
Abstract
Astaxanthin is commercially sold as a pigment for animal feed and as an antioxidant for the nutraceutical sector. Astaxanthin is predominantly manufactured synthetically from petrochemicals but is also obtained from the chlorophyte Haematococcus pluvialis (Haematococcus lacustris). The petrochemical-derived synthetic alternative has conventionally been used, attributable to its lower cost ($1300–1800 kg−1). However, it is inferior as an antioxidant, prohibited for direct human consumption, and may cause toxicity in the final product. Conventionally, astaxanthin from H. lacustris is produced in a two-stage production process, incorporating a green and red stage for maximising growth and astaxanthin production, respectively, but a one-stage process has been proposed. The H. lacustris-derived astaxanthin industry has been a commercial success, but several constraints have arisen including contamination issues, relatively low biomass and astaxanthin productivities, high downstream processing costs, and photobleaching issues in the red stage. These constraints need to be addressed for the production of astaxanthin from H. lacustris for the aquaculture sector. Alternatively, through the exploitation of an alternative life cycle stage, red motile macrozooids can be formed lacking the thick walls of aplanospores. It is envisaged that the red motile macrozooids could be harvested and fed as a whole-cell product directly to the aquaculture sector rich in astaxanthin and polyunsaturated fatty acids, bypassing the cell disruption and extraction steps to deliver bioavailable astaxanthin as a biobased feed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
1.1 Carotenoids and Their Chemistry
Carotenoids are a family of greater than 600 naturally occurring pigments synthesised by higher plants, algae, fungi, and bacteria (Yaakob et al. 2014). Around 40 carotenoids are typically present in the human diet (BCC Research 2015). The chemical structure of carotenoids is derived from the carotenoid lycopene (C40H56). Carotenoids are mainly hydrocarbons which have two terminal rings joined by a conjugated double-bond chain or polyene system (Yuan et al. 2011). Two major groups of carotenoids have been characterised on the basis of their chemical structure: the carotenes (composed of carbon and hydrogen) and the xanthophylls (oxygenated derivatives). Astaxanthin, a xanthophyll, was found to be closely related to the other carotenoids, β-carotene, zeaxanthin, and lutein, and has many of the physiological and metabolic functions associated with carotenoids (Guerin et al. 2003). However, the presence of the hydroxyl and keto endings on each ionone ring reflect the unique properties, such as the ability to be esterified, a more polar configuration, and a higher antioxidant activity (inhibiting oxidation of other molecules) (Guerin et al. 2003). Each double bond from the polyene chain has been found to exist in two different configurations as geometric isomers cis or trans. Cis isomers are known to be thermodynamically less stable than trans isomers (Higuera-Ciapara et al. 2006) and in nature, most carotenoids predominate in the trans form (Stahl and Sies 2003).
Rodríguez-Sáiz et al. (2010) determined that astaxanthin contains two chiral centres and is present in three configurational isomers of the trans form (all-E isomer) (3R,3′R), (3R,3′S), and (3S,3′S). The (3S,3′S) form is the most abundant astaxanthin isomer in nature (Mont et al. 2010) and has been observed to be of the highest biotechnological value (Al-Bulishi 2015). Synthetic astaxanthin is generally composed of the three enantiomers (3R,3′R), (3R,3′S), and (3S,3′S) with a ratio of 1:2:1 and is unesterified, whereas astaxanthin from H. pluvialis is of the (3S,3′S stereoisomer), and 70% is in the monoester form, 10–15% in the diester form and 4–5% in the free form (Higuera-Ciapara et al. 2006; Ranga Rao et al. 2010; Young et al. 2017), which is also the main form in wild salmon (47.1–90%) (Young et al. 2017). The (3S,3′S) stereoisomer has been reported to impart a higher pigmentation in rainbow trout (Oncorhynchus mykiss) than other astaxanthin isomers and has been stated as the preferred additive for aquaculture (Choubert and Heinrich 1993). The 3S,3′S isomer has also been reported to have contributed to human health benefits, whereas the other forms have not been proven to have had positive biological effects (Capelli et al. 2013a; Guerin et al. 2003). Depending on their origin, astaxanthin can be found in association with other compounds such as proteins and biological lipids. In the case of Haematococcus pluvialis, up to 95% of astaxanthin molecules can be esterified with fatty acids (FAs) (commonly oleic, palmitic, and linoleic acid) (Lorenz and Cysewski 2000), with oleic acid as the major FA which is conjugated to astaxanthin molecules (Holtin et al. 2009). The synthetic form is found in the free, unesterified form as is astaxanthin derived from the yeast Xanthophyllomyces dendrorhous (Capelli et al. 2013a).
1.2 The Astaxanthin Market
The global carotenoid market reached US $1.5 billion in 2017 and is scheduled to reach US $2 billion by 2019 due to rising consumer awareness regarding the health benefits offered by the wide variety of carotenoids (BCC Research 2018). Panis and Rosales (2016) stated that in 2014 global astaxanthin production was 280 metric tons with a valuation of US $447, and the forecast for 2020 is US $1.5 billion (Panis and Rosales 2016; Allewaert et al. 2017; Molino et al. 2018). From industry reports, H. pluvialis-derived astaxanthin represents 5–8 tons (Pers. Com. Brevel Ltd.). Currently, 95% of the astaxanthin available in the market is generated synthetically from petrochemicals, <1% is produced from H. pluvialis, and the remainder is produced from the bacterium Paracoccus carotinifaciens, and the yeast Xanthophyllomyces dendrorhous (Koller et al. 2014; Panis and Rosales 2016; Shah et al. 2016). In 2009, 91% of commercial astaxanthin was used for animal feed pigments and 9.1% was used for nutraceuticals, with supply dominated by the synthetic form (Oilalgae 2015). It has been determined that the highest market share in 2016 (40%) was for the animal feed market (Market Watch 2019).
The market for astaxanthin has significantly grown from when it was first approved by the US Food and Drug Administration (FDA) in 1987 for its use as a feed additive in aquaculture and, over a decade later, when natural astaxanthin was subsequently approved to be used as a nutraceutical (Guerin et al. 2003). H. pluvialis-derived astaxanthin as a colour additive has been approved for salmonid feeds and additionally as a dietary supplement for human consumption in several European countries, the USA, and Japan (Yuan et al. 2011). To date, there is no European Food Safety Authority (EFSA) approval for the therapeutic application of H. pluvialis-derived astaxanthin. In line with EU 2015/2283, astaxanthin has been registered as a novel food and can be used to fortify foods equivalent to a maximum intake of 8 mg/day, but this is currently under review (https://www.efsa.europa.eu/sites/default/files/consultation/callsfordata/Callfordata-safetyassessmentofAstaxanthin.pdf). H. pluvialis-derived astaxanthin extracted using supercritical CO2 has been granted Novel Food status by the UK Foods Standard Agency (FSA), and the US FDA has granted astaxanthin from H. pluvialis GRAS certified (Generally Recognised as Safe) (Shah et al. 2016). EU regulation 2015/1415 has limited synthetic astaxanthin to <100 ppm/kg of fish feed, whereas natural astaxanthin is widely accepted as safe (FDA GRAS Notice. No. GRN 000294). In the salmon farming industry, 10–15% of the total feed cost is attributable to astaxanthin (Mann et al. 2000; Nguyen 2013).
The major synthetic producers for astaxanthin are BASF, Royal DSM, and Zhejiang NHU Co. Ltd with a selling price of $2000 (Koller et al. 2014), but current costs can be as low as $1300 kg−1 for pure astaxanthin for the aquaculture industry ($130 kg−1 for 10% astaxanthin) (Pers. Com. Brevel Ltd.). Natural astaxanthin can range from $2500–7150 kg−1 (Kim et al. 2016; Koller et al. 2014), but from industrial reports, the price of nutraceutical grade pure astaxanthin from H. pluvialis is $6000 kg−1 (Pers. Com. Brevel Ltd.). Currently, the estimated cost of production for synthetic astaxanthin is around $1000 kg−1 compared to H. pluvialis-derived astaxanthin costing around $3000–$3600 kg−1 (Li et al. 2011). Concerns have been raised for synthetic astaxanthin use in human consumption due to it being derived from petrochemicals, making astaxanthin from natural sources a preferred choice (Li et al. 2011). Furthermore, there is concern that synthetically synthesised astaxanthin could be linked to cancer (Newsome 1986), but this has not been substantiated to date. Nevertheless, synthetic astaxanthin has not undergone safety testing for direct human use and has not been recorded to provide health benefits to humans; thus, it has not been registered with regulatory authorities for direct human use in any country (Capelli et al. 2013a), with the exception of DSM’s AstaSana™. The level of synthetic food additives which are legally allowed into the market has steadily decreased due to suspected roles as promoters of carcinogenesis with additional claims of liver and renal toxicities (Guedes et al. 2011) creating stricter regulations for the human supplement market. Due to the high cost of production for H. pluvialis-derived astaxanthin and a requirement for low-cost astaxanthin in the animal feed sector, many H. pluvialis astaxanthin producers have targeted the higher value nutraceutical and pharmaceutical markets due to the numerous reported health benefits for natural astaxanthin (Guerin et al. 2003). However, H. pluvialis-derived astaxanthin has been found to be effective for animal feed through improved pigmentation of the flesh and skin, enhanced antioxidant potential, improved fish egg quality, increased growth, and survival of sea bream, rainbow trout, yellow croaker, and salmonid fry compared with the synthetic type (Li et al. 2014; Sheikhzadeh et al. 2012).
In the future, astaxanthin has the potential as a functional food, for example as a partial substitution of flour in cookies (Hossain et al. 2017). To be utilised in food matrices, further innovations in maintaining stability, preservation, encapsulation, and storage are required to avoid degradation and chemical changes (Martínez-delgado et al. 2017). The nutraceutical market today is dominated by astaxanthin from H. pluvialis, but one ‘nature identical’ synthetic product has entered the market, AstaSana™, manufactured by DSM. The Natural Algae Astaxanthin Association (NAXA) promotes H. pluvialis-derived astaxanthin (AlgaTechnologies Ltd., Cyanotech Corporation, Beijing Gingko Group (BGG) and Atacama Bio Natural), and they have been aiming to educate the public about the health benefits of natural astaxanthin and the major differences between the natural and synthetic form. A selection of the products are showcased in Fig. 6.1.

Examples of products from (a) Cyanotech Corporation and (b) Atacama Bio Natural. Astaxanthin capsules are typically sold with 4–12 mg of astaxanthin as a 10% oleoresin with astaxanthin extracted from the dried powder and formulated in edible oils. (a) BioAstin® Hawaiian Astaxanthin®. (Courtesy of Nutrex Hawaii). (b) NatAxtin™ ME: microencapsulated oleoresin powder and NatAxtin™ oil: astaxanthin-rich 10% oleoresin from supercritical CO2. (Courtesy of Atacama Bio Natural Products S.A., Chile). Astaxanthin from H. pluvialis cultivated in the Atacama Desert
High demand by consumers has led to many new companies (at least 22 companies from 13 countries) entering the market in recent years (Fig. 6.2), and producers in China such as BGG are projected to become leaders in the market (Capelli 2018). Additionally, research is being undertaken on other sources of astaxanthin from natural sources such as Chromochloris zofingiensis (Chen et al. 2017), with the intention to compete with synthetic, yeast, bacterial and H. pluvialis-derived astaxanthin.

Manufacturing locations of companies producing astaxanthin from H. pluvialis. The orange, red, and blue lines indicate the Tropic of Cancer, Equator, and the Tropic of Capricorn, respectively. There are 22 production companies with 77% of companies manufacturing outside of the tropics
1.3 Astaxanthin Health Benefits
Astaxanthin has been reported to be one of the most potent compounds in terms of its antioxidant activity, 65 times more powerful than vitamin C; 10 times stronger than β-carotene, canthaxanthin, zeaxanthin, and lutein; and 100 times more effective than alpha-tocopherol (Capelli et al. 2013a; Miki 1991). The free radical quenching ability is attributable to the conjugated structure of astaxanthin which allows the molecule to intercalate within the phospholipid bilayers of the biological membranes and the terminal hydroxylated ring structures which remain exposed to the outer and inner surfaces of the membrane (Riccioni et al. 2012). The demand for natural astaxanthin as a nutraceutical has exponentially increased due to a growing clinical evidence base including early-stage human trials with many reported health benefits, most notably a high antioxidant potential (Guerin et al. 2003). Dr Joseph Mercola, one of the world’s most followed physicians, had declared that astaxanthin was ‘the #1 supplement you’ve never heard of that you should be taking’. Natural astaxanthin, with a molecular structure containing polar hydrophilic ends, can move throughout the entire body (Yuan et al. 2011) and cross the blood-brain barrier to bring antioxidant protection to the brain and eyes (Fig. 6.3), crossing the phospholipid bilayer, a unique characteristic, which only a few other carotenoids possess, including lutein and zeaxanthin (Minatelli 2008). In rats, astaxanthin has been found to accumulate in the hippocampus and cerebral cortex (Manabe et al. 2018).

Astaxanthin has the capacity to cross the blood-brain barrier to provide antioxidant protection to the brain and eyes. The blood-brain barrier is a semipermeable membrane which separates the blood from the cerebrospinal fluid forming a barrier to the passage of cells and large molecules but allowing the diffusion of hydrophobic and small polar molecules. Tight junctions create the barrier, and lipophilic substances can pass through the membrane
It has been clearly revealed that the source of astaxanthin can have a positive impact on health with extensive preclinical (in vitro and animal models) and clinical studies (Yuan et al. 2011). Natural H. pluvialis-derived astaxanthin has been showcased to be over 50 times more effective than synthetic astaxanthin in singlet oxygen quenching and 20 times more effective in free radical elimination (Capelli et al. 2013a). It has been found that the bioaccessibility of H. pluvialis-derived astaxanthin in supplements was higher than in aquaculture-derived salmon where synthetic astaxanthin is used for pigmentation (Chitchumroonchokchai and Failla 2017). Research by Nishida et al. (2007) revealed that astaxanthin has a singlet oxygen quenching capability 800 times greater than ubiquinone (antioxidant present in most cells in the body). These initial results have led to further studies, which have concluded antilipid peroxidation activities in vitro (Leite et al. 2010), anticancer properties in vitro and in vivo with rodent models (Tanaka et al. 2012), immune system boosting activity (Bolin et al. 2010), eye health maintenance (Guerin et al. 2003; Piermarocchi et al. 2012), alleviation of arthritic symptoms (Capelli et al. 2013b), and protection against cognitive decline (Katagiri et al. 2012; Satoh et al. 2009). In mice models, H. pluvialis astaxanthin has been shown to inhibit the growth of Helicobacter pylori (common cause of peptic ulcers) and reduces bacterial load in the infected cells, but further research is needed to determine if this is the case in human patients (Kang and Kim 2017).
Most studies of H. pluvialis-derived astaxanthin have been in vitro and in animal models, and the efficacy has been well proven (Guerin et al. 2003; Visioli and Artaria 2017; Yuan et al. 2011). In addition, the health benefits of astaxanthin in human patients have been extensively reported: improvements in muscle endurance through reduced lactic acid and increased respiratory and sympathetic nervous system activities (Capelli et al. 2013b), antioxidant potential in bilateral cataract patients (Hashimoto et al. 2014), improved immune response and a reduction in inflammation and oxidative stress (Park et al. 2010), cognitive improvements with increased response time and accuracy of completing tasks (Katagiri et al. 2012; Satoh et al. 2009), cosmetic benefits through improvements in skin elasticity and a reduction in wrinkles (Tominaga et al. 2012), and improvements in semen quality with an associated increase in pregnancy rates (Elgarem et al. 2002; Comhaire et al. 2005). However, with regard to anticancer, cardiovascular health claims for the alleviation of oxidative stress in humans, and benefits in ocular health that have been reported in vitro and in vivo, no conclusive statements can be deduced, and further study is warranted.
