated the BR effects on abiotic stress resistance in cucumber against polyethylene glycol (PEG), cold, and salt.
Previous studies have reported that BRs can increase ethylene production and induce the alternative oxidase (AOX) pathway. Results showed that the transcription levels of ethylene-mediated biosynthesis genes such as 1-aminocyclopropane-1-carboxylate oxidase2 (CSACO2), ripening-related ACC synthase1 (CSACS1), CSAOX, ripening-related ACC synthase2 (CSACS2), 1-aminocyclopropane-1-carboxylate oxidase1 (CSACO1), and ACC synthase3 (CSACS3) were enhanced after BR treatment. Furthermore, salicylhydroxamic acid (SHAM, AOX inhibitor) and an inhibitor of ethylene biosynthesis like aminooxyacetic acid (AOA) application reduced plant tolerance to different environmental stresses and factors. This process is accomplished by blocking respiration or cellular process, which is induced by BRs. This study concluded the role of ethylene in BR-induced AOX activity, which is involved in abiotic stress resistance (Wei et al., 2015).
The transpiration rate depends on the opening and closing of stomata in plants, and stomata play a significant role in protecting the plant against stress conditions like water stress and pathogens. The stomatal movement pattern depends on different reversible alterations, including turgor pressure and water stomata flow in stomata. This stage is induced by many exogenous and endogenous stimuli. Therefore, the analysis of the opening and closing of stomata mechanism is essential to understand how the plants protect themselves against water and pathogens stress (Roelfsema & Hedrich, 2005). The opening and closing of stomata are regulated by different plant hormones involved in a complex signaling pathway network. Previously the most linked plant hormone for stomatal closure was abscisic acid only, but recent studies have suggested that BRs and ethylene affect the stomata activity (Shi et al., 2015).
Another study based on the interaction of ethylene and BRs in plants for salt stress highlighted different mechanisms. In this study, the BRs that induce salt tolerance in tomato plants were examined. In this study, the induce levels of ethylene and H2O2 in brassinolide-treated tomato seedlings were investigated. Results revealed that H2O2 and ethylene are intricate in BR-induced stress tolerance, and both BRs and ethylene could stimulate H2O2 production (Zhu et al., 2016).
The salt stress adversely affects the plant by reducing its leaf area, root and shoot length, membrane stability, accumulation of dry matter, relative water content, root weight, and reducing carbon dioxide assimilation, ultimately affecting plants’ fruit production. Calcium acts as a second messenger and plays an important role in intervening mechanisms induced in response to different abiotic stresses in plants (Kader & Lindberg, 2010). It enhanced the growth of salt stress in plant and its subsequent signaling which control ion homeostasis pathways. Calcium ions restrict the entry of sodium ions in plant cells under salt stress conditions (Hussain et al., 2010). The most harmful effect of salinity stress is the accumulation of Na+ and Cl− ions in the plant tissues, which are highly exposed to soil with a high concentration of NaCl. When these ions enter the cell, it results in a severe ionic imbalance, which causes important physiological disorders in plants. The increased amount of calcium increases the growth and germination of the salt-stressed plant (Fig. 7.2).

Ethylene interaction with brassinosteroids (BRs) and abiotic stressors (i.e., salinity, drought, chilling, and oxidative stress)
Arrows show positive interaction, while blunt head arrows show negative interaction
Salt stress, the most adverse stress among abiotic stresses, reduces the oxidative stress, ion toxicity, and water unattainability apart from obstructing plant growth and productivity. Different other activities are involved, which ultimately leads to minimizing the plant productivity and growth (Parvin et al., 2015). The stress conditions in plants can result in oxidative damage. Therefore, the cells of plants require a sophisticated and delicate central antioxidant system. Glutathione (GSH) and ascorbic acid (AA) connection plays a significant role in this antioxidant system, which ultimately protects these plants against different oxidative damages. Ascorbic acid has various physiological functions such as photosynthesis regulation and increased cell growth in plants (Yasin et al., 2018b; Locato et al., 2013).
