
Abstract
Global climate change accelerates the coincidence of a variety of abiotic stresses, viz., salinity, drought, flooding, high and low temperature, and biotic stresses, viz. phytopathogens which degrade agricultural productivity. In such circumstances, plant growth-promoting rhizobacteria (PGPR) are eco-friendly and sustainable candidates to combat these stresses. Several PGPR with the ability to support plant growth under various stressed conditions have been commercialized. The current chapter is mainly restricted to beneficial effects of PGPR on plant growth and development under environmental and biotic stresses. It begins with the description of various abiotic and biotic stress factors affecting plant growth and their tolerance achieved by both physiological and molecular mechanisms of adaptation. The use of PGPR helps ameliorate these stresses in rhizosphere by using several mechanisms and has beneficial effects on plant growth after efficiently colonizing the root surface. Plant growth stimulation through PGPR is the net result of multiple mechanisms of action that may be activated simultaneously. Such bacteria are more likely to be used for stress tolerance to fulfill the need for food production under extreme environmental conditions. The bacterial inoculants also enhance nutrient uptake and crop growth. They are also involved in biocontrol so they may be good supplements to chemical fertilizers and agrochemicals. This chapter discusses the potential and key mechanisms used by PGPR under stress conditions for sustainable agricultural productivity followed by their prospects.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- ACC-deaminase activity
- Beneficial bacteria
- Environmental sustainability
- Integrated stress
- Phytohormones
- Plant tolerance
2.1 Introduction
Global warming leads to abiotic and biotic stresses that are affecting crop productivity all over the globe. Abiotic stresses include salinity, drought, flooding, heavy metals, heat, and cold stress, while biotic stresses are caused by the attack of insects, pests, and microbial infection. Such abiotic and biotic stresses are collectively known as integrated stresses (Flynn 2003; Brown et al. 2017). Abiotic stresses are also known as environmental stresses that drastically reduce plant growth by disrupting metabolic and biochemical attributes of a plant (Suzuki et al. 2014). Abiotic stresses induce plant responses through modification in cellular metabolism, reduction in plant growth, and modulation in biochemical attributes (Singh 2015). Many studies have illustrated the effect of selective abiotic stresses such as salinity (Mumtaz et al. 2018; Sapre et al. 2021), heavy metals (Thakare et al. 2021), drought (Mumtaz et al. 2020; Ansari et al. 2021), flooding (Bal and Adhya 2021), heat (Sarkar et al. 2021), and cold (Vega-Celedón et al. 2021) on the growth of different plants.
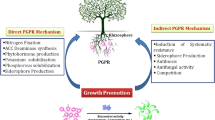
Microorganisms can be handy in mitigation of abiotic and biotic stresses. Beneficial bacteria which constitute up to 2–5% of the total bacterial population in soil are beneficial for plant growth even under adverse environments (Nazli et al. 2020). Millions of microorganisms colonizing the rhizosphere form a complex biological community with a plant root system and stimulate plant growth through their metabolic activities (Schmidt et al. 2014; Zvinavashe et al. 2021). Plant growth promoting bacteria (PGPR) improve plant nutrient availability and help in sustained crop yields (Hayat et al. 2010) through efficient nutrient solubilization and mobilization, providing biocontrol against phytopathogens, improving soil structure and quality by sequestrating toxic salt ions, and even degrading various xenobiotic compounds (Braud et al. 2009; Koskey et al. 2021). The application of such inoculants is a promising strategy for the amelioration of various stresses and improving plant health (Ahmad et al. 2013a). Bioaugmentation of PGPR modulates environmental stresses through direct and indirect mechanisms that induce systemic tolerance (Yang et al. 2009). Production of phytohormones and 1-amino-cyclopropane-1-carboxylic acid (ACC) deaminase activity by various PGPR has a crucial role in plant growth under environmental as well as biotic stresses. It activates the radicular system and promotes root elongation through the development of apical meristem and lateral roots, thus improving plant access to soil nutrients (Vessey 2003; Liu et al. 2021). In the current chapter, we have discussed the status of various abiotic and biotic stresses and their effects on plant growth. We have also discussed the potential of PGPR for remediation of these stresses. Abiotic stress induced by salt, drought, flooding, heat, and cold factors and biotic stress due to fungal, bacterial, and nematode pathogen have been reviewed. Various direct and indirect mechanisms used by PGPR for amelioration of these stresses have also been discussed.
2.2 Plant Responses to Stress
Plant stress is a condition in which plants are developed in non-ideal circumstances for their growth. It could be due to various external biological and non-biological factors that lead to a direct effect on plant growth. Biological stresses are also known as biotic stress, observed on plants due to plant interactions with pathogenic bacteria, fungi, virus, nematodes, weeds, and/or insects. Non-biological stress could be due to physical factors such as deficiency or toxicity of nutrients, high or low temperature, limited water availability or excess of water conditions, etc. (McKersie and Lesheim 1994). Non-biological factors result in non-favorable conditions to plants and enforce them under a state of stresses known as abiotic or environmental stresses. Mostly, abiotic stresses have a huge impact on plants by causing up to a 50% reduction in growth. Both biotic and abiotic stresses could affect plant growth for short or long periods depending on the exposure level (Mosa et al. 2017). Plants are exposed to abiotic and biotic stress factors, collectively called stressors that deviate plant’s optimal growth conditions. Stressors damage plant growth up to certain tolerance limits and, after that, may cause death. Stressors are interconnected to each other and affect plants by causing osmotic stress, ion imbalance, disrupting plant metabolism, production of ethylene, reactive oxygen species (ROS), and cell homeostasis. Many groups of genes have also shown changes in their expression patterns due to various stressors (Mittler 2006; Ramegowda et al. 2020; Kusale et al. 2021; Sofy et al. 2021).
Plants use a stress-sensing mechanism that depends on plant species and type of stress. Plant response to stress can be categorized into alarming, resistance, exhaustion, and regeneration (Mosa et al. 2017). In initial stress exposure, distinguishable changes from optimal plant growth in terms of functional deviation, declining vitality, and increase in catabolism take place. If stressors continue, these affect plant growth and productivity, and plant responds through stress adaptation, repair, and hardening processes. To minimize damage caused by various stresses, plant activates a complex signaling cascade of defense and specific ion channels; generates ROS and phytohormones, and expresses various stress-related genes that improve plant stress tolerance (Ramegowda et al. 2020; Tanveer and Ahmed 2020; Salvi et al. 2021). Every plant species has a certain limit for resistance against stresses that cannot be extended through external modification. Further increase in stress dosage beyond the plant resistance will cause loss of vitality, severe cell damage, and plant death (Mosa et al. 2017). Before complete senescence, plants can regenerate their physiological standards that allow plants to survive under various stresses. However, if stressors cause permanent damage, then it will not recover. A plant can survive under stress conditions but with limited growth and reproduction ability. These responses usually involve physiological-, biochemical-, and molecular-level actions (Kranner et al. 2010; Brunetti et al. 2019; Ansari et al. 2021). Plant response to stress can be modified/improved through exogenous application of various chemicals, phytohormones, and/or by modification of growing conditions. Rhizosphere microbiome engineering has also been studied as an eco-friendly, economical, and sustainable approach by the soil microbiologists.
2.3 Strategies for Stress Amelioration
Several strategies have been proposed by plant scientists to ameliorate stress. These strategies are comprehensively reviewed and available in literature (Glick et al. 2007; Saharan and Nehra 2011; Nadeem et al. 2014). Certain chemicals are used to reduce the impact of stresses on plant growth. These chemicals can reduce the negative impact of ethylene on plant growth by inhibiting its excess production under stress. Important chemicals reported in the literature are silver ions (Ag+), cobalt ions (Co2+), and aminoethoxyvinylglycine (AVG) which improve plant growth by suppressing ethylene production (Coupland and Jackson 1991; Mckeon et al. 1995; Kim and Mulkey 1997). Savvides et al. (2016) reported that plant priming with chemical agents, hydrogen peroxide, melatonin, polyamines, sodium hydrosulfide, and sodium nitroprusside, can improve multiple-stress tolerance. Priming with γ-amino butyric acid and β-amino butyric acid also induces promising tolerance in plants against abiotic stresses (Vijayakumari et al. 2016). Soaking seed with these priming agents shows greater stress tolerance by improving metabolic events and shows early seed germination and seedling emergence as compared to non-primed seeds (Ashraf et al. 2018). These chemicals also improve antioxidative enzymes, proline, malondialdehyde, and improve stress-responsive proteins (Thakur et al. 2019; Buttar et al. 2020).
The use of chemicals in agriculture is, however, expensive and deteriorates the plant, soil, and human health (Dodd et al. 2004; Pahalvi et al. 2021). These chemicals can increase crop yield but reduce farm profitability. Scientists have also raised certain environmental issues due to the persistence and toxicity of these chemicals for the natural habitat (Ahmadi et al. 2009). The use of osmoprotectants and potassium to reduce the negative effects has also been reported. However, the use of chemicals cannot reduce the negative effects of ion toxicity caused by salinity or root desiccation under osmotic stress. Although plants employ some specific mechanisms to cope with stresses, the presence of beneficial microbes can be helpful to combat the negative effects of stresses on plant growth. Rhizosphere engineering by using PGPR can therefore be exploited as a useful strategy to ameliorate the negative effects on plant growth. It could be the most important approach to remediate multiple stresses for sustainable agriculture. Recently, such beneficial bacteria have been acknowledged by scientists for their capability to improve crop productivity as well as raise stress tolerance in various crops (Goyal et al. 2019; Njeru and Koskey 2021).
2.4 Role of Plant Growth-Promoting Rhizobacteria to Combat Stresses in Plants
Plants are associated with soil microorganisms in a symbiotic relationship; thus there is a strong plant-microbe interaction in the rhizosphere. Such symbiotic free-living soil microorganisms inhabiting the rhizosphere of many plant species having diverse beneficial effects on host plants are known as PGPR. Their interactions with plants are commercially explored and have biotechnological applications for sustainable agriculture to improve plant health in an eco-friendly manner (Gonzalez et al. 2015; Gouda et al. 2018). Biological activities of soil microbiome in the ecosystem make it vigorous for sustainable agriculture production. The PGPR dwell in plant roots and improve plant growth through various mechanisms including nitrogen (N) fixation (Kuan et al. 2016; Aasfar et al. 2021), solubilization of phosphorus (P) (Selvakumar et al. 2009; Kusale et al. 2021); potassium (K) (Ahmad et al. 2016) and zinc (Zn) solubilization (Mumtaz et al. 2017, 2019); production of siderophores (Gouda et al. 2018); ACC-deaminase (Misra et al. 2017; Ansari et al. 2021; Vega-Celedón et al. 2021); and phytohormones, e.g. indole-3-acetic acid (IAA) (Kim et al. 2017; Khan et al. 2020a, b; Kusale et al. 2021), cytokinins (CK) (Liu et al. 2013; Ilyas et al. 2020), gibberellins (GA) (Gutiérrez-Mañero et al. 2001; Khan et al. 2020a, b), abscisic acid (ABA) (Belimov et al. 2014; Kang et al. 2019), hydrogen cyanide (HCN) (Kremer and Souissi 2001), and antibiotics and/or lytic enzymes (Gouda et al. 2018; Migunova and Sasanelli 2021). They also possess more specific plant growth-promoting (PGP) traits that can ameliorate/reduce environmental stresses such as heavy metals, salinity, and drought stress and biological control of phytopathogens (Egamberdieva and Lugtenberg 2014). Such PGPR are classified into two main types: (1) the extracellular PGPR that can colonize the rhizoplane and (2) intracellular PGPR that colonize inside the root cortex or nodule cells (Bhattacharyya and Jha 2012).
The PGPR possess several PGP characteristics that are involved in enhancing plant physiology under normal as well as stress conditions through various mechanisms (Ahmad et al. 2011; Zakry et al. 2012; Asghari et al. 2020). They produce volatile organic compounds (VOCs) and enzymes for biocontrol of phytopathogens and neutralize abiotic and biotic stresses. However, different PGPR have different modes of action which differ under certain circumstances according to their host plant, abiotic and biotic factors, plant defense and developmental stages, and presence of competition for microflora (Garcia et al. 2015). They can support plant growth and development through enhancing nutrient availability by nitrogen fixation, solubilization of minerals, mineralization of nutrients, and production of phytohormones and siderophores (Bhardwaj et al. 2014). Such mechanisms improve plant growth directly and may contribute according to the ability of microbial strains. In the presence of PGPR, improved individual ion fluxes at the root surface enrich nutrient uptake (Gouda et al. 2018). The PGPR may indirectly improve plant growth and development by producing suppressive compounds that prevent the phytopathogen attack on plants (Singh and Jha 2015; Migunova and Sasanelli 2021). This process actively helps plants grow under biotic stresses (Akhgar et al. 2014). In this mechanism, PGPR contribute through the production of chitinases, cellulases, proteases, and other hydrolytic enzymes and production of antibiotics, siderophores, VOCs, exopolysaccharides (EPS), and induced systemic resistance (ISR) in response to a plant pathogen (Gupta et al. 2015). The occurrence of abiotic and biotic stresses drastically affects crop productivity over the globe. So, there is a need to explore unique PGP characteristics of soil microbiome under extreme environmental conditions, their relations with crop plants, genetic diversity, and successful application methods for increased agriculture production (Grover et al. 2011).
2.4.1 Abiotic Stress
External conditions of either physical or biological nature that unfavorably disturb plant growth are known as stresses. Physical stresses, viz., salinity, drought, waterlogging, heat, cold and heavy metal contamination, etc., are termed as abiotic stresses (Brown et al. 2017), while biological agents like microbial and insect and pest attacks are known as biotic stresses (Flynn 2003). According to an estimate, between 2003 and 2013, 78 types of natural disasters caused 140 billion dollars worth of losses in developing countries (FAO 2015). Abiotic stresses impose their hazardous effects on plant growth by altering the metabolic, biochemical, and physiological attributes of plant growth (Suzuki et al. 2014; Sapre et al. 2021). Abiotic stresses initiate plant responses in terms of cellular metabolism, changes in growth and yield attributes, modulation in metabolites and biochemical attributes, and alteration in gene expression (Singh 2015; Sofy et al. 2021).
Abiotic stresses impair photosynthetic machinery in plants such as photosystem I and II, stomatal conductance, carbon (C) fixation, electron transport, and ATP-generating system (Nouri et al. 2015; Chatterjee et al. 2020; Sharma et al. 2020). These may modify the structure of chloroplasts and thylakoid complexes, cause changes in metabolites concentration, and generate ROS (Gan et al. 2019). For example, salinity stress causes loss of photosynthetic pigments and stomatal closure due to reduced nutrient uptake (Stepien and Kłbus 2006). It also reduces the carbon dioxide (CO2) availability and carbon (C) fixation that generate ROS, thus causing cellular damage through oxidation of lipids, proteins, and nucleic acids (Lopez-Raez 2016). Abiotic stresses adversely affect plant growth by decreasing water availability that causes an imbalance between enzymatic and non-enzymatic reactions taking place in chloroplasts, mitochondria, and electron transport reactions (Koyro et al. 2012; Adak et al. 2020). Various crop plants differ in their ability to withstand most of the abiotic stresses. The plant tolerates abiotic stresses through genetic and physiological adaptive mechanisms and interacts symbiotically with endophytic and rhizospheric bacteria. Many studies have demonstrated genetic adaptation to abiotic stresses including molecular mechanism, identification of overexpressed genes, and development of stress-tolerant transgenic plants (Zang et al. 2017; Datir et al. 2019; Woo et al. 2020). Several studies have also reported plant-microbe interactions in abiotic stress tolerance. Pre-treatment of plant seed or seedlings with PGPR can improve plant survival during abiotic stresses which are being discussed in subsequent sections for each abiotic stress.
2.4.1.1 Salt Stress
Agriculture is moving into regions with lower rainfall and saline soil that occupy 6% of the world’s land area due to the increased need for food production (Munns 2005). High salinity is one of the major abiotic factors that adversely affect crop growth, development, and productivity (Mumtaz et al. 2018). It converts fertile fields to marginal lands and causes desertification. Soil salinization is the buildup of dissolved salts in agricultural soils over a prolonged time. It may arise due to weathering of rocks and minerals, precipitation, wind-borne salts, the influx of seawater, and cultivation operations, viz. unnecessary irrigation, land clearing, and insufficient drainage. The dryland salinity originated due to salt deposition in the topsoil through water evaporation (Rengasamy 2002, 2010; Pitman and Läuchli 2002; Shrivastava and Kumar 2015). Salinity effects are more harmful under the raised groundwater table, arid, and coastal areas, and only salt-tolerant plants called halophytes can grow (Rengasamy 2002; Flowers and Colmer 2008). The salinity-sensitive plants called glycophytes eventually die as their physiological and biochemical attributes are susceptible to salt-induced stress (Hajiboland et al. 2018).
Salinity stress hinders crop production by causing various physiological and metabolic changes. It suppresses plant growth through osmotic stress and ion toxicity (Rozema and Flowers 2008; Rahnama et al. 2010). Osmotic stress due to higher salt buildup in soil and plants causes a reduction in water absorption ability, accelerated water loss from leaves, and various physiological changes, viz. nutrient imbalance, membrane interruption, inability to reduce ROS, and alteration in antioxidant enzymes (Munns and Tester 2008; Rahnama et al. 2010; Sagar et al. 2020). Salinity causes accumulation of sodium (Na+) and chloride (Cl-) ions in plant tissues that cause severe ion imbalance and physiological disorders. Accumulation of higher Na+ contents retard the uptake of potassium ions (K+) that is necessary for plant growth and development and may cause plant death (James et al. 2011). Due to salt stress, oxidative stress elevates the production of ROS which leads to oxidative damages in proteins, lipids, and DNA and interrupts vital cellular functions of plants (Xie et al. 2014; Kusale et al. 2021).
Plants tolerate high salt stress by developing various physiological adaptations viz. ion homeostasis, transport, and compartmentalization, synthesis of osmoprotectants, compatible solutes, polyamines, nitric oxide, and activation of antioxidant enzymes and hormone modulation (Gupta and Huang 2014; Dubey and Verma 2019; Sofy et al. 2021). There is a need to apply an attenuated approach to remediate the effects of salt stress on crops. Due to reduced agricultural production under salinity stress, the production of salinity-resistant crops is a desirable scientific achievement. For successful saline soil reclamation, effective soil management practices are needed (Munns and Tester 2008). The sustainable management practices to facilitate future agriculture production provide more support regarding crop production. Various plant growth-promoting microorganisms (PGPM) play a beneficial role to promote agricultural productivity in an ecofriendly manner (Rodriguez and Redman 2008). Such PGPM associated with the stressed plant augment to adapt their microenvironments (Paul and Lade 2014). Various crops under salinity-stressed conditions respond to PGPM that improves plant growth. Such biotechnology can help to develop salinity-tolerant technologies for growing crops. Augmented PGPR colonizes plant’s rhizospheric soil and alleviates stress in crops. This remediation strategy could be successful due to a wide range of salinity-tolerant microbes, their associated interactions with plants, potential PGP metabolites, and prospective application methods.