The health benefits and published evidence suggest H. pluvialis astaxanthin is safe and orally bioavailable (Fassett and Coombes 2012), whilst having no provitamin A activity (which can lead to hypervitaminosis A) (Olaizola and Huntley 2003), and consequently, more clinical trials should be conducted (Fassett and Coombes 2012). The recommended dosage is 4–8 mg/day for normal health maintenance, and for athletes, 12 mg has been observed to be more effective (Capelli 2018). The FDA has approved H. pluvialis-derived astaxanthin for direct human consumption (up to 12 mg/day), and if taken for less than 30 days, 24 mg can be taken (Visioli and Artaria 2017). As the number of clinical studies and promotion of natural astaxanthin have increased, market demand has increased leading to a situation where demand is greater than the current ability to supply.
1.4 Sources of Astaxanthin
As outlined above, commercially, the main sources of astaxanthin are synthetically derived from natural astaxanthin X. dendrorhous and H. pluvialis. H. pluvialis is the best known natural producer of astaxanthin (Table 6.1). To date, only one higher plant species has produced astaxanthin (Adonis annua) with only 1% dry weight (DW) being observed in the petals (Renstrøm and Liaaen-Jensen 1981). However, despite the potential, the plant has relatively small flowers preventing suitable commercial production (Cunningham and Gantt 2011). H. pluvialis-derived astaxanthin is readily accepted for the human food market and of all living organisms has been reported to have the highest concentration of astaxanthin, with reports regularly revealing around 4% (Aflalo et al. 2007) and up to 7.7% at laboratory scale (Kang et al. 2005). The purity of astaxanthin as total carotenoids in H. pluvialis is much higher than other microalgae and can be up to 95% of the total carotenoids (Harker et al. 1996), with most reports revealing an average of 85% (Capelli et al. 2013a; Dore and Cysewski 2003). Comparatively, C. zofingiensis has been reported to contain approximately 50% astaxanthin of the total carotenoids, the other main carotenoids being canthaxanthin and adonixanthin (Liu et al. 2014b). Interestingly, in a direct comparison on the basis of astaxanthin esterification, C. zofingiensis was observed to contain a higher percentage of astaxanthin diesters (76.3% of the total astaxanthin), but with a significant reduction in astaxanthin monoesters (18% of total astaxanthin) compared with H. pluvialis (35.5% of total astaxanthin as diesters and 60.9% as monoesters) (Yuan et al. 2011). For other microalgae to become commercially competitive, the extracts would need to be purified, adding cost to the process. The commercial sector producing H. pluvialis-derived astaxanthin is well established, and there have been no adverse effects associated with the administration of H. pluvialis-derived astaxanthin reported to date (Fassett and Coombes 2012; Satoh et al. 2009; Spiller and Dewell 2003).
1.5 Large-Scale Production of Astaxanthin
The commercial scale-up of H. pluvialis has been and currently is difficult, with cultures requiring strict environmental conditions in the green stage (Olaizola and Huntley 2003). The first large-scale study investigating astaxanthin production from Haematococcus in a commercial facility (500,000 L bioreactor, 4500 m2) in California was in 1987 by Microbio Resources Inc. for the production of a powder (1% DW astaxanthin), marketed under the name Algaxan Red (Bubrick 1991). This dry algal powder was utilised for the aquaculture sector with a production cost of <US $20 kg−1 for the astaxanthin biomass product and able to compete with the synthetic form on price US $2000 kg−1 (Bubrick 1991). To date, there are several astaxanthin companies who have been unsuccessful including Fuji Chemicals with their BioDome™ system (Hawaii, USA), Aragreen (UK), and Maui Tropical Algae Farms (Hawaii, USA). Large-scale cultivation often results in low biomass densities susceptible to contamination issues, with high cell die-off (photobleaching) when transferred to the red stage under high light, and overall, it is a costly production process requiring extraction of the mechanically and chemically resistant, thick-walled aplanospores. Furthermore, there are issues with biofouling from cells forming in ‘dead areas’ with poor circulation which can lead to reduced light penetration and can cause considerable downtime, thus increasing the annual cost in large-scale cultivation. Companies such as Varicon Aqua Solutions Ltd. (PhycoFlow™) (http://www.variconaqua.com) along with individuals (Van De Ven and Van de Ven 2009) have patented technologies with automated self-cleaning systems, and these technologies offer the ability to reduce biofouling and downtime. The production capacity of H. pluvialis outdoors is further constrained by an intrinsically slow growth rate, low cell density, ease of contamination by other microorganisms, and susceptibility to adverse weather conditions (Ip et al. 2004). Most companies follow a production process similar to the schematic shown in Fig. 6.4.

Current process used on a commercial scale in the two-stage production process
Most companies have utilised H. pluvialis strains from culture collections which have been investigated in the literature such as CCAP 34/12, NIES-144, and SCCAP k-0084 (BCC Research 2015) that have been maintained in an artificial environment for a long period of time since their initial isolation from the natural environment. A few companies have used H. pluvialis strains, isolated directly from the natural environment that are unavailable in culture collections, including Cyanotech utilising H2B (Cifuentes et al. 2003) and MC Biotech utilising a local Brunei isolate. At a commercial level, astaxanthin is commonly produced using a two-stage culture system involving a green stage, for maximal biomass production, and a red stage, for maximising the product astaxanthin (Aflalo et al. 2007; Olaizola 2003; Olaizola and Huntley 2003), but a three-stage culture system (green, starvation, and red stage) is implemented by Algalif, Iceland. In the outdoor two-stage production process, astaxanthin productivities can reach 8–10 mg/L/day over a 10 day cycle (4 day green stage and 6 day red stage) with astaxanthin accounting for up to 4% DW under high light and nitrate-depleted conditions in the red stage (Aflalo et al. 2007). Alternatively, a continuous, one-stage production process has been demonstrated at pilot scale which produces astaxanthin from a mixed culture of motile macrozooids and palmelloids, resulting in almost twice the astaxanthin productivity (20.8 mg/L/day), formed under nitrate-deficient conditions (Del Río et al. 2008), but this process is yet to be adopted on a commercial scale.
Currently, the manufacture of astaxanthin is mainly conducted outdoors (Fig. 6.2), primarily due to the high light intensities and temperatures required for astaxanthin induction in the red stage which would be uneconomical indoors, but surprisingly most of the production occurs outside the tropics. In indoor cultivation, the red stage can account for 59% of the electricity costs, mainly due to the high light costs (Li et al. 2011). Only two companies employ a completely indoor production process to the author’s knowledge, Fuji Chemicals and Algalif. Fuji Chemicals abandoned H. pluvialis culture outdoors in its BioDome™ system in Hawaii due to contamination issues and subsequently continued with indoor mixotrophic culture in Sweden and Washington (Algae Industry Magazine 2015). Commercial production of H. pluvialis-derived astaxanthin in temperate zones is constrained due to unsuitable weather conditions for astaxanthin production outdoors, and consequently, only indoor culture is feasible. Aragreen in Gloucestershire was investigating astaxanthin production from H. pluvialis, but the company filed for bankruptcy in 2017 (Aragreen 2015).
Industrially, there are a wide range of cultivation methods, and most are aiming to utilise a more sustainable production process. In the case of Cyanotech, in the green stage H. pluvialis is cultivated indoors under strictly regulated culture conditions and then transferred outdoors for astaxanthin induction in the red stage in open ponds. AlgaTechnologies in Israel conducts their whole process outdoors in photobioreactors (PBRs) to exploit natural sunlight and utilises photovoltaic cells (Algae Industry Magazine 2015). Comparatively, Algalif utilises geothermal energy for an entire production process indoors using light-emitting diodes (LEDs). Most companies are focussing on phototrophic cultivation, but mixotrophic cultivation is being explored, for example, at Fuji Chemicals. Lorenz and Cysewski (2000) reported that astaxanthin induction can take 3–5 days, and during this stage, the encystment process results in the formation of aplanospores (cysts).
After cultivation and astaxanthin induction, the aplanospores are harvested by gravitational settlement and further concentrated by ultracentrifugation. The biomass is then dried, conventionally by spray drying as this is more economical than freeze drying and drum drying (Dore and Cysewski 2003; Shah et al. 2016). The dried thick-walled aplanospores are subjected to an extraction process to disrupt the cell walls and make the astaxanthin bioavailable. The walls of aplanospores resist digestion by animals (in feed applications) and by humans (nutraceutical applications), and therefore, the aplanospores must be disrupted for astaxanthin to become bioavailable (Olaizola and Huntley 2003). Sommer et al. (1991) observed that intact astaxanthin-rich aplanospores of H. pluvialis on ingestion do not result in pigmentation in salmonids. Care has to be taken in the extraction process to limit oxygen exposure and high temperatures which can damage the astaxanthin and result in losses in the process (Bustos-Garza et al. 2013). Extraction of astaxanthin on a commercial scale is most commonly by supercritical fluid extraction (SFE) with CO2 (ScCO2) (Shah et al. 2016). After extraction, the dried product is usually mixed with a preservative and shipped to feed manufacturers where it is incorporated into formulated feed (Olaizola and Huntley 2003). Alternatively, the astaxanthin is encapsulated and formulated for nutraceuticals which is discussed in Sect. 3.4.
2 Biology of H. pluvialis/H. lacustris
H. pluvialis Flotow belongs to the Chlorophyceae, order Volvocales, and family Haematococcaceae, and in the past, this species has been referred to as Haematococcus lacustris or Sphaerella lacustris (Shah et al. 2016). Currently, H. pluvialis and H. lacustris are synonymous and the correct terminology is H. lacustris; therefore, this taxonomy will be followed hereafter (Buchheim et al. 2013; Mazumdar et al. 2018; Nakada and Ota 2016).
It has been determined that Haematococcus is non-monophyletic with two distinct Haematococcus lineages by using nuclear-encoded small (18S) and large (26S) subunit rRNA combined with internal transcribed spacer 2 (ITS2) genes (Buchheim et al. 2013). It has been determined that H. pluvialis (H. lacustris) is the only member of the Haematococcus genus (albeit with at least five distinct lineages A–E from bootstrap data), with motile macrozooids with ‘delicate’ cytoplasmic strands and the formation of aplanospores with copious amounts of astaxanthin. Buchheim et al. (2013) stated the other Haematococcus species (H. buetschlii, H. capensis, H. zimbabwiensis, and H. droebakensis) should be designated to the second lineage, the Balticola genus (cytoplasmic strands thickened at the base) as previously proposed by Droop (1956). Allewaert et al. (2015) reported three species of Haematococcus from European isolates (H. pluvialis, H. rubens, and H. rubicundus) with H. pluvialis having the lowest maximum growth rate. Mazumdar et al. (2018) have reported four Haematococcus lineages with five valid species: H. lacustris, H. rubicundus, H. rubens, H. carocellus, and H. alpinus. The H. alpinus species was recently isolated from an alpine zone in New Zealand and identified as a new species with no known relatives (Mazumdar et al. 2018).
H. lacustris is regarded as the ‘birdbath’ alga which is distinct from other species of Haematococcus due to its ability to produce a vegetative resting stage (cyst/haematocyst/aplanospore) and is known to accumulate high amounts of the carotenoid astaxanthin (Buchheim et al. 2013; Droop 1955). H. lacustris differs morphologically from other species of Haematococcus, having uniformly thin cytoplasmic strands compared to strands which are thickened at the base (Buchheim et al. 2013) and possessing three or more pyrenoids compared to only two in other species of the Haematococcus genera (Allewaert et al. 2015). H. lacustris is primarily a freshwater species, commonly found in ephemeral rain pools, natural and manmade ponds, and birdbaths (Burchardt et al. 2006). H. lacustris is circumglobal and has been found on every continent with the exception of Antarctica (Guiry 2010). The ability of H. lacustris to encyst allows this species to survive in extreme conditions: high light, temperature, and salinity (Proctor 1957a).
2.1 Life Cycle
The first reports in the literature on the life cycle of H. lacustris were written in the middle of the nineteenth century (Flotow 1844) (Peebles 1909; Elliot 1934), and there was a further surge of interest in the late 1950s (Droop 1956; Proctor 1957a, b). H. lacustris has a complex life cycle, typically encompassing four life cycle stages: with the green stage containing vegetative cells, green motile macrozooids (flagellates), and non-motile palmelloids (zoospores), the red stage containing non-motile aplanospores (haematocysts), and a gamete stage with microzooids (Elliot 1934; Triki et al. 1997; Han et al. 2012). Triki et al. (1997) observed that microzooids were formed from aplanospores maintained in nitrate-starved conditions for over a month when transferred to a medium high in nutrients. Under favourable conditions, green motile macrozooids predominate (Triki et al. 1997), but when conditions become unfavourable, e.g. low nutrients, high light, and salinity stress, aplanospore formation occurs in conjunction with astaxanthin accumulation (Harker et al. 1996).
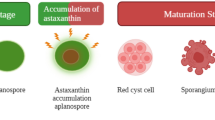
Wayama et al. (2013) reported that the life cycle of H. lacustris was more complex than originally perceived, describing the life cycle in more detail. It was reported that when H. lacustris aplanospores were resuspended in fresh medium, they germinated and released up to 32 green motile macrozooids by cytokinesis. After 3–5 days, these were noted to form green coccoid cells (palmelloids), and as aging progressed, the cells were transformed into intermediate cells (palmelloids) and aplanospores (Wayama et al. 2013). Even though a considerable body of work has been undertaken, the life cycle of H. lacustris is still not fully understood along with the morphotypes involved (Figs. 6.5 and 6.6). To date, little is known on what conditions contribute to palmelloid formation other than culture ageing and strain disposition (Allewaert et al. 2017). In addition, red motile macrozooids have also been observed in some strains, but it has not been fully elucidated why they are formed rather than aplanospores (Fig. 6.5) (Brinda et al. 2004; Butler et al. 2017; Del Río et al. 2005; Grünewald et al. 1997; Hagen et al. 2000; Tocquin et al. 2012).

Life cycle of H. lacustris: (a) green motile macrozooid, (b) early-stage palmelloids, (c) late-stage palmelloids, (d) aplanospore, (e) green motile microzooid, and (f) red motile macrozooid. The black lines indicate known interactions between life cycle stages, and the red lines indicate proposed interactions

Reproduction in H. lacustris is still a contentious issue as it is unknown whether H. lacustris undergoes sexual reproduction, and more direct evidence is still warranted (Chunhui et al. 2017). During division, sporocysts are formed which can contain 16 or 8 cells in the green and red stage, respectively (Figs. 6.7 and 6.8). H. lacustris is reported to be capable of sexual reproduction, but it is considered unusual (Triki et al. 1997). Triki et al. (1997) did not observe sexual reproduction in green motile macrozooids and reported that this was due to H. lacustris being heterothallic within culture collections, with populations derived from a single mating type (Droop 1956). Zheng et al. (2017) stated there is no convincing evidence that H. lacustris undergoes sexual reproduction, and it is unknown if Figs. 6.7d and 6.8b show the onset of sexual reproduction or whether the cells separate without fusing. For syngamy to occur, two comparable mating types have to be present (Triki et al. 1997). In the presence of a single clone, or numerous incompatible clones of H. lacustris, syngamy would not be possible (Triki et al. 1997). Determining if sexual reproduction occurs in H. lacustris would be of biotechnological interest because then mating trials could be conducted for selective breeding.

