
Abstract
During the last decade, there has been growing interest in the role of trehalose metabolism in tolerance to abiotic stress in higher plants, especially cold stress. So far, this metabolism has not yet been studied in Vitis vinifera L., despite the economic importance of this crop. The goal of this paper was to investigate the involvement of trehalose metabolism in the response of grapevine to chilling stress, and to compare the response in plants bacterised with Burkholderia phytofirmans strain PsJN, a plant growth-promoting rhizobacterium that confers grapevine chilling tolerance, with mock-inoculated plants. In silico analysis revealed that the V. vinifera L. genome contains genes encoding the enzymes responsible for trehalose synthesis and degradation. Transcript analysis showed that these genes were differentially expressed in various plant organs, and we also characterised their response to chilling. Both trehalose and trehalose 6-phosphate (T6P) were present in grapevine tissues and showed a distinct pattern of accumulation upon chilling. Our results suggest a role for T6P as the main active molecule in the metabolism upon chilling, with a possible link with sucrose metabolism. Furthermore, plants colonised by B. phytofirmans and cultivated at 26°C accumulated T6P and trehalose in stems and leaves at concentrations similar to non-bacterised plants exposed to chilling temperatures for 1 day. Overall, our data suggest that T6P and trehalose accumulate upon chilling stress in grapevine and might participate in the resistance to chilling stress conferred by B. phytofirmans.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Trehalose is a multi-faceted disaccharide present in most living organisms (Elbein et al. 2003; Paul et al. 2008). In higher plants, trehalose is (1) synthesised via the phosphorylated intermediate trehalose-6-phosphate (T6P) in a two-step pathway involving trehalose phosphate synthase (TPS) and trehalose phosphate phosphatase (TPP) and (2) degraded by trehalase (TRE; Paul et al. 2008). Trehalose is a key sugar in microorganisms (Argüelles 2000; Paul et al. 2008) and is particularly involved in stress resistance. It protects several bacteria and fungi against osmotic (Purvis et al. 2005), oxidative (Cao et al. 2008) and cold stresses (Kandror et al. 2002, 2004). In contrast to these organisms, trehalose was, until recently, considered to be an unimportant sugar in most vascular plants. With the exception of some drought-resistant “resurrection plants”, such as Myrothamnus flabellifolius, Selaginella lepidophylla or Selaginella tamariscina (Drennan et al. 1993; Goddijn and van Dun 1999; Liu et al. 2008), its concentration in other species was barely detectable (Paul et al. 2008; Fernandez et al. 2010).
There are large families of TPS and TPP genes in Arabidopsis thaliana (Leyman et al. 2001; Avonce et al. 2006; Lunn 2007). To date, in all plants whose TPS genes were analysed, only one isoform has been shown to encode an active TPS enzyme (AtTPS1, OsTPS1, SlTPS1; van Dijck et al. 2002; Vandesteene et al. 2010; Li et al. 2011). In contrast, recent evidence suggests that all of the A. thaliana TPP genes encode active enzymes with specific expression patterns in different plant tissues (Vandesteene 2009). During the last decade, transgenic approaches have been used to increase trehalose synthesis in crop species by over-expression of bacterial, yeast or plant enzymes for trehalose synthesis (for review, see Fernandez et al. 2010). Most of the resulting transgenic plants displayed improved resistance against abiotic stress, especially drought and cold. In these plants, concentrations of both T6P and trehalose were usually higher than in wild-type controls (for review, see Fernandez et al. 2010). However, so far, no clear explanation of the involvement of T6P and/or trehalose in triggering these resistance traits has been provided. Nevertheless, it seems clear that at least one of these two molecules displays signalling properties (Reignault et al. 2001; Bae et al. 2005a, b; Lunn et al. 2006; Renard-Merlier et al. 2007; Paul et al. 2008; Fernandez et al. 2010). Recently, strong evidence has emerged to suggest that T6P inhibits the sucrose-non-fermenting-1-related kinase 1 (SnRK1), a key transcriptional regulator that responds to carbon and energy supply (Zhang et al. 2009). In addition, T6P may act as a downstream signal of sucrose cytoplasmic concentration (Lunn et al. 2006). Surprisingly, not much is known about either T6P or trehalose levels in non-transgenic plants or the regulation of their synthesis following application of stresses, especially in woody plants under cold stress.
Despite global warming, low temperatures are still a major limitation to agricultural productivity, causing massive losses to crops worldwide (Boyer 1982; Bray et al. 2000; Ruelland et al. 2009). The types of damage caused by cold have been extensively reviewed in Ruelland et al. (2009). It is common to distinguish stress imposed by freezing (below 0°C) and chilling (above 0°C; Ruelland et al. 2009). The latter generally triggers a phenomenon known as cold acclimation (Ruelland et al. 2009; Janská et al. 2010), even if long-term exposure to chilling can threaten the plant’s survival. Cold acclimation is characterised by modification of several physiological traits (reviewed in Ruelland et al. 2009), including accumulation of soluble sugars as cryoprotectants (Steponkus 1984; Hirsh 1987; Crowe 2007), as well as scavengers of reactive oxygen species (Couée et al. 2006) and signalling molecules (Ruelland et al. 2009).
Vitis vinifera L. is cultivated worldwide, mainly in temperate and cool climates. Chilling markedly reduces its growth and photosynthesis, and delays flowering (Hendrickson et al. 2004; Bertamini et al. 2005; Ait Barka et al. 2006; Bertamini et al. 2006). Cold acclimation in grapevine is characterised by the synthesis of proline and phenolic compounds, but also by simultaneous accumulation of starch and soluble sugars (Ait Barka et al. 2006). Improving resistance to chilling stress would be of considerable interest to winegrowers. The use of plant growth-promoting rhizobacteria (PGPR) is one of the most recent technologies developed to protect crops against damage caused by various forms of stress (Yang et al. 2009). We have previously shown that the PGPR Burkholderia phytofirmans strain PsJN is able to colonise grapevine roots (Ait Barka et al. 2000), and that this association increases soluble sugar concentrations and reduces the damage caused to grapevine by chilling (Ait Barka et al. 2006).
In this paper, our goal was to identify the active isoforms of TPS, TPP and TRE and then investigate transcript level regulation of their expression upon chilling (4°C) in V. vinifera L., as well as the fluctuations of both T6P and trehalose. In addition, we measured levels of these metabolites in the chilling-tolerant grapevine plants when bacterised with B. phytofirmans strain PsJN, to assess whether T6P and/or trehalose have any role in this induced resistance.
Materials and methods
Plant material
In vitro growth conditions
In vitro growth conditions were chosen to prevent contamination by microorganisms, which can contain high concentrations of trehalose and so interfere with the measurement of the relatively low amounts of trehalose found in plants. In vitro plants of V. vinifera L. cv. Chardonnay clone 7535 (obtained from Moet & Chandon, Épernay, France) were micro-propagated from nodal explants grown on 15 ml agar medium in 25-mm culture tubes as described previously (Ait Barka et al. 2006). Cultured plants were grown in a growth chamber at constant 26°C, under white fluorescent light (200 μmol quanta m−2 s−1) with 16 h light per day.
Bacterial inoculum and plant bacterisation
The bacterial inoculum was produced by transferring two loops of Burkholderia phytofirmans strain PsJN (obtained from Dr. E. Ait Barka, University of Reims, France) to 100 ml of King’s B liquid medium in a 250-ml Erlenmeyer flask incubated at 20°C at 150 rpm for 48 h. Bacteria were collected by centrifugation (3,000g for 15 min) and washed twice with phosphate-buffered saline (PBS, 10 mM Na2HPO4, 150 mM NaCl, pH 6.5). The pellet was re-suspended in PBS and the bacterial concentration was estimated by measuring optical density at 600 nm. The concentration was adjusted with PBS to 3 × 108 CFU ml−1 (Pillay and Nowak 1997) and the resulting solution was used as the inoculum. Roots of 2-week-old in vitro plants were immersed in inoculum for 20 s, whereas control mock-inoculated plants were immersed in PBS solution for the same length of time. After inoculation, plants were grown as described above for 4 weeks before the experiments.