The PGPR have been investigated under salt-induced stress for their role in improving the efficiency of photosynthesis, ion homeostasis, and plant-water relations regulated by a signaling network that mitigates the stress (Wang et al. 2016; Mahadik and Kumudini 2020; Taj and Challabathula 2021). They optimize water uptake and stomatal opening by improving the transpiration rate and hydraulic conductivity. Marulanda et al. (2010) reported that inoculation with Bacillus megaterium improved root hydraulic conductivity in maize under 2.59 dS m-1 salt stress that correlated with increased ZmPIP expression. Arabidopsis thaliana treated with Bacillus subtilis encourages proline production with the expression of proBA genes that conferred salt tolerance to withstand the initial osmotic shocks (Chen et al. 2007). In another study, Bacillus amyloliquefaciens strain SN13 inoculated in rice augmented salt stress of 200 mM NaCl through upregulation of stress-related genes (Nautiyal et al. 2013). Upadhyay and Singh (2015) reported that inoculation with Bacillus aquimaris promoted total sugar contents in wheat that related to increased NPK and biomass accumulation and reduction of Na+ in leaves. del Amor and Cuadra-Crespo (2012) reported that pepper plants inoculated with Azospirillum brasilense and Pantoea dispersa improved salinity resistance in pepper and recorded higher plant dry matter correlated with enhanced stomatal conductance and photosynthetic rate. The PGPR induces accumulation of osmoprotectants and compatible solutes, viz., glycine, proline, and trehalose having more potency than their linked host plants under salt stress (Arif and Ghoul 2018; Taj and Challabathula 2021). These compatible solutes are absorbed by plant roots that maintain osmotic balance. The bean crop overexpressed trehalose-6-phosphate gene when co-inoculated by Paenibacillus polymyxa and Rhizobium tropici that promoted nodulation and N accumulation. The upregulation of stress tolerance genes through R. tropici and P. polymyxa induced salinity tolerance in beans (Figueiredo et al. 2008).
The PGPR trap cations in their EPS matrix and limit salt uptake by plants that improve their root structure and regulate ion affinity transporter’s expression. They relieve nutrient imbalance by reducing Na+ and Cl- influx and increased exchange of macronutrients and micronutrients through mineralizing the nutrients, lowering rhizospheric pH, and producing siderophores (Nadeem et al. 2016; Etesami and Alikhani 2019; Ji et al. 2020). They maintain ion homeostasis by boosting activity of K+ transporters, improving K+:Na+ ratios, decreasing Na+ and Cl- buildup, and increasing Na+ exclusion. Maize inoculation with IAA-producing Azotobacter strains C5 and C9 showed improved Na+ exclusion and K+ uptake (Rojas-Tapias et al. 2012). Under 200 mM NaCl stress, Puccinellia tenuiflora inoculated with B. subtilis strain GB03 exhibited reduced Na+ buildup (Niu et al. 2016). PGPR produce exogenous phytohormones and enzymes that may help ameliorate salt stress. During salinity stress, plants also synthesize phytohormones in the response of plant-microbe relations (Dodd et al. 2010; Kusale et al. 2021).
Several PGPR can produce phytohormones, viz., IAA, CK, GA, and ABA, proliferate roots hairs, and enhance uptake of water and nutrients under salt stress. Salinity stress reduces the endogenous IAA and ABA levels in plants which can be maintained at their normal level through PGPR inoculation. For example, B. amyloliquefaciens strain SQR9 stimulated ABA production of inoculated plants under salinity stress (Chen et al. 2016). Similarly, Kim et al. (2014) inoculated Enterobacter sp. strain EJ01 in Arabidopsis thaliana under salt stress and reported increased expression of salt stress-responsive genes. They also reported that strain EJ01-inoculated plant showed increase in ROS scavenging activities through production of ascorbate peroxidase (APX) under salinity stress. PGPR produce CK under salt stress; however their role in salt stress tolerance is still under question (Arkhipova et al. 2005). Egamberdieva (2009) inoculated wheat with CK-producing Pseudomonas strains TSAU22, TSAU6, and TSAU20 that enhanced wheat growth under salt stress. PGPR modulate ABA biosynthesis under salinity stress and support plant growth. In another study, Dietzia natronolimnaea strain STR1 induced salt-tolerance in wheat by modulating ABA signaling cascade and upregulation of salt stress-induced gene. These strains contributed to antioxidant enzymes and proline content gene expression that enhanced salt tolerance (Bharti et al. 2016). Cucumber inoculation with strains of Burkholderia, Promicromonospora, and Acinetobacter showed a significant increase in biomass, water potential, and decreased electrolyte leakage at 120 mM NaCl stress. These strains showed downregulation of ABA and increased levels of salicylic acid and GA contents in inoculated plant (Kang et al. 2014). Similarly, improvement in maize growth has also been reported on compacted saline-sodic soil through seed inoculation with Pseudomonas syringae and Pseudomonas fluorescens (Zafar-ul-Hye et al. 2018).
Under salinity stress, ethylene levels increase, which causes inhibition in auxin transcription and constrain plant growth. PGPR produce ACC-deaminase that inhibit the biosynthesis of ethylene in plants. Soil bacteria produce ACC-deaminase enzyme that converts ACC to ammonia and a-ketobutyrate and enhance salt tolerance and promote plant growth (Glick et al. 2007). Tomato seedlings inoculated with Pseudomonas putida strain UW4 grown under 90 mM NaCl stress showed increased shoot growth due to the expression of upregulated Toc GTPase that import stress response proteins (Yan et al. 2014). Okra inoculation with Enterobacter sp. strain UPMR18 ameliorated salt stress due to escalating antioxidant activities (Habib et al. 2016). Nadeem et al. (2009) reported salt tolerance in maize due to co-inoculation of ACC-deaminase-producing Pseudomonas and Enterobacter spp. The ACC-deaminase- and IAA-producing PGPR can ameliorate the negative effects of salt stress effectively. The accumulation of IAA stimulates the ACC synthase genes, resulting in a higher ACC amount that leads to ethylene production. The ACC-deaminase-producing PGPR break down the excess ACC and lower the toxic level of ethylene, and simultaneously IAA production promotes plant growth (Glick 2012; Ahmad et al. 2013b). In another study, the combined use of ACC-deaminase-containing PGPR strains Agrobacterium fabrum and B. amyloliquefaciens along with biochar improved wheat productivity under drought-stressed conditions (Danish and Zafar-ul-Hye 2019; Zafar-ul-Hye et al. 2019).
The IAA- and ACC-deaminase-producing Arthrobacter sp. and Bacillus sp. inoculation promoted proline contents under osmotic stress and revealed downregulation of stress-inducible genes in sweet pepper (Sziderics et al. 2007). In another study, Panwar et al. (2016) inoculated IAA- and ACC-deaminase-producing P. dispersa strain PSB3 in chickpea under 150 mM NaCl stress and reported significant improvement in plant biomass and grain yield attributes due to reduced Na+ uptake, electrolyte leakage, and enhanced K+ uptake. The extracellular secretions from PGPR could also manipulate signaling pathways and alleviate salt stress (Bhattacharyya et al. 2015; Zhou et al. 2016; Kumawat et al. 2020). PGPR produce EPS responsible for bacterial attachment to soil particles. EPS improves soil structure and water-holding capacity by binding soil particles (Upadhyay et al. 2011). Qurashi and Sabri (2012) inoculated chickpea with Halomonas variabilis strain HT1 and Planococcus rifietoensis strain RT4 and reported salt tolerance by improving plant growth and soil aggregation. P. polymyxa ANM59 and Paenibacillus sp. ANM76 have also been reported as potential bioinoculants to improve abiotic stress tolerance of crop plants (Ahmad et al. 2019a). The mechanisms of action adopted by PGPR to mitigate salt stress are summarised in Fig. 2.1 and Table 2.1.

Proposed salt-tolerance mechanisms mediated by PGPR. Rhizobacteria colonize the root surface and produce extracellular polysaccharides to ameliorate salinity stress. They manipulate phytohormones level and improve nutritional balance and photosynthetic activity through various physiological and biochemical processes
2.4.1.2 Drought Stress
Drought is a condition of an uncommon drop in soil moisture content due to a prolonged period of low rainfall. It is the utmost devastating abiotic stress that restricts crop growth and productivity worldwide. The arable lands affected by drought have doubled during the period from 1970 to 2000 and is expected to increase further by 2050, posing serious threat to crop productivity (Vinocur and Altman 2005). In the last four decades, drought caused reduction in cereal yield by up to 10% and is estimated to affect crop production in over 50% of cultivated land by 2050 (Lesk et al. 2016). Drought may be of moderate, short, extremely severe, and prolonged duration (Bottner et al. 1995). It causes a reduction in plant growth by altering physiological traits of a plant (Rahdari and Hoseini 2012; Mumtaz et al. 2020). Growth reduction due to drought stress has been described in cereal crops such as barley (Hellal et al. 2018), maize (Avramova et al. 2015), rice (Mumtaz et al. 2020), wheat (Kosová et al. 2016), etc. Among growth attributes, drought commonly causes a reduction in water content and fresh weight of plants (Jaleel et al. 2009; Meenakshi et al. 2019). Due to limited water availability, drought causes a reduction in nutrient availability and transportation (Selvakumar et al. 2012). Free radicals induced under drought stress cause oxidative stress that affects antioxidant defense and ROS production. The ROS generation damages lipid peroxidation and degradation of membrane, protein, and lipids in plants (Hendry 2005; Chiappero et al. 2019). Drought stress causes a reduction in chlorophyll content, photosynthetic and transpirational rates, and stomatal conductance in various crops (Ohashi et al. 2006; Mumtaz et al. 2020).
Drought as multi-dimensional stress affects various subcellular compartments, cell organs, and whole plant levels (Horn and Chapman 2012). Drought destructively upsets the quantity and quality of plants. Drought stress alleviation is important to fulfill food demand. To address this global challenge, there is a dire need to develop improved crop germplasm and technologies to increase water use efficiency (WUE) (Passioura 2007). Recently, the application of beneficial microorganisms has got attention for the mitigation of drought by improving WUE (Marulanda et al. 2009; Ngumbi and Kloepper 2016). Under drought stress, the plant-associated bacterial community in the rhizosphere alleviates drought stress and improves plant health by improving the resistance in plants against stressors (Cherif et al. 2015; Aslam et al. 2020; Kour et al. 2020). The PGPR in the water-limited rhizosphere with repeated dry periods is more stress-tolerant and increases plant growth as compared to PGPR population of the well-irrigated rhizosphere (Mayak et al. 2004). Drought tolerance mediated by PGPR could be through the production of phytohormones, viz., IAA, ABA, and GA, production of ACC-deaminase to reduce ethylene production, and induced drought tolerance through the production of metabolites, viz., siderophore and EPS (Timmusk and Nevo 2011; Meenakshi et al. 2019; Ansari et al. 2021).
Plants produce phytohormones such as IAA, ABA, GA, and CK which play important role in drought stress alleviation (Egamberdieva 2013; Fahad et al. 2015; Kumar et al. 2019a). PGPR are reported to produce these phytohormones under drought stress that can improve plant growth (Khan et al. 2020c). The IAA-producing PGPR alleviate drought stress by improving lateral root development and water uptake (Mantelin and Touraine 2004). For example, Dimkpa et al. (2009) reported that Azospirillum (IAA producing) promoted drought stress tolerance in plants. Nitric oxide (NO, signaling molecule) produced by A. brasilense inoculation in tomato showed induction of IAA pathway and development of adventitious roots. Its inoculation in maize grown under drought stress also improved water content in comparison to uninoculated control plants (Molina-Favero et al. 2008). Wheat treated with A. brasilense strain Sp245 improved drought stress tolerance through increased grain yield; Mg, K, and Ca concentration; water content; and lowering volumetric cell wall elasticity (Creus et al. 2004). The IAA-producing Bacillus thuringiensis inoculation in Lavandula dentata showed drought stress amelioration and improved plant metabolic activities, nutrition, and physiology (Armada et al. 2014). GA- and ABA-producing PGPR also help plants withstand drought stress. Soybean inoculation with P. putida strain H-2-3 (GA producing) showed an increase in plant growth under drought stress (Sang-Mo et al. 2014). Similarly, inoculation of maize with Azospirillum lipoferum ameliorated drought stress in plants (García et al. 2017). The ABA is a stress hormone, and its biosynthesis is induced during cellular dehydration under water deficit conditions. The inoculation of CK-producing B. subtilis in Platycladus orientalis improved drought stress alleviation by increasing ABA levels (Egamberdieva et al. 2017).
Drought stress enhances the production of ethylene that causes retarded plant growth (Hardoim et al. 2008). The ACC-deaminase-producing bacteria sequester and degrade plant ACC and improve plant growth, supply N and energy, and reduce the deleterious effect of ethylene in plants (Glick 2005; Nadeem et al. 2017; Danish and Zafar-ul-Hye 2019). Mayak et al. (2004) inoculated ACC-deaminase-producing Achromobacter piechaudii in tomato and pepper and reported increase in plant growth under drought stress by reducing ethylene production. The ACC-deaminase-containing PGPR enhance grain yield and N accumulation in grains and repair depressed nodulation in plants (Dodd et al. 2004; Danish et al. 2020). Inoculation with ACC-deaminase-producing P. fluorescens in Pisum sativum under drought stress showed longer roots due to increased water uptake (Zahir et al. 2008). Variovorax paradoxus strain 5C-2 improved pea productivity through increase in WUE due to ABA production in drought stress (Belimov et al. 2009). PGPR promoted wheat growth due to better water and nutrient acquisition over uninoculated control (Shakir et al. 2012; Ansari et al. 2021). Sharma et al. (2013) reported that co-inoculation of Bacillus, Pseudomonas, and Mesorhizobium strains (ACC-deaminase producing) in Cicer arietinum under drought conditions promoted seed germination and seedling biomass over uninoculated control (Hui and Kim 2013).
2.4.1.3 Flooding Stress
Waterlogging and submergence are abiotic stresses, collectively called flooding stresses. Flooding stress is a global issue that influences plant composition and productivity (Phukan et al. 2016). Under flooding conditions, soil becomes completely saturated with water and develops an anaerobic environment. Plant roots become hypoxic which impairs the growth and development of the plants (Nishiuchi et al. 2012; Glick 2014). According to an estimate, flooding affected nearly 13% of the country’s land and nearly 10% of agricultural lands globally (Cramer et al. 2011). Flooding stress causes yield loss up to 15–80% in various crops (Patel et al. 2014).
Under flooding conditions, poorly adapted plant species are severely affected by excess water that results in 104-fold slower oxygen diffusion as compared to air (Armstrong and Drew 2002). Oxygen-deficient conditions lead to higher production of plant hormone “ethylene” that increases ROS production, destruction of macromolecules, and inhibition of photochemical and ultimately causes plant death (Grichko and Glick 2001a; Glick 2005; Ahmed et al. 2006). Under flooding conditions, plants exhibit symptoms of chlorosis, necrosis, and low productivity (Paul et al. 2016). Various wetland plant species are highly effective due to a combination of various physiological adaptations to withstand flooding stress conditions (Ravanbakhsh et al. 2017). Such plants acclimatize flooding stress through a physical method including escape from submerged conditions, prevention of oxygen deficiency via internal aeration, anoxia tolerance, and avoiding oxidative damage (Armstrong et al. 1994; Gibbs and Greenway 2003; Voesenek et al. 2003). Under flooding stress, plant roots produce a larger amount of ethylene that modifies phytohormone cascade of ABA, GA, and IAA to encourage the emergence of adventitious rooting in plants (Vidoz et al. 2010; Dawood et al. 2016; Ali et al. 2020). Root development also requires ethylene that persuades the development of ROS in epidermal cells leading to cell death by dropping antioxidant activity (Steffens and Sauter 2009; Yamauchi et al. 2011).
Under flooding stress, ACC-deaminase-producing bacteria play a key role in reducing excess ethylene production by catabolizing ACC to ammonia and α-ketobutyric acid (Ali and Kim 2018; Sasidharan et al. 2018; Bal and Adhya 2021). Limited literature is available to describe PGPR’s role in flooding stress mainly through their ability to produce ACC-deaminase. Transgenic tomato developed through ACC-deaminase expressing transcriptional control of rolD promoter from Agrobacterium rhizogenes and 35S constitutive expression was evaluated under flooding stress (Grichko and Glick 2001b). Development of transgenic plants with the insertion of rolD promoter showed ACC-deaminase expression and increased flooding tolerance by improving seedling biomass, epinasty, leaf chlorophyll, and protein content and reducing the ethylene production (Grichko and Glick 2001b). Under flooding stress, ACC synthase is produced in a higher amount in the plant tissues that induces the production of ACC-deaminase by bacteria. Symbiotic bacteria can reduce ethylene stress up to 60–90% by producing ACC-deaminase enzyme (Saleem et al. 2007; Jung et al. 2018; Ali et al. 2018). Grichko and Glick (2001a) inoculated tomato with Enterobacter cloacae (strains UW4 and CAL2) and P. putida (strains pRKACC) and reported that the expression of ACC-deaminase genes in these strains promoted substantial flooding stress tolerance by improving plant growth and physiological attributes (Grichko and Glick 2001a).
Farwell et al. (2007) inoculated ACC-deaminase-producing P. putida strain UW4 in ACC-deaminase-producing transgenic canola and non-transformed canola under flooding and nickel (Ni) stress. They reported that transgenic canola inoculated with strain UW4 showed better shoot biomass as compared to non-transformed canola under flooding conditions (Farwell et al. 2007). Barnawal et al. (2012) investigated the effect of Achromobacter xylosoxidans strain Fd2, Herbaspirillum seropedicae strain Oci9, Ochrobactrum rhizosphaerae strain Oci13, and Serratia ureilytica strain Bac5 on Ocimum sanctum. They reported that bacterial treatment alleviated waterlogging stress and reduced ethylene production, lipid peroxidation, proline contents, and improved chlorophyll content and nutrient uptake. Strain Fd2 exhibited maximum waterlogging tolerance with 46.5% higher growth and herb yield and a 53% reduction in ACC concentration over uninoculated waterlogged plants (Barnawal et al. 2012). Habibzadeh et al. (2012) treated canola plants with biofertilizers through foliar and seed application and reported their effect on physiological and morphological attributes of canola under flooding conditions. The biofertilizers were AAP (strains of Azotobacter, Azospirillum, and Pseudomonas) and APB (strains of Azospirillum, Pseudomonas, Bacillus). Both foliar and seed treatment with these biofertilizers significantly alleviated flooding stress by improving superoxide dismutase (SOD), catalase (CAT), and peroxidases (POD) activities and by reducing ethylene and lipid peroxidation (Habibzadeh et al. 2012).