Asexual reproduction in green motile macrozooids: (a) 16 daughter cells, (b) four daughter cells, (c) released green motile macrozooids after the breakdown the of extracellular matrix, and (d) pairing head to head in green motile macrozooids

(a) Asexual reproduction with eight daughter cells formed and (b) pairing head to head in red motile macrozooids
During the life cycle of H. lacustris, ultrastructural changes occur within the cell. These have been well documented by Wayama et al. (2013) and reviewed by Shah et al. (2016). The green motile macrozooids are surrounded by an extracellular matrix (Wayama et al. 2013). During the onset of encystment, H. lacustris cell walls thicken up to 2 μm, and the cells develop conspicuous pyrenoids with many starch grains located around the pyrenoids (Wayama et al. 2013). Circular oil droplets with various sizes containing astaxanthin are located around the nucleus (Wayama et al. 2013). As astaxanthin accumulates, the chloroplast reduces in volume, and the chloroplasts degenerate and are localised in the interspace between oil droplets, but the photosynthetic activity of the cell is maintained (Wayama et al. 2013). It is still unknown what happens to the ultrastructure of the red motile macrozooids and if it is the same as in aplanospores.
2.2 Biochemical Components: Proteins, Lipid, Carbohydrates, Pigments
Morphogenesis from the green motile macrozooid to the aplanospore cell stage results in profound changes within the cell, including changes in cell wall structure which can be detected by electron microscopy and cytochemistry (Wayama et al. 2013). The green motile macrozooids exhibit an extracellular matrix (mainly consisting of glycoproteins and lacking cellulose or acetolysis-resistant material) around the cell (Hagen et al. 2002). In ageing green motile macrozooids, a two-layered primary cell wall forms (containing β-1,4-glycosidic linkages), which is subsequently followed by a loss of motility and the development of palmelloid cells (Hagen et al. 2002). After completion of the primary cell wall, the formation of a trilaminar sheath is observed containing cellulose in the palmelloids (Damiani et al. 2006). During encystment, the H. lacustris cell develops a secondary cell wall containing algaenan, a sporopollenin-like material, which is highly resistant to chemical and mechanical breakage (Han et al. 2013a). Montsant et al. (2001) identified that the cell wall of aplanospores was two- to threefold thicker than green motile macrozooids using transmission electron microscopy (TEM). The composition of the aplanospores cell walls is 70% carbohydrate (89.4% mannose), 6% protein, 3% cellulose, and 3% acetolysis-resistant material (Hagen et al. 2002). The biochemical composition of H. lacustris varies depending on the life cycle stage and the environmental conditions.
H. lacustris in the green stage typically has a biochemical composition of 13.8–48.0% protein (higher protein related to a higher nitrogen content) (Gacheva et al. 2015; Sipaúba-Tavares et al. 2015), 39.0–64.2% carbohydrate, and 8.3–16.2% lipid (as a function of DW) (Lorenz 1999; Gacheva et al. 2015). The primary FAs in the green stage were linolenic (18:3 (n − 3)) and palmitic acid (16:0) (26.4% and 18.9% of the total FAs, respectively) (T. Butler, unpublished data). With regard to the pigment fraction, the green vegetative cells can contain chlorophylls a and b, and the carotenoids lutein (70%), neoxanthin (12%), violaxanthin (10%), β-carotene (8%) with zeaxanthin are also reported (Harker and Tsavalos 1996; Harker et al. 1996).
In varying stages of the red phase, H. lacustris had a proximate composition (on a dry basis): 14–26% crude protein, 2.6–26.3% lipid, 6.30–48.8% carbohydrate, 2.0–4.0% ash, and an approximate gross energy of 24.1 kJ/g (Boussiba and Vonshak 1991; Choubert and Heinrich 1993; Sarada et al. 2006; Kim et al. 2015; Molino et al. 2018). It was reported that the lipid was composed of 88.3% FAs with 48.20% as polyunsaturated fatty acids (PUFAs) (the main fatty acids were linoleic acid (18:2 (n − 6)), palmitic, and oleic acid (18:1) encompassing 74.25%) (Kim et al. 2015; Molino et al. 2018). The amino acid profile is comprised of glutamic acid, aspartic acid, leucine, and alanine with a total amino acid content of 10% DW and 46% of the amino acids in H. lacustris defined as essential (Kim et al. 2015). The monosaccharides were mostly glucose and mannose (46% and 40.9% of the composition, respectively) (Kim et al. 2015). In a commercial product, Cyanotech previously reported that spray-dried aplanospore biomass had an astaxanthin content of >1.5% with the biomass containing 20–30% protein, 30–40% carbohydrate, 5–15% ash (dry weight (DW)), and 4–9% moisture, with a particle size of 5–25 μm (Dore and Cysewski 2003).
Generally, it has been observed in the red stage that the carbohydrate content increases dramatically with up to 74% as starch observed (Boussiba and Vonshak 1991). When aplanospores are formed, cytoplasmic lipid droplets (FAs as mono- or diesters) can account for 40% DW and can contain 4% astaxanthin (Aflalo et al. 2007; Saha et al. 2013a). The neutral lipid fraction predominates in the green and red stages, and in the transition to the red stage, the neutral lipid fraction as triacylglycerides (TAGs) increases along with the glycolipid content (Damiani et al. 2010). In the red phase, the aplanospores are rich in palmitic (C16:0), linoleic (18:2), and linolenic (18:3), whereas in the red motile macrozooid stage, oleic acid (18:1) was also found to be abundant (Butler et al. 2017). The biochemical composition largely depends on several factors including cultivation conditions, e.g. light, temperature, nutrients, carbon dioxide, and the genetics of the cell. Nutrient starvation is well known to increase lipid content, and the FAs formed are suitable for biodiesel (Damiani et al. 2010; Saha et al. 2013a). In the red stage, the pigment composition undergoes a dramatic shift with astaxanthin comprising 80–99% of the total carotenoids (Dragos et al. 2010). The ratio of carotenoids to chlorophyll is around 0.2 in the green stage but increases to 2–9 in the red stage (Shah et al. 2016). In the aplanospores and red motile macrozooids, astaxanthin is not in the free form but is often esterified with mono- or diesters of palmitic (16:0), linoleic (18:2), or oleic acid (18:2) (Shah et al. 2016; Butler et al. 2017). In aplanospores, approximately 70% of the astaxanthin is monoesters, 25% diesters, and only 5% free astaxanthin (Johnson and Schroeder 1995; Solovchenko and Chekanov 2014), but in red motile macrozooids, 77% of the astaxanthin occurs as a monoester, 18% as diesters, and 1.4% as free astaxanthin (Butler et al. 2017).
2.3 Bioprospecting for Commercially Relevant Strains
H. lacustris is a ubiquitous freshwater green alga with members of this species found circumglobally, and to date, >150 strains have been isolated, with the majority from the northern hemisphere (Fig. 6.9). Of these unique strains, the location of isolation of at least 44% remains unknown (Alam and Wang 2019).

Geographical mapping of 153 H. pluvialis/H. lacustris strains obtained from 18 culture collections and the published literature. From the 18 culture collections 65 unique strains were obtained. It was observed that for most of the strains the location of isolation was unknown (44.4%)
In the past, morphological traits have been used to determine the species and strains of H. lacustris. It has been identified that some culture collection strains predominantly grow as green motile macrozooids and others as predominantly green palmelloids (Han et al. 2012). However, morphology alone proves difficult to observe differences between strains, and genetics play an increasingly important role for differentiation, and therefore, a rapid DNA barcoding method is required for cataloguing new strains. Barcoding is important for identifying and cataloguing strains with desirable characteristics, including high growth rate and astaxanthin production and the capability to survive extremophilic conditions (an advantage in large-scale culture to avoid culture crashes from contaminants). Conventionally, microalgal cells are lysed using heating and cetyl trimethylammonium bromide (CTAB) (Doyle and Doyle 1990) to release DNA that is then amplified through the polymerase chain reaction (PCR), run on a gel, purified, and sequenced (Mostafa et al. 2011). As a pretreatment for DNA extraction, bead beating has been incorporated as it is notoriously difficult to extract high-yielding DNA (Peled et al. 2011). Recently, a simple colony PCR process has been established for H. lacustris with a simple heating step in a PCR machine for 10 min with PCR buffer, followed by subsequent PCR amplification (with improved band intensity with increasing cycle number) (Liu et al. 2014a).
To date, no universal barcode has been established for eukaryotic microorganisms, but the hypervariable V4 region of the 18S rDNA was proposed as a prebarcode (Łukomska-Kowalczyk et al. 2016). Mostafa et al. (2011) used inter simple sequence repeat (ISSR) and random-amplified polymorphic DNA (RAPD) molecular markers to observe the genetic diversity between ten strains of H. lacustris, four sourced from Iran and the other six from CCAP which successfully produced a dendrogram showing the correct strains based on their geographical origin. Allewaert et al. (2015) used ITS2 and then complete ITS rDNA and rbcL molecular phylogenies (with ITS being more powerful for species/strain separations) to determine the relationship between European H. lacustris isolates (seven strains) and those in common culture collections. It was determined that six lineages could be resolved from the ITS rDNA phylogenetic data corresponding to three out of five of the ITS2 rDNA lineages (A, C, and E) reported by Buchheim et al. (2013). Denaturing gradient gel electrophoresis (DGGE) was also identified as a method for rapid identification of European temperate strains and has been highlighted as a method that could be used in the future (Allewaert et al. 2015).
Bioprospecting for new strains has been fairly commonplace for H. lacustris as the large, red aplanospore cells are easy to see, and isolates have been obtained globally from Temperate Zones over Europe (Allewaert et al. 2015) to Torrid Zones such as India (Prabhakaran et al. 2014) to Frigid Zones such as Svalbard (Klochkova et al. 2013). Many strains of H. lacustris in culture collections including the Culture Collection of Algae and Protozoa (CCAP) have been found in ephemeral pools (CCAP 2015). Other strains of H. lacustris have been found in extreme environments outside of their conventional niche. An isolate has been found at high altitude in India (Prabhakaran et al. 2014). Recently, a cold-tolerant strain of H. lacustris was isolated from the high Arctic in Blomstrand Halvøya, Svalbard, which has been found to exhibit growth between 4 and 15 °C and can produce astaxanthin at 4–10 °C (Kim et al. 2011; Klochkova et al. 2013). H. lacustris has even been recorded in samples from a nuclear-fuel storage pond in Sellafield, UK (Groben 2007). A thermophilic strain with the ability to grow at temperatures up to 41.5 °C has also been isolated (Gacheva et al. 2015). In one study that screened 30 natural isolates and compared them with culture collection strains, it was identified that the culture collection strains had a lower astaxanthin productivity which might have been attributed to a loss in photoprotective capacity during longer-term cultivation (Allewaert et al. 2017). Bioprospecting offers significant potential for identifying new strains of H. lacustris with desirable characteristics for biotechnological exploitation. Elucidating the diversity in H. lacustris species is essential for biotechnological applications as potential fast-growing and astaxanthin-hyperaccumulating strains can be identified, in conjunction with determining strains suitable to local climatic conditions.
An Arctic strain of biotechnological significance is BM1, found on coastal rocks off the coast of a Russian island, with the ability to tolerate salinities up to 25% (Chekanov et al. 2014). Typically, a salinity of 8% causes the cessation of growth in H. lacustris (Boussiba and Vonshak 1991). This strain could be cultivated in brackish water which would reduce the cost of production, would minimise the environmental burden, and would be suitable for areas with a limited supply of freshwater. In addition, astaxanthin accumulation was detected after only 10 days of cultivation (Chekanov et al. 2014). After 6 days of resuspension in distilled water, 27 °C, 480 μmol photons/m2/s, and continuous light, the astaxanthin content reached 5–5.5% DW (99% of the total carotenoids).
Much of the scientific literature has been based on specific strains of H. lacustris from culture collections, including CCAP 34/7 (Harker and Tsavalos 1996; Mendes-Pinto et al. 2001; Mostafa et al. 2011; Rioboo et al. 2011), CCAP 34/8 (García-Malea et al. 2005, 2006, 2009), SCCAP k-0084 (Montsant et al. 2001; Peled et al. 2012; Wayama et al. 2013), and the highest number of publications on NIES-144 (Kobayashi et al. 1991, 1993, 1997a, b; Kang et al. 2005; Yoo et al. 2012; Wan et al. 2014a). There are few reports on other strains held in culture collections including CCAP strains 34/1D, 34/13, and 34/14. Published papers for these strains have mainly been restricted to the study of evolutionary relationships (Mostafa et al. 2011). Revisiting these strains could provide a quick method for identifying strains with suitable properties for commercial production of astaxanthin which could alleviate some of the issues currently found within the industry such as identifying red motile macrozooids with the highest reported astaxanthin content to date (2.74% DW) (Butler et al. 2017). To date, it has been observed that the highest astaxanthin content has been observed in NIES-144 and the highest astaxanthin productivity in CCAP 34/8.
To date, only two publications have comprehensively compared H. lacustris strains. From 25 strains from various culture collections, it was identified that CCAC 0125 was the optimal strain with a total biomass and astaxanthin yield of 91.2 g/m2 and 1.4 g/m2, respectively, with an associated astaxanthin content of 1.5% DW (Kiperstok et al. 2017). All 25 strains were capable of immobilised growth in a biofilm attaining biomass and astaxanthin yields of between 73 and 112 g/m2, 0.74 g/m2 and 2.1 g/m2, respectively. Allewaert et al. (2017) undertook a screen of 30 strains which included recently isolated strains and those maintained in culture collections. It was concluded that these recently isolated strains generally had a higher astaxanthin productivity and there was a 15-fold difference, with BE02_09 having the highest astaxanthin productivity (4.59 mg/L/day). In future strain selections it would be beneficial to conduct a high-throughput screening method to identify astaxanthin-hyperproducing mutants. Fourier-transform infrared spectroscopy has been suggested rather than conventional high-performance liquid chromatography (HPLC) (Liu and Huang 2016).
2.4 PBR Development and Cultivation Mode
To date, H. lacustris has been cultivated in open ponds, plastic bags, fermentors, and PBRs (flat plate, horizontal/tubular, bubble columns, and airlift) and attached to a membrane system (Table 6.2). The highest astaxanthin productivity (20.8 mg/L/day) was obtained in an indoor system using a one-stage process (Del Río et al. 2008). The production process can range from 10 to 90 days, with biomass productivities in the green and red stages ranging from 0.04 to 1.58 g/L/day and 0.02–1.90 g/L/day, respectively, with astaxanthin productivities from 0.12 to 20.9 mg/L/day, and with an astaxanthin content of 0.20–7.72% DW (Table 6.3). It has previously been reported that tubular airlift systems are preferable over bubble column PBRs for the outdoor production of biomass and astaxanthin productivity due to an optimal average lighting irradiance of 130 μmol photons/m2/s with nitrate decreasing to <5 mM for astaxanthin induction over the 16-day period (Table 6.3) (García-Malea López et al. 2006). In a direct comparison between a bubble column and an airlift PBR, the airlift PBR resulted in a 18% higher biomass concentration (4.8 g/L DW, 7 × 106 cells/mL) and a 16% higher astaxanthin yield (480 mg/L), attributable to the regular light/dark cycles and laminar flow in the downcomer of the airlift PBR (Ranjbar et al. 2008). In terms of the optimal light path, it has been observed that a 6 cm light path in the green stage, followed by a 3 cm light path in the red stage resulted in the highest astaxanthin productivity (20.1 mg/L/day) (Wang et al. 2019). In terms of the optimal bioreactor size, it has been determined that the smaller flat plate bioreactor system (17 L) resulted in a 97% higher cell density than in a 200 L system (Issarapayup et al. 2011) showcasing the difficulties of scaling up. Maximising hydrodynamic mixing through an optimised sparger and PBR shape is also critical (metal, 0.2 vvm, 1.3 cm diameter sparger, 60° V-shaped bottom) with a resultant 1.7-fold increase in astaxanthin productivity without the adherence of cells (Yoo et al. 2012). However, with 60+ years of PBR research (317 studies of algal reactors), it has been identified that there is little difference between the system used and the biomass productivity overall, but it was suggested that intermediate volume bioreactors with higher surface area-to-volume ratios could provide higher yields whilst simultaneously reducing the environmental footprint with lower energy consumption (Granata 2017).