Different abiotic conditions, including drought or salinity, affect the symbioses relationship between plants and microorganisms. This symbioses relationship is important in the uptake of essential nutrients in plants. Studies reported ethylene signaling mutant of pea Psein2 and examined the interaction between BRs and ethylene and its effect on mycorrhizal development (Weller et al., 2015; Khan et al., 2017b). Rice is the essential crop worldwide, and different mechanisms to induce plant resistance to stress have been discovered. The study identified a gene, OsSta2, which expression induces oxidative and salt stress tolerance in rice crops. The results from this study suggested that OsSta2 gene plays a key role in the complex network of the ABA signaling pathway throughout the stress response (Kumar et al., 2017). The stress response mechanism and molecular breeding can be better understood by the genome-wide studies for different gene identification and their role in stress responses. These studies revealed that AP2/EREBP gene families were identified and classified in the Cucurbitaceae species. These families of genes play crucial roles in controlling different environmental stresses (Lee et al., 2017).
Biotic Stresses
Biotic stress is also one of the important constraints in plant productivity. Plants suffer from different stress conditions, which can be biotic or abiotic factors. The stress factors in plants can be abiotic like temperature or drought and biotic like pathogens and different pests like nematodes, insects, and fungi. Biotic stress is when there is damage to plants from a living organism such as parasites, bacteria, fungi, viruses, and harmful as well as beneficial insects. The defensive system of plants provides resistance to these biotic and abiotic factors. The defensive mechanism of plants includes physical or chemical barriers and functions effectively to decrease the harmful impact of biotic factors. These defensive mechanisms also involve complex pathways of complex phytohormones, including BRs, ABA, and ethylene (Ahmad et al., 2014c; Akram et al., 2014).
The involvement of these biotic and agrochemical factors is important if there is no genetically based resistance to confirm high productivity. Plants can develop morphological and physiological adaptations to survive in harsh environments. To grow in high salt stress, halophytes can excrete extra salt with the help of their secretory glands (Zaheer et al., 2017; Anjum et al., 2017; Akram et al., 2013). A previous study worked to understand the tomato plant responses like genetic control and signaling pathways to abiotic and biotic stress, including salinity and pathogen stresses (Bai et al., 2018). The research revealed that application of BR at low concentration improves the plant growth, quality, and production and induces resistance to different fungal and viral pathogens in various plants such as tomato, tobacco, and cucumber (Wang et al., 2015).
A recent study showed that in the fungal disease of cedar-apple rust, the expression levels were increased for flavonoid compounds (e.g., anthocyanin and catechin), and MYB genes (MYB30), specifically in the fungus-infected tissues. The study also suggested that plant hormones, including SA, ABA, JA, BR, and ETH, were found to be highest in infected plants of apple (Bashir et al., 2016; Lu et al., 2017). In the study on BR-treated pepper plant exposed to cold stress, the plant hormones such as SA, ETH, and JA levels were found to be significantly increased (Li et al., 2016; Ahmad et al., 2013). The experimental work results suggested that BR functions by interacting with SA, JA, and ETH signaling hormones, especially for cold stress response. This further highlights that BRs play a crucial role in response to biotic stress tolerance by activating transcriptional factors, enzymes, hormones, biotic resistance genes, antioxidants, and signaling pathways to reduce biotic stresses of plants.