Etesami et al. (2014) reported that rice inoculation with endophytic ACC-deaminase-producing P. fluorescens strain REN1 promoted root growth under flooding stress. The Streptomyces sp. strain GMKU inoculation in mung bean alleviated flooding stress by improving shoot and root growth and chlorophyll content and reducing ethylene formation (Jaemsaeng et al. 2018). Similarly, Ali et al. (2018) reported that Sesamum indicum inoculation with Pseudomonas veronii strain KJ showed flooding stress mitigation by improving plant growth and physiological attributes. Production of IAA by PGPR induces adventitious root growth that also helps plant maintain their growth and ameliorate flooding stress. Rice seedling inoculation with IAA-producing strain REN1 also showed better root development under flooding conditions (Etesami et al. 2014). Similarly, Kim et al. (2017) inoculated Klebsiella variicola strain AY-13 in soybean under waterlogged conditions and reported its potential to induce adventitious root growth, improved plant growth, and flooding tolerance (in soybean). Strain AY-13 was able to produce IAA up to 84.27±3.55 μg mL-1 as analyzed through gas chromatography-mass spectrometry selected ion monitoring (GC-MS/SIM) as reported by Kim et al. (2017).
2.4.1.4 Heat Stress
Temperature is a primary factor affecting plant growth and development (Hatfield and Prueger 2015). Each plant species requires a certain temperature regime characterized as a minimum, maximum, and optimal temperature range for its growth and productivity (Hatfield et al. 2011). Global warming is becoming a major issue worldwide due to elevated carbon dioxide that is causing economic losses. It is predicted that greenhouse gas emission is causing a steady increase in average temperature worldwide. The increase in average temperature linked with climate changes usually aggravates the heat stress in agriculture (Carbonell-Bojollo et al. 2019; Zhang et al. 2019). Heat stress negatively affects plant growth and development by causing changes in plant morphology and physiology (Prasad et al. 2008). Repeated extremely high-temperature events have caused phenological disturbances in different plant species (Li et al. 2017). In the early crop growth stages, heat stress inhibits seed germination and seedling’s emergence (Wahid et al. 2008). It can damage photosynthetic pigments, carbon metabolism, and organic solute transport (Hasanuzzaman et al. 2013; Balal et al. 2016). El-Daim et al. (2014) reported that temperature at 45 °C caused mortality of wheat seedlings up to 80–95%. The impact of heat stress on the reproductive phase of crops generally leads to variation in mineral nutrition, deactivation of the immune system, and the successive generation of ROS (Zhang et al. 2016; Lanza and dos Reis 2021). Heat stress elevation disrupts the rate of photosynthesis, stomatal conductance, and transpiration (Fahad et al. 2016). The activity of photosystem II (PSII) was inhibited due to heat stress that ultimately reduced photosynthetic rate which is the major cause of the inhibition of chlorophyll production (Chen et al. 2007; Killi et al. 2020).
Plants tolerate heat stress by altering osmotic pressure, antioxidants, plant hormone level, and regulation of HSPs. Plant regulates the production of certain plant hormones, such as ABA, GA, JA, and IAA, to improve heat resistance (Abdelrahman et al. 2017; Khan et al. 2020c). Among these phytohormones, IAA and GA improve plant development processes, while JA and ABA can alleviate the heat stress (Ahammed et al. 2016). Hydrogen sulfide production also improves resistance to heat stress in plants (Chen et al. 2016). The HSPs protect the natural protein and improve protein stability, which in turn improve the thermal tolerance in the plant (Wei et al. 2021). The accumulation of HSP in terms of ZmHSP17.2 in corn and OsHSP26 in rice showed a strong correlation in heat stress tolerance (Lee et al. 2007; Liu et al. 2013; Sarkar et al. 2021). Maize leaves under heat stress expressed sHSP17.2, sHSP17.4, and sHSP26, while these proteins were also regulated by ABA (Hu et al. 2010). HSFA1 is a major regulator of signal perception, transduction, and control of the expression of stress-sensitive genes in tomato plants. The expression of HSP26, HSP17.4, and HSP17.2 also played an important role in heat tolerance in corn (Asada 2006). A multifunctional cytoplasmic protein known as eukaryotic elongation factor 1α (eEF1A) is responsible for the heat resistance in tomatoes (Momčilović et al. 2016). Plants also increase the activity of antioxidant enzymes, including alpha-tocopherol; glutathione; ascorbic acid; CAT, SOD, APX, and POD; and glutathione reductase, to repair the oxidative damage caused by heat stress (Asada 2006; Khan et al. 2020a, b).
Plant inoculation with PGPR improves heat tolerance by enhancing the photosynthetic rate, water use efficiency, antioxidant enzymatic activity, and uptake of water and nutrients (Zhu et al. 2012; Maya and Matsubara 2013; Kang et al. 2019; Sarkar et al. 2021). The literature on the mechanism of heat stress tolerance due to bacterial inoculation is very limited; however, studies are available to support that bacterial augmentation can ameliorate heat stress by producing ACC-deaminase, EPS, and phytohormones which are involved in the expression of HSP. Application of ACC-deaminase-producing PGPR under heat stress could ease unfavorable conditions. Production of ACC-deaminase by PGPR reduces the inhibitory effect of stress hormone “ethylene” generated under heat stress through degrading ACC into ammonia and α-ketobutyrate (Barnawal et al. 2012). The detailed mechanism of conversion of ACC into ammonia and α-ketobutyrate by the action of ACC-deaminase produced by PGPR has already been reported in the above abiotic stress sections of this chapter. The PGPR uses a similar mechanism of action in all abiotic stress to reduce the lethal concentration of ethylene. Recently, Mukhtar et al. (2020) inoculated PGP Bacillus cereus having the ability to produce ACC-deaminase up to 0.76 to C0.9 μM/mg protein/h in tomato under heat stress. They reported a significant improvement in growth and physiological and biochemical attributes of tomato plants due to inoculation with EPS-producing, ACC-deaminase-containing B. cereus. Wu and Yang (2019) reported that ethylene-mediated signaling under heat stress was involved in the reduction of oxidative damage, maintenance of chlorophyll content, and enhancement of thermo-tolerance in rice seedlings. They also reported that HSP and ethylene-signaling-related genes were involved in complex network regulation that conferred thermo-tolerance to rice seedlings.
Srivastava et al. (2008) reported that high-temperature (40 °C up to 5 days) tolerant P. putida strain NBRI0987 isolated from chickpea rhizosphere was affected with drought stress. Strain NBRI0987 was able to tolerate heat stress due to its ability of biofilm formation and overexpression of sigma factor genes. Ali et al. (2009) inoculated sorghum with thermo-tolerant Pseudomonas sp. strain AKM-P6 under elevated temperatures (47–50 °C for day and 30–33 °C for a night). They reported that inoculation with strain AKM-P6 ameliorated the elevated temperature stress through well-established growth on the root surface and promoted root biomass, shoot biomass, amino acids, sugar, proline, chlorophyll, and induced synthesis of protein (Ali et al. 2009). El-Daim et al. (2014) reported the heat tolerance in wheat through inoculation with B. amyloliquefaciens strain UCMB5113 and A. brasilense strain NO40. They reported that inoculation with these strains improved heat tolerance through activating ascorbate-glutathione redox cycle, reducing generation of ROS and cell damage (El-Daim et al. 2014).
Ali et al. (2011) investigated heat tolerance in wheat through inoculation with thermo-tolerant PGPR P. putida strain AKMP7. They reported that strain AKMP7 promoted growth of wheat in terms of root length, shoot length, biomass, tiller, and spikelet number and promoted grain formation through reducing membrane injury and improving the activity of SOD, APX, and CAT and various metabolites like chlorophyll, proline, protein, and carbohydrates (Ali et al. 2011). The heat shock protein could be a key factor in maintaining the functional conformation of the protein under heat (Wang et al. 2004). Among HSP, HSP17.8 is highly upregulated in wheat seedlings under heat stress; however, the response to bacterial treatment may vary between different cultivars and the bacterial strains. Interestingly, the expression of HsfA3 was downregulated in Bacillus-treated wheat seedlings compared to untreated heat-loaded seedlings (Suzuki et al. 2008). In Arabidopsis, HsfA3 improved the production of several HSPs (Ikeda and Ohme-Takagi 2009) and regulated two independent pathways, suggesting that they play an integrated role in stress management (Liu and Charng 2013). El-Daim et al. (2014) reported that inoculation of wheat with these strains B. amyloliquefaciens strain UCMB5113 and A. brasilense strain NO40 improved heat tolerance through the transcript level of a stress-related gene. They reported that the preactivation of heat shock transcription factors had an important role in heat stress tolerance (El-Daim et al. 2014).
2.4.1.5 Cold Stress
Low temperature is one of the most important abiotic stresses also known as cold or chilling stress. It limits crop productivity by damaging various physiological and biochemical cell functioning with visible symptoms (Ruelland and Zachowski 2010). Cold stress slows down the plant metabolism by damaging cellular macromolecules and membrane function, clotting of the cell membrane, and alteration in lipid composition and induces synthesis of compatible solutes, proteins, and carbohydrate metabolism (Welti et al. 2002; Baek and Skinner 2003). Chilling stress forms extracellular ice crystals leading to frozen dehydration and concentration of cellular juice and causes several other mechanical effects (Mishra et al. 2012).
Adaptation to cold stress tolerance encompasses a range of biochemical, molecular, and metabolic processes (Zhu et al. 2007; Ritonga and Chen 2020). Plants in the first attempt try to resist changes in cellular metabolism due to lower temperatures. Further, plant regulatory mechanisms are activated and restore levels and normal flow of metabolites (Fernie et al. 2005). They maintain homeostasis to gain tolerance to freezing and implying gene expression (Cook et al. 2004). The cold stress adaptation process involves changes in gene expression, membrane lipids, protein content, soluble proteins, sugar, proline, total phenols, deoxygenase, low leakage of ions from the cell membrane; changes in the accumulation of anthocyanins; and production of metabolites, including osmotic protective agents, and allows osmosis (Kasuga et al. 2004; Barka et al. 2006; Tiryaki et al. 2019). Plant cells sense cold stress by cold-responsive membrane hardening induced by genes that contribute to cold acclimation of alfalfa and Brasicca napus (Sangwan et al. 2001).
A major portion of the earth is covered with a low-temperature environment that acts as a reservoir of cold-tolerant (psychrophilic) bacteria having a low metabolic activity (Kumar et al. 2019b). Various psychrophilic bacterial strains including Acinetobacter rhizosphaerae, Arthrobacter nicotianae, Bacillus sp., Bordetella sp., Brevundimonas terrae, Exiguobacterium acetylicum, Mesorhizobium sp., Pseudomonas cedrina, P. fluorescens, P. putida, Providencia sp., Pseudomonas corrugata, Pseudomonas fragi, Rhizobium leguminosarum, Serratia marcescens, Sinorhizobium meliloti, Stenotrophomonas, etc. having the ability to increase P and iron (Fe) availability under cold stress by producing siderophore and solubilizing P were reported previously (Prevost et al. 1999, 2003; Pandey et al. 2002, 2006; Katiyar and Goel 2004; Negi et al. 2005; Trivedi and Sa 2008; Gulati et al. 2009; Selvakumar et al. 2008, 2009; Verma et al. 2015; Yadav et al. 2015; Zubair et al. 2019; Araya et al. 2020; Tapia-Vázquez et al. 2020; Vega-Celedón et al. 2021). Recently, Awasthi et al. (2019), Gautam et al. (2019b), Kumar et al. (2019b), and Tiryaki et al. (2019) also reported cold-tolerant bacteria including Arthrobacter humicola, Brevibacillus invocatus, Brevibacterium frigoritolerans, Pseudomonas chlororaphis, P. fluorescens, Pseudomonas fragi, Pseudomonas proteolytica, Pseudomonas helmanticensis, Pseudomonas koreensis, Pseudomonas mandelii, and Viridibacillus arenosi isolated from various cold regions.
The psychrophilic bacteria alleviate cold injury in plants using various mechanisms. Such bacteria could be efficient enough to synthesize antifreeze proteins and have a strong affinity for the promotion of root growth (Sun et al. 1995). Cold-tolerant Bradyrhizobium japonicum was reported for its ability to promote nodule formation and nitrogen fixation in soybean under cold stress (Mishra et al. 2009). Barka et al. (2006) reported improvement in cold tolerance in grapevine seedlings due to inoculation with Burkholderia phytofirmans that reduce electrolyte leakage. Inoculation with endophytic B. phytofirmans strain PsJN improved cold stress tolerance through the process of antioxidant scavenging, modulation in carbohydrate metabolism, and trehalose metabolism in grapevine (Fernandez et al. 2012; Theocharis et al. 2012). Mishra et al. (2011) and Barka et al. (2006) reported that psychrophilic bacterial inoculation also improved sugar metabolism pathway, photosynthetic activity, total phenolic contents, and activation of genes related to C-repeat binding factor. Inoculation with strains of Azospirillum, Bacillus, and Raoultella spp. improved cold tolerance in wheat and barley by reducing freezing injury and sustaining antioxidant enzyme activity (Turan et al. 2013). Tomato inoculated with Pseudomonas frederiksbergensis strain OS261 and Pseudomonas vancouverensis strain OB155 showed improved cold tolerance due to enhanced cold-acclimatized gene expression, antioxidant activity, proline synthesis, and reduction in membrane damage (Subramanian et al. 2015, 2016). Production of phytohormones by Serratia nematodiphila improved cold tolerance in pepper seedling (Kang et al. 2015). Inoculation with strains of Bacillus and Pseudomonas spp. in Phaseolus vulgaris has been reported to improve cold tolerance in plants by regulating freezing injury, ice-nucleating activity, lipid peroxidation, and activities of SOD, CAT, and POD (Tiryaki et al. 2019). So, it is concluded that plant interaction with cold-tolerant bacteria under cold stress environment can sustainably engineer cold stress (Kushwaha et al. 2020).
2.4.2 Biotic Stress
The PGPR help in improving the plant growth by suppressing the growth of phytopathogens, and weeds thus acting as a biocontrol and bioherbicide agents (Adetunji et al. 2019; Khan et al. 2020c). They use the indirect mechanisms of action to promote plant growth through alleviating biotic stresses due to growth of deleterious organisms (Arya et al. 2018). PGPR as biocontrol agents are environment-friendly supplements to treat plant diseases and to reduce the use of chemicals in agriculture (Gerhardson 2002). They control pathogens through pathogen exclusion based on bacterial capacity to colonize faster and effectively and by reducing the nutrient availability to harmful pathogens (Table 2.2). PGPR, for example, limit Fe availability to pathogenic microbes by production of siderophores that chelate Fe, making it unavailable for use by pathogens (Singh et al. 2011). They also produce a heterogeneous group of organic compounds called antibiotics which are lethal to various microorganisms even at low concentrations (Haas and Defago 2005). They reduce the pathogen attack by provoking a defense response in plants known as induced systemic resistance (ISR) involving synthesis of various defense metabolites that modify physical and biochemical properties of host without causing disease (Bent 2006; Bukhat et al. 2020).
PGPR produce antibiotics that are bioactive metabolites having the ability to inhibit growth of disease-causing agents. Under stress, PGPR secrete antibiotics having varying degrees of specificity and mode of action (Pathak et al. 2017). Pseudomonas sp., for example, can produce antibiotics including 2,4-diacetylphloroglucinol, amphisin, phenazine, phenzine, pyoluteorin, pyrrolnitrin, tensin, and tropolone (Compant et al. 2005; Showkat 2012). Many Pseudomonas strains produce pyrrolnitrin antibiotic which is a broad-spectrum antifungal compound. Similarly, in another study, Perneel et al. (2008) reported that Pseudomonas aeruginosa and P. chlororaphis can produce phenazine, which is a heterocyclic, broad-spectrum antibiotic used in biocontrol against phytopathogens. Two broad-spectrum antibiotics called kanosamine and zwittermicin are produced by B. cereus that act against damping-off of the alfalfa seedlings (Lozano et al. 2016). Iturins with β-amino fatty acid produced by strains of Bacillus sp. exhibit strong antifungal activity and rely predominantly on the ability to augment membrane permeability due to strong interaction with the phospholipids and sterols (Alvarez et al. 2012). Microbial HCN is involved in biocontrol as well as sequestration of metal ions and consequently increases the availability of nutrients which is beneficial both for the rhizobacteria and plant host (Rijavec and Lapanje 2016; Meena et al. 2020).
Microbial production of siderophores has a central role in the development of a plant (Beneduzi et al. 2012). Siderophores are one of the strongest carriers of Fe3+ in combination with enterobactin (Saharan and Nehra 2011). The production of siderophores is affected by the type of iron ion; pH; enough availability of N, C, and P; and other trace elements (Compant et al. 2005). Pseudobactin or pyoverdine have been reported to be produced by P. fluorescens, which have a high affinity for sequestration of Fe2+. The ferripyoverdins (a complex of pyoverdine with Fe3+) are reported to be combined with the receptors of the outer membrane and transported to the cytoplasm and ultimately get reduced to Fe2+ (Haas and Defago 2005). The plant growth is directly increased by production of the siderophore as they make Fe available for plants. It has been reported that PGPR also consumes Fe for effective rhizospheric colonization. PGPR produce lytic enzymes to degrade cells of the pathogens (Compant et al. 2005). The synergistic effects of PGPR metabolites such as antibiotics and lytic enzymes are responsible for the degradation of cell walls of microbial pathogen and weeds (Dunne et al. 1998).