Chilling treatment
A 6-week-old plants, bacterised or non-bacterised with B. phytofirmans, were transferred to a growth chamber at 4°C, with 16 h light (white fluorescent light, 200 μmol quanta m−2 s−1) and 8 h dark, whereas the control plants grew at 26°C under the same light conditions.
Before performing experiments described in this manuscript, all samples were previously controlled by real-time RT-PCR for positive induction of VvCBF4 (6, 9 and 24 h after cold exposure), a major cold response regulator in grapevine (Xiao et al. 2008; data not shown).
Sampling
A minimum of six plants was harvested for each treatment, which was replicated at least three times. For sugar analysis, sampling was performed at the same hour (8:00 a.m. after 2 h of illumination, except for the 3 and 9 h samples). Roots, stems and leaves were collected separately from the harvested plants and immediately frozen in liquid nitrogen. Plant material was manually ground to a fine powder at liquid nitrogen temperature using a pestle and mortar. Aliquots of 100 mg were taken for RNA extraction, and 20 mg for sugar extraction.
Analysis of gene expression
Sampling, DNase treatment, RNA extraction and synthesis of cDNA
Total RNA was extracted from 100 mg of leaf powder using RNA Plant Purification Reagent (Invitrogen), according to the manufacturer’s instructions. The RNA pellet was re-suspended in 20 μl of RNase-free water. Genomic DNA was removed by RNase-free DNase treatment (Promega) using 1 U of enzyme per 5 μl of RNA according to manufacturer’s instructions. RNA purity and concentration were assessed by determining the absorbance of the samples at 260 and 280 nm and the A 260/A 280 ratio. RNA quality was evaluated from the integrity of the 28S and 18S rRNA bands on a 1% agarose gel after electrophoresis in 0.5× TAE (Tris–acetate–EDTA), staining with ethidium bromide and visualisation under UV light. Reverse transcription of RNA was performed with 150 ng of total RNA, using M-MLV reverse transcriptase (Invitrogen) following the manufacturer’s protocol.
Real-time RT-PCR analysis
PCR reactions were carried out in duplicates in 96-well plates (15 μl per well) in a reaction buffer containing 1× SYBR Green I mix (PE Biosystems, Foster City, CA, USA; including Taq polymerase, dNTPs, SYBR Green dye), 280 nM primers (forward and reverse) and a 1:30 dilution of reverse transcribed RNA. PCR conditions were 95°C for 15 s (denaturation) and 60°C for 1 min (annealing/extension) for 40 cycles using a GeneAmp 5700 sequence Detection System (Applied Biosystems). Transcript level was calculated using the comparative Ct method (ΔΔCt) with the Ef1α gene as internal control for normalisation (Terrier et al. 2005) and non-bacterised plants grown at 26°C as the reference sample (1× expression level). For TPPs expression analysis, transcript number was calculated using the standard curve method and normalised with the Ef1α gene as internal control. Primers used for analysis of VvTPS1, VvTPPs and VvTRE expression are given in Table 1.
Sequence alignment and phylogenetic tree
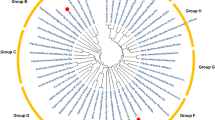
All protein sequences used for alignment were extracted from the appropriate database (http://www.ncbi.nlm.nih.gov/genbank/). Sequence alignments and phylogenetic analysis were performed with ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/) with the following parameters (tree format: phylip; excludes gaps: false; percent identity matrix: false; distance correction: false; clustering methods: neighbour-joining). The phylogenetic tree was drawn in a radial form using TreeView software (http://www.taxonomy.zoology.gla.ac.uk/rod/treeview.html).
Yeast growth complementation assay
We used the yeast (Saccharomyces cerevisiae) W303-1A wild-type strain (Matα leu2-3, 112 ura3-1 trp1-1 his3-11,15 ade2-1 can1-100 GAL SUC2), and the tps2Δ deletion strain YSH448 (W303-1A, tps2Δ::HIS3) for the complementation assays (Hohmann et al. 1993). All yeasts were from the Institute of Botany, KU Leuven, Belgium, Prof. F. Rolland. The VvTPPA sequence was amplified from cDNA synthesised by reverse transcription of RNA extracted from leaves of V. vinifera L. cv. Chardonnay clone 7535 (primers, VvTPPA-A: catgcc-ATGGATCTGAAGTCCAATCATTC; VvTPPA-B: tcccccggg-TAGTGCACTTG ACTTCTTCC) and cloned in a yeast multicopy pYX212 plasmid with an HXT7 promoter and URA3 marker. The VvTPPA coding region was inserted without a stop codon and in frame with a C-terminal double HA tag. As a control, the tps2Δ strain was transformed with empty pYX212 vector, or with the PYX212 vector containing the ScTPS2 gene (Ramon et al. 2009) or the Arabidopsis thaliana AtTPPA cDNA (AtTPPA-A: ggaagatct-ATGGACATGAAATCTGGTCACTC; AtTPPA-B: aaggcct-ACCCATTGATCTCTTCCAT GTCA). Yeast transformation was performed using the one-step method as described by Chen et al. (1992). For drop assays, cultures of the transformed tps2Δ and WT control strains were grown overnight at 28°C on SD-ura containing 2% glucose and drop assays were performed (spotted at an OD600 of 1). Transformants were incubated at 28 (control) or 38.6°C and analysed after 2 days. For Western blot analysis of the heterologous protein expression, cells of 50 ml overnight cultures were harvested and resuspended in 500-μl ice-cold lysis buffer (1× PBS, 0.1% Triton X-100, 10% glycerol, 2.5 mM MgCl2, 1 mM EDTA, pH8) containing protease inhibitors (Complete EDTA-free, Roche; see Ramon et al. 2009 and Vandesteene et al. 2010 for details). Amounts of immuno-precipitated reactive protein did not vary by more than a factor 2 between samples. Equal amounts of solubilised proteins were loaded on a Mini-protean precast gel (Bio-Rad) together with the SeeBlue pre-stained protein standard (Invitrogen) in a Tris/glycine/SDS running buffer and analysed using immunoblotting. For detection of HA-tagged proteins, we used HRP-coupled anti-HA high affinity rat antibodies (1/1,000; Roche). Proteins indicated as full-length have about the predicted size of full-length tagged proteins.
Analyses of trehalose and T6P
Sugar extraction
Sugars were extracted according to Lunn et al. (2006). A 20 mg of frozen powder were mixed in 2-ml tubes (pre-cooled with liquid N2) with 350 μl of ice-cold CHCl3/CH3OH (3:7, v/v), vigorously shaken and incubated at −20°C for 2 h. Afterwards, 350 μl of ice-cold ultrapure water (resistivity 18.2 MΩ) were added. Samples were vortexed and centrifuged at 14,000g for 10 min at 4°C. The upper aqueous phase was transferred to a 2-ml screw-capped tube and the chloroform phase was re-extracted with 300 μl ice-cold water. The aqueous fractions were combined and evaporated to dryness using a centrifugal vacuum dryer and then redissolved in 350 μl ultrapure water (resistivity 18.2 MΩ). Before further analysis, large molecular weight compounds were removed by centrifugal filtration using a Multiscreen Ultracel-10 filter plate (10 kDa cutoff; Millipore).