Recently, there has been a large transition in the materials utilised for PBRs; traditionally, plastic tubes have been utilised due to their apparent low cost. Recently, many microalgal companies have partnered with Schott AG (e.g. A4F, Varicon Aqua Solutions Ltd., Ecoduna, and Heliae) and are replacing plastic PBRs with Schott glass due to higher resistance to UV radiation and chemicals, reductions in biofilm formation, and cost savings over a longer-term period (Schott 2019). It has been identified that over a 12-month period using a total tube length of 12 km, a 10% higher biomass and astaxanthin productivity was attained using a wall thickness of 1.8 mm compared with 2.5 mm in Israel, attributable to higher sunlight penetration and more stable temperatures (Schott 2019).
Novel modes of cultivation have included attached cultivation on a membrane and utilising perfusion culture. In attached cultivation, the cells are cultivated in the water column and then seeded on a membrane to increase the light surface area, and this reduces the harvesting costs as the cells are already dewatered, resulting in up to a 90% reduction in water consumption (Zhang et al. 2014). Other benefits include overall energy savings from the lack of mixing/pumping and a reduction in contamination, particularly single-celled protozoa (Wan et al. 2014a). Furthermore, when attached cultivation is employed in the red stage, the astaxanthin induction is faster than in column PBRs (Wan et al. 2014a). Utilising a two-stage system with attached cultivation for the red stage, biomass and astaxanthin productivities of 3.7 g/m2/day and 65.8 mg/m2/day, respectively, were obtained utilising strain NIES-144 (12 day) (2.8- and 2.4-fold higher than conventional suspended bioreactor cultivation, respectively) (Wan et al. 2014a). The optimal temperatures for maximising biomass and astaxanthin production were conflicting, with the highest astaxanthin content (1.5% DW) at 15 °C but the maximum biomass and astaxanthin productivities at 25 °C (Wan et al. 2014a). Zhang et al. (2014) reported a slightly modified version of attached cultivation utilising strain SAG 34-1b with a different medium (BG-11 vs. NIES-N) with nitrate deprivation (1.8 mM) rather than depletion, resulting in an astaxanthin productivity of 164.5 mg/m2/day. Through a strain screening process (25 strains) and utilising the optimal strain (CCAC 0125) for a twin-layer two-stage immobilisation system, a high biomass (19.4 g/m2/day at 1015 μmol photons/m2/s) and astaxanthin productivity (0.39 g/m2/day at 1015 μmol photons/m2/s) was obtained with 1% CO2 supplementation, 14/10 photoperiod, and 28.5 °C (Kiperstok et al. 2017). Utilising a one-stage process resulted in similar biomass and astaxanthin productivities but in half the time (8 days) (Kiperstok et al. 2017).
Alternatively, perfusion culture in a fermentor has been demonstrated where the medium (NIES-C with 11.98 mM acetate, 2.58 mM nitrate, and 0.147 mM phosphate) is continuously replaced, removing inhibitory metabolites formed during cultivation and replenishing nutrients (Park et al. 2014). This process resulted in a biomass and astaxanthin yield of 12.3 g/L (0.18 g/L/day) and 602 mg/L, respectively, through stepwise increased light irradiance (150–450 μmol photons/m2/s) (Park et al. 2014). However, this method required 54% more energy than a fed-batch stepwise photoautotrophic process and 24.5 additional days (Kang et al. 2010).
To date, most systems have focussed on phototrophic production using a two-stage strategy as the conditions for maximising growth and astaxanthin productivity are mutually exclusive (Table 6.2). The aim of the green stage is to maximise biomass productivity and the second red stage is to induce astaxanthin formation. Utilising this method, a biomass yield of almost 20 g/L could be attained after 21 days in the green stage using stepwise increases in irradiance (25–100–500 μmol photons/m2/s) with an astaxanthin productivity of 11.5 mg/L/day (Aflalo et al. 2007). Wang et al. (2019) have obtained a biomass yield of 20.1 g/L (1.34 g/L/day) in the green stage and 27.3 g/L DW (0.91 g/L/day) after the red stage, the highest biomass yield to date in a H. lacustris photoautotrophic system. A single-stage astaxanthin production system has been devised (utilising an impinging irradiance of 1000 μmol photons/m2/s, a specific average irradiance of 93.4 μmol photons/m2/s, dilution rate of 0.9 μ/day, and 2.2 mM nitrate) which resulted in a biomass productivity after the red stage of 1.9 g/L/day and an astaxanthin productivity of 20.8 mg/L/day, the highest to date (Del Río et al. 2008). The technical feasibility of this process outdoors in summer (50 L tubular PBR) has been showcased with a biomass and astaxanthin productivity of 0.7 g/L/day and 8 mg/L/day, respectively, and further increases were believed to be attainable through increasing the availability of light (>53.45 μmol photons/m2/s) to the cells (García-Malea et al. 2009). Furthermore, it has been proposed that night-time losses of biomass and astaxanthin could be reduced through identifying the optimised control temperature (Wan et al. 2014a). It was observed that 2.9 and 5-fold increases in biomass and astaxanthin productivities could be obtained with NIES-144 when the night temperature was maintained below 28 °C, but this will differ for each strain and the specific cultivation conditions (Wan et al. 2014a). However, it must be noted that several drawbacks of this system have been highlighted including the lower astaxanthin content compared to the two-stage process (0.9–1.1% vs. 3.8% DW), the requirement for artificial illumination at night which is economically unattractive, vulnerability to grazers, and difficulties in harvesting the heterogeneous cells (compared to gravitational settlement for the aplanospores) (Aflalo et al. 2007).
H. lacustris is capable of mixotrophic growth with sodium acetate and ribose appearing to be the most suitable substrates (Kobayashi et al. 1991; Pang and Chen 2017). It is well known that acetate is a suitable organic carbon source for maximum growth (4–12 mM) and for enhancing astaxanthin accumulation in the red stage (Cifuentes et al. 2003; Göksan et al. 2010; Gong and Chen 1997; Kang et al. 2007). Higher biomass and astaxanthin productivities (0.35 g/L/day and 4.54 mg/L/day, respectively) have been obtained when utilising NSII medium supplemented with 50 mM sodium acetate throughout the whole process, but with the addition of 100 mM potassium acetate in the red stage, a higher astaxanthin productivity could be attained (10.21 mg/L/day) with a 2.5–4.3% astaxanthin content, albeit with a reduced biomass productivity (0.24 g/L/day) (Pan-utai 2017). Using a fed-batch mode with sodium acetate results in a biomass yield of 1.77 g/L after 9 days (93.9% higher than a batch culture), and it was determined that the culture should be fed at night (Sun et al. 2015). Utilising ribose (9.66 mM) as a C5 carbon substrate has been suggested to prolong the green stage, increase the specific growth rate and biomass yields, and reduce the risk of contamination (Pang and Chen 2017). It has been stated that organic carbon feeding should be done at night under 16 °C, to minimise the loss of enzymatic activity (Sun et al. 2015).
H. lacustris has also been reported to be capable of heterotrophic growth using sodium acetate as a carbon source in the green stage, but the organism has slow metabolic growth (0.20–0.22 μ/day), and contamination issues have been encountered (Droop 1955; Hata et al. 2001). Comparatively, photoautotrophic production in the green stage results in a growth rate of 0.56 μ/day (García-Malea et al. 2005) and even up to 1.30 μ/day (Boussiba and Vonshak 1991). Heterotrophic growth (10–30 mM sodium acetate) offers the potential for producing high biomass yields in the green stage with Hata et al. (2001) obtaining a biomass yield of 7 g/L DW and Wan et al. (2015) attaining a biomass yield of 26 g/L DW with the highest biomass productivity (1.58 g/L/day) to date in the green stage (Table 6.2). Hata et al. (2001) revealed that after the third repeated fed batch, culture contamination became an issue (Hata et al. 2001), but Wan et al. (2015) did not report contamination issues. To date, there have been no confirmed reports of astaxanthin production in a commercial heterotrophic process; however, Kobayashi et al. (1997a, b) reported carotenoid formation in heterotrophic cultures of H. lacustris in the laboratory, but this was not confirmed to be astaxanthin, and to date, this remains unknown. A sequential heterotrophic-photoautotrophic production process has been suggested where astaxanthin formation was induced using nitrate deprivation, and subsequently, 5% CO2 was supplied to the culture resulting in the highest cellular astaxanthin content to date of 7.72% DW (6.25 mg/L/day) (Kang et al. 2005). Commercially, some companies have been experimenting with heterotrophic cultivation of H. lacustris, but astaxanthin production in the red stage is currently insufficient for commercialisation. Utilising the alga’s heterotrophic ability to produce high biomass yields in the green stage, followed by optimal induction in the red stage through photoautotrophic induction, could be a suitable method for attaining high astaxanthin productivities.
2.5 Astaxanthin Induction
Astaxanthin is accumulated outside the plastid in cytoplasmic lipid vesicles (Grunewald et al. 2001). Astaxanthin has been reported to have several functions within H. lacustris aplanospores, acting as a ‘sunshade’ protecting the photosynthetic apparatus, protection from photooxidative stress, and minimising oxidation of storage lipids (Han et al. 2012). To date, it is not fully known how astaxanthin acts to protect H. lacustris, and further studies need to be conducted to fully elucidate its role in the protection of H. lacustris cells in unfavourable conditions.
Astaxanthin synthesis was originally proposed to be induced by the cessation of cell division and only occur in the resting stage (Boussiba and Vonshak 1991; Kobayashi et al. 1997a, b). However, synthesis of astaxanthin has been demonstrated to be independent of cell division and can occur in vegetative cells (motile macrozooids and palmelloids) (Brinda et al. 2004; Butler et al. 2017; Del Río et al. 2005, 2008; Hagen et al. 2001). A range of factors have been investigated to explore astaxanthin synthesis by H. lacustris, including high light, high temperature, and nutrient deprivation/depletion (e.g. nitrate and phosphate). High light was suggested to have been one of the most effective factors in the stimulation of astaxanthin synthesis (Choi and Park 2002). High-temperature treatments have been reported to have resulted in greater levels of astaxanthin, but temperatures >30 °C have been found to reduce the biomass yield (Tjahjono et al. 1994). Salinity stress by the addition of NaCl (0.1–0.5%) has also been used to increase astaxanthin levels, but concentrations of 0.6–0.8% NaCl can cause severe cell mortality (Cifuentes et al. 2003; Harker et al. 1996; Sarada et al. 2002).
It has been proposed and is well known that nitrate limitation is the critical factor inducing astaxanthin accumulation with high light and dilution rate as factors responsible for enhancing the astaxanthin content, but which alone are not inducers of the pigment itself (nitrate > dilution rate > light) (García-Malea et al. 2009). Christian et al. (2018) further validated the effect of light and identified that high light intensity alone had little effect on inducing astaxanthin production. It has been identified that nitrate limitation (<5–8 mM) results in the formation of astaxanthin with 2.2 mM nitrate being optimal for astaxanthin productivities (Ranjbar et al. 2008; Del Río et al. 2008). In the two-stage process under nitrate deprivation (4 mM), the astaxanthin content is 2.7% DW, but with nitrate depletion, the content is increased to 3.8% DW (Wang et al. 2013). In the case of urea as a nitrogen source, 3 mM resulted in the highest astaxanthin content (2.4% DW) (Wang et al. 2019). Other reports have stated that a nitrate concentration of 0.6 mM is the concentration necessary to induce astaxanthin accumulation, whilst avoiding culture washout (García-Malea et al. 2009). The specific concentration of nitrate required for induction is likely dependent on the PBR, cultivation conditions, and the strain employed.
2.6 Astaxanthin Biosynthesis
Astaxanthin is produced in the chloroplast, accumulates around the nucleus to protect the ultrastructures from reactive oxygen species (ROS), is esterified in the endoplasmic reticulum, and spreads into cytoplasmic lipids, with recent models revealing that astaxanthin and lipid biosynthesis are not synchronous, with lipid droplets accumulating faster than astaxanthin (Collins et al. 2011; Saha et al. 2013b; Solovchenko et al. 2013; Cheng et al. 2017a). It has been found that photorespiration can accelerate astaxanthin accumulation which is speculated to be through increasing glycerate-3-phosphate (PGA), a precursor to glyceraldehyde 3-phosphate (G3P) (Fig. 6.10) during the Calvin cycle (Zhang et al. 2016). It has been identified that isopentenyl pyrophosphate (IPP) is an essential intermediate of carotenoid synthesis and this molecule can originate from the mevalonate pathway (MVA) or non-mevalonate pathway (MEP) in the chloroplast (Lemoine and Schoefs 2010). In the MEP pathway, 1-deoxy-d-xylulose-5-phosphate is formed in the first stage. IPP then undergoes isomerisation to dimethylallyl diphosphate (DMAPP); however, it remains unknown which enzyme is responsible for this conversion (Shah et al. 2016). The isoprenoid chain is then elongated, initiated with DMAPP with a subsequent linear addition of three molecules of IPP, and is catalysed by geranylgeranyl pyrophosphate synthase (GGPS); then finally, geranylgeranyl pyrophosphate (GGPP) is formed, a C20 compound (Shah et al. 2016).

Astaxanthin biosynthesis in H. lacustris. (Modified from Shah et al. 2016). The enzyme abbreviations are defined as follows: HDR, 4-hydroxy-3-methylbut-2-enyl diphosphate reductase; IPI, isopentenyl pyrophosphate isomerase; PSY, phytoene synthase; GGPS, geranylgeranyl pyrophosphate synthase; PDS, phytoene desaturase; ZDS, ζ-carotene desaturase; LCYE, lycopene ε-cyclase; LCYB, lycopene β-cyclase; CrtR-b, β-carotene 3,3′-hydroxylase; BKT, β-carotene ketolase
For the carotenoid synthesis, phytoene synthase (PSY) is the catalyst and initiates a head-to-tail condensation of two GGPP molecules to form the C40 compound, phytoene which acts as a precursor to astaxanthin (Cunningham and Gantt 2011). It is well known that PSY is upregulated in the transition from green to red stage cultures (Gwak et al. 2014). It has recently been proposed that a mutation in the PSY of H. pluvialis had an essential role in the evolution of hypercarotenogenesis (Pick et al. 2019). Lycopene is formed through four desaturation steps (increasing the number of conjugated carbon-carbon double bonds) involving two phytoene desaturases (PDS) and a ζ-carotene desaturase (ZDS) as the catalysts (Li et al. 2010; Nawrocki et al. 2015). Plastid terminal oxidases (PTOX1 and PTOX2) are cofactors involved in carotenoid desaturation, and PTOX1 is coregulated with astaxanthin synthesis, but the function is still undefined (Wang et al. 2009). Following the desaturation stages, the two termini of lycopene undertake cyclisation through competing pathways regulating metabolic flux; primary carotenoid formation is catalysed by lycopene ε-cyclase (LCYE), and secondary carotenoids are synthesised by lycopene β-cyclase (LCYB) resulting in β-carotene formation (Gwak et al. 2014; Lao et al. 2017). It has been found that β-carotene is exported out of the chloroplast and is enzymatically converted to astaxanthin in the cytoplasm (Fig. 6.10) (Pick et al. 2019). The cyclisation of lycopene is an important regulatory branch in astaxanthin biosynthesis. Upregulation of LCYB at the transcriptional, proteomic, and metabolomic levels could result in elevated concentrations of astaxanthin. It has also been identified that the two final oxidation steps catalysed by β-carotene ketolase (BKT) and β-carotene hydroxylase (CrtR-b) are rate-limiting steps of astaxanthin synthesis (Shah et al. 2016).