Ethylene and Pathogenesis
Ethylene biosynthesis has been reported at accelerated rates during the progressive events of pathogenicity. In this process, there is little or no discrimination of the pathogen type (e.g., bacteria, fungi, viruses, or nematodes) or the pathogenic species. Ross and Williamson first highlighted the topic during 1951 by recording the elevated ethylene contents in virus-infected plants (Ross & Williamson, 1951). The enhanced ethylene contents fall under the early biochemical communications of plant cells with the other cells in the vicinity. Generally, it is associated with the cell necrosis leaving localized lesions on the plant surfaces. Bacterial pathogens have been well investigated for the boosted ethylene contents and the characteristic lesion development. Viral pathogens also adopt the same pattern as the bacterial pathogens, but their own characteristic symptoms. Ethylene biosynthesis is increased with the viral disease progress. Fungal pathogens also drive plant cells toward an ethylene peak formation, while the height of the peak correlates with the amount of tissue damage (Ahmad et al., 2019, 2020c; Ahmed et al., 2017).
Ethylene Biosynthesis During Infections
Ethylene production does not require physical damage by the pathogens, but it is also elicited due to the pathogen-origin elicitors. The physical invasions of microbes are the secondary factors leading to the lesion formations, if detected by the hypersensitive defense systems (Khan et al., 2018). Pathogen elicitors that are difficult to be detected by the plant defense machinery cause a delayed excitation of the ethylene biosynthesis. Thus, it leads to much more damages to the photosynthetic and physiological systems of the cells. However, an interesting fact about the hormone was revealed to the researchers when some pathogenic bacteria and fungi produced ethylene by themselves under in vitro conditions. However, their ability to produce ethylene is of more assistance to trigger ACC synthase than the elicitation of the plant defense cascade. This abrupt excitation of the ACC synthase causes the stunted growth of plants, a characteristic feature of the biotic stress. Therefore, ethylene synthesis in plants is the best and the most optimized measure to alleviate plant biotic stress.
Regulation of Ethylene Under Post-Infection Conditions
A MET-ACC-independent pathway has been extensively reported for ethylene biosynthesis by the host plant during progressing disease establishment. The poor incorporation of radioactive MET derived the conclusion into plant-produced ethylene. The results proved that MET was not associated with the main C2H4 biosynthetic pathway in the infected plant tissues. Furthermore, the conclusion was supported by AVG application, a MET inhibitor, which could not reduce ethylene production. Another MET inhibitor Co+2 failed to inhibit or reduce ethylene biosynthesis in infected tissues. Similarly, ACC application, an intermediate of the MET-ACC pathway, could not enhance ethylene contents. All these facts concluded that ethylene production was involved in a mechanism other than the MET-ACC-dependent pathway.
Ethylene and Disease Spread
It is very hard to draw a generalized and precise relationship between the ethylene production and disease development. The role of ethylene during the infection process becomes more complex when it interacts with other growth hormones (e.g., auxins), pathogen-derived toxins, and arthropods associated herbivory. However, in a broader area, an interconnection between the host-derived ethylene and disease development can only be made by ignoring the other factors, that is, the negative interrelation. However, in some cases, ethylene inhibition caused a significant reduction in the disease development. On the other hand, the exogenous application of ethylene has been proved a useless strategy for plant protection programs because it promoted the disease development rather than to control the pathogen. Pathogen-derived ethylene doubles symptom severity if compared with non-ethylene-producing pathogen strains.
Ethylene Interrelation with Toxins
Toxins are classified among the plant stressors promoting plant diseases. Several phytotoxins have been reported negatively impacting plant health, causing diseases, and deteriorating the edible quality of plants. Toxins are also directly related to ethylene, which concomitantly reduces plant growth by the ACC synthase pathway. Fusicoccum amygdali is famous for fusicoccin production, which is responsible for developing disease symptoms on almond and peach. It stimulates the conversion of ACC to ethylene. Another example of the phytotoxin interacting with ethylene is coronatine produced by Pseudomonas syringae. The toxin bears the tendency to increase the ethylene release from the different plants. Similarly, Pseudomonas phaseolicola produces a toxin named phaseolotoxin in addition to the production of ethylene. This joint production of both the stressors is lethal for the plants and proves a supporting effect of toxins to the ethylene production and downstream biotic stress responses (Fig. 7.3).