Trehalose measurement
Trehalose was measured enzymatically by fluorescence spectrometry in a 96-well microtitre plate as described by Mollo et al. (2011).
T6P measurement
T6P was assayed in plant extracts using a Dionex HPLC system coupled to an AB SCIEX Q-Trap5500 apparatus (AB SCIEX) according to Lunn et al. (2006). Before analysis, samples were diluted 40 times with ultrapure water and spiked with a deuterated-T6P standard Lunn et al. (2006).
Statistical analyses
All the results were replicated in three independent experiments. Figures display the means of three biological replicates ±SD. Student’s t test and Tukey’s test were performed using Kyplot 2.0 software (http://www.kyenslab.com/en/products/kyplot.html). Differences at P < 0.05 were considered significant.
Results
Analysis of trehalose-related genes in grapevine
Grapevine TPS, TPP and TRE genes were identified by sequence similarity-based searches of the V. vinifera L. genome database: (http://www.ncbi.nlm.nih.gov/sites/entrez?db=bioproject&cmd=Retrieve&dopt=Overview&list_uids=12992). The deduced protein sequences were compared with those of the various isoforms from other plant species, to identify which of the grapevine genes are likely to encode catalytically active enzymes of trehalose metabolism.
TPS genes
Grapevine TPS genes were identified by searching the database using the TBLASTN algorithm with AtTPS1 as a query sequence. AtTPS1 is the only TPS isoform in A. thaliana with demonstrated TPS activity (Blázquez et al. 1998; Vandesteene et al. 2010). Seven TPS genes were found in the V. vinifera L. genome (Table 2). All of the deduced protein sequences contain the typical glycosyltransferase domain found in TPS enzymes (cd03788; Fig. 1). Like their A. thaliana counterparts, the C-terminal region of these V. vinifera TPS proteins contained a TPP-like domain (pfam02358; Fig. 1).

Alignment of V. vinifera L. TPS and A. thaliana AtTPS1 protein sequences. The TPS domain is underlined in blue and TPP domain in green. Amino acid residues that are similar in all sequences are shaded in grey, while letters representing those that are identical in VvTPS1 and AtTPS1 are in red. The black arrows mark amino acids that are predicted to be important for substrate binding (according to Lunn 2007; Vandesteene et al. 2010) in catalytically active TPS enzymes, and all of these residues are perfectly conserved in both AtTPS1 and VvTPS1
Phylogenetic analysis showed that only one V. vinifera L. TPS (VvTPS1) clustered with class I TPS protein from A. thaliana and particularly AtTPS1, the active TPS isoform in A. thaliana (Fig. S1; Vandesteene et al. 2010). Furthermore, VvTPS1 showed much greater similarity to AtTPS1 (86%, Table 2; Fig. 1) than did the VvTPS2-7 proteins (54–56%). VvTPS1 also contained an N-terminal extension that is characteristic of the catalytically active TPSs, such as AtTPS1 (Fig. 1), OsTPS1 and SlTPS1 (Fig. S2; van Dijck et al. 2002; Li et al. 2011) and absent in VvTPS2-7 (Fig. 1). Finally, all of the residues predicted to be involved in substrate (G6P and UDPG) binding and catalysis in AtTPS1, OsTPS1 and SlTPS1, based on the crystal structure of the Escherichia coli TPS (otsA; Lunn 2007; Vandesteene et al. 2010), are conserved in VvTPS1 (Fig. 1, S2). Based on these observations, it was concluded that VvTPS1 is the only gene encoding an active TPS enzyme in grapevine. Phylogenetic analysis also revealed that the six other TPS (VvTPS2-7) clustered with the so-called class II TPS protein from plants (Fig. S1), whose function is still unknown (Lunn 2007; Vandesteene et al. 2010).
TPP genes
Using a similar approach, six TPP genes were identified in the V. vinifera L. genome (Table 2). The encoded proteins exhibit at least 60% of similarity with their respective closest A. thaliana homologues (Table 2). Each of them contained a putative TPP domain (pfam02358) and the three typical amino acid motifs of the l-2-haloacid dehalogenase (HAD) superfamily of enzymes (Lunn 2007). Recent evidence suggests that all 10 of the TPP genes in A. thaliana encode active enzymes (Vandesteene 2009). Alignment with Thermoplasma acidophilum TPP (TaTPP), whose crystal structure has been resolved (Rao et al. 2006), revealed that residues involved in binding the trehalose moiety of T6P were conserved in all grapevine TPP isoforms, apart for the conservative substitution of Lys to Arg in four isoforms (VvTPPA, VvTPPB, VvTPPC and VvTPPE; Fig. S3), which is commonly observed in TPP enzymes from plants, including several isoforms from A. thaliana that are known to be catalytically active (Lunn 2007). This suggests that all six of the grapevine TPP genes encode active enzymes. Therefore, we decided to analyse relative transcript level of all isoforms in grapevine organs by RT-qPCR (Fig. S4). Three isoforms were poorly expressed both in non-chilled and chilled plants: VvTPPF could not be detected in any grapevine organs (Fig. S4) VvTPPC and VvTPPD were only detected in leaves and stems, respectively, but at a very low level (Fig. S4). Concerning the three other isoforms, VvTPPB, VvTPPE and VvTPPA mRNAs were the most abundant, respectively, in roots, stems and leaves, both in non-chilled and chilled plants (Fig. S4), but VvTPPA expression was the most induced in chilled plants in the three organs (Fig. 2). VvTPPE induction by chilling was similar when compared with VvTPPA but VvTPPB expression was poorly modified (Fig. 2). Furthermore, VvTPPA was the principal TPP isoform in leaves (Fig. S4), which account for the majority of the tissue in the in vitro grapevine plants (>60%; O. Fernandez, unpublished results). Consequently, we decide to focus on VvTPPA for detailed study as this was the most highly expressed TPP gene in grapevine and showed a strong response to chilling, making it a good marker for the effects of chilling on TPP activity.

Relative transcript accumulation of VvTPPs in chilled plant organs. Relative transcript accumulation was calculated using the comparative Ct method (ΔΔCt) with the Ef1α gene as internal control for normalisation. The results represent the mean fold increase (±SD) of mRNA level of three independent experiments over non-bacterised plants grown at 26°C as the reference sample (referred as 1× expression level). Plants were exposed to chilling during 9 h, a time point when all VvTPPs gene induction peaked. n.a. not applicable
TRE gene
Only one TRE gene sharing significant similarity with known plant TRE genes was identified in the V. vinifera L. genome, indicating that there is a single isoform of trehalase in grapevine (Table 2). Primers were designed for analysis of VvTRE expression by qRT-PCR.
VvTPPA encodes a functional trehalose phosphate phosphatase
The VvTPPA gene was the most widely expressed TPP gene in leaves and the most induced upon chilling in grapevine (Fig. 2), therefore, it was of interest to test if this gene encodes a catalytically active TPP enzyme. Using the 1,618-bp full-length cDNA sequence of VvTPPA from V. vinifera L. cv. Pinot Noir (XM_002265643.1) available in database, flanking primers were designed to amplify the 1,158-bp ORF of the VvTPPA gene from V. vinifera L. cv. Chardonnay by PCR. The resulting amplicon was purified and sequenced. Alignment revealed that the cv. Chardonnay sequence (Fig. S5) shared 99.4% identity with the cv. Pinot Noir sequence (XM_002265643.1), with only five differences in the nucleic acid sequence (Fig. S5). Similar differences from the XM_002265643.1 sequence were found in several VvTPPA ESTs from cv. Cabernet Sauvignon, only one of which leads to a change in the amino acid sequence; namely, the conservative substitution of Leu 310 (CTT) for Val 310 (GTT) within the TPP domain (Fig. S5).