Recent transcriptomic profiling on the effect of LEDs on the astaxanthin biosynthetic pathway has been conducted, and it has been revealed that blue LED irradiation results in an increased expression of the enzymes BKT and the carotenoid hydroxylase (CHY) compared to white light as the baseline, but comparatively, red LED irradiation results in downregulation (Lee et al. 2018). Upregulation of PSY, LCY, carotenoid ketolase (Crt-o), and CrtR-b was also observed when blue light was used for astaxanthin induction (Ma et al. 2018). The astaxanthin synthesis pathway is complex, and multiple regulatory mechanisms at the transcriptional, translational, and posttranslational level are involved with five key enzymes critical in the process: isopentenyl pyrophosphate isomerase (IPI), PSY, PDS, Crt-o, and CrtR-b (Li et al. 2010). It is essential for the pathway of astaxanthin to be understood before genetic engineering can be undertaken (Fig. 6.10).
2.7 Genetic Engineering
Genetic engineering of microalgae has been reported in over 30 species, but the toolbox available for H. lacustris is limited. To date, the chloroplast genome (1.35 Mb) has been sequenced by the Synthetic Genomics group for H. lacustris UTEX 2505 (close relative of UTEX 16, a descendent of NIES-2264), and it has been identified as the largest assembled chloroplast of any plant or alga to date, but more coverage is required for the nuclear genome to be sequenced (Bauman et al. 2018; Buchheim et al. 2013; Smith 2018). It was reported that the sequencing of the chloroplast genome leaves many unanswered questions; >90% of the DNA was non-coding, it has a non-standard genetic code, it only encodes 12 tRNAs (less than half of a typical plastome), and it is one of the few sequenced plastids that is not biased in adenine and thymine (Smith 2018).
Currently, most genetic improvements in H. lacustris have been limited to classic random mutagenesis due to the lack of an available nuclear genome and a poorly annotated chloroplast genome. To date, UV mutagenesis (Sun et al. 2008; Tripathi et al. 2001) and chemical mutagenesis using N-methyl-N-nitro-N-nitrosoguanidine (MNNG) (Hu et al. 2008) and ethyl methanesulphonate (EMS) (Sun et al. 2008; Tripathi et al. 2001) have been trialled for elevating astaxanthin content with the aim of inducing 85–95% mortality. Chemical mutagenesis has been more successful because of the ability of H. lacustris to tolerate light damage, and using MNNG has resulted in improvements of volumetric astaxanthin up to threefold (Hu et al. 2008). For screening these mutants, herbicides have typically been used such as nicotine and norflurazon (Shah et al. 2016). With astaxanthin mutants, the colonies are potentially easier to pick because of their brighter red colouration. A Chilean H. lacustris mutant (mutated with EMS) was cultivated in a commercial-sized open pond of 125,000 L, and the astaxanthin content was 30% greater than the wild-type strain on a DW basis and 72% greater on a per culture volume basis (Gómez et al. 2013). Irradiating FACHB-872 with 4000 Gy 60Co-γ and then cultivation under high light with 15% CO2 resulted in a 1.7-fold increase in astaxanthin compared with a wild-type strain, and importantly, 56% of the genes were significantly upregulated in the mutant cells including pyruvate kinase providing a feedstock for astaxanthin and phytoene synthase, lycopene beta-cyclase, and ZDS for β-carotene conversion to astaxanthin (Cheng et al. 2016, 2017a, b).
An emphasis has been on targeting rate-limiting steps in astaxanthin biosynthesis with the key enzymes being localized in the chloroplast, and it has been observed that PDS is a key target (Grünewald et al. 2001). A focus has been on nuclear transformations with conventional mutagenesis such as overexpression of a PDS with a point mutation for norflurazon resistance with transgenic lines possessing a 36% higher astaxanthin content after 2 days of high light induction (Steinbrenner and Sandmann 2006). Recently, the carotenoid biosynthesis pathway has been genetically modified through a plasmid transformation in the chloroplast with the endogenous PDS nuclear gene to overproduce astaxanthin (67% increase in astaxanthin content in transformed strains compared to the wild type) with induction under high light and nitrogen depletion without an adverse effect on growth or biomass productivity (Galarza et al. 2018).
Insertional mutagenesis has been investigated for producing high-yielding astaxanthin strains using Agrobacterium-mediated transformation (Kathiresan et al. 2009), biolistics (particle bombardment) (Steinbrenner and Sandmann 2006), and electroporation (Sharon-Gojman et al. 2015), but until recently they have lacked efficacy. Now stable transformation of the chloroplast and nuclear genome is possible with the introduction of two transgenes without the requirement of an additional antibiotic resistance gene (Gutiérrez et al. 2012; Sharon-Gojman et al. 2015). The PDS variant is used as a selection marker that confers resistance to the herbicide norflurazon with a single point mutation (L504A) (Shah et al. 2016). Enhancement of astaxanthin in H. lacustris could be achieved by upregulating the PSY and CrtR-b genes which are often defined as being rate limited for astaxanthin production and have previously been upregulated by high light (Han et al. 2013b; Li et al. 2008, 2010). To bring a disruptive change in astaxanthin yields from H. lacustris, advanced genetic engineering methods with a greater emphasis on targeted mutations are required, utilising homologous recombination and clustering regularly interspaced short palindromic repeat (CRISPR) technology, which is an important new platform for generating RNA-guided nucleases (RGNs), such as Cas9 which is an RNA-guided DNA nuclease employed to introduce targeted mutations into eukaryotic genomes (Brodie et al. 2017). This may become a reality once the H. lacustris nuclear genome has been sequenced with detailed annotations and more transformation tools are available. Future emphasis should be on developing a toolbox for genetically engineering H. lacustris targeting the astaxanthin biosynthetic pathway, potentially through upregulation of the rate-limiting enzymes BKT and CrtR-b (Shah et al. 2016).
Recently, a high-throughput method has been devised for screening and selecting hyperproducing mutants under UV mutagenesis (40 mJ/cm2 with 32 min of UV exposure time) using 50 μm sodium azide to accelerate astaxanthin induction (Eui et al. 2018). Using a soybean oil-based extraction method combined with spectrophotometric analysis (OD 470 nm) enabled the detection of 31 strains (88.5% of the cells) with higher astaxanthin production than the wild type (NIES-144) with the M13 strain exhibiting an astaxanthin yield 1.59 times higher than the wild type (174.7 mg/L) (Eui et al. 2018). Utilising this high-throughput method can help identify transformants, and this information can be supported through transcriptomics, proteomics, and metabolomics to further target bottlenecks in the production of astaxanthin. If a genetically engineered strain is to be utilised for commercial production, caution is warranted because there are regulatory hurdles to overcome such as Directive 2001/18/EC in the European Union (EU) where the approval procedure can take 4–6 years and cost 7–15 million Euros (Hartung et al. 2014).
3 Commercial Constraints of Astaxanthin Production from H. lacustris
Commercial astaxanthin production has been successful, and a number of companies are successfully operating. However, the market is saturating and the price of astaxanthin is overinflated. As supply has increased, the price of astaxanthin has fallen. To date, the focal point of astaxanthin has been as a nutraceutical and functional food. However, the bulk of astaxanthin is used for aquaculture which is dominated by the synthetic form and by X. dendrorhous. In order to compete with these sources, the cost of production needs to decrease, and there is a requirement for challenges to be overcome in the production process, including improvements in biomass and astaxanthin productivity, mitigating contamination, and the requirement for green chemistry and engineering. There needs to be more collaboration between academia and industry to advance knowledge. More investigations are required to look into the commercial feasibility of astaxanthin from H. lacustris. Shah et al. (2016) identified that there are three key areas to target for further improvements: cultivation efficiency and cost, good cultivation practice with the control of predators, and extraction and purification of astaxanthin. In this section, the authors believe improving biomass/astaxanthin productivity, minimising cell die-off through photobleaching in the red stage, costly downstream processing methods, contamination, and the total cost of the process are critical problems to address.
3.1 Improved Biomass Productivity
Currently, the growth rate of H. lacustris is slow, and during cultivation as it is a flagellated form, it is vulnerable to shearing, attributable to hydrodynamic stress in the PBRs. There are difficulties in maintaining green motile macrozooids in the green stage without transitions to the palmelloid form which results in slower growing cells.
The optimal cultivation conditions have been determined from a range of studies. The optimal temperature for growth is strain dependent but is typically between 20 and 28 °C (Allewaert et al. 2015; Giannelli et al. 2015). Temperatures greater than 30 °C induce encystment and the formation of aplanospores within 2 days, resulting in the cessation of growth (Allewaert et al. 2015; Tjahjono et al. 1994). The optimal pH for H. lacustris has been reported in the range of 7.00–7.85 (Hata et al. 2001; Sarada et al. 2002). Typically, the optimal irradiance for H. lacustris is 70–177 μmol photons/m2/s with a saturation index of 250 μmol photons/m2/s (Zhang et al. 2014; Giannelli et al. 2015), but this has not been conclusive, and often lower irradiances have been investigated (Park et al. 2014). It is likely that other factors play a role in a multifactorial process. The impact of photoperiod has been preliminarily investigated with continuous illumination appearing to be optimal for higher-density cultures, but only a 12:12 and 24:0 photoperiod was compared (Domínguez-Bocanegra et al. 2007). The optimal light seems to be warm-white light, but this needs further investigation (Saha et al. 2013a). As an inorganic carbon source, CO2 is commonly utilised to increase biomass productivity and ranges from 1% to 5% in the green stage (Kaewpintong et al. 2007). It has been determined that CO2 supplied at 5% is beneficial for growth (3.3 g/L DW) but increases to 10% and results in a deterioration in growth, photosynthesis, and the assimilation of carbon, determined by PSII yield, NPQ activity, chlorophyll a content, and biomass yield (Chekanov et al. 2017). It has been observed that a γ-ray-irradiated mutant cultivated under CO2 at 6% in conjunction with high light (108 μmol photons/m2/s) had the highest biomass productivity (0.16 g/L/day) with a maximum growth rate of 0.6 μ/day (Cheng et al. 2016). Higher concentrations of CO2 at 10% or 20% resulted in a decrease in growth (Chekanov et al. 2017). A further emphasis needs to be on carbon uptake and assimilation by the cells rather than the CO2 input alone which has seldom been explored.
Biomass accumulation has been a major bottleneck in the two-stage process of astaxanthin production, and further optimisation of the growth media is required. It is clear that a wide range of media and their derivatives have been utilised for optimal H. lacustris cultivation (Table 6.3). Commonly, BBM or 3N-BBM has been used for H. lacustris culture (Oncel et al. 2011; Qinglin et al. 2007; Suh et al. 2006; Tocquin et al. 2012), along with BG-11 (Aflalo et al. 2007; Kiperstok et al. 2017; Torzillo et al. 2003; Zhang et al. 2009). It has been identified that BBM is more effective than 3N-BBM and BG-11 for attaining higher cell densities (Nahidian et al. 2018). Fábregas et al. (2000, 2001) utilised an optimised Haematococcus medium (OHM) with a final cell density of 3.77 × 105 cells/mL (3 times higher than BBM) with higher yields (1.62 g/L DW) attained by utilising high light (235 μmol photons/m2/s). Through media optimisation, BBM with threefold the phosphate concentration (5.16 mM) and an N/P ratio of 1.71:1 resulted in an 86% increase in the growth rate, and supplementation with 0.185 mM boron or 0.046 mM iron appeared beneficial for growth (Nahidian et al. 2018). However, a conflicting study by Fábregas et al. (2000) identified that boron was a nonessential nutrient for the growth of H. lacustris along with iodine, zinc, and vanadium. The addition of vitamin B12 has also been reported to increase algal growth by up to 55% in comparison to media without (Kaewpintong et al. 2007).
Utilising commercially viable media has also been investigated. A commercial plant fertiliser (£0.24 ton−1) with an N-P-K 20:20:20 resulted in 0.9 g/L DW after 12 days (Dalay et al. 2007). A common hydroponics fertiliser, FM:FB, was screened, and the optimised formulation resulted in a cell density of >1 × 106 cells/mL (Tocquin et al. 2012). Tocquin et al. (2012) attributed the increased growth to its low N/P ratio of 0.6:1 (1.00 mM nitrate and 1.63 mM phosphate), much lower than other media that have been tested (Table 6.3). Reformulating the medium could further increase biomass productivities, and only certain studies have investigated this in detail (Fábregas et al. 2000; Tocquin et al. 2012; Tripathi et al. 1999; Wang et al. 2019). An emphasis should be on the macronutrients with the concentrations of carbon, nitrate, and phosphate and the C/N/P ratio.
With regard to the nitrogen source, sodium nitrate has been determined to be optimal (Sarada et al. 2002). Feng et al. (2017) deduced that according to the elevated activity of nitrate reductase in the H. lacustris metabolism, the concentrations of sodium nitrate, monopotassium phosphate, and sodium acetate could be determined (3.53 mM, 0.33 mM, and 13.16 mM, respectively) with nitrate being the main influencing factor. However, by utilising a newly isolated strain (JNU35), it was determined that BBM with sodium nitrate replaced by urea (18 mM) was optimal for biomass productivity, attributable to providing a nitrogen and carbon source (Wang et al. 2019). Furthermore, an interesting observation was noted where the strain was observed to have increased growth when resuspended in the red stage in nitrate-depleted BBM medium, attributed to the synthesis of storable nitrogen compounds in the green stage (Wang et al. 2019). This was also noted by Butler et al. (2017) with cells initially cultivated in BG-11, followed by resuspension in nitrate-depleted 3N-BBM + V.
3.2 Improved Astaxanthin Content
Astaxanthin induction factors have been well characterised, and nitrate deprivation has been determined as critical with high light enhancing the rate of astaxanthin accumulation (Christian et al. 2018; Del Río et al. 2008; García-Malea et al. 2009). To date, the highest astaxanthin content (7.72% DW) has been attained using a sequential heterotrophic-photoautotrophic production process where astaxanthin induction was initiated under nitrate deprivation and 5% CO2 (Kang et al. 2005), but the highest astaxanthin productivity has been attained with the one-stage process, albeit with a low content of astaxanthin (1.1% DW) (Del Río et al. 2008). Utilising the one-stage process, the astaxanthin content (0.8–1.1% DW) is lower than the two-stage process (4% DW) with a lower astaxanthin proportion of the total carotenoid fraction (65% compared with 95%) (Aflalo et al. 2007). If a two-stage process is adopted, the initial biomass density in the red stage plays a critical role in increasing astaxanthin content with 0.8 g/L DW being optimal (Wang et al. 2013). With an initial density of 0.1 g/L, severe photoinhibition was observed, and using 2.7 g/L DW resulted in light limitation for astaxanthin production in outdoor cultivation (Wang et al. 2013).
Regarding other parameters, it has been identified that the optimal temperature for astaxanthin production is 27 °C (Evens et al. 2008). Increased temperatures have been hypothesised to result in the synthesis of astaxanthin through oxygen radical formation (Tjahjono et al. 1994). Abiotic stresses generally cause the generation of reactive oxygen species (ROS) within the cell and induce astaxanthin as a defence strategy (Eui et al. 2018). The addition of 0.45 mM Fe2+ in the form of ferrous sulphate can significantly increase the biosynthesis of carotenoids attributable to the formation of hydroxyl radicals (Kobayashi et al. 1993). Adding 0.45 mM Fe2+, 2.25 mM sodium acetate, and high temperature (30 °C) results in a further increase in carotenoids (Kobayashi et al. 1993; Tjahjono et al. 1994). This has also been validated using transcriptome analysis where high light and sodium acetate addition (25 mM) resulted in expression of essential genes related to carotenoid biosynthesis and FA elongation, but this was not observed for Fe2+ (20 μM) which showed a decrease in gene expression related to photosynthesis-antenna proteins (He et al. 2018).