In response to phytopathogens, PGPR help plants stimulate a defense mechanism having a hypersensitive reaction called ISR (Compant et al. 2005). Numerous Bacillus sp. trigger ISR in biotic stress that improves plant growth through ultrastructural changes and cytochemical alteration in plants (Kloepper et al. 2004). The bacteriocins produced by PGPR are different from narrow-spectrum antibiotics that can kill closely related bacterial strains (Beneduzi et al. 2012). The bio-surfactants are considered to be effective for the biocontrol of phytium species (Perneel et al. 2008). For example, P. fluorescens is capable of protecting plants from disease-causing pathogenic fungi. It secretes a varying degree of exoproducts through antibiosis, phytohormone activities, lytic activities, and antimicrobial compounds to be considered as a primary mechanism to suppress disease in root area (Péchy-Tarr et al. 2008). Such PGPR play a major role in biocontrol by producing a wide range of secondary metabolites including the production of volatile compounds, antibiotics, siderophores, HCN, and other disease-suppressing compounds (Showkat 2012).
Numerous microorganisms have been reported as potential bioherbicides. PGPR are the most suitable biocontrol agents as compared to other microorganisms due to their rapid growth and genetic variation via mutagenesis (Johnson et al. 1996; Li et al. 2003). Various strains of P. fluorescens have shown their benefits as well as inhibitory effects on plants (Banowetz et al. 2008). Their suppressive effects were reported due to production of extracellular metabolites that suppress plant germination (Banowetz et al. 2008). Growth inhibition of various grassy weeds including downy brome was observed through inoculation with P. fluorescens strain D7 (Kennedy et al. 2001). These strains showed plant growth inhibition due to combined production of extracellular peptides and lipopolysaccharides (Gurusiddaiah et al. 1994). However, their mechanism for weed growth suppression has not yet been described. P. fluorescens strain WH6 showed growth inhibition of 21 monocot and 8 dicot plant species; however it was unable to affect the germination of hybrid corn (Banowetz et al. 2008). Its ability to suppress weed germination was reported due to production of germination arrest factor (GAF) composed of 4-formylaminooxy-L-vinylglycine and oxyvinylglycines due to synthesis of their regulatory genes (Banowetz et al. 2008; McPhail et al. 2010; Halgren et al. 2013; Okrent et al. 2014). The strain BRG100 showed suppression of Setaria viridis due to the production of extracellular metabolites (pseudophomin A and B) having phytotoxic effects (Caldwell et al. 2012).
Pseudomonas strains produced phytotoxins including phenazine-1-carboxylic acid, 2-aminophenoxazone, and 2-aminophenol that have allelopathic effects against plant growth (Adetunji et al. 2019). The strains of Xanthomonas campestris were also reported as a potential weed control agent. Its strain JT-P482 showed specific control against grasses Poa annua and Poa attenuate (Imaizumi et al. 1997). The strain LVA-987 also showed weed control against horseweed (Boyette and Hoagland 2015). No information on phytotoxic compounds produced by these strains is available in support of its application as a bioherbicide. Such strains spread in their host plant through colonizing plant xylem. They interact with host plant through their receptor proteins that can recognize pathogen-associated molecular patterns that activates plant defense responses in terms of production of ROS and programmed cell death (Guy et al. 2013; Dugé de Bernonville et al. 2014). Such allelopathic bacteria may also produce enzymes and broad-spectrum antibiotics that have inhibitory effects on plant growth (Abbas et al. 2017). Production of elevated IAA levels by PGPR also possesses allelopathic effects (Sarwar and Kremer 1995). The HCN production by rhizobacteria is a major suppressor of weed growth (Kremer and Souissi 2001).
Recent developments in bioherbicides support the potential use of HCN-producing bacterial strains along with multiple PGP traits for improving crop growth by suppression of weeds. In a recent study, Dar et al. (2020) reported Pseudomonas strains with potential Pseudomonas for both potential to control Phalaris minor and Avena fatua in wheat. They reported four Pseudomonas strains having HCN-producing ability along with multiple PGP traits such as P solubilization, ACC-deaminase activity, siderophores production, and oxidase activity during in vitro studies. In the bioassay, they reported these strains as strong phytotoxic for lettuce seedling having a mortality rate of up to 73.3%. Consortia of these strains were more effective than sole inoculation in the biocontrol of Pectoralis minor and Avena fatua seedlings. In addition to biocontrol of weeds, these strains were also effective in improving wheat growth. They argued that the use of these strains as bioherbicide may improve human and environmental health. So, inoculation with allelopathic bacteria could support the existing herbicides that not only control weeds but also alleviate the allelopathic effects of weeds. The mechanisms of biotic stress amelioration by PGPR have been summarized in Fig. 2.2.

Mechanisms of biocontrol by plant growth-promoting rhizobacteria
2.4.3 Integrated Stress Amelioration
Abiotic and biotic stresses due to climate change are a major cause of productivity loss in major crops (Anderson et al. 2020). The effects of abiotic and biotic stresses on plant growth and their amelioration by PGPR have already been discussed in the above sections. Under field conditions, crops face multiple stresses which drastically decline crop productivity (Pandey et al. 2017; Hussain et al. 2018). Managing abiotic and biotic stresses with competent and cost-effective approaches is a major challenge. Such approaches including the development of stress-tolerant varieties and application of chemical fertilizers and pesticides are cost-intensive. Recently, the application of PGPR to ameliorate various environmental and biotic stresses has been revealed as a cost-effective and efficient approach in integrated stress management (Grover et al. 2011). We have proposed a possible mechanism of action in the amelioration of integrated stresses (Fig. 2.3).

Proposed mechanisms of action for integrated stress amelioration by plant growth-promoting rhizobacteria
As described in the above sections, the PGPR having the ability to produce ACC-deaminase, phytohormones, antioxidant enzymes, siderophores, EPS, VOCs, and solubilization of nutrients can ameliorate all the abiotic stresses. Similarly, in biotic stress control, the HCN-producing PGPR along with production of antibiotics, lytic enzymes, siderophores, and ISR are effective (Fig. 2.3).
The PGPR with multiple PGP traits can be effective in integrated stress amelioration (ISA). As a combination of stresses enhance the ethylene production, ACC-deaminase-producing PGPR with multiple PGP traits can be more effective in ISA. For example, recent studies reported the effectiveness of ACC-deaminase-producing PGPR in ISA (Barnawal et al. 2017; Srinivasan et al. 2017; Saikia et al. 2018; Orozco-Mosqueda et al. 2020; Li et al. 2020). Some mechanisms of stress mitigation by PGPR including the production of siderophores, nutrient mineralization, production of VOCs, and EPS are not well understood in all abiotic stresses; however, these metabolic activities have been reported to be effective in various environmental stresses (Talaat and Shawky 2017; Khan and Bano 2019). PGPR release secondary metabolites including antibiotics, siderophores, hydrolytic enzymes, and HCN that mitigate the biotic stresses in terms of infection by pathogens and allelopathic effects of weed as well as help improve crop productivity (Jha 2018). As reported by Dar et al. (2020), HCN-producing rhizobacteria with multiple PGP traits can be strong candidates for integrated stress amelioration.
2.5 Conclusion and Prospects
Under natural field conditions, plants face several abiotic and biotic stresses which negatively affect plant growth. These stresses result in the enhanced biosynthesis of ethylene and damage the plant physiological and biochemical attributes. The application of PGPR is well reported for mitigation of these stresses. These beneficial rhizobacteria directly mitigate integrated stresses by metabolic activities in terms of production of phytohormones, ACC-deaminase, siderophores, and EPS and ensure nutrient availability through improving their solubility. PGPR have an indirect role as well in stress alleviation by producing antibiotics, HCN, and hydrolytic enzymes and induce ISR that help plant tolerate biotic stresses.
Improvement in PGPR population and diversity through rhizosphere engineering or use of microbial inoculants and/or their metabolites should be practiced for better agroecosystem and soil health. There is a need to study interactive effects of various stresses on a plant to know about impacts of combined stresses on crop production. Future research is needed to evaluate the role of PGPR under multiple stress conditions.
Future research should also be focused on understanding the mechanisms involved in PGPR-induced stress tolerance in crop plants. There is also a need to investigate why certain PGPR with specific PGP traits could not induce stress tolerance in crop plants under soil (field) and environmental conditions. Strategies should be developed to study and improve plant-microbe interactions using bioinformatics, modeling tools, and other molecular techniques to enhance crop stress tolerance and for sustainable agroecosystems.
References
Aasfar A, Bargaz A, Yaakoubi K, Hilali A, Bennis I, Zeroual Y, Meftah Kadmiri I (2021) Nitrogen fixing azotobacter species as potential soil biological enhancers for crop nutrition and yield stability. Front Microbiol 12:354. https://doi.org/10.3389/fmicb.2021.628379
Abbas T, Zahir ZA, Naveed M (2017) Bioherbicidal activity of allelopathic bacteria against weeds associated with wheat and their effects on growth of wheat under axenic conditions. Bio Control 62(5):719–730
Abd El-Daim IA, Bejai S, Meijer J (2014) Improved heat stress tolerance of wheat seedlings by bacterial seed treatment. Plant Soil 379(1):337–350
Abdelrahman M, El-Sayed M, Jogaiah S, Burritt DJ, Tran LSP (2017) The “STAY-GREEN” trait and phytohormone signaling networks in plants under heat stress. Plant Cell Rep 36(7):1009–1025
Adak MK, Sarkar B, De AK, Saha I, Ghosh A (2020) Alleviation of abiotic stress by nonconventional plant growth regulators in plant physiology. In: Rakshit A, Singh HB, Singh AK, Singh US, Fraceto L (eds) New frontiers in stress management for durable agriculture. Springer, Singapore, pp 197–211
Adetunji CO, Oloke JK, Bello OM, Pradeep M, Jolly RS (2019) Isolation, structural elucidation and bioherbicidal activity of an eco-friendly bioactive 2-(hydroxymethyl) phenol, from Pseudomonas aeruginosa (C1501) and its ecotoxicological evaluation on soil. Environ Technol Innov 13:304–317
Ahammed GJ, Li X, Zhou J, Zhou YH, Yu JQ (2016) Role of hormones in plant adaptation to heat stress. In: Ahammed GJ, Yu JQ (eds) Plant hormones under challenging environmental factors. Springer, Dordrecht, pp 1–21
Ahmad M, Zahir ZA, Asghar HN, Asghar M (2011) Inducing salt tolerance in mung bean through co-inoculation with Rhizobium and PGPR containing ACC-deaminase. Can J Microbiol 57(7):578–589
Ahmad M, Zahir ZA, Khalid M, Nazli F, Arshad M (2013a) Efficacy of Rhizobium and Pseudomonas strains to improve physiology, ionic balance and quality of mung bean under salt-affected conditions on farmer’s fields. Plant Physiol Biochem 63:170–176
Ahmad M, Zahir ZA, Nazli F, Akram F, Arshad M, Khalid M (2013b) Effectiveness of halo-tolerant, auxin producing Pseudomonas and Rhizobium strains to improve osmotic stress tolerance in mung bean (Vigna radiata L.). Braz J Microbiol 44(4):1341–1348
Ahmad M, Nadeem S, Naveed M (2016) Potassium-solubilizing bacteria and their application in agriculture. In: Meena VS, Maurya BR, Verma JP, Meena RS (eds) Potassium solubilizing microorganisms for sustainable agriculture. Springer, New Delhi, pp 293–313
Ahmad M, Naseer I, Hussain A, Mumtaz MZ, Mustafa A, Hilger TH, Zahir ZA, Xu M (2019a) Appraising endophyte-plant symbiosis for improved growth, nodulation, nitrogen fixation and abiotic stress tolerance: an experimental investigation with chickpea (Cicer arietinum L.). Agronomy 9:621. https://doi.org/10.3390/agronomy9100621
Ahmad I, Zaib S, Alves PC, Luthe DS, Bano A, Shakeel SN (2019b) Molecular and physiological analysis of drought stress responses in Zea mays treated with plant growth promoting rhizobacteria. Biol Plant 63:536–547
Ahmadi N, Mibus H, Serek M (2009) Characterization of ethylene-induced organ abscission in F1 breeding lines of miniature roses (Rosa hybrida L.). Postharvest Biol Technol 52:260–266
Ahmed S, Nawata E, Sakuratani T (2006) Changes of endogenous ABA and ACC, and their correlations to photosynthesis and water relations in mungbean (Vigna radiata L.) Wilczak cv. KPS1 during waterlogging. Environ Exp Bot 57(3):278–284
Akhgar R, Arzanlou M, Bakker PA, Hamidpour M (2014) Characterization of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-containing Pseudomonas sp. in the rhizosphere of salt-stressed canola. Pedosphere 24:161–468
Al-Hussini HS, Al-Rawahi AY, Al-Marhoon AA, Al-Abri SA, Al-Mahmooli IH, Al-Sadi AM, Velazhahan R (2019) Biological control of damping-off of tomato caused by Pythium aphanidermatum by using native antagonistic rhizobacteria isolated from Omani soil. J Plant Pathol 101(2):315–322
Ali S, Kim WC (2018) Plant growth promotion under water: decrease of waterlogging-induced ACC and ethylene levels by ACC deaminase-producing bacteria. Front Microbiol 9:1096
Ali SZ, Sandhya V, Grover M, Kishore N, Rao LV, Venkateswarlu B (2009) Pseudomonas sp. strain AKM-P6 enhances tolerance of sorghum seedlings to elevated temperatures. Biol Fertil Soils 46:45–55
Ali SZ, Sandhya V, Grover M, Linga VR, Bandi V (2011) Effect of inoculation with a thermotolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticum spp.) under heat stress. J Plant Interact 6(4):239–246
Ali S, Khan MA, Kim WC (2018) Pseudomonas veronii KJ mitigates flood stress-associated damage in Sesamum indicum L. Appl Biol Chem (5):575–585
Ali Q, Shahid S, Nazar N, Hussain AI, Ali S, Chatha SA, Perveen R, Naseem J, Haider MZ, Hussain B, Hussain SM (2020) Use of phytohormones in conferring tolerance to environmental stress. In: Hasanuzzaman M (ed) Plant ecophysiology and adaptation under climate change: mechanisms and perspectives II. Springer, Singapore, pp 245–355
Ali M, Lou Y, Hafeez R, Li X, Hossain A, Xie T, Lin L, Li B, Yin Y, Yan J, An Q (2021) Functional analysis and genome mining reveal high potential of biocontrol and plant growth promotion in nodule-inhabiting bacteria within Paenibacillus polymyxa complex. Front Microbiol 11:3627
Alvarez F, Castro M, Principe A, Borioli G, Fischer S, Mori G, Jofre E (2012) The plant-associated Bacillus amyloliquefaciens strains MEP218 and ARP23 capable of producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in biocontrol of sclerotinia stem rot disease. J Appl Microbiol 112(1):159–174
Amna DBU, Sarfraz S, Xia Y, Kamran MA, Javed MT, Sultan T, Munis MF, Chaudhary HJ (2019) Mechanistic elucidation of germination potential and growth of wheat inoculated with exopolysaccharide and ACC-deaminase producing Bacillus strains under induced salinity stress. Ecotoxicol Environ Saf 183:109466
Anderson R, Bayer PE, Edwards D (2020) Climate change and the need for agricultural adaptation. Curr Opin Plant Biol 56:197–2020. https://doi.org/10.1016/j.pbi.2019.12.006
Ansari FA, Jabeen M, Ahmad I (2021) Pseudomonas azotoformans FAP5, a novel biofilm-forming PGPR strain, alleviates drought stress in wheat plant. Int J Environ Sci Technol 3:1–6
Araya MA, Valenzuela T, Inostroza NG, Maruyama F, Jorquera MA, Acuña JJ (2020) Isolation and characterization of cold-tolerant hyper-acc-degrading bacteria from the rhizosphere, endosphere, and phyllosphere of Antarctic vascular plants. Microorganisms 8(11):1788
Arif F, Ghoul M (2018) Halotolerance of indigenous fluorescent Pseudomonads in the presence of natural osmoprotectants. Annu Res Rev Biol 24(4):1–11
Arkhipova T, Veselov S, Melentiev A, Martynenko EV, Kudoyarova GR (2005) Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 272(1–2):201–209
Armada E, Roldan A, Azcon R (2014) Differential activity of autochthonousbacteria in controlling drought stress in native Lavandula and Salvia plants species under drought conditions in natural arid soil. Microb Ecol 67:410–420
Armstrong W, Drew MC (2002) Root growth and metabolism under oxygen deficiency. In: Waisel Y, Eshel A, Kafkafi U (eds) Plant roots: the hidden half, 3rd edn. Marcel Dekker, New York, pp 729–761
Armstrong W, Brändle R, Jackson MB (1994) Mechanisms of flood tolerance in plants. Acta Bot Neerl 43(4):307–358
Arya B, Komala BR, Sumalatha NT, Surendra GM, Gurumurthy PR (2018) PGPR induced systemic tolerance in plant. Int J Curr Microbiol App Sci 7:453–462
Asada K (2006) Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 141(2):391–396