A yeast complementation assay was used to test whether the VvTPPA coding sequence from cv. Chardonnay encodes a functional TPP enzyme. The VvTPPA PCR product was cloned into a yeast expression vector and introduced into a tps2∆ yeast mutant, a TPP knock-out yeast mutant unable to grow at high temperature (Fig. 3). All transformants were able to growth at non-stringent temperature of 28°C (Fig. 3). In contrast to tps2∆ transformed with the empty vector, the tps2∆ transformed with VvTPPA was able to grow at a high temperature of 38.6°C, like the positive controls, WT and AtTPPA-transformed yeasts (Fig. 3). This yeast growth complementation assay demonstrates that VvTPPA displays heterologous TPP activity. Therefore, any changes in VvTPPA transcript abundance that are translated into changes in the amount of VvTPPA protein will alter the TPP activity in the plant.

VvTPPA displays heterologous TPP activity. a Heterologous expression of VvTPPA leads to the complementation of the yeast tps2Δ heat sensitivity at 38.6°C. ScTPS2- and AtTPPA-transformed tps2Δ strains were added as positive controls; empty vector-transformed tps2Δ as negative control. b To exclude complications with growth phase differences, quantitative spot assay was performed with overnight grown samples, diluted to and spotted with OD6001. Western blot analysis was performed to confirm the expression of the HA-tagged proteins. Asterisks indicate the full-length tagged protein, based on the predicted mass and based on the pre-stained protein standard
Fluctuations of trehalose metabolism following chilling in grapevine tissues
To follow the response of trehalose metabolism upon chilling, we performed qRT-PCR analysis to determine changes in the expression of the VvTPS1 and VvTRE genes, which are likely to encode the only catalytically active TPS and trehalase enzymes in grapevine, and the VvTPPA gene, which was shown to encode a functional TPP enzyme, and is the most widely expressed TPP isoform in this species (Fig. 4). To complement this expression analysis, the T6P and trehalose contents of roots, stems and leaves were measured (Fig. 5).

Expression of V. vinifera L. trehalose-related genes upon chilling (4°C). Levels of VvTPS1 (a, d), VvTPPA (b, e), VvTRE (c, f) transcripts were measured by qRT-PCR in roots (open square), stems (filled square), and leaves (filled square) of grapevine plants non-bacterised (a–c) or bacterised (d–f). The results represent the mean fold increase (±SD) of mRNA level over non-bacterised plants grown at 26°C (referred as 1× expression level) of three independents experiments

T6P and trehalose accumulation in V. vinifera L. plants upon chilling (4°C). T6P content was measured by LC–MS/MS in roots (a), stems (b) and leaves (c). Trehalose content was measured by fluorimetry in stems (d) and leaves (e). Roots trehalose levels were below detection limit. Both metabolites were quantified in non-chilled control plants (open square) and chilled plants (filled square). Values are mean ± SD of samples from three independent experiments. Asterisks indicate significant difference (P < 0.05). n.d. not determined due to excessive background fluorescence in the trehalose assay
Trehalose-related gene expression in grapevine organs
No change in the expression level of VvTPS1 was detected in grapevine organs upon chilling stress (Fig. 4a). In contrast, a substantial increase in the abundance of the VvTPPA transcript was detected in roots, stems and leaves within 6 h of chilling treatment. The expression of VvTPPA reached a maximum in roots and stems after 9 h (6- and 9-fold higher than non-chilled controls, respectively, Fig. 4b), falling back after 24 h. In leaves, induction was lower (maximum 3-fold) but constant during the first 24 h (Fig. 4b). Interestingly, the VvTRE transcripts showed an opposite response in leaves, with a transient down-regulation during the first 9 h (Fig. 4c), but no change in roots and stems (data not shown).
T6P and trehalose accumulation
Roots, stems and leaves from non-stressed plants contained very low amounts of T6P: 0.2, 0.2 and 0.4 nmol g−1 FW, respectively (Fig. 5a–c). In roots and stems, a significant increase in T6P was detected after 48 h at 4°C (Fig. 5a, b). T6P also increased in leaves, but even more quickly, with a more than twofold increase within 24 h after chilling exposure (Fig. 5c). In all organs, T6P concentration attained a maximum after 72 h chilling, reaching 0.5, 0.8 and 1.9 nmol g−1 FW in roots, stems and leaves, respectively. Since the earliest and strongest increases in T6P were detected in leaves, T6P was also measured in leaf samples from plants exposed to shorter periods of chilling. After 3 and 9 h of chilling exposure, the level of T6P in the leaves had reached 0.6 ± 0.05 and 1.2 ± 0.09 nmol g−1 FW, respectively (i.e. 1.5- and 3-fold higher than in control leaves). Interestingly, the levels of T6P were highly correlated with those of sucrose in both chilled and non-chilled plants (Fig. S6).
Trehalose could not be detected in roots of either control or chilling treated plants, most likely because concentrations were below the detection limit (0.5 nmol g−1 FW), rather than due to interference with the trehalose assay by something in the root extract, because we were able to detect trehalose in roots of bacterised plants (see below). In stems and leaves of non-chilled plants, the level of trehalose was 10 times higher than that of T6P (Fig. 5d, e). The trehalose content of the stems was the same in both chilled and control plants (Fig. 5d), but increased 2.5-fold in leaves after 72 h at 4°C (Fig. 5e).
Fluctuations of trehalose metabolism in chilled plants when bacterised with B. phytofirmans strain PsJN
Trehalose-related gene expression
Bacterisation with B. phytofirmans leads to improved chilling tolerance in grapevine. We investigated the possibility that this bacteria induced chilling tolerance might be linked to changes in trehalose metabolism in the plants by analysing the expression of the VvTPS1, VvTPPA and VvTRE genes in bacterised plants upon chilling. No differences in the levels of expression of these genes were detected in bacterised versus non-bacterised plants at 4 weeks after inoculation (data not shown). As observed previously for non-bacterised plants (Fig. 4a), chilling exposure did not modify expression of VvTPS1 (Fig. 4d) but did lead to up-regulation of VvTPPA (Fig. 4e), although the increase was delayed compare to non-bacterised plants (Fig. 4b, e). In contrast, VvTRE showed the opposite response to chilling in bacterised plants as compared to non-bacterised plants, being twofold up-regulated in the former, but repressed in the latter (Fig. 4c, f).
T6P and trehalose accumulation
T6P and trehalose were measured in bacterised plants 24 and 120 h after chilling exposure, when differences in these metabolites were clearly noticeable in non-bacterised plants, especially in leaves. Before chilling exposure, the amounts of T6P were similar in roots from both bacterised and non-bacterised plants (Fig. 6a), whereas higher levels of T6P were detected in both stems and leaves of bacterised plants as compared to the non-bacterised controls (Fig. 6b, c). Interestingly, leaves of bacterised plants that had not been exposed to chilling had a similar T6P content to those from non-bacterised plants treated at 4°C for 24 h (Fig. 6a). After 120 h of chilling, T6P increased in roots, stems and leaves of bacterised plants, reaching similar levels to those observed in non-bacterised plants (Fig. 6a–c).