Ethylene interaction with brassinosteroids (BRs), pathogens, and phytotoxins
The interaction leads to some physical outcome in the plant’s stature. Black arrows show positive interaction, while blunt head arrows show negative interaction. Red arrows show an abrupt elevation in the contents due to some external stimuli/applications. Reception signals of pathogen or phytotoxins have been shown with dotted arrows
References
Abbas, H. M. K., et al. (2020). Metabolic and transcriptomic analysis of two Cucurbita moschata germplasms throughout fruit development. BMC Genomics, 21, 365.
Ahmad, A., & Ashraf, Y. (2016). In vitro and in vivo management of Alternaria leaf spot of Brassica campestris L. Journal of Plant Pathology & Microbiology, 7, 1000365.
Ahmad, A., Shafique, S., & Shafique, S. (2013). Cytological and physiological basis for tomato varietal resistance against Alternaria alternata. Journal of the Science of Food and Agriculture, 93, 2315–2322.
Ahmad, A., Shafique, S., & Shafique, S. (2014a). Intracellular interactions involved in induced systemic resistance in tomato. Scientia Horticulturae (Amsterdam), 176(127–133).
Ahmad, A., Shafique, S., Shafique, S., & Akram, W. (2014b). Penicillium oxalicum directed systemic resistance in tomato against Alternaria alternata. Acta Physiologiae Plantarum, 36, 1231–1240.
Ahmad, A., Shafique, S., & Shafique, S. (2014c). Molecular basis of antifungal resistance in tomato varieties. Pakistan Journal of Agricultural Sciences, 51, 683–687.
Ahmad, A., et al. (2019). Benzenedicarboxylic acid upregulates O48814 and Q9FJQ8 for improved nutritional contents of tomato and low risk of fungal attack. Journal of the Science of Food and Agriculture, 99, 6139–6154.
Ahmad, A., et al. (2020a). Dopamine alleviates hydrocarbon stress in Brassica oleracea through modulation of physio-biochemical attributes and antioxidant defense systems. Chemosphere, 128633. https://doi.org/10.1016/j.chemosphere.2020.128633
Ahmad, A., et al. (2020b). Metabolic and proteomic perspectives of augmentation of nutritional contents and plant defense in Vigna unguiculata. Biomolecules, 10, 224.
Ahmad, A., et al. (2020c). First report of Fusarium nelsonii causing early-stage fruit blight of cucumber in Guangzhou, China. Plant Disease, 104, 1542.
Ahmad, A., et al. (2021a). Functional and structural analysis of a novel acyltransferase from pathogenic Phytophthora melonis. ACS Omega. https://doi.org/10.1021/acsomega.0c03186
Ahmad, A., et al. (2021b). Synergistic effects of nitric oxide and silicon on promoting plant growth, oxidative stress tolerance and reduction of arsenic uptake in Brassica juncea. Chemosphere, 262, 128384.
Ahmed, S., et al. (2017). Characterization of anti-bacterial compounds from the seed coat of Chinese windmill palm tree (Trachycarpus fortunei). Frontiers in Microbiology, 8, 1894.
Akhtar, S. S., Mekureyaw, M. F., Pandey, C., & Roitsch, T. (2020). Role of cytokinins for interactions of plants with microbial pathogens and pest insects. Frontiers in Plant Science, 10, 1777.
Akram, W., Anjum, T., Ali, B., & Ahmad, A. (2013). Screening of native bacillus strains to induce systemic resistance in tomato plants against fusarium wilt in split root system and its field applications. International Journal of Agriculture and Biology, 15, 1289.
Akram, W., Anjum, T., Ahmad, A., & Moeen, R. (2014). First report of Curvularia lunata causing leaf spots on Sorghum bicolor from Pakistan. Plant Disease, 98, 1007.
Anjum, T., Akram, W., Shafique, S., Sahfique, S., & Ahmad, A. (2017). Metabolomic analysis identifies synergistic role of hormones biosynthesis and phenylpropenoid pathways during fusarium wilt resistance in tomato plants. International Journal of Agriculture and Biology, 19, 1073–1078.