For astaxanthin induction in the red stage, it has been reported that H. lacustris cannot be cultivated heterotrophically in the dark and the production of astaxanthin should adopt a photosynthetic mode (Guedes et al. 2011). Using high light (950 μmol photons/m2/s) in an indoor enclosed system for the red stage is costly and not economically feasible for indoor production (Olaizola 2000; Guedes et al. 2011). It has been determined that 300 μmol photons/m2/s as an incident light intensity is the optimal for astaxanthin production with light saturation reached at 500 μmol photons/m2/s, and 600 μmol photons/m2/s resulted in a lower astaxanthin content with increased cell death (Evens et al. 2008; Giannelli et al. 2015; Li et al. 2010). It has been suggested that inducing astaxanthin in the red stage in a flat plate bioreactor (3 cm light path) would elevate the astaxanthin content to 5.6% after 15 days compared with 3.7% DW in a glass column (6 cm light path) (Wang et al. 2019). Alternatively utilising different wavelengths of light could reduce the light intensity required for astaxanthin induction. Blue light (380–470 nm) is well known to induce the transition to encystment and can be utilised as part of a two-stage strategy (red light for the green stage and blue light at a high light intensity for astaxanthin induction) with an astaxanthin content up to 5.5% DW within 12.5 days (Katsuda et al. 2004; Lababpour et al. 2004). Sun et al. (2015) determined that blue and white light (3:1) at 95 μmol photons/m2/s increased the astaxanthin yield by 11.8% compared with blue light alone (91.8 mg/L) and reduced the encystment time.
Plant hormones have been investigated for elevating yields including jasmonic acid, abscistic acid, and methyl jasmonate and have been reviewed elsewhere (Shah et al. 2016). In addition, growth regulators such as salicylic acid, gibberellic acid, and 2,4-epibrassinolide were found promising for increasing astaxanthin content as reviewed by Shah et al. (2016). With these compounds, astaxanthin genes were upregulated (tenfold increase). The highest improvement of astaxanthin was with 50 mg/L salicylic acid under low light conditions (25 μmol photons/m2/s) elevating the astaxanthin content sevenfold, but this yield was comparatively low compared to other reports. It was observed that higher levels of the hormones/growth regulators adversely affected growth and astaxanthin accumulation (Gao et al. 2012). In addition, micronutrients such as selenium have been observed to result in an increase in astaxanthin but have resulted in declines in the biomass yield (Zheng et al. 2017).
Utilising γ-ray-irradiated mutants cultivated under CO2 at 6% in conjunction with high light (108 μmol photons/m2/s) resulted in 2.4-fold higher astaxanthin content than with the wild-type strain (Cheng et al. 2016). The same mutant could be cultivated with 15% CO2 resulting in 5.8 times higher astaxanthin accumulation than under aeration with astaxanthin induction occurring after 24 h (Li et al. 2017). Christian et al. (2018) observed that H. lacustris cells cultivated under 15% CO2 and high light (300 μmol photons/m2/s) turned orange within 1 day, and by day 2, astaxanthin was accumulated with a final astaxanthin content of 3.62% DW after 4 days, but a reduction in cell density was observed, presumably attributable to photobleaching.
To date, the most successful parameters for inducing astaxanthin in the red stage have been nitrate deprivation/depletion combined with high light and an increase in the C/N ratio. Further improvements have been proposed using PBR developmental work, blue light, elevated temperatures, plant hormones, and micronutrients and through genetic engineering. It is likely that these effects when utilised in combination will have a synergistic effect on the astaxanthin content. A particularly interesting aspect would be on developing a process with high astaxanthin accumulation in the red stage whilst the cells are still continuing to divide as revealed by Wang et al. (2019).
3.3 Minimising Cell Die-Off Through Photobleaching in the Red Stage
In the green stage, controlled culture conditions are provided with low light intensity for maximising growth as discussed above. Upon attainment of a suitable biomass concentration, the culture is subjected to unfavourable conditions, usually involving high light in the red stage to induce astaxanthin biosynthesis (Olaizola 2000). During the initial transition to the red stage, mass cell die-off (photobleaching) has been noted ranging from 20% to 80% depending on the strain, PBR, and induction conditions (Wang et al. 2013). After 24 h of high light, cells have been noted to lose their flagella, and after 48 h the cell density has been observed to decrease by 41% (Gu et al. 2014). The surviving cells have been observed to undergo profound biochemical and cellular changes, with transformations in the life cycle from the vegetative to the aplanospore stage (Wang et al. 2014). It has been observed that H. lacustris cells exposed to higher irradiance accumulated more astaxanthin, but exhibited higher cell mortality (Li et al. 2010). The exact cause of cell death when cells have been transferred from the green to the red stage remains unknown (Wang et al. 2014), but there is a necessity to reduce die-off between the green and the red stage to minimise losses in biomass productivity.
It has been suggested that optimising the initial cell concentration for the red stage (Wang et al. 2013) and applying palmelloid cells rather than green motile macrozooids to the red stage for the induction of astaxanthin may present a promising strategy for greater biomass and astaxanthin production (Wang et al. 2014). Another strategy is to use stepwise irradiance allowing the cells to acclimate (Park et al. 2014). Of the vegetative cells, the green motile macrozooid stage has been determined to have a higher susceptibility to photooxidative stress than palmelloids when transferred to the red stage (Han et al. 2012; Harker et al. 1996; Sarada et al. 2002). In most H. lacustris strains, there is an overall tendency to rapidly transform to palmelloid stages, and these morphotypes are often preferred over green motile macrozooids because they are believed to be more resistant to imposed stressors (Allewaert et al. 2017; Wang et al. 2013). Choi et al. (2011) determined that transferring these palmelloids (senescent cells) to the red stage resulted in a higher capacity to accumulate astaxanthin. However, there is no evidence that strains with more palmelloids will have a higher associated astaxanthin productivity (Allewaert et al. 2017).
3.4 Downstream Processing
Downstream processing encompasses harvesting, cell disruption, drying, extraction, encapsulation, and formulation of the compound. Depending on the application, downstream processing can account for 20–40% of the costs of the production process (’t Lam et al. 2018). H. lacustris aplanospores are large (>50 μm) and, therefore, can be harvested by gravitational settlement (6–8 h in a pond, then 12–24 h for tank sedimentation) resulting in a concentration factor of 5.33, and subsequently, centrifugation (disk-stack) is undertaken resulting in a total suspended solid content of 15% (Li et al. 2011; Olaizola and Huntley 2003; Panis and Rosales 2016). However, as the harvesting process progresses, the biochemical components including the astaxanthin can change affecting the product, especially in tropical locations with high light and heat; therefore, this stage requires further development work and optimisation.
For the cell disruption and extraction stage, this can either be conducted using a wet or dry method. To obtain dry biomass, freeze drying, spray drying, and drum drying have been employed (Kamath and Sarada 2007), but spray drying has been concluded to be the most appropriate in the case of H. lacustris-derived astaxanthin taking into account cost and recovery rate (95–100%), resulting in a 5% moisture content (Panis and Rosales 2016; Pérez-López et al. 2014). Care has to be taken during cell disruption and extraction not to damage the carotenoids which are vulnerable to thermal degradation and oxidation. Carotenoids such as astaxanthin are unstable due to their structural configuration (3-hydroxy, 4-keto end-group) (Mendes-Pinto et al. 2001). It has to be noted that drying biomass in conjunction with mechanical disruption results in enhanced extraction efficiency, but the energy burden increases along with the financial cost (Panis and Rosales 2016).
The aim of cell disruption and extraction is to release the astaxanthin from the thick trilaminar sheath and algaenan cell walls. Methods investigated for cell disruption and extraction of dried haematocysts are generally categorised as mechanical and nonmechanical (Fig. 6.11) (Mendes-Pinto et al. 2001; Kim et al. 2016; Shah et al. 2016; Molino et al. 2018; Liu et al. 2018; Kapoore et al. 2018). Bead milling has been suggested as the most effective and energy-efficient method for the extraction of astaxanthin, and dried biomass is used as the feedstock with a dried algal cake between 100 and 200 g/L being optimal (Greenwell et al. 2010). Alternative methods manipulating the life cycle of H. lacustris have been investigated by utilising cell germination (12–18 h), combined with ionic liquid treatment (Praveenkumar et al. 2015). Adding nitrate to the medium of formed aplanospores resulted in zooids being released without rigid cell walls, resulting in 19.2 pg astaxanthin/cell after 1 min ionic liquid extraction with 1-ethyl-3-methyllimidazolium ethylsulfate at room temperature (Praveenkumar et al. 2015). This method had several associated advantages including less-toxic solvent use, a lower energy input, and the avoidance of thermal stress, but future challenges to address are improving the germination rate and how to recover the expensive ionic liquids.

Downstream processing for the production of astaxanthin from H. lacustris: (a) Aplanospore cells with desired astaxanthin product within the cell, (b) harvesting and drying, (c) cell disruption and extraction, and (d) encapsulation and formulation
Today, there is a desire for natural and environmentally friendly methods of extraction to be developed because traditional solvent extraction requires large quantities of organic solvents and is labour intensive, and the labile pigments can be exposed to excessive light, heat, and oxygen (Denery et al. 2004). There is a requirement for a nontoxic method which is inexpensive in utilising a green chemistry approach. A successful alternative has been SFE, and several companies such as Phasex Corporation are now operating specifically in the SFE field for the extraction of microalgae for the production of high-value nutraceuticals (Phasex Corporation 2015). SFE is the use of a substance such as CO2 or water at a temperature and pressure above their critical point between the typical liquid and gas phase. Compared to most solvents, CO2 has been found to be relatively cheap, nontoxic, chemically inert, and stable (Guedes et al. 2011). It has been found that ScCO2 is industrially scalable and has been proven for coffee decaffeination, the extraction of hops, and for astaxanthin extraction from H. lacustris (Kwan et al. 2018).
Supercritical fluids have their own physicochemical properties similar to both gases and liquids such as high compressibility, high diffusibility, low viscosity, and a low surface tension allowing the fluid to easily diffuse through the natural extractant matrix, achieving higher-quality extractions compared to conventional liquid solvents (Pan et al. 2012). CO2 is commonly used as the SFE solvent (Nobre et al. 2006), but CO2 is not a good solvent for extraction with nonpolar molecules such as astaxanthin, and to improve the solvating power requires a cosolvent such as ethanol or olive oil to improve the polarity of CO2 (Cheng et al. 2018; Wang et al. 2012). The challenge to overcome with ScCO2 is to ensure high and consistent recovery rates of astaxanthin, but in some cases only 50% post cell disruption has been observed (Cheng et al. 2018; Nobre et al. 2006). Furthermore, ScCO2 has a high investment cost (Kadam et al. 2013) and is a labour-intensive step of the production process requiring several hours for the extraction of astaxanthin from the biomass (Michalak and Chojnacka 2014). A way of mitigating this cost could be utilising a biorefinery approach with ScCO2 for the extraction of astaxanthin and TAG simultaneously based on density sequential extraction (Kwan et al. 2018). Di Sanzo et al. (2018) extracted multiple products of interest with ScCO2 with ball milling as a pretreatment with the maximum recovery of astaxanthin and lutein (98.6% and 52.3%, respectively) at 50 °C, 500 bar, and the maximum recovery of FA (93.2%) at 65 °C, 550 bar. Interestingly, at lower recovery rates, the purity of these compounds was higher (Di Sanzo et al. 2018).
To reduce the cost of the production process, wet processing methods have recently been explored. If a wet processing method is used, the extracted astaxanthin-containing biomass must be processed within a few hours to avoid spoilage (Shah et al. 2016). A new method of wet cell disruption has been developed involving hydrothermal disruption (200 °C, 10 min, 6 MPa) which has resulted in near-complete astaxanthin extraction from the biomass and is a more environmentally friendly method as water evaporation is avoided, reducing the energy input (Cheng et al. 2017b). However, it should be noted that although total astaxanthin content was similar to other treatments, this method did result in a loss of astaxanthin diesters, and changes to the stereoisochemistry were not investigated which warrants further investigation. Utilising a wet extraction method with hydrothermal disruption in combination with ScCO2 extraction (8 MPa, 55 °C, with ethanol as a cosolvent, CO2:ethanol (4:1), flow rate 100 μL/min and 25 μL/min) resulted in a 98% recovery rate of astaxanthin in a total of 30 s (Cheng et al. 2018). The high cost of cell disruption and extraction in conjunction with the process being time consuming has led to studies on cell wall-deficient mutants, MT 537 and MT 2978, which were obtained by chemical mutagenesis with reduced thickness of secondary cell walls to overcome the need for extraction, potentially reducing the costing of a cell wall disruption process (Wang et al. 2005), but further experimentation has not been undertaken, and more work is warranted before taking this research to pilot scale.
Although harvesting and extraction of astaxanthin from H. lacustris are well studied, the encapsulation and formulation of astaxanthin have received limited attention. The methods for encapsulation have been reviewed in detail by Khalid and Barrow (2018). Typical methods for encapsulation include high-pressure homogenisation, emulsification, phase inversion, nanoparticles, and microencapsulation (Fig. 6.11). Difficulties have been observed with maintaining the stability and functionality of astaxanthin during the final product formulation stage. There is a requirement for low energy methods for encapsulation, but scalability is the limiting factor in the success of emulsification technologies (Khalid and Barrow 2018). The stability of the encapsulated astaxanthin can be affected by the matrix composition, emulsifier type, and the stabilisers used, but the ingredients need to ensure functionality of the product and have the ability to satisfy regulatory requirements (Khalid and Barrow 2018). Further work in this area will result in a greater product range of astaxanthin products including beverages and creams (Fig. 6.11).
3.5 Contamination Threats
H. lacustris cultures obtained from the environment are often heavily contaminated by other organisms, including protozoa, other algae, fungi, and bacteria (Kim et al. 2011; González et al. 2009). Extensive successive isolation steps are required for obtaining an axenic culture which requires great expertise. To date, micropipetting, differential centrifugation, dilution, phototaxis, purification using UV light, and antibiotic treatment have been investigated (Andersen 2005; Allewaert et al. 2015). Cho et al. (2013) developed a comprehensive protocol for yielding axenic strains from environmental samples by using ultrasonication, cell sorting, and micropicking using agar, without the need for antibiotics or time-consuming micropipetting. Ultrasonication was found to reduce bacterial and fungal loading by detaching them from flocs of microalgae (Cho et al. 2013). Cho et al. (2013) reported that fluorescent-activated cell sorting (FACS) resulted in the removal of 99.5% of the bacteria, but it was difficult to completely remove the attached bacteria from all of the life stages of H. lacustris, especially palmelloids and aplanospores. FACS requires expensive equipment, is costly to maintain, and requires trained personnel; therefore, it is not suitable for small biotechnology companies.