T6P and trehalose accumulation in V. vinifera L. plants bacterised with B. phytofirmans strain PsJN upon chilling (4°C). T6P content was measured by LC–MS/MS in roots (a), stems (b) and leaves (c). Trehalose content was measured by fluorimetry in roots (d), stems (e) and leaves (f). Both metabolites were quantified in non-bacterised (open square) and bacterised plants (filled square). Values are mean ± SD of samples from three independent experiments. Asterisks indicate significant difference (P < 0.05). n.d. not determined due to excessive background fluorescence in the trehalose assay, b.d. below detection limit
In roots of bacterised plants, whether chilled or not, we detected more than 90 nmol g−1 FW of trehalose (Fig. 6d), but we were unable to detect trehalose in the corresponding stem samples (Fig. 6e). Leaves of bacterised plants that had not been exposed to chilling had more than twice the amount of trehalose than leaves of non-bacterised plants maintained at 26°C (Fig. 6f). Upon chilling, the level of trehalose fell by half in leaves of bacterised plants after 24 h at 4°C, and then increased again after 120 h of chilling exposure, reaching similar values to those observed in leaves of non-bacterised plants after the same cold treatment (Fig. 6f).
Discussion
Transcriptional regulation of trehalose metabolism in grapevine in response to chilling
As expected from previous surveys of trehalose-related genes in plants (Leyman et al. 2001; Avonce et al. 2006; Lunn 2007), the genome of V. vinifera L. contains multiple TPS and TPP genes. However, with only seven TPS and six TPP genes, these gene families are somewhat smaller than those found in other species, such as A. thaliana (11 TPS/10 TPP), rice (Oryza sativa; 10/11) and Populus trichocarpa (12/10; Lunn 2007). The profusion of TPS and TPP genes in these plants is thought to have originated from several duplication events, including relatively recent whole genome duplications (Lunn 2007; Paul et al. 2008), but the genome of V. vinifera L. shows no evidence of a recent duplication (Jaillon et al. 2007), which may account for the smaller number of TPS and TPP genes in this species. V. vinifera L. contains a single TRE gene, like many other diploid angiosperms, with a few exceptions, such as P. trichocarpus which has three TRE genes (Lunn 2007). The number of TPS and TPP genes in the grapevine genome is subject to upward revision as remaining gaps in the sequence are filled and the assembly and annotation of the genome sequence is updated.
No evidence was found for transcriptional regulation of VvTPS1 in response to chilling. An in silico analysis performed by Iordachescu and Imai (2008) revealed similar results for the A. thaliana orthologue AtTPS1. In agreement with our findings, it was recently demonstrated that the rice orthologue, OsTPS1, is also not induced by cold (Li et al. 2011). The fact that T6P accumulated in grapevine upon exposure to chilling, whereas the VvTPS1 transcript level remain unchanged suggests that the rise in T6P was not brought about by transcriptional regulation of VvTPS1.
Chilled plants are characterised by a global rise in TPP relative transcript number in all plant organs (Fig. 2). As a good marker of this rise in TPP transcript level, VvTPPA was induced in all parts of the plant by chilling stress, but to varying degrees in different organs. TPP genes are known to be induced in plants by several abiotic stresses, such as cold, heat, salt and osmotic stress (Iordachescu and Imai 2008). In rice, two VvTPPA homologues, OsTPP1 and OsTPP2, are transiently induced upon chilling (Pramanik and Imai 2005). In A. thaliana, both AtTPPA and AtTPPD are cold inducible (Iordachescu and Imai 2008).
The induction of VvTPPA by chilling was higher in roots and stems than in leaves. Differential responses of TPP genes to abiotic stresses have previously been reported in maize, with drought leading to induction of TPP expression in ears but repression in tassels (Zhuang et al. 2007). An intriguing finding in grapevine was the strong induction of VvTPPA (Figs. 2, 4) together with a similarly strong induction of VvTPPE transcripts in stems (Fig. 2), where no trehalose accumulation was detected. Assuming that induction of these two isoforms transcript abundance in stems gave rise to higher TPP activity and increased synthesis of trehalose, the lack of accumulation of this sugar in stems might be explained by export to other parts of the plant via the phloem, or by trehalase-mediated hydrolysis of the trehalose into glucose.
VvTPPA (as well as other TPPs transcripts) and VvTRE transcript variations could account for the chilling induced accumulation of trehalose in leaves. Chilling treatment led to a moderate, but sustained induction of VvTPPA in leaves, whereas VvTRE expression was transiently down-regulated. Both of these responses are consistent with the observed accumulation of trehalose in the leaves, suggesting that this response is brought about by transcript level regulation. To our knowledge, there are no previous reports of a TRE gene expression being regulated by cold. However, the MtTRE gene from Medicago truncatula was found to be down-regulated upon salt stress (López et al. 2008).
T6P and trehalose concentrations in non-chilled plants
T6P levels were slightly higher in leaves from non-chilled grapevine plants than in leaves of soil-grown A. thaliana plants (Lunn et al. 2006). The grapevine plants were grown on sucrose-containing media, which might account for their higher T6P, as it has previously been reported that exogenous supply of sucrose to A. thaliana seedlings dramatically increased the level of T6P (Lunn et al. 2006). In contrast, trehalose levels in non-chilled grapevine samples were somewhat lower than those previously measured in A. thaliana (Avonce et al. 2004; Miranda et al. 2007; Veyres et al. 2008). The latter measurements were made using HPLC-based methods, which are susceptible to over-estimation of trehalose if other sugars are not fully resolved from trehalose during the chromatographic separation, whereas the fluorimetric assay we used to measure trehalose enzymatically is very specific. This difference is illustrated by comparison of the trehalose content of A. thaliana plants (grown at 20°C), which were found to be in the range of 6–9 nmol g−1 FW using the fluorimetric assay, but more than 30 nmol g−1 FW using high pressure anion exchange chromatography with pulsed amperometric detection (R. Feil, unpublished data).
The potential role of T6P as a signal molecule in the response to chilling
It has been postulated that T6P acts as a signal metabolite in plants, responding in particular to changes in sucrose content (Lunn et al. 2006). The levels of T6P in roots, stems and leaves of grapevine were all increased in response to chilling, changing more consistently, and more rapidly than trehalose. The response was particularly rapid in leaves, where T6P was found to have increased within 3 h of exposure to chilling. We observed a strong correlation between T6P and sucrose content in all plant organs from both chilled and non-chilled control plants (Fig. S6). This suggests that the rise in T6P levels in plants exposed to chilling might be due to cold induced accumulation of sucrose, although a direct, sucrose-independent response of T6P to the cold treatment cannot be excluded. Irrespective of the mechanism behind the rise in T6P, it is likely that this signal metabolite will affect its downstream targets. For example, high T6P is likely to inhibit the SnRK1 protein kinase, leading to multiple effects on transcription, protein synthesis and enzymatic activities. Although the physiological significance of the cold induced increase in T6P is uncertain, it seems reasonable to conclude that this signal metabolite could play some part in the plant’s response to chilling, although its exact function is uncertain.
The role of trehalose during chilling in grapevine remains unclear with a substantial increase in trehalose content being observed only in leaves upon exposure to chilling. Even in cold-treated plants, the total amount of trehalose was low (<15 nmol g−1 FW), indicating that the intracellular concentration of trehalose would be too low to make a major contribution to overall osmoregulation (Gibon et al. 1997). However, almost nothing is known about the subcellular distribution of trehalose in plants, so we cannot exclude the possibility that trehalose is localised within a specific organelle or subcellular compartment, and acts as a compatible osmolyte within that part of the cell (Fernandez et al. 2010). The observation that chloroplast specific over-expression of enzymes for trehalose synthesis in tobacco led to improved drought tolerance supports this hypothesis (Karim et al. 2007). Localised high concentrations of trehalose might also act to protect membranes from the effects of cold induced dehydration (Crowe 2007; Ruelland et al. 2009) or oxidative damage as reported in wheat exposed to heat stress (Luo et al. 2008). Trehalose accumulation was restricted to the leaves and only occurred after 72 h of cold exposure, which was much later than the increase in T6P, which argues against trehalose having a general signalling function during the chilling response.