Ayub, R. A., Reis, L., Lopes, P. Z., & Bosetto, L. (2018). Ethylene and brassinosteroid effect on strawberry ripening after field spray Efeito do etileno e do brassinoestereoide no amadurecimento do morango após aplicação no campo. Revista Brasileira de Fruticultura, 40, 1–6.
Azzi, L., et al. (2015). Fruit growth-related genes in tomato. Journal of Experimental Botany, 66, 1075–7086.
Bai, Y., Kissoudis, C., Yan, Z., Visser, R. G. F., & van der Linden, G. (2018). Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. The Plant Journal, 93, 781–793.
Bashir, Z., et al. (2013). Quantification of cellulose contents by transmission spectra of plant tissues. Cellulose Chemistry and Technology, 47, 509.
Bashir, Z., et al. (2016). Tomato plant proteins actively responding to fungal applications and their role in cell physiology. Frontiers in Physiology, 7, 257.
Bergonci, T., Silva-Filho, M. C., & Moura, D. S. (2014). Antagonistic relationship between AtRALF1 and brassinosteroid regulates cell expansion-related genes. Plant Signaling & Behavior, 9, 16–18.
Chen, I. J., et al. (2013). A chemical genetics approach reveals a role of brassinolide and cellulose synthase in hypocotyl elongation of etiolated Arabidopsis seedlings. Plant Science, 209, 46–57.
Cherian, S., Figueroa, C. R., & Nair, H. (2014). ‘Movers and shakers’ in the regulation of fruit ripening: A cross-dissection of climacteric versus non-climacteric fruit. Journal of Experimental Botany, 65, 4705–4722.
Deslauriers, S. D., & Larsen, P. B. (2010). FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in arabidopsis hypocotyls. Molecular Plant, 3, 626–640.
Fariduddin, Q., Yusuf, M., Ahmad, I., & Ahmad, A. (2014). Brassinosteroids and their role in response of plants to abiotic stresses. Biologia Plantarum, 58, 9–17.
Flowers, T. J. (2004). Improving crop salt tolerance. Journal of Experimental Botany, 55, 307–319.
Greco, M., Chiappetta, A., Bruno, L., & Bitonti, M. B. (2012). In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. Journal of Experimental Botany, 63, 695–709.
Grierson, C., Nielsen, E., Ketelaarc, T., & Schiefelbein, J. (2014). Root hairs. Arabidopsis Book, 12, e0172.
Hafeez, M., et al. (2019). Gossypol-induced fitness gain and increased resistance to deltamethrin in beet armyworm, Spodoptera exigua (Hübner). Pest Management Science, 75, 683–693.
Hashemi, L., Golparvar, A. R., Nasr Esfahani, M., & Golabadi, M. (2019). Correlation between cucumber genotype and resistance to damping-off disease caused by Phytophthora melonis. Biotechnology and Biotechnological Equipment, 33, 1494–1504.
Hoque, T. S., et al. (2016). Methylglyoxal: An emerging signaling molecule in plant abiotic stress responses and tolerance. Frontiers in Plant Science, 7, 1341.
Hossain, M. Z., Hossain, M. D., & Fujita, M. (2006). Induction of pumpkin glutathione S-transferases by different stresses and its possible mechanisms. Biologia Plantarum, 50, 210–218.
Hu, S., et al. (2020). Regulation of fruit ripening by the brassinosteroid biosynthetic gene SlCYP90B3 via an ethylene-dependent pathway in tomato. Horticulture Research, 7, 163.
Hussain, K., et al. (2010). What molecular mechanism is adapted by plants during salt stress tolerance? African Journal of Biotechnology, 9, 416–422.