Asghari B, Khademian R, Sedaghati B (2020) Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci Hort 263:109132
Ashraf MA, Akbar A, Askari SH, Iqbal M, Rasheed R, Hussain I (2018) Recent advances in abiotic stress tolerance of plants through chemical priming: an overview. In: Rakshit A, Singh HB (eds) Advances in seed priming. Springer, Singapore, pp 51–79
Aslam F, Ali B (2018) Halotolerant bacterial diversity associated with Suaeda fruticosa (L.) forssk. Improved growth of maize under salinity stress. Agronomy 8:131
Aslam H, Ahmad SR, Anjum T, Akram W (2018) Native halotolerant plant growth promoting bacterial strains can ameliorate salinity stress on tomato plants under field conditions. Int J Agric Biol 20:315–322
Aslam MU, Raza MA, Saleem MF, Waqas M, Iqbal R, Ahmad S, Haider I (2020) Improving strategic growth stage-based drought tolerance in quinoa by rhizobacterial inoculation. Commun Soil Sci Plant Anal 51(7):853–868
Avramova V, AbdElgawad H, Zhang Z, Fotschki B, Casadevall R, Vergauwen L, Knapen D, Taleisnik E, Guisez Y, Asard H, Beemster GT (2015) Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol 169(2):1382–1396
Awasthi S, Sharma A, Saxena P, Yadav J, Pandiyan K, Kumar M, Singh A, Chakdar H, Bhowmik A, Kashyap PL, Srivastava AK (2019) Molecular detection and in silico characterization of cold shock protein coding gene (cspA) from cold adaptive Pseudomonas koreensis. J Plant Biochem Biotechnol 28:405–413
Baek KH, Skinner DZ (2003) Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci 165(6):1221–1227
Bal HB, Adhya TK (2021) Alleviation of submergence stress in rice seedlings by plant growth-promoting rhizobacteria with ACC-deaminase activity. Front Sust Food Syst 5:36
Balal RM, Shahid MA, Javaid MM, Iqbal Z, Anjum MA, Garcia-Sanchez F, Mattson NS (2016) The role of selenium in amelioration of heat-induced oxidative damage in cucumber under high temperature stress. Acta Physiol Plant 38(6):158. https://doi.org/10.1007/s11738-016-2174-y
Bangash N, Khalidi A, Mahmood T, Siddique TM (2013) Screening rhizobacteria containing ACC deaminase for growth promotion of wheat under water stress. Pak J Bot 45(SI):91–96
Banowetz GM, Azevedo MD, Armstrong DJ, Halgren AB, Mills DI (2008) Germination-Arrest Factor (GAF): biological properties of a novel, naturally-occurring herbicide produced by selected isolates of rhizosphere bacteria. Biol Control 46:380390
Barka EA, Nowak J, Clément C (2006) Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl Environ Microbiol 72(11):7246–7252
Barnawal D, Bharti N, Maji D, Chanotiya CS, Kalra A (2012) 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant Physiol Biochem 58:227–235
Barnawal D, Bharti N, Pandey SS, Pandey A, Chanotiya CS, Kalra A (2017) Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol Plant 161(4):502–514
Belimov AA, Dodd IC, Hontzeas N, Theobald JC, Safronova VI, Davies WJ (2009) Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylatedeaminase increase yield of plants grown in drying soil via both local andsystemic hormone signaling. New Phytol 181:413–423
Belimov AA, Dodd IC, Safronova VI, Dumova VA, Shaposhnikov AI, Ladatko AG, Davies WJ (2014) Abscisic acid metabolizing rhizobacteria decrease ABA concentrations in planta and alter plant growth. Plant Physiol Biochem 74:84–91
Beneduzi A, Ambrosini A, Passaglia LMP (2012) Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet Mol Biol 35(4):1044–1051
Bent E (2006) Induced systemic resistance mediated by plant growth-promoting rhizobacteria (PGPR) and fungi (PGPF). In: Tusuz S, Bent E (eds) Multigenic and induced systemic resistance in plants. Springer, Boston, MA, pp 225–258
Bhardwaj D, Ansari MW, Sahoo RK, Tuteja N (2014) Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb Cell Fact 3(66):1–10
Bharti N, Pandey SS, Barnawal D, Patel VK, Kalra A (2016) Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci Rep 6:34768. https://doi.org/10.1038/srep34768
Bhattacharyya PN, Jha DK (2012) Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microbiol Biotechnol 28:1327–1350
Bhattacharyya D, Garladinne M, Lee YH (2015) Volatile indole produced by rhizobacterium Proteus vulgaris JBLS202 stimulates growth of Arabidopsis thaliana through auxin, cytokinin, and brassinosteroid pathways. J Plant Growth Regul 34:158–168
Bottner P, Couteaux MM, Vallejo VR (1995) Soil organic matter in Mediterranean-type ecosystems and global climatic changes: a case study-thesoils of the Mediterranean basin. In: Jose M, Oechel WC (eds) Global change and mediterranean-type ecosystems. Springer-Verlag, New York, pp 306–325
Boyette CD, Hoagland RE (2015) Bioherbicidal potential of Xanthomonas campestris for controlling Conyza canadensis. Biocontrol Sci Technol 25:229–237
Braud A, Jézéquel K, Bazot S, Lebeau T (2009) Enhanced phytoextraction of an agricultural Crand Pb-contaminated soil by bioaugmentation with siderophore-producing bacteria. Chemosphere 74:280–286
Brown AJP, Cowen LE, di Pietro A, Quinn J (2017) Stress adaptation. Fungal Kingdom 1:463–485
Brunetti C, Tattini M, Guidi L, Velikova V, Ferrini F, Fini A (2019) An integrated overview of physiological and biochemical responses of Celtis australis to drought stress. Urban For Urban Green 46:126480. https://doi.org/10.1016/j.ufug.2019.126480
Bruno LB, Karthik C, Ma Y, Kadirvelu K, Freitas H, Rajkumar M (2020) Amelioration of chromium and heat stresses in Sorghum bicolor by Cr6+ reducing-thermotolerant plant growth promoting bacteria. Chemosphere 244:125521
Bukhat S, Imran A, Javaid S, Shahid M, Majeed A, Naqqash T (2020) Communication of plants with microbial world: exploring the regulatory networks for PGPR mediated defense signaling. Microbiol Res 1:126486
Buttar ZA, Wu SN, Arnao MB, Wang C, Ullah I, Wang C (2020) Melatonin suppressed the heat stress-induced damage in wheat seedlings by modulating the antioxidant machinery. Plants 9(7):809
Caldwell CJ, Hynes RK, Boyetchko SM, Korber DR (2012) Colonization and bioherbicidal activity on green foxtail by Pseudomonas fluorescens BRG100 in a pesta formulation. Can J Microbiol 58:1–9
Carbonell-Bojollo R, Veroz-Gonzalez O, Ordoñez-Fernandez R, Moreno-Garcia M, Basch G, Kassam A, Repullo-Ruiberriz de Torres MA, Gonzalez-Sanchez EJ (2019) The effect of conservation agriculture and environmental factors on CO2 emissions in a rainfed crop rotation. Sustainability 11(14):3955. https://doi.org/10.3390/su11143955
Chatterjee P, Kanagendran A, Samaddar S, Pazouki L, Sa TM, Niinemets Ü (2020) Influence of Brevibacterium linens RS16 on foliage photosynthetic and volatile emission characteristics upon heat stress in Eucalyptus grandis. Sci Total Environ 700:134453
Chen M, Wei H, Cao J, Liu R, Wang Y, Zheng C (2007) Expression of Bacillus subtilis proBA genes and reduction of feedback inhibition of proline synthesis increases proline production and confers osmotolerance in transgenic Arabidopsis. J Biochem Mol Biol 40:396–403
Chen L, Liu Y, Wu G, Veronican NK, Shen Q, Zhang N, Zhang R (2016) Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol Plant 158:34–44
Chen ZY, Abuduaini X, Mamat N, Yang QL, Wu MJ, Lin XR, Wang R, Lin RR, Zeng WJ, Ning HC, Zhao HP (2021) Genome sequencing and functional annotation of Bacillus sp. strain BS-Z15 isolated from cotton rhizosphere soil having antagonistic activity against Verticillium dahliae. Arch Microbiol 5:1–1
Cherif H, Marasco R, Rolli E, Ferjani R, Fusi M, Soussi A, Mapelli F, Blilou I, Borin S, Boudabous A, Cherif A (2015) Oasisdesert farming selects environment-specific date palm root endophyticcommunities and cultivable bacteria that promote resistance to drought. Environ Microbiol Rep 7:668–678
Chiappero J, del Rosario CL, Alderete LG, Palermo TB, Banchio E (2019) Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind Crop Prod 139:111553
Compant S, Duffy B, Nowak J, Clément C, Barka EA (2005) Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71(9):4951–4959
Cook D, Fowler S, Fiehn O, Thomashow MF (2004) A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis. Proc Natl Acad Sci 101(42):15243–15248
Coupland D, Jackson MB (1991) Effects of mecoprop (an auxin analogue) on ethylene evolution and epinasty in two biotypes of stellaria media. Ann Bot 68:167–172
Cramer GR, Urano K, Delrot S, Pezzotti M, Shinozaki K (2011) Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biol 11(1):163. https://doi.org/10.1186/1471-2229-11-163
Creus CM, Sueldo RJ, Barassi CA (2004) Water relations and yield in Azospirillum-inoculated wheat exposed to drought in the field. Can J Bot 82:273–281
Cui L, Yang C, Wei L, Li T, Chen X (2020) Isolation and identification of an endophytic bacteria Bacillus velezensis 8-4 exhibiting biocontrol activity against potato scab. Biol Control 141:104156
Danish S, Zafar-ul-Hye M (2019) Co-application of ACC-deaminase producing PGPR and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Sci Rep 9(1):1–3
Danish S, Zafar-ul-Hye M, Mohsin F, Hussain M (2020) ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS One 15(4):e0230615
Dar A, Zahir ZA, Asghar HN, Ahmad R (2020) Preliminary screening of rhizobacteria for biocontrol of little seed canary grass (Phalaris minor Retz.) and wild oat (Avena fatua L.) in wheat. Can J Microbiol 66:368–376
Daranas N, Roselló G, Cabrefiga J, Donati I, Francés J, Badosa E, Spinelli F, Montesinos E, Bonaterra A (2019) Biological control of bacterial plant diseases with Lactobacillus plantarum strains selected for their broad-spectrum activity. Ann Appl Biol 174(1):92–105
Datir SS, Kochle M, Jindal S (2019) Molecular approaches and salt tolerance mechanisms in leguminous plants. In: Akhtar MS (ed) Salt stress, microbes, and plant interactions: mechanisms and molecular approaches. Springer, Singapore, pp 49–67
Dawood T, Yang X, Visser EJ, Te Beek TA, Kensche PR, Cristescu SM, Lee S, Floková K, Nguyen D, Mariani C, Rieu I (2016) A coopted hormonal cascade activates dormant adventitious root primordia upon flooding in Solanum dulcamara. Plant Physiol 170:2351–2364
del Amor FM, Cuadra-Crespo P (2012) Plant growth-promoting bacteria as a tool to improve salinity tolerance in sweet pepper. Funct Plant Biol 39:82–90
Dimkpa C, Weinand T, Asch F (2009) Plant-rhizobacteria interactions alleviateabiotic stress conditions. Plant Cell Environ 32:1682–1694
Dodd IC, Belimov AA, Sobeih WY, Safronova VI, Grierson D, Davies WJ (2004) Will modifying plant ethylene status improve plant productivity in water-limited environments. In: Handbook and Abstracts for the 4th International Science Congress, Brisbane, Australia, p 134
Dodd IC, Zinovkina NY, Safronova VI, Belimov AA (2010) Rhizobacterial mediation of plant hormone status. Ann Appl Biol 157:361–379
Dubey A, Verma AK (2019) Salinity stress-dependent coordination of metabolic networks in relation to salt tolerance in plants. In: Giri B, Verma A (eds) Microorganisms in saline environments: strategies and functions. Springer, Cham, pp 401–422
Dugé de Bernonville T, Noël LD, SanCristobal M, Danoun S, Becker A, Soreau P, Arlat M, Lauber E (2014) Transcriptional reprogramming and phenotypical changes associated with growth of Xanthomonas campestris pv. campestris in cabbage xylem sap. FEMS Microbiol Ecol 89:527–541
Dunne C, Moënne-Loccoz Y, McCarthy J, Higgins P, Powell J, Dowling DN, O’Gara F (1998) Combining proteolytic and phloroglucinol-producing bacteria for improved biocontrol of Pythium-mediated damping-off of sugar beet. Plant Pathol 47(3):299–307
Egamberdieva D (2009) Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol Plant 31:61–864
Egamberdieva D (2013) The role of phytohormone producing bacteria inalleviating salt stress in crop plants. In: Miransari M (ed) Biotechnological techniques of stress tolerance in plants. Stadium Press LLC, Houston, TX, pp 21–39
Egamberdieva D, Lugtenberg B (2014) Use of plant growth-promoting rhizobacteria to alleviate salinity stress in plants. PGPR to alleviate salinity stress on plant growth. In: Miransari M (ed) Use of microbes for the alleviation of soil stresses. Springer, New York, pp 73–96
Egamberdieva D, Wirth SJ, Alqarawi AA, AbdAllah EF, Hashem A (2017) Phytohormones and beneficial microbes: essential components for plants to balance stress and fitness. Front Microbiol 8:2104. https://doi.org/10.3389/fmicb.2017.02104
Eida AA, Bougouffa S, L’Haridon F, Alam I, Weisskopf L, Bajic VB, Saad MM, Hirt H (2020) Genome insights of the plant-growth promoting bacterium Cronobacter muytjensii JZ38 with volatile-mediated antagonistic activity against Phytophthora infestans. Front Microbiol 11:369
El-Daim IAA, Bejai S, Meijer J (2014) Improved heat stress tolerance of wheat seedlings by bacterial seed treatment. Plant Soil 379(1–2):337–350
Etesami H, Alikhani HA (2019) Halotolerant plant growth-promoting fungi and bacteria as an alternative strategy for improving nutrient availability to salinity-stressed crop plants. In: Kumar M, Estesami H, Kumar V (eds) Saline soil-based agriculture by halotolerant microorganisms. Springer, Singapore, pp 103–146
Etesami H, Hosseini HM, Alikhani HA (2014) Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol Mol Biol Plants 20(4):425–434
Fahad S, Hussain S, Bano A, Saud S, Hassan S, Shan D, Khan FA, Khan F, Chen Y, Wu C, Tabassum MA (2015) Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: consequences for changing environment. Environ Sci Pollut Res 22:4907–4921
Fahad S, Hussain S, Saud S, Hassan S, Tanveer M, Ihsan MZ, Shah AN, Ullah A, Khan F, Ullah S, Alharby H (2016) A combined application of biochar and phosphorus alleviates heat-induced adversities on physiological, agronomical and quality attributes of rice. Plant Physiol Biochem 103:191–198
Fallahzadeh-Mamaghani V, Golchin S, Shirzad A, Mohammadi H, Mohamadivand F (2021) Characterization of Paenibacillus polymixa N179 as a robust and multifunctional biocontrol agent. Biol Control 154:104505
FAO Food and Agriculture Organization of the United Nations (2015) The impact of natural hazards and disasters on agriculture and food security and nutrition: a call for action to build resilient livelihoods. FAO, Rome
Farwell AJ, Vesely S, Nero V, Rodriguez H, McCormack K, Shah S, Dixon DG, Glick BR (2007) Tolerance of transgenic canola plants (Brassica napus) amended with plant growth-promoting bacteria to flooding stress at a metal-contaminated field site. Environ Pollut 147(3):540–545
Fernandez O, Vandesteene L, Feil R, Baillieul F, Lunn JE, Clément C (2012) Trehalose metabolism is activated upon chilling in grapevine and might participate in Burkholderia phytofirmans induced chilling tolerance. Planta 236(2):355–369
Fernie AR, Geigenberger P, Stitt M (2005) Flux an important, but neglected, component of functional genomics. Curr Opin Plant Biol 8(2):174–182
Figueiredo MV, Burity HA, Martínez CR, Chanway CP (2008) Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by coinoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl Soil Ecol 40:182–188
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 1:945–963
Flynn P (2003) Biotic vs. abiotic—distinguishing disease problems from environmental stresses. Hortic Home Pest News 489:22
Gan P, Liu F, Li R, Wang S, Luo J (2019) Chloroplasts- beyond energy capture and carbon fixation: tuning of photosynthesis in response to chilling stress. Int J Mol Sci 20(20):5046. https://doi.org/10.3390/ijms20205046
Garcia FP, Menendez E, Rivas R (2015) Role of bacterial bio fertilizers in agriculture and forestry. AIMS Bioeng 2:183–205
García JE, Maroniche G, Creus C, Suárez-Rodríguez R, Ramirez-Trujillo JA, Groppa MD (2017) In vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growth of maize under drought stress. Microbiol Res 202:21–29
Gautam S, Chauhan A, Sharma R, Sehgal R, Shirkot CK (2019a) Potential of Bacillus amyloliquefaciens for biocontrol of bacterial canker of tomato incited by Clavibacter michiganensis ssp. michiganensis. Microb Pathog 130:196–203
Gautam N, Sharma P, Rana JC, Singh M (2019b) Plant growth promoting traits of a novel psychrotrophic bacterium Virdibacillus arenosi PH15 isolated from rhizosphere of Podophyllum hexandrum. R Acad J Med Plants 7(1):013–019
Gerhardson B (2002) Biological substitutes for pesticides. Trends Biotechnol 20:338–343
Ghosh D, Sen S, Mohapatra S (2017) Modulation of proline metabolic gene expression in Arabidopsis thaliana under water-stressed conditions by a drought-mitigating Pseudomonas putida strain. Ann Microbiol 67(10):655–668
Gibbs J, Greenway H (2003) Mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Funct Plant Biol 30:1–47
Glick BR (2005) Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol Lett 251(1):1–7
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:963401. https://doi.org/10.6064/2012/963401
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169(1):30–39
Glick BR, Cheng Z, Czarny J, Duan J (2007) Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur J Plant Pathol 119:329–339
Gonzalez AJ, Larraburu EE, Llorente BE (2015) Azospirillum brasilense increased salt tolerance of Jojoba during in vitro rooting. Ind Crop Prod 76:41–48
Gouda S, Kerry RG, Das G, Paramithiotis S, Shin HS, Patra JK (2018) Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol Res 206:131–140