T6P and trehalose might participate in chilling resistance induced by B. phytofirmans strain PsJN
The rhizobacterium B. phytofirmans has been shown to induce chilling resistance in grapevine (Ait Barka et al. 2006). It has also been reported that association with both symbiotic and pathogenic bacteria can influence trehalose metabolism in plants (Brodmann et al. 2002; Streeter and Gomez 2006; López et al. 2007; Gravot et al. 2011). Therefore, we were interested to know whether B. phytofirmans might also be affecting trehalose metabolism, and thereby cold tolerance, in grapevine.
Bacterisation with B. phytofirmans was found to modify the expression of VvTRE upon chilling. Before chilling stress the expression levels of trehalose-related genes were similar in both bacterised and non-bacterised plants. After chilling exposure, both VvTPS1 and VvTPPA exhibited qualitatively similar responses in bacterised and non-bacterised plants. In marked contrast, expression of VvTRE was slightly up-regulated in the leaves of bacterised plants but clearly down-regulated in non-bacterised ones (Fig. 4c, f). Increased expression of VvTRE might be expected to increase trehalase activity in the leaves of bacterised plants, which would be entirely consistent with the transient reduction in trehalose content observed in these leaves after 24 h of chilling exposure (Fig. 6f).
B. phytofirmans also stimulated plant T6P synthesis. In non-chilled plants maintained at 26°C, no difference in T6P content was detected in root tissues from bacterised plants as compared to non-bacterised ones. It has been shown that the endophytic bacterium B. phytofirmans establishes large populations within the roots of grapevine when growing in association with this species (Compant et al. 2005) and we measured amount of trehalose higher than 90 nmol g−1 FW in roots of bacterised plants and such a high level of trehalose is unlikely to be of plant origin. Our finding that T6P levels in roots were unchanged by bacterisation suggest that the B. phytofirmans cells contribute very little to the overall amount of T6P detected in bacterised plants, and that association with this bacterium does not lead to accumulation of T6P by the root cells. However, accumulation of T6P was observed in both stems and leaves of bacterised plants before chilling, indicating a stimulation of T6P synthesis or inhibition of T6P degradation caused by the bacterial association. This increase in T6P might be linked to the increase in total soluble sugars, especially sucrose that has been reported to occur upon bacterisation (Ait Barka et al. 2006). Supporting this assertion, a positive correlation between T6P and sucrose concentrations in bacterised plants has been found (Fig. S6d).
Association with B. phytofirmans did not compromise the further increase in T6P content induced by chilling, with T6P reaching similar levels in both bacterised and non-bacterised plants after 120 h exposure to chilling (4°C). Furthermore, T6P concentrations in non-chilled bacterised plants were similar to those of non-bacterised ones after 24 h at 4°C (Fig. 6a–c). Altogether, these observations suggest that any influence of B. phytofirmans on T6P content depends on its effect on the plants before exposure to chilling.
B. phytofirmans modified the trehalose content of both roots and leaves, which were higher in bacterised plants than in non-bacterised plants that had not been exposed to chilling. Accumulation of trehalose in plants associated with beneficial micro-organisms has been documented especially in rhizobial nodules (Farias-Rodriguez et al. 1998; Streeter and Gomez 2006) as well as in roots associated with ectomycorrhizae (López et al. 2007), and has been correlated with drought resistance (Farias-Rodriguez et al. 1998). The trehalose accumulated in the roots of bacterised grapevine plants is probably of bacterial origin since the bacterial population is high in roots and closely associated with the grapevine cells (Compant et al. 2005; Bordiec et al. 2010). In contrast, the leaves are poorly colonised by the bacteria; hence, trehalose accumulation in leaves of bacterised plants is more likely to result from stimulation of trehalose metabolism, which is consistent with the higher T6P concentration in these leaves. Therefore, stimulation of trehalose metabolism may represent a part of the mechanism by which B. phytofirmans strain PsJN confers chilling tolerance to grapevine.
Upon chilling, we observed a transient fall in the trehalose content of leaves from bacterised plants after 24 h at 4°C, followed later by a recovery after 120 h trehalose to levels seen in leaves of non-bacterised plants. These data suggest that the transient reduction in trehalose content, together with up-regulation of the expression of VvTRE, might be an induced response in plants to avoid interference with T6P/trehalose signalling as it was recently proposed (Brodmann et al. 2002; Gravot et al. 2011).
In this paper, we present the first set of data showing significant fluctuations in trehalose metabolism of grapevine during cold stress. Variations in trehalose and T6P content, as well as related gene regulation are organ dependent whether they occur in plants colonised with a PGPR or not. In non-bacterised plants, our results are consistent with a role for T6P as a signal molecule in the response to chilling stress, possibly linked to changes in sucrose concentration (Lunn et al. 2006). Nevertheless, since T6P and trehalose accumulation are induced in chilling-tolerant bacterised plants, we cannot rule out a participation of both metabolites in triggered cold tolerance.
Abbreviations
- HAD:
-
l-2-Haloacid dehalogenase
- PGPR:
-
Plant growth-promoting rhizobacteria
- SnRK1:
-
Sucrose-non-fermenting-1-related kinase 1
- T6P:
-
Trehalose-6-phosphate
- TPS:
-
Trehalose phosphate synthase
- TPP:
-
Trehalose phosphate phosphatase
- TRE:
-
Trehalase
References
Ait Barka E, Belarbi A, Hachet C, Nowak J, Audran J-C (2000) Enhancement of in vitro growth and resistance to gray mould of Vitis vinifera co-cultured with plant growth-promoting rhizobacteria. FEMS Microbiol Lett 186:91–95
Ait Barka E, Nowak J, Clément C (2006) Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl Environ Microbiol 72:7246–7252
Argüelles JC (2000) Physiological roles of trehalose in bacteria and yeasts: a comparative analysis. Arch Microbiol 174:217–224
Avonce N, Leyman B, Mascorro-Gallardo JO, van Dijck P, Thevelein JM, Iturriaga G (2004) The Arabidopsis trehalose-6-P synthase AtTPS1 gene is a regulator of glucose, abscisic acid, and stress signaling. Plant Physiol 136:3649–3659
Avonce N, Mendoza-Vargas A, Morett E, Iturriaga G (2006) Insights on the evolution of trehalose biosynthesis. BMC Evol Biol 6:109
Bae H, Herman E, Bailey B, Bae H-J, Sicher R (2005a) Exogenous trehalose alters Arabidopsis transcripts involved in cell wall modification, abiotic stress, nitrogen metabolism, and plant defense. Physiol Plant 125:114–126
Bae H, Herman E, Sicher R (2005b) Exogenous trehalose promotes non-structural carbohydrate accumulation and induces chemical detoxification and stress response proteins in Arabidopsis thaliana grown in liquid culture. Plant Sci 168:1293–1301