Ibrahim, A., Shahid, A. A., & Ahmad, A. (2017). Evaluation of carrier materials to develop Bacillus subtilis formulation to control root knot nematode infection and promote agroeconomic traits in eggplant. Journal of Animal and Plant Sciences, 27, 1321–1330.
Iqbal, N., et al. (2017). Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Frontiers in Plant Science, 8, 1–19.
Jiang, K., & Asami, T. (2018). Chemical regulators of plant hormones and their applications in basic research and agriculture. Bioscience, Biotechnology, and Biochemistry, 82, 1265–1300.
Kader, M. A., & Lindberg, S. (2010). Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signaling & Behavior, 5, 233–238.
Khan, W. U., Ahmad, S. R., Yasin, N. A., Ali, A., & Ahmad, A. (2017a). Effect of Pseudomonas fluorescens RB4 and Bacillus subtilis 189 on the phytoremediation potential of Catharanthus roseus (L.) in Cu and Pb-contaminated soils. International Journal of Phytoremediation, 19, 514.
Khan, W. U., et al. (2017b). Role of Ni-tolerant Bacillus spp. and Althea rosea L. in the phytoremediation of Ni-contaminated soils. International Journal of Phytoremediation, 19, 470.
Khan, W. U., et al. (2018). Role of Burkholderia cepacia CS8 in Cd-stress alleviation and phytoremediation by Catharanthus roseus. International Journal of Phytoremediation, 20, 581.
Khan, T. A., et al. (2019). Proteomic and physiological assessment of stress sensitive and tolerant variety of tomato treated with brassinosteroids and hydrogen peroxide under low-temperature stress. Food Chemistry, 289, 500–511.
Kumar, R., Khurana, A., & Sharma, A. K. (2014). Role of plant hormones and their interplay in development and ripening of fleshy fruits. Journal of Experimental Botany, 65, 4561–4575.
Kumar, M., Choi, J., An, G., & Kim, S. R. (2017). Ectopic expression of OsSta2 enhances salt stress tolerance in rice. Frontiers in Plant Science, 8, 1–14.
Lee, S.-C., et al. (2017). Genome-wide identification of the dehydrin genes in the cucurbitaceae species. Plant Breeding and Biotechnology, 5, 282–292.
Li, J., et al. (2016). Transcriptome analysis of pepper (Capsicum annuum) revealed a role of 24-epibrassinolide in response to chilling. Frontiers in Plant Science, 7, 1–17.
Li, G., et al. (2021). Hydrogen sulfide mitigates cadmium induced toxicity in Brassica rapa by modulating physiochemical attributes, osmolyte metabolism and antioxidative machinery. Chemosphere, 263, 127999.
Lin, Z., Zhong, S., & Grierson, D. (2009). Recent advances in ethylene research. Journal of Experimental Botany, 60, 3311–3336.
Locato, V., Cimini, S., & De Gara, L. (2013). Strategies to increase vitamin C in plants: From plant defense perspective to food biofortification. Frontiers in Plant Science, 4, 1–12.
Lu, Y., et al. (2017). Flavonoid accumulation plays an important role in the rust resistance of Malus plant leaves. Frontiers in Plant Science, 8, 1–13.
Lv, B., et al. (2018). Brassinosteroids regulate root growth by controlling reactive oxygen species homeostasis and dual effect on ethylene synthesis in Arabidopsis. PLoS Genetics, 14, 1–26.
Manzano, S., et al. (2011). The role of ethylene and brassinosteroids in the control of sex expression and flower development in Cucurbita pepo. Plant Growth Regulation, 65, 213–221.
Mazzella, M. A., Casal, J. J., Muschietti, J. P., & Fox, A. R. (2014). Hormonal networks involved in apical hook development in darkness and their response to light. Frontiers in Plant Science, 5, 1–13.
McAtee, P., Karim, S., Schaffer, R., & David, K. (2013). A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Frontiers in Plant Science, 4, 1–7.