H. lacustris is extremely susceptible to contamination in both the green and red stages, and predators can eliminate 90% of the Haematococcus biomass in <72 h (Bubrick 1991). It has been concluded that large-scale single-phase open-pond systems have proved unsatisfactory for the production of H. lacustris due to difficulties with contamination and control (Bubrick 1991; Margalith 1999). Contamination in microalgal cultures has become arguably the greatest threat to the industry (Day 2013; Han et al. 2013a, b). Proctor (1957c) found that of the microalgae investigated, H. lacustris was the most sensitive and Chlamydomonas reinhardtii was found to readily outcompete it within 3–5 days after inoculation, presenting a major threat of outcompeting H. lacustris when cultivated in open ponds in large-scale culture. Vampyrellids (common freshwater amoeba) may also present a threat to the H. lacustris industry, known to perforate algal cell walls to extract the cell contents (Carney and Lane 2014), increasing biomass loss and leakage of astaxanthin. The antibacterial/antiparasitic agent metronidazole has been investigated, and 3 μg/mL was determined to be effective for eliminating protozoa in H. lacustris cultures (Torres-Carvajal et al. 2017). To date, there have been limited publications on the issues of contamination in the large-scale production of H. lacustris (Poonkum et al. 2015; Torzillo et al. 2003). It has been argued that Cyanotech’s red stage process of astaxanthin induction in open ponds for 5–6 days is too short for a contaminant to impact the system and the culture conditions are unsuitable for the growth of any possible contaminant (Olaizola and Huntley 2003). In industry, cases of contamination remain unreported, with the exception of Fuji Chemicals BioDome™ system in Hawaii (Algae Industry Magazine 2015). This may be due to insufficient monitoring of stocks, and commercial sensitivity definitely has a role. It has been identified in a commercial sample from AlgaeLabs Ltd. that the main microalgal contaminant was the Chlorophyte Coelastrella sp. on the basis of ITS ½ fragment sequencing (Dawidziuk et al. 2017).
Since 2008, there have been several publications on a new chytrid contaminant of H. lacustris cultures (Gutman et al. 2009; Hoffman et al. 2008), described as the most serious hurdle responsible for reductions in astaxanthin productivities and frequent culture collapse (Han et al. 2013a, b). The source of the H. lacustris chytrid remains unknown, but it has been identified as Paraphysoderma sedebokerensis and is closely related to the plant pathogen Physoderma (Gutman et al. 2009). Glucose in the medium was found to increase susceptibility to infection (Gutman et al. 2009; Hoffman et al. 2008), revealing a potential disadvantage of mixotrophic production. An interesting characteristic of the chytrid is the high level of resistance against desiccation, with the chytrid remaining viable even after being placed in an evacuated desiccator for 3 weeks which may allow it to spread to H. lacustris cultures in the air (Hoffman et al. 2008). Hoffman et al. (2008) reported that when the chytrid was placed in cultures of various strains of H. lacustris (20 strains tested), it infected them all in the palmelloid and aplanospore stages, including NIES-144, SAG 34/1b, SAG 192.80, CCAP 34/19, and SCCAP k-0084.
To date, only H. lacustris SCCAP k-0084 has been investigated in depth for its susceptibility to infection by P. sedebokerensis (Gutman et al. 2011, 2009; Hoffman et al. 2008). Green H. lacustris cultures infected by the chytrid will turn dark brown and will clump (Hoffman et al. 2008). Hoffman et al. (2008) observed that the chytrid infected palmelloids and aplanospores, but motile macrozooids remained unaffected. Agitation of cultures resulted in infection rates of up to 100% after 3 days in the green stage (Hoffman et al. 2008). Gutman et al. (2009) revealed that after 4 days of incubation, all H. lacustris cells in the aplanospores stage had P. sedebokerensis attached. H. lacustris cells were thought to be more susceptible to infection when the cultures were nitrogen starved (Gutman et al. 2009) and to be a representative of an unfavourable environment for H. lacustris when aplanospore formation occurs. Gutman et al. (2009) revealed that of the 13 Chlorophytes studied, the chytrid appeared to be Haematococcus specific with infection occurring both in the green and red stages. Information on the pathogen’s life cycle is scarce, and little data exists on its nutritional requirements and mode of infection (Strittmatter et al. 2016). It is hypothesised that P. sedebokerensis transitions between a vegetative and resting phase depending on favourable or unfavourable growth conditions (Strittmatter et al. 2016). Future research on dissemination of the amoeboid and flagellated propagules of P. sedebokerensis warrants investigation as these have been identified as the most vulnerable to adverse environmental conditions, but the life cycle is complex and has not been fully elucidated (Strittmatter et al. 2016). Further emphasis should be on an early prevention or elimination strategy to avoid significant culture collapses of this important commercial alga. Currently, the pathogen’s nutritional requirements are being determined through integrated metabolomics and transcriptomics, and H. lacustris cultures are being screened for random clones resistant to infection and with high astaxanthin productivities (A4F 2015).
To date, there is a lack of effective solutions to prevent or treat microbial contamination of mass cultures on a commercial scale, and most methods are reliant on microscopy and staining for early detection. Other methods have included flow cytometry, but this is a high capital investment for small microalgal biotechnology companies and cannot be used to identify microalgal contamination (Carney and Lane 2014). More recently, a high-resolution melting (HRM) analysis has been developed to detect fungal and microalgal contaminants and can identify contaminants with low levels of DNA in 5 h (2.5 ng/mL for fungi and 1.25 ng/mL for microalgae) (Dawidziuk et al. 2017). Natural, algal-mediated chemicals exist such as abscisic acid, and chemical agents have been observed to be effective against chytrids such as copper sulphate and Triton-N (Carney and Lane 2014). To date, five patents have been filed in relation to P. sedebokerensis, but the methods either rely on an early detection method using qPCR or epifluorescence microscopy which is not easily accessible in the field or easily available to small biotechnology companies, or through the use of fungicides, but the efficacy is unknown (WO2013127280A1, CN106755393A, AU2013353154B2, CN103857785A, CN202519240U). Developing methods of control which could lead to the eradication of this ‘pest’ are critical. In addition, crop protection strategies against other grazers and predators are also required which remain environmentally inert.
3.6 The Question of Economics
Producing astaxanthin from H. lacustris is more costly than other species of algae such as Spirulina due to the necessity for PBRs in the green stage, high electricity consumption, and the requirement for extraction of astaxanthin from the thick-walled cells, adding to the overall production cost (Issarapayup et al. 2011; Li et al. 2011). As outlined above in terms of overall economics, H. lacustris-derived astaxanthin is more costly to produce than the synthetic form, with production costs for H. lacustris-derived astaxanthin at large companies such as Cyanotech, Alga Technologies, and Mera Pharmaceuticals estimated at up to $3600 kg−1 (Li et al. 2011).
Several studies have produced detailed techno-economic assessments for the economical production of astaxanthin in different locations: Europe, the Middle East, and the Far East. When H. lacustris was cultivated in a flat plate airlift PBR, it was determined that the reactor size and the cost of production were directly correlated, but a biomass reduction was simultaneously observed (Issarapayup et al. 2011). Issarapayup et al. (2011) identified that one of the major costs of cultivation of H. lacustris was the high electricity costs (40% of total operating costs); however, it was determined that artificial illumination results in a 107% higher productivity. Using a life cycle assessment (LCA) in Ireland, it was determined that electricity dominated the environmental burdens (Pérez-López et al. 2014). In Ireland, astaxanthin can only be produced outdoors for 5 months of the year with indoor production essential for the remainder of the calendar year (Pérez-López et al. 2014). Lower light intensities and changes in bioreactor design could aid in reducing the cost of the production process and limit the environmental burden. Reusing the medium was not an effective option for reducing costs because the productivity of the system decreased by 30% (Issarapayup et al. 2011), but utilising wastewater could offer a cost-effective alternative.
A process design and an economic assessment were conducted for astaxanthin production in Lebanon, and it was determined that at a market price higher than US $1500 kg−1, a production process could be economically feasible, and if the astaxanthin market price was US $6000 kg−1, a payback period of 1 year and a return on capital employed (ROCE) of 113% were possible (Zgheib et al. 2018). This was based on an annual production of 2592 kg of astaxanthin taking into account the harvesting costs (gravity sedimentation and disk-stack centrifuge), cell disruption (bead miller), drying (spray dryer), extraction (supercritical CO2), and the fact that the residual biomass would be fed into an anaerobic digester and used for biogas and as a biofertiliser (Zgheib et al. 2018). The cost of the upstream processing was not taken into account and, therefore, represents an extreme underestimate.
Based on a modelling approach simulating large-scale production of astaxanthin, it has been proposed that the cost of producing astaxanthin in Livadeia, Greece, and Amsterdam, Netherlands, could be €1536 kg−1 astaxanthin (426 kg/year for a 2 ha site) and €6403 kg−1 (143 kg/year for a 2 ha site), respectively (Panis and Rosales 2016). For the upstream process, horizontal tubular PBRs (5 cm diameter) were chosen for the green stage (pH 7.5) and open raceway ponds for the red stage (pH 8.0) with an assumption that each system occupied 1 ha. For the CO2 supply, flue and flaring gases were selected (2.2 mg/L CO2—5–10% CO2). For the downstream process, harvesting encompassed gravitational settlement followed by subsequent disk-stack centrifugation. The biomass was then spray dried resulting in a moisture content of 5%, and subsequent supercritical fluid extraction was applied (60 °C, 30 MPa with ethanol as a cosolvent (9.4%)) resulting in a 10–20% astaxanthin oleoresin. It was elucidated from the model that June–August were the most productive months in terms of astaxanthin production. It was determined that temperature was the most sensitive parameter for astaxanthin productivity. It was found that using the residual biomass as a biofertiliser ($30–60 kg−1) could be a more economical process. Water consumption was found to be high, and water recycling in the red stage has been suggested, resulting in an improvement in the economics and footprint (Panis and Rosales 2016). Tubular PBR cooling was observed to be the most energy-intensive process (Panis and Rosales 2016). It was observed that cultivation in the PBRs was the highest cost of the process with tubular PBR cooling being the major costing. Even with these cost savings, it was concluded that H. lacustris-derived astaxanthin in Greece and the Netherlands could not compete with synthetic production of astaxanthin (€880 kg−1) for the feed market (Panis and Rosales 2016). It was further concluded that cultivation in equatorial regions such as Hawaii and Israel was more favourable (Panis and Rosales 2016).
For H. lacustris-derived astaxanthin to be cost competitive with the synthetic form, significant developments in the streamlining of the upstream (culturing) and downstream processing (dewatering, cell disruption, and extraction) will be required. Currently, the easiest way to reduce overall cost is to transfer production to a low-cost site, which is why a large number of companies are operating in China such as BGG. Li et al. (2011) published a report on the potential production cost of H. lacustris-derived astaxanthin outdoor operation in Kunming, China, using airlift tubular bioreactors and a raceway pond. It was assumed 33 tons of biomass per year (2.5% astaxanthin DW) could be produced equating to 914 kg of astaxanthin, and assuming a 10-year depreciation on fixed capital cost, the direct production cost for biomass and astaxanthin was estimated at $18 kg−1 and £718 kg−1, respectively.
In addition to the production process being economical, it also needs to be environmentally sustainable reducing the carbon footprint. A key way to do this is through energy reduction, utilising renewable energy sources, and through reductions in water consumption. To date, only one LCA has been published on H. lacustris-derived astaxanthin, and electricity represented the major contributor to the environmental burden, particularly in the green stage for cultivation (Pérez-López et al. 2014). Companies such as AlgaTechnologies have implemented solar power as the primary source of energy (specified at 250 W, 15% conversion efficiency, 1.65 m2) (AlgaTechnologies 2015; Panis and Rosales 2016). In terms of water consumption, industrial cultivation of H. lacustris for astaxanthin requires 1000–1500 tons of freshwater for the production of 1 ton of H. lacustris biomass, but attached cultivation in the red stage has already been showcased to be effective for astaxanthin induction, and the water consumption is 30% of that in an open pond (Wan et al. 2014a).
Utilising a biorefinery approach using a ‘high-value product first’ principle where astaxanthin, phytosterols, and PUFAs are all produced (Bilbao et al. 2016), with the residual biomass being used as a protein source or a biofertiliser, could offer a future sustainable production process. Utilising waste streams such as carbon dioxide from flue gases and carbonates from the soft drink industry could offer cost reductions as well as environmentally sustainable solutions. It has been identified that if flue gases are used, a buffering system is required to the conventional and expensive HEPES, and Choi et al. (2017) determined that utilising a bicarbonate and phosphate buffer could be suitable using 10 mM KOH and 0.1 mM phosphate enabling a pH of 7 to be maintained.
4 Exploiting an Alternative H. lacustris Life Cycle Stage and the Future Direction for the Astaxanthin Industry
To target the global aquaculture market, around 130–1000 tons of astaxanthin is required annually to feed the salmonids with a diet containing 50 mg astaxanthin/kg (Zhang et al. 2009; Solovchenko and Chekanov 2014). Cerón et al. (2007) stated that the evaluation of the quality of the H. lacustris biomass should take into account both astaxanthin content and the FA profile. It has been identified that aquaculture-derived salmon contains ten times less astaxanthin than wild salmon and the bioaccessibility of astaxanthin in wild salmon is greater (Chitchumroonchokchai and Failla 2017). It has been found that during in vitro digestion of salmon flesh, >80% of the astaxanthin is recovered in the human body (Chitchumroonchokchai and Failla 2017).
Sommer et al. (1991) had clearly revealed that H. lacustris intact cells failed to achieve satisfactory pigmentation in salmonids and had to be disrupted which was further validated by Mendes-Pinto et al. (2001). Cell disruption in a scalable pressure treatment system (5000 psi for bioavailable astaxanthin) was deemed effective for lysing H. lacustris cells, and ethoxyquin was added to minimise oxidation (Young et al. 2017). It was shown that the lysed cells should be spray dried, incorporated into the feed, and frozen at −20 °C before being fed to salmonids for commercially acceptable pigmentation (Young et al. 2017). However, it is well known that the cell disruption and extraction costs are high. To successfully incorporate H. lacustris-derived astaxanthin into aquaculture feeds, a series of developments need to be made. One idea is to increase the biomass and astaxanthin productivities in the two-stage system through PBR development, optimised cultivation, and induction parameters in conjunction with operating in a low-cost locality for an economically favourable process. In addition, a biorefinery process could be implemented where the PUFAs are sold in an aquaculture feed in conjunction with a defatted microalga for partial fishmeal protein replacement (12.5% has been showcased in shrimp feeding trials) (Shah et al. 2018).
Another alternative is exploiting the accumulation of astaxanthin in red motile macrozooids. Synthesis of astaxanthin is not induced by the cessation of cell division and is independent of aplanospore formation (Butler et al. 2017; Hagen et al. 2000). Red motile macrozooids have been formed that contain up to 2.74% DW astaxanthin (78.4% of total carotenoids) and are rich in PUFAs; thus, it is envisioned that they could be directly fed to aquaculture species’ (Butler et al. 2017). This is a process that is being explored at Brevel Ltd., Israel (Fig. 6.12). Currently, the biomass yield is low, and therefore, this could be incorporated into the one-stage process as devised by Del Río et al. (2008). Adding value to the H. lacustris biomass through the development of an oral vaccine such as against furunculosis in salmonids and immune-boosting supplements to reduce the mortality rates (which can be as high as 27%) (Overton et al. 2018) could increase the supply of H. lacustris to the aquaculture sector. Selenium is an example of an immune-boosting supplement and is incorporated into selenoproteins that have antioxidant and anti-inflammatory effects, and selenium is also highly protective against mercury which has been found to bioaccumulate in fish (Ralston et al. 2014; Rayman 2012). Micronutrients such as selenium could be added directly to the medium to incorporate into the cell, and decreases in biomass have not been observed at 3 mg/L (17.3 μM), and total selenium could accumulate up to 646 μg/g with 380 μg/g organic selenium (Zheng et al. 2017). Many developments have occurred in increasing the biomass/astaxanthin productivities and cost reductions from H. lacustris-derived astaxanthin, and the future bodes well for the replacement of synthetic astaxanthin by this biobased source.