Bertamini M, Muthuchelian K, Rubinigg M, Zorer R, Nedunchezhian N (2005) Low-night temperature (LNT) induced changes of photosynthesis in grapevine (Vitis vinifera L.) plants. Plant Physiol Biochem 43:693–699
Bertamini M, Muthuchelian K, Rubinigg M, Zorer R, Velasco R, Nedunchezhian N (2006) Low-night temperature increased the photoinhibition of photosynthesis in grapevine (Vitis vinifera L. cv. Riesling) leaves. Environ Exp Bot 57:25–31
Blázquez MA, Santos E, C-l Flores, Martínez Zapater JM, Salinas J, Gancedo C et al (1998) Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J 13:685–689
Bordiec S, Paquis S, Lacroix H, Dhondt S, Ait Barka E, Kauffmann S, Jeandet P, Mazeyrat-Gourbeyre F, Clément C, Baillieul F, Dorey S et al (2010) Comparative analysis of defence responses induced by the endophytic plant growth-promoting rhizobacterium Burkholderia phytofirmans strain PsJN and the non-host bacterium Pseudomonas syringae pv. pisi in grapevine cell suspensions. J Exp Bot 62:595–603
Boyer JS (1982) Plant productivity and environment. Science 218:443–448
Bray E, Bailey-Serres J, Weretilnyk E (2000) Responses to abiotic stresses. In: Buchanan B, Gruissem W, Jones R (eds) Biochemistry and molecular biology of plants. American Society of Plant Physiology, Rockville, pp 1158–1203
Brodmann D, Schuller A, Ludwig-Muller J, Aeschbacher RA, Wiemken A, Boller T, Wingler A (2002) Induction of trehalase in Arabidopsis plants infected with the trehalose-producing pathogen Plasmodiophora brassicae. Mol Plant Microbe Interact 15:693–700
Cao Y, Wang Y, Dai B, Wang B, Zhang H, Zhu Z, Xu Y, Cao Y, Jiang Y, Zhang G (2008) Trehalose is an important mediator of Cap1p oxidative stress response in Candida albicans. Biol Pharm Bull 31:421–425
Chen D-C, Yang B-C, Kuo T–T (1992) One-step transformation of yeast in stationary phase. Curr Genet 21:83–84
Compant S, Reiter B, Sessitsch A, Nowak J, Clement C, Ait Barka E et al (2005) Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl Environ Microbiol 71:1685–1693
Couée I, Sulmon C, Gouesbet G, El Amrani A (2006) Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J Exp Bot 57:449–459
Crowe JH (2007) Trehalose as a “chemical chaperone” fact and fantasy. In: Back N, Cohen IR, Lajtha A, Lambris JD, Paoletti R (eds) Molecular aspects of the stress response: chaperones, membranes and networks. Springer, New York, pp 143–158
Drennan PM, Smith MT, Goldsworth D, Van Staden J (1993) The occurence of trehalose in the leaves of the desiccation-tolerant angiosperm Myrothamnus flabellifolius welw. J Plant Physiol 142:493–496
Elbein AD, Pan YT, Pastuszak I, Carroll D (2003) New insights on trehalose: a multifunctional molecule. Glycobiology 13:17–27
Farias-Rodriguez R, Mellor RB, Arias C, Pena-Cabriales JJ (1998) The accumulation of trehalose in nodules of several cultivars of common bean (Phaseolus vulgaris) and its correlation with resistance to drought stress. Physiol Plant 102:353–359
Fernandez O, Béthencourt L, Quero A, Sangwan RS, Clément C (2010) Trehalose and plant stress responses: friend or foe? Trends Plant Sci 15:409–417
Gibon Y, Bessieres MA, Larher F (1997) Is glycine betaine a non-compatible solute in higher plants that do not accumulate it? Plant Cell Environ 20:329–340
Goddijn OJM, van Dun K (1999) Trehalose metabolism in plants. Trends Plant Sci 4:315–319
Gravot A, Grillet L, Wagner G, Jubault M, Lariagon C, Baron C, Deleu C, Delourme R, Bouchereau A, Manzanares-Dauleux MJ (2011) Genetic and physiological analysis of the relationship between partial resistance to clubroot and tolerance to trehalose in Arabidopsis thaliana. New Phytol 191:1083–1094
Hendrickson L, Ball MC, Wood JT, Chow WS, Furbank RT (2004) Low temperature effects on photosynthesis and growth of grapevine. Plant Cell Environ 27:795–809
Hirsh AG (1987) Vitrification in plants as a natural form of cryoprotection. Cryobiology 24:214–228
Hohmann S, Neves MJ, Koning W, Alijo R, Ramos J, Thevelein JM (1993) The growth and signalling defects of the ggs1 (fdp1/byp1) deletion mutant on glucose are suppressed by a deletion of the gene encoding hexokinase PII. Curr Genet 23:281–289
Iordachescu M, Imai R (2008) Trehalose biosynthesis in response to abiotic stresses. J Integr Plant Biol 50:1223–1229
Jaillon O, Aury J-M, Noel B, for The French-Italian Public Consortium for Grapevine Genome Characterization, et al (2007) The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 449:463–467
Janská A, Maršík P, Zelenková S, Ovesná J (2010) Cold stress and acclimation—what is important for metabolic adjustment? Plant Biol 12:395–405
Kandror O, DeLeon A, Goldberg AL (2002) Trehalose synthesis is induced upon exposure of Escherichia coli to cold and is essential for viability at low temperatures. Proc Natl Acad Sci USA 99:9727–9732
Kandror O, Bretschneider N, Kreydin E, Cavalieri D, Goldberg AL (2004) Yeast adapt to near-freezing temperatures by STRE/Msn2,4-dependent induction of trehalose synthesis and certain molecular chaperones. Mol Cell 13:771–781
Karim S, Aronsson H, Ericson H, Pirhonen M, Leyman B, Welin B, Mäntylä E, Palva E, van Dijck P, Holmström K-O (2007) Improved drought tolerance without undesired side effects in transgenic plants producing trehalose. Plant Mol Biol 64:371–386
Leyman B, van Dijck P, Thevelein JM (2001) An unexpected plethora of trehalose biosynthesis genes in Arabidopsis thaliana. Trends Plant Sci 6:510–513
Li H-W, Zang B-S, Deng X-W, Wang X-P (2011) Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 234:1007–1018
Liu M-S, Chien C-T, Lin T-P (2008) Constitutive components and induced gene expression are involved in the desiccation tolerance of Selaginella tamariscina. Plant Cell Physiol 49:653–663
López MF, Männer P, Willmann A, Hampp R, Nehls U (2007) Increased trehalose biosynthesis in Hartig net hyphae of ectomycorrhizas. New Phytol 174:389–398
López M, Tejera NA, Iribarne C, Lluch C, Herrera-Cervera JA (2008) Trehalose and trehalase in root nodules of Medicago truncatula and Phaseolus vulgaris in response to salt stress. Physiol Plant 134:575–582
Lunn JE (2007) Gene families and evolution of trehalose metabolism in plants. Funct Plant Biol 34:550–563
Lunn JE, Feil R, Hendriks JHM, Gibon Y, Morcuende R, Osuna D, Scheible W-R, Carillo P, Hajirezaei M-R, Stitt M (2006) Sugar-induced increases in trehalose 6-phosphate are correlated with redox activation of ADP-glucose pyrophosphorylase and higher rates of starch synthesis in Arabidopsis thaliana. Biochem J 397:139–148
Luo Y, Li W-M, Wang W (2008) Trehalose: protector of antioxidant enzymes or reactive oxygen species scavenger under heat stress? Environ Exp Bot 63:378–384
Miranda J, Avonce N, Suárez R, Thevelein J, van Dijck P, Iturriaga G (2007) A bifunctional TPS–TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta 226:1411–1421
Mollo L, Martins MCM, Olivera VF, Nievola CC, Figuereido-Ribeiro RCL (2011) Effects of low temperature on growth and non-structural carbohydrates of the imperial bromeliad Alcantarea imperialis cultured in vitro. Plant Cell Tissue Organ Cult. doi:10.1007/s11240-011-9966-y
Paul MJ, Primavesi LF, Jhurreea D, Zhang Y (2008) Trehalose metabolism and signaling. Annu Rev Plant Biol 59:417–441