Montoya, T., et al. (2005). Patterns of Dwarf expression and brassinosteroid accumulation in tomato reveal the importance of brassinosteroid synthesis during fruit development. The Plant Journal, 42, 262–269.
Müller, R., & Stummann, B. M. (2003). Postharvest physiology | ethylene. Encyclopedia of Rose Science, 557–564. https://doi.org/10.1016/b0-12-227620-5/00064-1
Naemi, F., Asghari, G., Yousofi, H., & Yousefi, H. A. (2014). Chemical composition of essential oil and anti trichomonas activity of leaf, stem, and flower of Rheum ribes L. extracts. Avicenna Journal of Phytomedicine, 4, 191–199.
Nolan, T. M., Vukasinović, N., Liu, D., Russinova, E., & Yin, Y. (2020). Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell, 32, 298–318.
Papadopoulou, E., & Grumet, R. (2005). Brassinosteriod-induced femaleness in cucumber and relationship to ethylene production. HortScience, 40, 1763–1767.
Parvin, K., Ahamed, K. U., Islam, M. M., & Haque, M. N. (2015). Response of tomato plant under salt stress: Role of exogenous calcium. Journal of Plant Sciences, 10, 222–233.
Roelfsema, M. R. G., & Hedrich, R. (2005). In the light of stomatal opening: New insights into ‘the Watergate’. The New Phytologist, 167, 665–691.
Ross, A., & Williamson, C. (1951). Physiologically active emanations from virusinfected plants. Phytopathology, 41, 431.
Saini, S., Sharma, I., & Pati, P. K. (2015). Versatile roles of brassinosteroid in plants in the context of its homoeostasis, signaling and crosstalks. Frontiers in Plant Science, 6, 1–17.
Schaller, G. E. (2012). Ethylene and the regulation of plant development. BMC Biology, 10, 9–11.
Shafaghat, A. (2011). Antioxidant, antimicrobial activities and fatty acid components of flower, leaf, stem and seed of Hypericum scabrum. Natural Product Communications, 6, 1739–1742.
Shafique, S., et al. (2014). Determination of molecular and biochemical changes in cotton plants mediated by mealybug. NJAS – Wageningen Journal of Life Sciences, 70–71, 39–45.
Shah, A. A., et al. (2020). Butanolide alleviated cadmium stress by improving plant growth, photosynthetic parameters and antioxidant defense system of brassica oleracea. Chemosphere, 261, 127728.
Shi, C., et al. (2015). Ethylene mediates brassinosteroid-induced stomatal closure via Gα protein-activated hydrogen peroxide and nitric oxide production in Arabidopsis. The Plant Journal, 82, 280–301.
Singh, M., Gupta, A., & Laxmi, A. (2014a). Glucose control of root growth direction in Arabidopsis thaliana. Journal of Experimental Botany, 65, 2981–2993.
Singh, M., Gupta, A., & Laxmi, A. (2014b). Glucose and phytohormone interplay in controlling root directional growth in Arabidopsis. Plant Signaling & Behavior, 9, 4–8.
Swanson, S., & Gilroy, S. (2010). ROS in plant development. Physiologia Plantarum, 138, 384–392.
Symons, G. M., et al. (2006). Grapes on steroids. Brassinosteroids are involved in grape berry ripening. Plant Physiology, 140, 150–158.
Tariq, M., Shah, A. A., Yasin, N. A., Ahmad, A., & Rizwan, M. (2020). Enhanced performance of Bacillus megaterium OSR-3 in combination with putrescine ammeliorated hydrocarbon stress in Nicotiana tabacum. International Journal of Phytoremediation, 1–11. https://doi.org/10.1080/15226514.2020.1801572
Ul Haq, M. Z., Zhang, Z., Wei, J., & Qiang, S. (2020). Ethylene biosynthesis inhibition combined with cyanide degradation confer resistance to quinclorac in echinochloa crus-galli var. Mitis. International Journal of Molecular Sciences, 21, 1573.