Goyal D, Prakash O, Pandey J (2019) Rhizospheric microbial diversity: an important component for abiotic stress management in crop plants toward sustainable agriculture. In: Singh JS, Singh DP (eds) New and future developments in microbial biotechnology and bioengineering. Elsevier, Amsterdam, pp 115–134
Grichko VP, Glick BR (2001a) Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol Biochem 39(1):11–17
Grichko VP, Glick BR (2001b) Flooding tolerance of transgenic tomato plants expressing the bacterial enzyme ACC deaminase controlled by the 35S, rolD or PRB-1b promoter. Plant Physiol Biochem 39(1):19–25
Grover M, Ali SZ, Sandhya V, Rasul A, Venkateswarlu B (2011) Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J Microbiol Biotechnol 27(5):1231–1240
Gulati A, Vyas P, Rahi P, Kasana RC (2009) Plant growth-promoting and rhizosphere-competent Acinetobacter rhizosphaerae strain BIHB 723 from the cold deserts of the Himalayas. Curr Microbiol 58:371–377
Gupta B, Huang B (2014) Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics 2014:701596. https://doi.org/10.1155/2014/701596
Gupta G, Parihar SS, Ahirwar NK, Snehi SK, Singh V (2015) Plant growth promoting Rhizobacteria (PGPR): current and future prospects for development of sustainable agriculture. J Microbiol Biochem 7:96–102
Gurusiddaiah S, Gealy DR, Kennedy AC, Ogg AG (1994) Isolation and characterization of metabolites from Pseudomonas fluorescens D7 for control of downy brome (Bromus tectorum). Weed Sci 42:492–501
Gutiérrez-Mañero FJ, Ramos-Solano B, Probanza AN, Mehouachi JR, Tadeo F, Talon M (2001) The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol Plant 111(2):206–211
Guy E, Lautier M, Chabannes M, Roux B, Lauber E, Arlat M, Noël LD (2013) xopAC-triggered immunity against Xanthomonas depends on Arabidopsis receptor-like cytoplasmic kinase genes PBL2 and RIPK. PLoS One 8:e73469. https://doi.org/10.1371/journal.pone.0073469
Haas D, Defago G (2005) Biological control of soil-borne pathogens by fluorescent Pseudomonads. Nat Rev Microbiol 3:307–319
Habib SH, Kausar H, Saud HM (2016) Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. Biomed Res Int 2016:6284547. https://doi.org/10.1155/2016/6284547
Habibzadeh F, Sorooshzadeh A, Pirdashti H, Modarres Sanavy SA (2012) A comparison between foliar application and seed inoculation of biofertilizers on canola (Brassica napus L.) grown under waterlogged conditions. Aust J Crop Sci 6(10):1435–1440
Hajiboland R, Bahrami-Rad S, Akhani H, Poschenrieder C (2018) Salt tolerance mechanisms in three Irano-Turanian Brassicaceae halophytes relatives of Arabidopsis thaliana. J Plant Res 131:1029–1046
Halgren A, Maselko M, Azevedo M, Mills D, Armstrong D, Banowetz G (2013) Genetics of germination-arrest factor (GAF) production by Pseudomonas fluorescens WH6: identification of a gene cluster essential for GAF biosynthesis. Microbiology 159:36–45
Hardoim PR, Overbeek LSV, Elsas JDV (2008) Properties of bacterialendophytes and their proposed role in plant growth. Trends Microbiol 16:463–471
Hasanuzzaman M, Nahar K, Gill SS, Fajita M (2013) Drought stress responses in plants, oxidative stress, and antioxidant defense. In: Tuteja N, Gill SS (eds) Climate change and plant abiotic stress tolerance. Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim, pp 209–250
Hatfield JL, Prueger JH (2015) Temperature extremes: effect on plant growth and development. Weather Clim Extremes 10:4–10
Hatfield JL, Boote KJ, Kimball BA, Ziska LH, Izaurralde RC, Ort D, Thomson AM, Wolfe D (2011) Climate impacts on agriculture: implications for crop production. Agron J 103(2):351–370
Hayat R, Ali S, Amara U, Khalid R, Ahmad I (2010) Soil beneficial bacteria and their role in plant growth promotion: a review. Ann Microbiol 60:579–598
Hellal FA, El-Shabrawi HM, El-Hady MA, Khatab IA, El-Sayed SA, Abdelly C (2018) Influence of PEG induced drought stress on molecular and biochemical constituents and seedling growth of Egyptian barley cultivars. J Genet Eng Biotechnol 16(1):203–212
Hendry GA (2005) Oxygen free radical process and seed longevity. Seed Sci J 3:141–147
Horn PJ, Chapman KD (2012) Lipidomics in tissues, cells and subcellular compartments. Plant J 70(1):69–80
Hu X, Li Y, Li C, Yang H, Wang W, Lu M (2010) Characterization of small heat shock proteins associated with maize tolerance to combined drought and heat stress. J Plant Growth Regul 29(4):455–464
Hui LJ, Kim SD (2013) Induction of drought stress resistance by multifunctional PGPR Bacillus licheniformis K11 in Pepper. Plant Pathol J 29:201–208
Hussain HA, Hussain S, Khaliq A, Ashraf U, Anjum SA, Men S, Wang L (2018) Chilling and drought stresses in crop plants: implications, cross talk, and potential management opportunities. Front Plant Sci 9:393. https://doi.org/10.3389/fpls.2018.00393
Ikeda M, Ohme-Takagi M (2009) A novel group of transcriptional repressors in Arabidopsis. Plant Cell Physiol 50:970–975
Ilyas N, Mumtaz K, Akhtar N, Yasmin H, Sayyed RZ, Khan W, Enshasy HAE, Dailin DJ, Elsayed EA, Ali Z (2020) Exopolysaccharides producing bacteria for the amelioration of drought stress in wheat. Sustainability 12(21):8876
Imaizumi S, Nishino T, Miyabe K, Fujimori T, Yamada M (1997) Biological control of annual bluegrass (Poa annua L.) with a Japanese isolate of Xanthomonas campestris pv. Poae (JT-P482). Biol Control 8:7–14
Issa A, Esmaeel Q, Sanchez L, Courteaux B, Guise JF, Gibon Y, Ballias P, Clément C, Jacquard C, Vaillant-Gaveau N, Aït Barka E (2018) Impacts of Paraburkholderia phytofirmans strain PsJN on tomato (Lycopersicon esculentum L.) under high temperature. Front Plant Sci 9:1397
Jaemsaeng R, Jantasuriyarat C, Thamchaipenet A (2018) Positive role of 1-aminocyclopropane-1-carboxylate deaminase-producing endophytic Streptomyces sp. GMKU 336 on flooding resistance of mung bean. Agric Nat Resour 52(4):330–334
Jaleel CA, Manivannan P, Wahid A (2009) Drought stress in plants: a review on morphologicalcharacteristics and pigments composition. Int J Agric Biol 11:100–105
Jamali H, Sharma A, Srivastava AK (2020) Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J Basic Microbiol 60(3):268–280
James RA, Blake C, Byrt CS, Munns S (2011) Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J Exp Bot 62:2939–2947
Jha Y (2018) Induction of anatomical, enzymatic, and molecular events in maize by PGPR under biotic stress. In: Meena VS (ed) Role of rhizospheric microbes in soil. Springer, Singapore, pp 125–141
Ji J, Yuan D, Jin C, Wang G, Li X, Guan C (2020) Enhancement of growth and salt tolerance of rice seedlings (Oryza sativa L.) by regulating ethylene production with a novel halotolerant PGPR strain Glutamicibacter sp. YD01 containing ACC deaminase activity. Acta Physiol Plant 42:42. https://doi.org/10.1007/s11738-020-3034-3
Jochum MD, McWilliams KL, Borrego EJ, Kolomiets MV, Niu G, Pierson EA, Jo YK (2019) Bioprospecting plant growth-promoting rhizobacteria that mitigate drought stress in grasses. Front Microbiol 10:2106
Johnson DR, Wyse DL, Jones KJ (1996) Controlling weeds with phytopathogenic bacteria. Weed Technol 10:621–624
Joshi B, Chaudhary A, Singh H, Kumar PA (2020) Prospective evaluation of individual and consortia plant growth promoting rhizobacteria for drought stress amelioration in rice (Oryza sativa L.). Plant Soil 457(1):225–240
Jung H, Ali S, Kim JY, Kim WC (2018) Transgenic Arabidopsis expressing acdS gene of Pseudomonas veronii-KJ alleviate the adverse effects of salt and water-logging stress. Plant Breed Biotechnol 6(3):221–232
Kakembo D, Lee YH (2019) Analysis of traits for biocontrol performance of Pseudomonas parafulva JBCS1880 against bacterial pustule in soybean plants. Biol Control 134:72–81
Kang SM, Khan AL, Waqas M, You YH, Kim JH, Kim JG, Hamayun M, Lee IJ (2014) Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J Plant Interact 9:673–682
Kang SM, Khan AL, Waqas M, You YH (2015) Gibberellin-producing Serratia nematodiphila PEJ1011 ameliorates low temperature stress in Capsicum annum L. Eur J Soil Biol 68:85–93
Kang SM, Khan AL, Waqas M, Asaf S, Lee KE, Park YG, Kim AY, Khan MA, You YH, Lee IJ (2019) Integrated phytohormone production by the plant growth-promoting rhizobacterium Bacillus tequilensis SSB07 induced thermotolerance in soybean. J Plant Interact 14(1):416–423
Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K (2004) A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought-and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 45(3):346–350
Katiyar V, Goel R (2004) Siderophore mediated plant growth promotion at low temperature by mutant of fluorescent pseudomonad. Plant Growth Regul 42:239–244
Kennedy AC, Johnson BN, Stubbs TL (2001) Host range of a deleterious rhizobacterium for biological control of downy brome. Weed Sci 49:792–797
Khan N, Bano A (2019) Rhizobacteria and abiotic stress management. In: Syed RZ, Arora NK, Reddy MS (eds) Plant growth promoting rhizobacteria for sustainable stress management. Springer, Singapore, pp 65–80
Khan N, Bano A, Rahman MA, Guo J, Kang Z, Babar MA (2019) Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci Rep 9(1):1–9
Khan MA, Asaf S, Khan AL, Jan R, Kang SM, Kim KM, Lee IJ (2020a) Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PLoS One 15(4):e0232228
Khan MA, Asaf S, Khan AL, Jan R, Kang SM, Kim KM, Lee IJ (2020b) Thermotolerance effect of plant growth-promoting Bacillus cereus SA1 on soybean during heat stress. BMC Microbiol 20(1):1–4
Khan N, Bano A, Ali S, Babar MA (2020c) Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul 90:189–203
Killi D, Raschi A, Bussotti F (2020) Lipid peroxidation and chlorophyll fluorescence of photosystem II performance during drought and heat stress is associated with the antioxidant capacities of C3 sunflower and C4 maize varieties. Int J Mol Sci 21(14):4846
Kim SY, Mulkey TJ (1997) Effect of ethylene antagonists on auxin-induced inhibition of intact primary root elongation in maize (Zea mays L.). J Plant Biol 40:256–260
Kim K, Jang YJ, Lee SM, Oh BT, Chae JC, Lee KJ (2014) Alleviation of salt stress by Enterobacter sp. EJ01 in tomato and Arabidopsis is accompanied by up-regulation of conserved salinity responsive factors in plants. Mol Cells 37:109–117
Kim AY, Shahzad R, Kang SM, Seo CW, Park YG, Park HJ, Lee IJ (2017) IAA-producing Klebsiella variicola AY13 reprograms soybean growth during flooding stress. J Crop Sci Biotechnol 20(4):235–242
Kloepper JW, Ryu CM, Zhang S (2004) Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 94(11):1259–1266
Koskey G, Mburu SW, Awino R, Njeru EM, Maingi JM (2021) Potential use of beneficial microorganisms for soil amelioration, phytopathogen biocontrol, and sustainable crop production in smallholder agroecosystems. Front Sustain Food Syst 5:606308. https://doi.org/10.3389/fsufs
Kosová K, Urban MO, Vítámvás P, Prášil IT (2016) Drought stress response in common wheat, durum wheat, and barley: transcriptomics, proteomics, metabolomics, physiology, and breeding for an enhanced drought tolerance. In: Hossain MA, Wani SH, Bhattacharjee S, Burritt DJ, Tran LSP (eds) Drought stress tolerance in plants, vol 2. Springer, Cham, pp 277–314
Kour D, Rana KL, Yadav AN, Sheikh I, Kumar V, Dhaliwal HS, Saxena AK (2020) Amelioration of drought stress in Foxtail millet (Setaria italica L.) by P-solubilizing drought-tolerant microbes with multifarious plant growth promoting attributes. Environ Sustain 3(1):23–34
Koyro HW, Ahmad P, Geissler N (2012) Abiotic stress responses in plants: an overview. In: Ahmad P, Prasad NV (eds) Environmental adaptations and stress tolerance of plants in the era of climate change. Springer, New York, pp 1–28
Kranner I, Minibayeva FV, Beckett RP, Seal CE (2010) What is stress? concepts, definitions and applications in seed science. New Phytol 188:655–673
Kremer RJ, Souissi T (2001) Cyanide production by rhizobacteria and potential for suppression of weed seedling growth. Curr Microbiol 43:182–186
Kuan KB, Othman R, Rahim KA, Shamsuddin ZH (2016) Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PLoS One 11(3):0152478. https://doi.org/10.1371/journal.pone.0152478
Kumar A, Patel JS, Meena VS, Srivastava R (2019a) Recent advances of PGPR based approaches for stress tolerance in plants for sustainable agriculture. Biocatal Agric Biotechnol 29:101271. https://doi.org/10.1016/j.bcab.2019.101271
Kumar S, Suyal DC, Yadav A, Shouche Y, Goel R (2019b) Microbial diversity and soil physiochemical characteristic of higher altitude. PLoS One 14(3):e0213844. https://doi.org/10.1371/journal.pone.0213844
Kumawat KC, Sharma P, Nagpal S, Gupta RK, Sirari A, Nair RM, Bindumadhava H, Singh S (2020) Dual microbial inoculation, a Game changer?–Bacterial biostimulants with multifunctional growth promoting traits to mitigate salinity stress in Spring Mungbean. Front Microbiol 11:600576. https://doi.org/10.3389/fmicb.2020.600576
Kusale SP, Attar YC, Sayyed RZ, Malek RA, Ilyas N, Suriani NL, Khan N, El Enshasy HA (2021) Production of plant beneficial and antioxidants metabolites by Klebsiella variicola under salinity stress. Molecules 26(7):1894
Kushwaha P, Kashyap PL, Kuppusamy P (2020) Microbes for cold stress resistance in plants: mechanism, opportunities, and challenges. In: Goel R, Soni R, Suyal DC (eds) Microbiological advancements for higher altitude agro-ecosystems and sustainability. Springer, Singapore, pp 269–292
Lanza MGDBR, dos Reis AR (2021) Roles of selenium in mineral plant nutrition: ROS scavenging responses against abiotic stresses. Plant Physiol Biochem 164:27–43. https://doi.org/10.1016/j.plaphy.2021.04.026
Lee DG, Ahsan N, Lee SH, Kang KY, Bahk JD, Lee IJ, Lee BH (2007) A proteomic approach in analyzing heat-responsive proteins in rice leaves. Proteomics 7(18):3369–3383
Lesk C, Rowhani P, Ramankutty N (2016) Influence of extreme weather disasters on global crop production. Nature 529:84–87
Li YQ, Sun ZL, Zhuang XF, Xu L, Chen SF, Li MZ (2003) Research progress on microbial herbicides. Crop Prot 22:247–252
Li J, McConkey BJ, Cheng Z, Guo S, Glick BR (2013) Identification of plant growth-promoting bacteria-responsive proteins in cucumber roots under hypoxic stress using a proteomic approach. J Proteomics 84:119–131
Li Z, Long R, Zhang T, Wang Z, Zhang F, Yang Q, Kang J, Sun Y (2017) Molecular cloning and functional analysis of the drought tolerance gene MsHSP70 from alfalfa (Medicago sativa L.). Int J Plant Res 130(2):387–396
Li Y, Shi H, Zhang H, Chen S (2019) Amelioration of drought effects in wheat and cucumber by the combined application of super absorbent polymer and potential biofertilizer. Peer J 7:e6073
Li X, Sun P, Zhang Y, Jin C, Guan C (2020) A novel PGPR strain Kocuria rhizophila Y1 enhances salt stress tolerance in maize by regulating phytohormone levels, nutrient acquisition, redox potential, ion homeostasis, photosynthetic capacity and stress-responsive genes expression. Environ Exp Bot 174:104023. https://doi.org/10.1016/j.envexpbot.2020.104023
Liu HC, Charng YY (2013) Common and distinct functions of Arabidopsis class A1 and A2 heat shock factors in diverse abiotic stress responses and development. Plant Physiol 163(1):276–290
Liu F, Xing S, Ma H, Du Z, Ma B (2013) Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl Microbiol Biotechnol 97(20):9155–9164
Liu CH, Siew W, Hung YT, Jiang YT, Huang CH (2021) 1-Aminocyclopropane-1-carboxylate (ACC) Deaminase gene in Pseudomonas azotoformans is associated with the amelioration of salinity stress in tomato. J Agric Food Chem 69(3):913–921
Lopez-Raez JA (2016) How drought and salinity affect arbuscular mycorrhizal symbiosis and strigolactone biosynthesis? Planta 243:1375–1385
Lozano GL, Holt J, Ravel J, Rasko DA, Thomas MG, Handelsman J (2016) Draft genome sequence of biocontrol agent Bacillus cereus UW85. Genome Announc 4(5):5–6
Maciag T, Krzyzanowska DM, Jafra S, Siwinska J, Czajkowski R (2020) The Great Five-an artificial bacterial consortium with antagonistic activity towards Pectobacterium spp. and Dickeya spp.: formulation, shelf life, and the ability to prevent soft rot of potato in storage. Appl Microbiol Biotechnol 104(10):4547–4561
Mahadik S, Kumudini BS (2020) Enhancement of salinity stress tolerance and plant growth in finger millet using fluorescent Pseudomonads. Rhizosphere 15:100226
Mantelin S, Touraine B (2004) Plant growth-promoting rhizobacteria and nitrateavailability: impacts on root development and nitrate uptake. J Exp Bot 55:27–34
Marin-Bruzos M, Grayston SJ, Forge T, Nelson LM (2021) Isolation and characterization of streptomycetes and pseudomonad strains with antagonistic activity against the plant parasitic nematode Pratylenchus penetrans and fungi associated with replant disease. Biol Control 158:104599
Marulanda A, Barea JM, Azcon R (2009) Stimulation of plant growth and drought tolerance by native microorganisms (AM Fungi and Bacteria) from dry environments: mechanisms related to bacterial effectiveness. J Plant Growth Regul 28:115–124
Marulanda A, Azcón R, Chaumont F, Ruiz-Lozano JM, Aroca R (2010) Regulation of plasma membrane aquaporins by inoculation with a Bacillus megaterium strain in maize (Zea mays L.) plants under unstressed and salt-stressed conditions. Planta 232:533–543
Maya MA, Matsubara YI (2013) Influence of arbuscular mycorrhiza on the growth and antioxidative activity in cyclamen under heat stress. Mycorrhiza 23(5):381–390
Mayak S, Tirosh T, Glick BR (2004) Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem 42(6):565–572