Pillay VK, Nowak J (1997) Inoculum density, temperature, and genotype effects on in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L.) seedlings inoculated with a pseudomonad bacterium. Can J Microbiol 43:354–361
Pramanik MHR, Imai R (2005) Functional identification of a trehalose 6-phosphate phosphatase gene that is involved in transient induction of trehalose biosynthesis during chilling stress in rice. Plant Mol Biol 58:751–762
Purvis JE, Yomano LP, Ingram LO (2005) Enhanced trehalose production improves growth of Escherichia coli under osmotic stress. Appl Environ Microbiol 71:3761–3769
Ramon M, De Smet IVE, Vandesteene L, Naudts M, Leyman B, van Dijck P, Rolland F, Beeckman TOM, Thevelein JM (2009) Extensive expression regulation and lack of heterologous enzymatic activity of the class II trehalose metabolism proteins from Arabidopsis thaliana. Plant Cell Environ 32:1015–1032
Rao KN, Kumaran D, Seetharaman J, Bonanno JB, Burley SK, Swaminathan S (2006) Crystal structure of trehalose-6-phosphate phosphatase–related protein: biochemical and biological implications. Protein Sci 15:1735–1744
Reignault PH, Cogan A, Muchembled J, Lounes-Hadj Sahraoui A, Durand R, Sancholle M et al (2001) Trehalose induces resistance to powdery mildew in wheat. New Phytol 149:519–529
Renard-Merlier D, Randoux B, Nowak E, Farcy F, Durand R, Reignault PH (2007) Iodus 40, salicylic acid, heptanoyl salicylic acid and trehalose exhibit different efficacies and defence targets during a wheat/powdery mildew interaction. Phytochemistry 68:1156–1164
Ruelland E, Vaultier M-N, Zachowski A, Hurry V, Kader J-C, Delseny M (2009) Cold signalling and cold acclimation in plants. Adv Bot Res 49:35–150
Steponkus PL (1984) Role of the plasma membrane in freezing injury and cold acclimation. Annu Rev Plant Physiol 35:543–584
Streeter JG, Gomez ML (2006) Three enzymes for trehalose synthesis in Bradyrhizobium cultured bacteria and in bacteroids from soybean nodules. Appl Environ Microbiol 72:4250–4255
Terrier N, Glissant D, Grimplet J, Barrieu F, Abbal P, Couture C, Ageorges A, Atanassova R, Léon C, Renaudin J-P, Dédaldéchamp F, Romieu C, Delrot S, Hamdi S (2005) Isogene specific oligo arrays reveal multifaceted changes in gene expression during grape berry (Vitis vinifera L.) development. Planta 222:832–847
van Dijck P, Mascorro-Gallardo JO, De Bus M, Royackers K, Iturriaga G, Thevelein JM (2002) Truncation of Arabidopsis thaliana and Selaginella lepidophylla trehalose-6-phosphate synthase unlocks high catalytic activity and supports high trehalose levels on expression in yeast. Biochem J 366:63–71
Vandesteene L (2009) Functional analysis of the trehalose metabolism gene family in Arabidopsis thaliana. PhD thesis, Katholieke Universiteit Leuven, Belgium
Vandesteene L, Ramon M, Le Roy K, van Dijck P, Rolland F (2010) A single active Trehalose-6-P synthase (TPS) and a family of putative regulatory TPS-like proteins in Arabidopsis. Mol Plant 3:406–419
Veyres N, Danon A, Aono M, Galliot S, Karibasappa YB, Diet A, Grandmottet F, Tamaoki M, Lesur D, Pilard S, Boitel-Conti M, Sangwan-Norreel BS, Sangwan RS (2008) The Arabidopsis sweetie mutant is affected in carbohydrate metabolism and defective in the control of growth, development and senescence. Plant J 55:665–686
Xiao H, Tattersall EAR, Siddiqua MK, Cramer GR, Nassuth A (2008) CBF4 is a unique member of the CBF transcription factor family of Vitis vinifera and Vitis riparia. Plant Cell Environ 31:1–10
Yang J, Kloepper JW, Ryu C-M (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci 14:1–4
Zhang Y, Primavesi LF, Jhurreea D, Andralojc PJ, Mitchell RAC, Powers SJ, Schluepmann H, Delatte T, Wingler A, Paul MJ (2009) Inhibition of SNF1-related protein kinase1 activity and regulation of metabolic pathways by trehalose-6-phosphate. Plant Physiol 149:1860–1871
Zhuang Y, Ren G, Yue G, Li Z, Qu X, Hou G, Zhu Y, Zhang J (2007) Effects of water-deficit stress on the transcriptomes of developing immature ear and tassel in maize. Plant Cell Rep 26:2137–2147
Acknowledgments
This work was supported by the Region Champagne-Ardennes. Vitis vinifera L. Chardonnay clone 7535 was a gift from Moët et Chandon. B. phytofirmans strain PsJN was kindly provided by Dr. Essaid Ait Barka. We thank Pr. Petronia Carillo for her help in optimisation of the trehalose assay for grapevine.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
425_2012_1611_MOESM1_ESM.ppt
Fig. S1 Phylogenetic tree of TPSs isoforms in V. vinifera L. and A. thaliana. A phylip phylogenetic tree was constructed using Clustwal W2—Phylogeny program. The neighbour-joining algorithm was used to construct tree from the distance matrix. Sequences were obtained from the NCBI Gene Bank. Tree was drawn in a radial form using TreeView software (http://www.taxonomy.zoology.gla.ac.uk/rod/treeview.html) (PPT 55 kb)
425_2012_1611_MOESM2_ESM.ppt
Fig. S2 Alignment of VvTPS1, AtTPS1, OsTPS1 and SlTPS1 protein sequences. Amino acid residues that are similar in all sequences are shaded in grey and the ones that are predicted to be important for substrate binding (according to Lunn 2007; Vandesteene et al. 2010) in catalytically active TPS are shaded in red. All of these residues are perfectly conserved in the four sequences (PPT 139 kb)
425_2012_1611_MOESM3_ESM.ppt
Fig. S3 Alignment of VvTPPA-F and TaTPP (from Thermoplasma acidophilum). Amino acid residues that are similar in all sequences are shaded in grey and the ones that are predicted to be important for substrate binding in catalytically active TPP enzymes (according to Lunn 2007) are shaded in red. All of these residues are perfectly conserved in the 6 VvTPP sequences (except one conservative substitution of Lys to Arg in four isoforms VvTPPA, VvTPPB, VvTPPC and VvTPPE) (PPT 105 kb)
425_2012_1611_MOESM4_ESM.ppt
Fig. S4 Relative transcript number of VvTPPs non-chilled (a) and chilled (b) grapevine plants. Transcript number was calculated using the standard curve method and normalized with the Ef1α gene as internal control. Results represent the mean (± SD) of transcript number analysed from three independent experiments. Plants were exposed to chilling during 9 h, a time point when all VvTPPs gene induction peaked. n.d not detected; ds: detected only in stems; dl : detected only in leaves (PPT 59 kb)
425_2012_1611_MOESM5_ESM.doc
Fig. S5 Protein coding region of the V. vinifera L. cv. Chardonnay TPPA cDNA and the deduced protein sequence. Differences from the orthologous V. vinifera L. cv. Pinot Noir cDNA and protein are highlighted in red (DOC 25 kb)
425_2012_1611_MOESM6_ESM.ppt
Fig. S6 Correlation between T6P and sucrose in roots, stems and leaves of V. vinifera L. plants. Correlation was evaluated in all samples (a), in non-chilled samples (b), in chilled samples (c) and in bacterised samples (d). All R² values are significant (P < 0.05) (PPT 59.5 kb)
Rights and permissions
About this article
Cite this article
Fernandez, O., Vandesteene, L., Feil, R. et al. Trehalose metabolism is activated upon chilling in grapevine and might participate in Burkholderia phytofirmans induced chilling tolerance. Planta 236, 355–369 (2012). https://doi.org/10.1007/s00425-012-1611-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-012-1611-4