Vardhini, B. V., & Anjum, N. A. (2015). Brassinosteroids make plant life easier under abiotic stresses mainly by modulating major components of antioxidant defense system. Frontiers in Environmental Science, 2, 1–16.
Verma, S., Mamdani, M. M., Al-Omran, M., Melo, M., & Rouleau, J. L. (2007). Angiotensin receptor blockers vs. angiotensin converting enzyme inhibitors and acute coronary syndrome outcomes in elderly patients: A population-based cohort study (UMPIRE study results). Journal of the American Society of Hypertension, 1, 286–294.
Vissenberg, K., Claeijs, N., Balcerowicz, D., & Schoenaers, S. (2020). Hormonal regulation of root hair growth and responses to the environment in Arabidopsis. Journal of Experimental Botany, 71, 2412–2427.
Wang, Y. H., & Irving, H. R. (2011). Developing a model of plant hormone interactions. Plant Signaling & Behavior, 6, 494–500.
Wang, M., et al. (2015). Functional analysis of GmCPDs and investigation of their roles in flowering. PLoS One, 10, 1–25.
Wei, L. J., et al. (2015). Ethylene is involved in brassinosteroids induced alternative respiratory pathway in cucumber (Cucumis sativus L.) seedlings response to abiotic stress. Frontiers in Plant Science, 6, 1–13.
Weller, J. L., et al. (2015). Ethylene signaling influences light-regulated development in pea. Plant Physiology, 169, 115–124.
Wu, J., et al. (2019). Cell membrane-interrupting antimicrobial peptides from Isatis indigotica fortune isolated by a Bacillus subtilis expression system. Biomolecules, 10, 30.
Yasin, N. A., et al. (2017). Imperative roles of halotolerant plant growth-promoting rhizobacteria and kinetin in improving salt tolerance and growth of black gram (Phaseolus mungo). Environmental Science and Pollution Research. https://doi.org/10.1007/s11356-017-0761-0
Yasin, N. A., et al. (2018a). Halotolerant plant-growth promoting rhizobacteria modulate gene expression and osmolyte production to improve salinity tolerance and growth in Capsicum annum L. Environmental Science and Pollution Research, 25, 23236–23250.
Yasin, N. A., et al. (2018b). The beneficial role of potassium in Cd-induced stress alleviation and growth improvement in Gladiolus grandiflora L. International Journal of Phytoremediation, 20, 274.
Yousaf, A., Qadir, A., Anjum, T., & Ahmad, A. (2015). Identification of microbial metabolites elevating vitamin contents in barley seeds. Journal of Agricultural and Food Chemistry, 63, 7304–7310.
Zaharah, S. S., Singh, Z., Symons, G. M., & Reid, J. B. (2012). Role of brassinosteroids, ethylene, abscisic acid, and indole-3-acetic acid in mango fruit ripening. Journal of Plant Growth Regulation, 31, 363–372.
Zaheer, M. M., et al. (2017). Amelioration of cadmium stress in gladiolus (Gladiolus grandiflora L.) by application of potassium and silicon. Journal of Plant Nutrition. https://doi.org/10.1080/01904167.2017.1385808
Zhu, T., et al. (2016). Ethylene and hydrogen peroxide are involved in brassinosteroid-induced salt tolerance in tomato. Scientific Reports, 6, 1–15.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Shahzadi, I., Ahmad, A., Noreen, Z., Akram, W., Yasin, N.A., Khan, W.U. (2022). Brassinosteroid and Ethylene-Mediated Cross Talk in Plant Growth and Development. In: Khan, M.T.A., Yusuf, M., Qazi, F., Ahmad, A. (eds) Brassinosteroids Signalling. Springer, Singapore. https://doi.org/10.1007/978-981-16-5743-6_7
Download citation
DOI: https://doi.org/10.1007/978-981-16-5743-6_7
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-5742-9
Online ISBN: 978-981-16-5743-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)