McKeon TA, Fernandez-Maculet JC, Yang SF (1995) Biosynthesis and metabolism of ethylene. In: Davies PJ (ed) Plant hormones physiology, biochemistry and molecular biology. Kluwer Academic, Dordrecht, pp 118–139
McKersie BD, Lesheim Y (1994) Stress and stress coping in cultivated plants, 1st edn. Springer, Dordrecht, p 256
McPhail KL, Armstrong DJ, Azevedo MD, Banowetz GM, Mills DI (2010) 4-Formylaminooxyvinylglycine, an herbicidal germination-arrest factor from Pseudomonas rhizosphere bacteria. J Nat Prod 73:1853–1857
Meena M, Swapnil P, Divyanshu K, Kumar S, Tripathi YN, Zehra A, Marwal A, Upadhyay RS (2020) PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: current perspectives. J Basic Microbiol 60(10):828–861
Meenakshi, Annapurna K, Govindasamy V, Ajit V, Choudhary DK (2019) Mitigation of drought stress in wheat crop by drought tolerant endophytic bacterial isolates. Vegetos 32(4):486–493
Migunova VD, Sasanelli N (2021) Bacteria as biocontrol tool against phytoparasitic nematodes. Plants 10(2):389
Mishra PK, Mishra S, Bisht SC, Selvakumar G, Kundu S, Bisht JK, Gupta HS (2009) Isolation, molecular characterization and -growth-promotion activities of a cold tolerant bacterium Pseudomonas sp. NARs9 (MTCC9002) from the Indian Himalaya. Biol Res 42(3):305–313
Mishra PK, Bisht SC, Ruwari P, Selvakumar G, Joshi GK, Bisht JK, Bhatt JC, Gupta HS (2011) Alleviation of cold stress in inoculated wheat (Triticum aestivum L.) seedlings with psychrotolerant Pseudomonads from NW Himalayas. Arch Microbiol 193(7):497–513
Mishra PK, Bisht SC, Bisht JK, Bhatt JC (2012) Cold-tolerant PGPRs as bioinoculants for stress management. In: Maheshwari DK (ed) Bacteria in agrobiology: stress management. Springer, Berlin, Heidelberg, pp 95–118
Misra S, Chauhan PS (2020) ACC deaminase-producing rhizosphere competent Bacillus spp. mitigate salt stress and promote Zea mays growth by modulating ethylene metabolism. 3 Biotech 10:119. https://doi.org/10.1007/s13205-020-2104-y
Misra S, Dixit VK, Khan MH, Mishra SK, Dviwedi G, Yadav S, Lehri A, Chauhan PS (2017) Exploitation of agro-climatic environment for selection of 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase producing salt tolerant indigenous plant growth promoting rhizobacteria. Microbiol Res 205:25–34
Misra S, Dixit VK, Mishra SK, Chauhan PS (2019) Demonstrating the potential of abiotic stress-tolerant Jeotgalicoccus huakuii NBRI 13E for plant growth promotion and salt stress amelioration. Ann Microbiol 69(4):419–434
Mitra S, Mukherjee A, Wiley-Kalil A, Das S, Owen H, Reddy PM, Ané J-M, James EK, Gyaneshwar P (2016) A rhamnose-deficient lipopolysaccharide mutant of Rhizobium sp. IRBG74 is defective in root colonization and beneficial interactions with its flooding-tolerant hosts Sesbania cannabina and wetland rice. J Exp Bot 67(19):5869–5884
Mittler R (2006) Abiotic stress, the field environment and stress combination. Trends Plant Sci 11:15–19
Molina-Favero C, Creus CM, Simontacchi M, Puntarulo S, Lamattina L (2008) Aerobic nitric oxide production by Azospirillum brasilense Sp245 and its influence on root architecture in tomato. Mol Plant Microbe Interact 2:1001–1009
Momčilović I, Pantelić D, Zdravković-Korać S, Oljača J, Rudić J, Fu J (2016) Heat-induced accumulation of protein synthesis elongation factor 1A implies an important role in heat tolerance in potato. Planta 244(3):671–679
Mosa KA, Ismail A, Helmy M (2017) Introduction to plant stresses. In: Mosa KA, Ismail A, Helmy M (eds) Plant stress tolerance. Springer, Cham, pp 1–19
Mukhtar T, Smith D, Sultan T, Seleiman MF, Alsadon AA, Ali S, Chaudhary HJ, Solieman TH, Ibrahim AA, Saad MA (2020) Mitigation of heat stress in Solanum lycopersicum L. by ACC-deaminase and exopolysaccharide producing Bacillus cereus: effects on biochemical profiling. Sustainability 12(6):2159. https://doi.org/10.3390/su12062159
Mumtaz MZ, Ahmad M, Jamil M, Hussain T (2017) Zinc solubilizing Bacillus spp. potential candidates for biofortification in maize. Microbiol Res 202:51–60
Mumtaz MZ, Saqib M, Abbas G, Akhtar J, Qamar ZU (2018) Genotypic variation in rice for grain yield and quality as affected by salt-affected field conditions. J Plant Nutr 41(2):233–242
Mumtaz MZ, Barry KM, Baker AL, Nichols DS, Ahmad M, Zahir ZA, Britz ML (2019) Production of lactic and acetic acids by Bacillus sp. ZM20 and Bacillus cereus following exposure to zinc oxide: a possible mechanism for Zn solubilization. Rhizosphere 12:100170. https://doi.org/10.1016/j.rhisph.2019.100170
Mumtaz MZ, Saqib M, Abbas G, Akhtar J, Qamar ZU (2020) Drought stress impairs grain yield and quality of rice genotypes by impaired photosynthetic attributes and K nutrition. Ric Sci 27(1):5–9
Munns R (2005) Genes and salt tolerance: bringing them together. New Phytol 167:645–663
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Nadeem SM, Zahir ZA, Naveed M, Arshad M (2009) Rhizobacteria containing ACC-deaminase confer salt tolerance in maize grown on salt-affected fields. Can J Microbiol 55:1302–1309
Nadeem SM, Ahmad M, Zahir ZA, Javaid A, Ashraf M (2014) The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol Adv 32:429–448
Nadeem SM, Ahmad M, Naveed M, Imran M, Zahir ZA, Crowley DE (2016) Relationship between in vitro characterization and comparative efficacy of plant growth-promoting rhizobacteria for improving cucumber salt tolerance. Arch Microbiol 198(4):379–387
Nadeem SM, Imran M, Naveed N, Khan MY, Ahmad M, Zahir ZA, Crowley DE (2017) Synergistic use of biochar, compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions. J Sci Food Agric 97:5139–5145
Nascimento F, Brígido C, Alho L, Glick BR, Oliveira S (2012) Enhanced chickpea growth-promotion ability of a Mesorhizobium strain expressing an exogenous ACC deaminase gene. Plant Soil 353:221–230
Nautiyal CS, Srivastava S, Chauhan PS, Seem K, Mishra A, Sopory S (2013) Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol Biochem 66:1–9
Nazli F, Najm-ul-Seher, Khan MY, Jamil M, Nadeem SM, Ahmad M (2020) Soil microbes and plant health. In: Haq IU, Ijaz S (eds) Plant disease management strategies for sustainable agriculture through traditional and modern approaches. Sustainability in plant and crop protection, vol 13. Springer, Cham, pp 111–135
Negi YK, Kumar J, Garg SK (2005) Cold-tolerant fluorescent Pseudomonas isolates from Garhwal Himalayas as potential plant growth promoting and biocontrol agents in pea. Curr Sci 89:2151–2156
Ngumbi E, Kloepper J (2016) Bacterial-mediated drought tolerance: current and future prospects. Appl Soil Ecol 105:109–125
Nishiuchi S, Yamauchi T, Takahashi H, Kotula L, Nakazono M (2012) Mechanisms for coping with submergence and waterlogging in rice. Rice 5:2. https://doi.org/10.1186/1939-8433-5-2
Niu SQ, Li HR, Paré PW, Aziz M, Wang SM, Shi H, Li J, Han QQ, Guo SQ, Li J, Guo Q (2016) Induced growth promotion and higher salt tolerance in the halophyte grass Puccinellia tenuiflora by beneficial rhizobacteria. Plant Soil 407:217–230
Njeru EM, Koskey G (2021) Using beneficial microorganisms to promote sustainable crop production and resilience of smallholder agroecosystems to changing climate. In: Mallappa VKH, Shirur M (eds) Climate change and resilient food systems. Springer, Singapore, pp 287–314
Nouri MZ, Moumeni A, Komatsu S (2015) Abiotic stresses: insight into gene regulation and protein expression in photosynthetic pathways of plants. Int J Mol Sci 16:20392–20416
Ohashi Y, Nakayama N, Saneoka H, Fujita K (2006) Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol Plant 50(1):138–141
Okrent RA, Halgren AB, Azevedo MD, Chang JH, Mills DI, Maselko M, Armstrong DJ, Banowetz GM, Trippe KM (2014) Negative regulation of germination-arrest factor production in Pseudomonas fluorescens WH6 by a putative extracytoplasmic function sigma factor. Microbiology 160:2432–2442
Orozco-Mosqueda MC, Glick BR, Santoyo G (2020) ACC deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiol Res 235:126439. https://doi.org/10.1016/j.micres.2020.126439
Ortiz-Ojeda P, Ogata-Gutiérrez K, Zúñiga-Dávila D (2017) Evaluation of plant growth promoting activity and heavy metal tolerance of psychrotrophic bacteria associated with maca (Lepidium meyenii Walp.) rhizosphere. AIMS Microbiol 3(2):279
Pahalvi HN, Rafiya L, Rashid S, Nisar B, Kamili AN (2021) Chemical fertilizers and their impact on soil health. In: Dar GH, Bhat RA, Mehmood MA, Hakeem KR (eds) Microbiota and biofertilizers, vol 2. Springer, Cham, pp 1–20
Pandey A, Palni LM, Mulkalwar P, Nadeem M (2002) Effect of temperature on solubilization of tricalcium phosphate by Pseudomonas corrugata. J Sci Ind Res 61:457–460
Pandey A, Trivedi P, Kumar B, Palni LM (2006) Characterization of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian central Himalaya. Curr Microbiol 53(2):102–107
Pandey P, Irulappan V, Bagavathiannan MV, Senthil-Kumar M (2017) Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front Plant Sci 8:537. https://doi.org/10.3389/fpls.2017.00537
Panwar M, Tewari R, Gulati A (2016) Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol Plant 38:278. https://doi.org/10.1007/s11738-016-2284-6
Passioura J (2007) The drought environment: physical, biological and agricultural perspectives. J Exp Bot 58:113–117
Patel PK, Singh AK, Tripathi N, Yadav D, Hemantaranjan A (2014) Flooding: abiotic constraint limiting vegetable productivity. Adv Plant Agric Res 1(3):96–103
Pathak R, Shrestha A, Lamichhane J, Gauchan D (2017) PGPR in biocontrol mechanisms and roles in disease suppression. Int J Agron Agric Res 11(1):69–80
Paul D, Lade H (2014) Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: a review. Agron Sustain Dev 34:737–752
Paul MV, Iyer S, Amerhauser C, Lehmann M, van Dongen JT, Geigenberger P (2016) Oxygen sensing via the ethylene response transcription factor RAP2 12 affects plant metabolism and performance under both normoxia and hypoxia. Plant Physiol 172(1):141–153
Péchy-Tarr M, Bruck DJ, Maurhofer M, Fischer E, Vogne C, Henkels MD, Donahue KM, Grunder J, Loper JE, Keel C (2008) Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ Microbiol 10(9):2368–2386
Perneel M, D’Hondt L, De Maeyer K, Adiobo A, Rabaey K, Höfte M (2008) Phenazines and biosurfactants interact in the biological control of soil-borne diseases caused by Pythium spp. Environ Microbiol 10(3):778–788
Phour M, Sindhu SS (2020) Amelioration of salinity stress and growth stimulation of mustard (Brassica juncea L.) by salt-tolerant Pseudomonas species. Appl Soil Ecol 149:103518
Phukan UJ, Mishra S, Shukla RK (2016) Waterlogging and submergence stress: affects and acclimation. Crit Rev In Biotech 36(5):956–966
Pitman MG, Läuchli A (2002) Global impact of salinity and agricultural ecosystems. In: Pitman MG, Lauchi A (eds) Salinity: environment-plants-molecules. Springer, Dordrecht, pp 3–20
Prasad PVV, Staggenborg SA, Ristic Z (2008) Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. Response of crops to limited water: understanding and modeling water stress effects on plant growth processes. In: Ahuja LR, Reddy VR, Saseendran SA, Yu Q (eds) Advances in agricultural systems modeling, vol 1, pp 301–355
Prasannakumar MK, Netravathi LM, Mahesh HB, Parivallal PB, Puneeth ME, Sathish A, Pramesh D, Middha SK, Das AJ, Rohit BS (2021) Comparative metagenomic analysis of rice soil samples revealed the diverse microbial population and biocontrol organisms against plant pathogenic fungus Magnaporthe oryzae. 3 Biotech 11(5):1–1
Prevost D, Drouin P, Antoun H (1999) The potential use of cold-adapted rhizobia to improve symbiotic nitrogen fixation in legumes cultivated in temperate regions. In: Margesin R, Schinner F (eds) Biotechnological applications of cold-adapted organisms. Springer, Berlin, Heidelberg, pp 161–176
Prevost D, Drouin P, Laberge S, Bertrand A, Cloutier J, Lévesque G (2003) Cold-adapted rhizobia for nitrogen fixation in temperate regions. Can J Bot 81(12):1153–1161
Qin S, Feng WW, Zhang YJ, Wang TT, Xiong YW, Xing K (2018) Diversity of bacterial microbiota of coastal halophyte Limonium sinense and amelioration of salinity stress damage by symbiotic plant growth-promoting actinobacterium Glutamicibacter halophytocola KLBMP 5180. Appl Environ Microbiol 84(19):e01533
Qurashi AW, Sabri AN (2012) Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz J Microbiol 43(3):1183–1191
Rahdari P, Hoseini SM (2012) Drought stress, a review. Int J Agron Plant Prod 3:443–446
Raheem A, Shaposhnikov A, Belimov AA, Dodd IC, Ali B (2018) Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch Agron Soil Sci 64(4):574–587
Rahnama A, James RA, Poustini K, Munns R (2010) Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Funct Plant Biol 37(3):255–263
Ramegowda V, Da Costa MV, Harihar S, Karaba NN, Sreeman SM (2020) Abiotic and biotic stress interactions in plants: a cross-tolerance perspective. In: Hossain MA, Liu F, Huang B (eds) Priming-mediated stress and cross-stress tolerance in crop plants. Academic Press, San Diego, CA 267-302
Ravanbakhsh M, Sasidharan R, Voesenek LA, Kowalchuk GA, Jousset A (2017) ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J Ecol 105(4):979–986
Řeháková K, Chroňáková A, Krištůfek V, Kuchtová B, Čapková K, Scharfen J, Čapek P, Doležal J (2015) Bacterial community of cushion plant Thylacospermum ceaspitosum on elevational gradient in the Himalayan cold desert. Front Microbiol 6:304
Rengasamy P (2002) Transient salinity and subsoil constraints to dryland farming in Australian sodic soils: an overview. Aust J Exp Agric 42:351–361
Rengasamy P (2010) Soil processes affecting crop production in salt-affected soils. Funct Plant Biol 37(7):613–620
Rijavec T, Lapanje A (2016) Hydrogen cyanide in the rhizosphere: not suppressing plant pathogens, but rather regulating availability of phosphate. Front Microbiol 7:1–14
Ritonga FN, Chen S (2020) Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 9(5):560
Rodriguez R, Redman R (2008) More than 400 million years of evolution and some plants still can’t make it on their own: plant stress tolerance via fungal symbiosis. J Exp Bot 59:1109–1114
Rojas-Tapias D, Moreno-Galván A, Pardo-Díaz S, Obando M, Rivera D, Bonilla R (2012) Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl Soil Ecol 61:264–272
Rozema J, Flowers T (2008) Ecology: crops for a salinized world. Science 322:1478–1480
Ruelland E, Zachowski A (2010) How plants sense temperature. Environ Exp Bot 69:225–232
Sachin N, Tsang A, Shaanker RU, Dayanandan S (2021) Genome Sequence Resource of Bacillus velezensis EB14, a native endophytic bacterial strain with biocontrol potential against the poplar stem canker causative pathogen, Sphaerulina musiva. Phytopathology 1:PHYTO09200433A
Sagar A, Sayyed RZ, Ramteke PW, Sharma S, Marraiki N, Elgorban AM, Syed A (2020) ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol Mol Biol Plants 26(9):1847–1854
Saharan BS, Nehra V (2011) Plant growth promoting rhizobacteria: a critical review. Life Sci Med Res 21(1):1–29
Saikia J, Sarma RK, Dhandia R, Yadav A, Bharali R, Gupta VK, Saikia R (2018) Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci Rep 8(1):1–16
Saleem M, Arshad M, Hussain S, Bhatti AS (2007) Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J Ind Microbiol Biotechnol 34(10):635–648
Salvi P, Manna M, Kaur H, Thakur T, Gandass N, Bhatt D, Muthamilarasan M (2021) Phytohormone signaling and crosstalk in regulating drought stress response in plants. Plant Cell Rep 22:1–25
Sang-Mo K, Radhakrishnan R, Khan AL, Min-Ji K, Jae-Man P, Bo-Ra K, Dong-Hyun S, In-Jung L (2014) Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol Biochem 84:115–124
Sangwan V, Foulds I, Singh J, Dhindsa RS (2001) Cold-activation of Brassica napus BN115 promoter is mediated by structural changes in membranes and cytoskeleton, and requires Ca2+ influx. Plant J 27(1):1–12
Sapre S, Gontia-Mishra I, Tiwari S (2018) Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol Res 206:25–32
Sapre S, Gontia-Mishra I, Tiwari S (2021) Plant growth-promoting rhizobacteria ameliorates salinity stress in pea (Pisum sativum). J Plant Growth Regul. https://doi.org/10.1007/s00344-021-10329-y
Sarkar J, Chakraborty B, Chakraborty U (2018) Plant growth promoting rhizobacteria protect wheat plants against temperature stress through antioxidant signalling and reducing chloroplast and membrane injury. J Plant Growth Regul 37(4):1396–1412
Sarkar J, Chakraborty U, Chakraborty B (2021) High-temperature resilience in Bacillus safensis primed wheat plants: A study of dynamic response associated with modulation of antioxidant machinery, differential expression of HSPs and osmolyte biosynthesis. Environ Exp Bot 182:104315
Sarwar M, Kremer RJ (1995) Enhanced suppression of plant growth through the production of L-tryptophan derived compounds by deleterious rhizobacteria. Plant Soil 172:261–269
Sasidharan R, Hartman S, Liu Z, Martopawiro S, Sajeev N, van Veen H, Yeung E, Voesenek LA (2018) Signal dynamics and interactions during flooding stress. Plant Physiol 176(2):1106–1117
Savvides A, Ali S, Tester M, Fotopoulos V (2016) Chemical priming of plants against multiple abiotic stresses: mission possible? Trends Plant Sci 21(4):329–340
Schmidt R, Köberl M, Mostafa A, Ramadan EM, Monschein M, Jensen KB, Bauer R, Berg G (2014) Effects of bacterial inoculants on the indigenous microbiome and secondary metabolites of chamomile plants. Front Microbiol 5:64. https://doi.org/10.3389/fmicb.2014.00064
Selvakumar G, Kundu S, Joshi P, Nazim S, Gupta AD, Mishra PK, Gupta HS (2008) Characterization of a cold tolerant plant growth-promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the North Western Indian Himalayas. World J Microbiol Biotechnol 24(7):955–960
Selvakumar G, Joshi P, Nazim S, Mishra P, Bisht J, Gupta H (2009) Phosphate solubilization and growth promotion by Pseudomonas fragi CS11RH1 (MTCC 8984), a psychrotolerant bacterium isolated from a high altitude Himalayan rhizosphere. Biologia 64(2):239–245
Selvakumar G, Panneerselvam P, Ganeshamurthy AN (2012) Bacterial mediated alleviation of abiotic stress in crops. In: Maheshwari DK (ed) Bacteria in agrobiology: stress management. Springer-Verlag, Berlin, Heidelberg, pp 205–224
Shakir MA, Asghari B, Arshad M (2012) Rhizosphere bacteria containing ACC deaminase conferred drought tolerance in wheat grown under semi-arid climate. Soil Environ 31:108–112
Sharma P, Khanna V, Kumar P (2013) Efficacy of aminocyclopropane-1-carboxylic acid (ACC)-deaminase producing rhizobacteria in ameliorating water stress in chickpea under axenic conditions. Afr J Microbiol Res 7:5749–5757
Sharma S, Joshi J, Kataria S, Verma SK, Chatterjee S, Jain M, Pathak K, Rastogi A, Brestic M (2020) Regulation of the Calvin cycle under abiotic stresses: an overview. In: Tripathi DK, Singh VP, Ramawat N (eds) Plant life under changing environment. Academic Press, San Diego, CA, pp 681–717
Shen YC, Korkor NL, Xiao R, Pu Q, Hu M, Zhang SS, Kong DD, Zeng G, Hu XF (2020) Antagonistic activity of combined bacteria strains against southern blight pathogen of Dendrobium officinale. Biol Control 151:104291
Showkat S (2012) Biological control of Fusarium oxysporum and Aspergillus sp. by Pseudomonas fluorescens isolated from wheat rhizosphere soil of Kashmir. IOSR Int J Pharm Biol Sci 1(4):24–32
Shrivastava P, Kumar R (2015) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22(2):123–131
Singh JS (2015) Biodiversity: current perspective. Clim Chang Environ Sustain 3(1):71–72
Singh RP, Jha PN (2015) Molecular identification and characterization of rhizospheric bacteria for plant growth promoting ability. Int J Curr Biotechnol 3:12–18
Singh RP, Jha PN (2017) Analysis of fatty acid composition of PGPR Klebsiella sp. SBP-8 and its role in ameliorating salt stress in wheat. Symbiosis 73(3):213–222
Singh JS, Pandey VC, Singh DP (2011) Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agric Ecosyst Environ 140:339–353
Singh RN, Gaba S, Yadav AN, Gaur P, Gulati S, Kaushik R, Saxena AK (2016) First high quality draft genome sequence of a plant growth promoting and cold active enzyme producing psychrotrophic Arthrobacter agilis strain L77. Stand Genomic Sci 11(1):1–9
Sofy MR, Aboseidah AA, Heneidak SA, Ahmad HR (2021) ACC deaminase containing endophytic bacteria ameliorate salt stress in Pisum sativum through reduced oxidative damage and induction of antioxidative defense systems. Environ Sci Pollut Res. https://doi.org/10.1007/s11356-021-13585-3
Srinivasan R, Mageswari A, Subramanian P, Maurya VK, Sugnathi C, Amballa C, Sa T, Gothandam KM (2017) Exogenous expression of ACC deaminase gene in psychrotolerant bacteria alleviates chilling stress and promotes plant growth in millets under chilling conditions. Indian J Exp Biol 55(7):463–468
Srivastava S, Yadav A, Seem K, Mishra S, Chaudhary V, Nautiyal CS (2008) Effect of high temperature on Pseudomonas putida NBRI0987 biofilm formation and expression of stress sigma factor RpoS. Curr Microbiol 56(5):453–457
Steffens B, Sauter M (2009) Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2 through an autoamplified signal pathway. Plant Cell 21:184–196
Stepien P, Kłbus G (2006) Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biol Plant 50(4):610. https://doi.org/10.1007/s10535-006-0096-z
Subramanian P, Mageswari A, Kim K, Lee Y, Sa T (2015) Psychrotolerant endophytic Pseudomonas sp. strains OB155 and OS261 induced chilling resistance in tomato plants (Solanum Lycopersicum Mill.) by activation of their antioxidant capacity. Mol Plant Microbe Interact 28:1073–1081
Subramanian P, Kim K, Krishnamoorthy R, Mageswari A, Selvakumar G, Sa T (2016) Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low temperatures. PLoS One 11(8):e0161592. https://doi.org/10.1371/journal.pone.0161592
Sun X, Griffith M, Pasternak JJ, Glick BR (1995) Low temperature growth, freezing survival, and production of antifreeze protein by the plant growth promoting rhizobacterium Pseudomonas putida GR12-2. Can J Microbiol 41(9):776–784
Sun Z, Yang LM, Han M, Han ZM, Yang L, Cheng L, Yang X, Lv ZL (2019) Biological control ginseng grey mold and plant colonization by antagonistic bacteria isolated from rhizospheric soil of Panax ginseng Meyer. Biol Control 138:104048
Suzuki N, Bajad S, Shuman J, Shulaev V, Mittler R (2008) The transcriptional co-activator MBF1c is a key regulator of thermotolerance in Arabidopsis thaliana. J Biol Chem 283(14):9269–9275
Suzuki N, Rivero RM, Shulaev V, Blumwald E, Mittler R (2014) Abiotic and biotic stress combinations. New Phytol 203:32–43
Sziderics AH, Rasche F, Trognitz F, Sessitsch A, Wilhelm E (2007) Bacterial endophytes contribute to abiotic stress adaptation in pepper plants (Capsicum annuum L.). Can J Microbiol 53:1195–1202
Taj Z, Challabathula D (2021) Protection of photosynthesis by halotolerant Staphylococcus Sciuri ET101 in tomato (Lycoperiscon esculentum) and rice (Oryza sativa) plants during salinity stress: Possible interplay between carboxylation and oxygenation in stress mitigation. Front Microbiol 11:547750. https://doi.org/10.3389/fmicb.2020.547750
Talaat NB, Shawky BT (2017) Microbe-mediated induced abiotic stress tolerance responses in plants. In: Singh DP, Singh HB, Prabha R (eds) Plant-microbe interactions in agro-ecological perspectives. Springer, Singapore, pp 101–133
Tanveer M, Ahmed HAI (2020) ROS signalling in modulating salinity stress tolerance in plants. In: Hasanuzzaman M, Tanveer M (eds) Salt and drought stress tolerance in plants, signaling networks and adaptive mechanisms. Springer, Cham, pp 299–314
Tapia-Vázquez I, Sánchez-Cruz R, Arroyo-Domínguez M, Lira-Ruan V, Sánchez-Reyes A, del Rayo S-CM, Padilla-Chacón D, Batista-García RA, Folch-Mallol JL (2020) Isolation and characterization of psychrophilic and psychrotolerant plant-growth promoting microorganisms from a high-altitude volcano crater in Mexico. Microbiol Res 232:126394
Thakare M, Sarma H, Datar S, Roy A, Pawar P, Gupta K, Pandit S, Prasad R (2021) Understanding the holistic approach to plant-microbe remediation technologies for removing heavy metals and radionuclides from soil. Curr Res Biotechnol 3:84–98. https://doi.org/10.1016/j.crbiot.2021.02.004
Thakur M, Sharma P, Anand A (2019) Seed priming-induced early vigor in crops: an alternate strategy for abiotic stress tolerance. In: Hasanuzzaman M, Fotopoulos V (eds) Priming and pretreatment of seeds and seedlings: implication in plant stress tolerance and enhancing productivity in crop plants. Springer, Singapore, pp 163–180
Theocharis A, Bordiec S, Fernandez O, Paquis S, Dhondt-Cordelier S, Baillieul F, Clément C, Barka EA (2012) Burkholderia phytofirmans PsJN primes Vitis vinifera L. and confers a better tolerance to low nonfreezing temperatures. Mol Plant Microbe Interact 25(2):241–249
Timmusk S, Nevo E (2011) Plant root associated biofilms. In: Maheshwari DK (ed) Bacteria in agrobiology: plant nutrient management, vol 3. Springer, Verlag, Berlin, pp 285–300
Tiryaki D, Ihsan A, Okkes A (2019) Psychrotolerant bacteria isolated from the leaf apoplast of coldadapted wild plants improve the cold resistance of bean (Phaseolus vulgaris L.) under low temperature. Cryobiology 86:111–119
Trivedi P, Sa T (2008) Pseudomonas corrugate (NRRL B-30409) mutants increased phosphate solubilisation, organic acid production, and plant growth at lower temperatures. Curr Microbiol 56:140–144
Turan M, Gulluce M, Cakmak R, Sahin F (2013) Effect of plant growth-promoting rhizobacteria strain on freezing injury and antioxidant enzyme activity of wheat and barley. J Plant Nutr 36:731–748
Upadhyay SK, Singh DP (2015) Effect of salt-tolerant plant growthpromoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol 17:288–293
Upadhyay SK, Singh JS, Singh DP (2011) Exopolysaccharide-producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 21:214–222
Vega-Celedón P, Bravo G, Velásquez A, Cid FP, Valenzuela M, Ramírez I, Vasconez IN, Álvarez I, Jorquera MA, Seeger M (2021) Microbial diversity of psychrotolerant bacteria isolated from wild flora of andes mountains and patagonia of chile towards the selection of plant growth-promoting bacterial consortia to alleviate cold stress in plants. Microorganisms 9(3):538
Verma P, Yadav AN, Khannam KS, Panjiar N, Kumar S, Saxena AK, Suman A (2015) Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann Microbiol 65(4):1885–1899
Vessey JK (2003) Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil 255(2):571–586
Vidoz ML, Loreti E, Mensuali A, Alpi A, Perata P (2010) Hormonal interplay during adventitious root formation in flooded tomato plants. Plant J 63:551–562
Vijayakumari K, Jisha KC, Puthur JT (2016) GABA/BABA priming: a means for enhancing abiotic stress tolerance potential of plants with less energy investments on defence cache. Acta Physiol Plant 38(9):230. https://doi.org/10.1007/s11738-016-2254-z
Vinocur B, Altman A (2005) Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr Opin Biotechnol 16:123–132
Voesenek LA, Benschop JJ, Bou J, Cox MC, Groeneveld HW, Millenaar FF, Vreeburg RA, Peeters AJ (2003) Interactions between plant hormones regulate submergence-induced shoot elongation in the flooding-tolerant dicot Rumex palustris. Ann Bot 91:205–211
Wahid A, Sehar S, Perveen M, Gelani S, Basra SM, Farooq M (2008) Seed pretreatment with hydrogen peroxide improves heat tolerance in maize at germination and seedling growth stages. Seed Sci Technol 36(3):633–645
Wang W, Vinocur B, Shoseyov O, Altman A (2004) Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 9(5):244–252
Wang Q, Dodd IC, Belimov AA, Jiang F (2016) Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase growth and photosynthesis of pea plants under salt stress by limiting Na+ accumulation. Funct Plant Biol 43(2):161–172
Wang X, Xiao C, Ji C, Liu Z, Song X, Liu Y, Li C, Yan D, Li H, Qin Y, Liu X (2021) Isolation and characterization of endophytic bacteria for controlling root rot disease of Chinese jujube. J Appl Microbiol 130(3):926–936
Wei Y, Zhu B, Liu W, Cheng X, Lin D, He C, Shi H (2021) Heat shock protein 90 co-chaperone modules fine-tune the antagonistic interaction between salicylic acid and auxin biosynthesis in cassava. Cell Rep 34(5):108717
Welti R, Li W, Li M, Sang Y, Biesiada H, Zhou HE, Rajashekar CB, Williams TD, Wang (2002) Profiling membrane lipids in plant stress responses role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J Biol Chem 277(35):31994–32002
Woo OG, Kim H, Kim JS, Keum HL, Lee KC, Sul WJ, Lee JH (2020) Bacillus subtilis strain GOT9 confers enhanced tolerance to drought and salt stresses in Arabidopsis thaliana and Brassica campestris. Plant Physiol Biochem 148:359–367
Wu YS, Yang CY (2019) Ethylene-mediated signaling confers thermotolerance and regulates transcript levels of heat shock factors in rice seedlings under heat stress. Bot Stud 60(1):1–12
Wu ZM, Yang Y, Li KT (2019) Antagonistic activity of a novel antifungalmycin N2 from Streptomyces sp. N2 and its biocontrol efficacy against Rhizoctonia solani. FEMS Microbiol Lett 366(3):fnz018
Wu YM, Chen X, Wang F, Hsiao CY, Yang CY, Lin ST, Wu LH, Chen YK, Liang YS, Lin YH (2021) Bacillus amyloliquefaciens strains control strawberry anthracnose through antagonistic activity and plant immune response intensification. Biol Control 157:104592
Xia Y, Farooq MA, Javed MT, Kamran MA, Mukhtar T, Ali J, Tabassum T, ur Rehman S, Munis MF, Sultan T, Chaudhary HJ (2020) Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol Biochem 151:640–649
Xie HT, Wan ZY, Li S, Zhang Y (2014) Spatiotemporal production of reactive oxygen species by NADPH oxidase is critical for tapetal programmed cell death and pollen development in Arabidopsis. Plant Cell 26:2007–2023
Yadav AN, Singh RN, Sachan SG, Kaushik R (2015) Prospecting cold deserts of north western Himalayas for microbial diversity and plant growth promoting attributes. J Biosci Bioeng 119(6):683–693
Yamauchi T, Rajhi I, Nakazono M (2011) Lysigenous aerenchyma formation in maize root is confined to cortical cells by regulation of genes related to generation and scavenging of reactive oxygen species. Plant Signal Behav 6:759–761
Yan J, Smith MD, Glick BR, Liang Y (2014) Effects of ACC deaminase containing rhizobacteria on plant growth and expression of Toc GTPases in tomato (Solanum lycopersicum) under salt stress. Botany 92:775–781
Yang J, Kloepper JW, Ryu CM (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci 14:1–4
Yang F, Zhang R, Wu X, Xu T, Ahmad S, Zhang X, Zhao J, Liu Y (2020) An endophytic strain of the genus Bacillus isolated from the seeds of maize (Zea mays L.) has antagonistic activity against maize pathogenic strains. Microb Pathog 142:104074
Yaseen R, Zafar-ul-Hye M, Hussain M (2019) Integrated application of ACC-deaminase containing plant growth promoting rhizobacteria and biogas slurry improves the growth and productivity of wheat under drought stress. Int J Agric Biol 21:869–878
Yaseen R, Aziz O, Saleem MH, Riaz M, Zafar-ul-Hye M, Rehman M, Ali S, Rizwan M, Nasser Alyemeni M, El-Serehy HA, Al-Misned FA (2020) Ameliorating the drought stress for wheat growth through application of ACC-deaminase containing rhizobacteria along with biogas slurry. Sustainability 12(15):6022
Zafar-ul-Hye M, Nasir A, Aon M, Hussain S, Ahmad M, Naz I (2018) Seed inoculation with Pseudomonas fluorescens and Pseudomonas syringae enhanced maize growth in a compacted saline-sodic soil. Phyton 87:25–31
Zafar-ul-Hye M, Danish S, Abbas M, Ahmad M, Munir TM (2019) ACC Deaminase producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with biochar improve wheat productivity under drought stress. Agronomy 9:343. https://doi.org/10.3390/agronomy9070343
Zahir ZA, Munir A, Asghar HN, Shahroona B, Arshad M (2008) Effectiveness of rhizobacteria containing ACC-deaminase for growth promotion of peas (P. sativum) under drought conditions. J Microbiol Biotechnol 18:958–963
Zakry FA, Shamsuddin ZH, Rahim KA, Zakaria ZZ, Rahim AA (2012) Inoculation of Bacillus sphaericus UPMB-10 to young oil palm and measurement of its uptake of fixed nitrogen using the N isotope dilution technique. Microbial Environ 27:257–262
Zang X, Geng X, Wang F, Liu Z, Zhang L, Zhao Y, Tian X, Ni Z, Yao Y, Xin M, Hu Z (2017) Overexpression of wheat ferritin gene TaFER-5B enhances tolerance to heat stress and other abiotic stresses associated with the ROS scavenging. BMC Plant Biol 17(1):14. https://doi.org/10.1186/s12870-016-0958-2
Zhang CX, Fu GF, Yang XQ, Yang YJ, Zhao X, Chen TT, Zhang XF, Jin QY, Tao LX (2016) Heat stress effects are stronger on spikelets than on flag leaves in rice due to differences in dissipation capacity. J Agron Crop Sci 202(5):394–408
Zhang X, Rong X, Cai M, Meng Q (2019) Collaborative optimization of emissions and abatement costs for air pollutants and greenhouse gases from the perspective of energy structure: an empirical analysis in Tianjin. Sustainability 11:3872. https://doi.org/10.3390/su11143872
Zhou C, Ma Z, Zhu L, Xiao X, Xie Y, Zhu J, Wang J (2016) Rhizobacterial strain Bacillus megaterium BOFC15 induces cellular polyamine changes that improve plant growth and drought resistance. Int J Mol Sci 17:976. https://doi.org/10.3390/ijms17060976
Zhu J, Dong CH, Zhu JK (2007) Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr Opin Plant Biol 10(3):290–295
Zhu XC, Song FB, Liu SQ, Liu TD, Zhou X (2012) Arbuscular mycorrhizae improve photosynthesis and water status of Zea mays L. under drought stress. Plant Soil Environ 58(4):186–191
Zicca S, De Bellis P, Masiello M, Saponari M, Saldarelli P, Boscia D, Sisto A (2020) Antagonistic activity of olive endophytic bacteria and of Bacillus spp. strains against Xylella fastidiosa. Microbiol Res 236:126467
Zubair M, Hanif A, Farzand A, Sheikh TM, Khan AR, Suleman M, Ayaz M, Gao X (2019) Genetic screening and expression analysis of psychrophilic Bacillus spp. reveal their potential to alleviate cold stress and modulate phytohormones in wheat. Microorganisms 7(9):337
Zvinavashe AT, Mardad I, Mhada M, Kouisni L, Marelli B (2021) Engineering the plant microenvironment to facilitate plant-growth-promoting microbe association. J Agric Food Chem. https://doi.org/10.1021/acs.jafc.1c00138
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Mumtaz, M.Z. et al. (2022). Role of Plant Growth-Promoting Rhizobacteria in Combating Abiotic and Biotic Stresses in Plants. In: Arora, N.K., Bouizgarne, B. (eds) Microbial BioTechnology for Sustainable Agriculture Volume 1. Microorganisms for Sustainability, vol 33. Springer, Singapore. https://doi.org/10.1007/978-981-16-4843-4_2
Download citation
DOI: https://doi.org/10.1007/978-981-16-4843-4_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-4842-7
Online ISBN: 978-981-16-4843-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)