Brevel Ltd. has a technology based on indoor internally illuminated photobioreactors which have a significantly lower surface area and are thus less prone to contamination and have better monitor and control processes. Light is either natural sunlight concentrated and transported into the cultivation tank via optical fibres or based on artificial lighting. Natural sunlight is spectrally filtered at the source in order to reduce heat and damage to cells by IR and UV, and thus, temperature control requirements are significantly reduced. The enclosed and fully controlled environment makes the possibility of mixotrophic cultivation viable and thus could further significantly increase yields and reduce cultivation costs. Under current estimates, Brevel will be capable of producing algal astaxanthin for the salmonid market by 2020 at a comparable retail price to the current synthetic product
References
’t Lam, G. P., Vermuë, M. H., Eppink, M. H. M., Wijffels, R. H., & Van Den Berg, C. (2018). Multi-product microalgae biorefineries: From concept towards reality. Trends in Biotechnology, 36, 216–227.
A4F. (2015). Project 1: Towards controlling chytrid pathogens in industrial cultures of Haematococcus sp. Retrieved May 1, 2019, from https://msc-alff.org/about/alff-students/noreen-hiegle/.
Aflalo, C., Meshulam, Y., Zarka, A., & Boussiba, S. (2007). On the relative efficiency of two- vs. one-stage production of astaxanthin by the green alga Haematococcus pluvialis. Biotechnology and Bioengineering, 98, 300–305.
Alam, M. A., & Wang, Z. (Eds.). (2019). Microalgae biotechnology for development of biofuel and wastewater treatment (pp. 1–655). New York, NY: Springer.
Al-Bulishi, M. S. M. (2015). Health aspects of astaxanthin: A review. Canadian Journal of Clinical Nutrition, 3, 71–78.
Algae Industry Magazine. (2015). Algae Industry Magazine. Retrieved October 10, 2015, from http://www.algaeindustrymagazine.com/?s=astaxanthin.
Algatechnologies. (2015). About Us. Retrieved June 17, 2015, from http://www.algatech.com/about.asp?cat=004.
Allewaert, C., et al. (2015). Species diversity in European Haematococcus pluvialis (Chlorophyceae, Volvocales). Phycologia, 54, 583.
Allewaert, C. C., Vanormelingen, P., Daveloose, I., Verstraete, T., & Vyverman, W. (2017). Intraspecific trait variation affecting astaxanthin productivity in two Haematococcus (Chlorophyceae) species. Algal Research, 21, 191–202.
Andersen, R. A. (Ed.). (2005). Algal culturing techniques. Amsterdam: Elsevier.
Aragreen. (2015). Aragreen. Retrieved May 20, 2015, from http://www.aragreen.com/home.asp.
Asker, D. (2017). Isolation and characterization of a novel, highly selective astaxanthin-producing marine bacterium. Journal of Agricultural and Food Chemistry, 65, 9101–9109.
Bauman, N., Akella, S., Hann, E., Morey, R., Schwartz, A. S., Brown, R., & Richardson, T. H. (2018). Next-generation sequencing of Haematococcus lacustris reveals an extremely large 1.35-megabase chloroplast genome. Genome Announcements, 6, e00181–e001818.
BCC Research. (2015). Boom in astaxanthin supplements boosting global carotenoid market, according to BCC Research. Retrieved July 23, 2015, from http://www.bccresearch.com/pressroom/fod/boom-in-astaxanthin-supplements-boosting-global-carotenoid-market-according-to-bcc-research.
BCC Research. (2018). The global market for carotenoids. Retrieved April 23, 2019, from https://www.bccresearch.com/market-research/food-and-beverage/the-global-market-for-carotenoids.html.
Bilbao, P. G. S., Damiani, C., Salvador, G. A., & Leonardi, P. (2016). Haematococcus pluvialis as a source of fatty acids and phytosterols: Potential nutritional and biological implications. Journal of Applied Phycology, 28, 3283–3294.
Bolin, A. P., Macedo, R. C., Marin, D. P., Barros, M. P., & Otton, R. (2010). Astaxanthin prevents in vitro auto-oxidative injury in human lymphocytes. Cell Biology and Toxicology, 26, 457–467.
Boussiba, S., & Vonshak, A. (1991). Astaxanthin accumulation in the green alga Haematococcus pluvialis. Plant & Cell Physiology, 32, 1077–1082.
Brinda, B. R., Sarada, R., Sandesh Kamath, B., & Ravishankar, G. A. (2004). Accumulation of astaxanthin in flagellated cells of Haematococcus pluvialis - Cultural and regulatory aspects. Current Science, 87, 1290–1295.
Brodie, J., Chan, C. X., De Clerck, O., Cock, J. M., Coelho, S. M., Gachon, C., Grossman, A. R., Mock, T., Raven, J. A., Smith, A. G., Yoon, H. S., & Bhattacharya, D. (2017). The algal revolution. Trends in Plant Science, 22, 726–738.
Bubrick, P. (1991). Production of astaxanthin from Haematococcus. Bioresource Technology, 38, 237–239.
Buchheim, M. A., Sutherland, D. M., Buchheim, J. A., & Wolf, M. (2013). The blood alga: Phylogeny of Haematococcus (Chlorophyceae) inferred from ribosomal RNA gene sequence data. European Journal of Phycology, 48, 318–329.
Burchardt, L., Balcerkiewicz, S., Kokocinski, M., Samardakiewicz, S., & Adamski, Z. (2006). Occurrence of Haematococcus pluvialis Flotow emend. Wille in a small artificial pool on the university campus of the collegium biologicum in Poznan (Poland). Resource Conservation, 1, 163–166.
Bustos-Garza, C., Yáñez-Fernández, J., & Barragán-Huerta, B. E. (2013). Thermal and pH stability of spray-dried encapsulated astaxanthin oleoresin from Haematococcus pluvialis using several encapsulation wall materials. Food Research International, 54, 641–649.
Butler, T., McDougall, G., Campbell, R., Stanley, M., & Day, J. (2017). Media screening for obtaining Haematococcus pluvialis Red motile macrozooids rich in astaxanthin and fatty acids. Biology (Basel), 7, 2.
Capelli, B. (2018). AstaZine™ natural astaxanthin: The supplement you can feel.
Capelli, B., Bagchi, D., & Cysewski, G. R. (2013a). Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods, 12, 145–152.
Capelli, B., Jenkins, U., & Cysewski, G. R. (2013b). Role of astaxanthin in sports nutrition. In Nutrition and enhanced sports performance (pp. 465–471). New York, NY: Academic Press.
Carney, L. T., & Lane, T. W. (2014). Parasites in algae mass culture. Frontiers in Microbiology, 5, 1–8.
CCAP. (2015). Haematococcus pluvialis. Retrieved September 10, 2015, from http://www.ccap.ac.uk/results2014.php.
Cerón, M. C., García-Malea, M. C., Rivas, J., Acien, F. G., Fernandez, J. M., Del Río, E., Guerrero, M. G., & Molina, E. (2007). Antioxidant activity of Haematococcus pluvialis cells grown in continuous culture as a function of their carotenoid and fatty acid content. Applied Microbiological Biotechnology, 74, 1112–1119.
Chekanov, K., Lobakova, E., Selyakh, I., Semenova, L., Sidorov, R., & Solovchenko, A. (2014). Accumulation of astaxanthin by a new Haematococcus pluvialis strain BM1 from the white sea coastal rocks (Russia). Marine Drugs, 12, 4504–4520.
Chekanov, K., Schastnaya, E., Solovchenko, A., & Lobakova, E. (2017). Effects of CO2 enrichment on primary photochemistry, growth and astaxanthin accumulation in the chlorophyte Haematococcus pluvialis. Journal of Photochemistry and Photobiology B: Biology, 171, 58–66.
Chen, T., & Wang, Y. (2013). Optimized astaxanthin production in Chlorella zofingiensis under dark condition by response surface methodology. Food Science and Biotechnology, 22, 1–8.
Chen, J., Liu, L., & Wei, D. (2017). Enhanced production of astaxanthin by Chromochloris zofingiensis in a microplate-based culture system under high light irradiation. Bioresource Technology, 245, 518–529.
Cheng, J., Li, K., Yang, Z., Zhou, J., & Cen, K. (2016). Enhancing the growth rate and astaxanthin yield of Haematococcus pluvialis by nuclear irradiation and high concentration of carbon dioxide stress. Bioresource Technology, 204, 49–54.
Cheng, J., Li, K., Zhu, Y., Yang, W., Zhou, J., & Cen, K. (2017a). Transcriptome sequencing and metabolic pathways of astaxanthin accumulated in Haematococcus pluvialis mutant under 15% CO2. Bioresource Technology, 228, 99–105.
Cheng, X., Riordon, J., Nguyen, B., Ooms, M. D., & Sinton, D. (2017b). Hydrothermal disruption of algae cells for astaxanthin extraction. Green Chemistry, 19, 106–111.
Cheng, X., Qi, Z., Burdyny, T., Kong, T., & Sinton, D. (2018). Low pressure supercritical CO2 extraction of astaxanthin from Haematococcus pluvialis demonstrated on a micro fluidic chip. Bioresource Technology, 250, 481–485.
Chitchumroonchokchai, C., & Failla, M. L. (2017). Bioaccessibility and intestinal cell uptake of astaxanthin from salmon and commercial supplements. Food Research International, 99, 936–943.
Cho, D. H., Ramanan, R., Kim, B. H., Lee, J., Kim, S., Yoo, C., Choi, G. G., Oh, H. M., & Kim, H. S. (2013). Novel approach for the development of axenic microalgal cultures from environmental samples. Journal of Phycology, 49(4), 802–810.
Choi, Y. E., & Park, J. M. (2002). Evaluation of factors promoting astaxanthin production by a unicellular green alga, Haematococcus pluvialis, with fractional factorial design. Biotechnology Progress, 18, 1170–1175.
Choi, Y. E., Yun, Y. S., Park, J. M., & Yang, J. W. (2011). Determination of the time transferring cells for astaxanthin production considering two-stage process of Haematococcus pluvialis cultivation. Bioresource Technology, 102, 11249–11253.
Choi, Y., Joun, J., Lee, J., Hong, M., Pham, H., Chang, W., & Sim, S. (2017). Development of large-scale and economic pH control system for outdoor cultivation of microalgae Haematococcus pluvialis using industrial flue gas. Bioresource Technology, 244, 1235–1244.
Choubert, G., & Heinrich, O. (1993). Carotenoid pigments of the green alga Haematococcus pluvialis: Assay on rainbow trout, Oncorhynchus mykiss, pigmentation in comparison with synthetic astaxanthin and canthaxanthin. Aquaculture, 112, 217–226.
Christian, D., Zhang, J., Sawdon, A. J., & Peng, C. (2018). Enhanced astaxanthin accumulation in Haematococcus pluvialis using high carbon dioxide concentration and light illumination. Bioresource Technology, 256, 548–551.
Chunhui, Z., Jianguo, L. I. U., & Litao, Z. (2017). Cell cycles and proliferation patterns in Haematococcus pluvialis. Chinese Journal of Oceanology and Limnology, 35, 1205–1211.
Cifuentes, A. S., González, M. A., Vargas, S., Hoeneisen, M., & González, N. (2003). Optimization of biomass, total carotenoids and astaxanthin production in Haematococcus pluvialis Flotow strain Steptoe (Nevada, USA) under laboratory conditions. Biological Research, 36, 343–357.
Collins, A. M., Jones, H. D. T., Han, D., Hu, Q., Beechem, T. E., & Timlin, J. A. (2011). Carotenoid distribution in living cells of Haematococcus pluvialis (Chlorophyceae). PLoS One, 6, 1–7.
Comhaire, F. H., El Garem, Y., Mahmoud, A., Eertmans, F., & Schoonjans, F. (2005). Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: A double blind randomized trial. Asian Journal of Andrology, 7, 257–262. https://doi.org/10.1111/j.1745-7262.2005.00047.x.
Cunningham, F. X., & Gantt, E. (2011). Elucidation of the pathway to astaxanthin in the flowers of Adonis aestivalis. Plant Cell, 23, 3055–3069.
Dalay, M. C., Imamoglu, E., & Demirel, Z. (2007). Agricultural fertilizers as economical alternative for cultivation of Haematococcus pluvialis. Journal of Microbiology and Biotechnology, 17, 393–397.
Damiani, M. C., Leonardi, P. I., Pieroni, O. I., & Cáceres, E. J. (2006). Ultrastructure of the cyst wall of Haematococcus pluvialis (Chlorophyceae): Wall development and behaviour during cyst germination. Phycologia, 45, 616–623.
Damiani, M. C., Popovich, C. A., Constenla, D., & Leonardi, P. I. (2010). Lipid analysis in Haematococcus pluvialis to assess its potential use as a biodiesel feedstock. Bioresource Technology, 101, 3801–3807.
Dawidziuk, A., Popiel, D., Luboinska, M., & Grzebyk, M. (2017). Assessing contamination of microalgal astaxanthin producer Haematococcus cultures with high-resolution melting curve analysis. Journal of Applied Genetics, 58, 277–285.
Day, J. G. (2013). Grazers: The overlooked threat to the sustained production of future algal biofuels. Biofuels, 4, 459–461.
Del Río, E., Acién, F. G., García-Malea, M. C., Rivas, J., Molina-Grima, E., & Guerrero, M. G. (2005). Efficient one-step production of astaxanthin by the microalga Haematococcus pluvialis in continuous culture. Biotechnology and Bioengineering, 91, 808–815.
Del Río, E., Acién, F. G., García-Malea, M. C., Rivas, J., Molina-Grima, E., & Guerrero, M. G. (2008). Efficiency assessment of the one-step production of astaxanthin by the microalga Haematococcus pluvialis. Biotechnology and Bioengineering, 100, 397–402.
Denery, J. R., Dragull, K., Tang, C. S., & Li, Q. X. (2004). Pressurized fluid extraction of carotenoids from Haematococcus pluvialis and Dunaliella salina and kavalactones from Piper methysticum. Analytica Chimica Acta, 501, 175–181.
Di Sanzo, G., Mehariya, S., Martino, M., Larocca, V., Casella, P., Chianese, S., Musmarra, D., Balducchi, R., & Molino, A. (2018). Supercritical carbon dioxide extraction of astaxanthin, lutein, and fatty acids from Haematococcus pluvialis microalgae. Marine Drugs, 16, 1–18.
Domínguez-Bocanegra, A. R., Ponce-Noyola, T., & Torres-Muñoz, J. A. (2007). Astaxanthin production by Phaffia rhodozyma and Haematococcus pluvialis: A comparative study. Applied Microbiology and Biotechnology, 75, 783–791.
Dore, J. E., & Cysewski, G. R. (2003). Haematococcus algae meal as a source of natural astaxanthin for aquaculture feeds. Kailua-Kona: Cyanotech Corporation.
Doyle, J. J., & Doyle, J. (1990). Isolation of plant DNA from fresh tissue. Focus, 12, 13.
Dragos, N., Bercea, V., Bica, A., Druga, B., Nicoara, A., & Coman, C. (2010). Astaxanthin production from a new strain of Haematococcus pluvialis grown in batch culture BT. Annals of the Romanian Society for Cell Biology, XV, 353–361.
Droop, M. R. (1955). Some factors governing encystment in Haematococcus pluvialis. Archiv für Mikrobiologie, 21, 267–272.
Droop, M. R. (1956). Haematococcus pluvialis and its allies. I. The sphaerellaceae. Revue Algologique, 2, 53–71.
Elgarem, Y., Lignell, A., & Comhaire, F. H. (2002). Supplementation with astaxanthin (astacarox) improves semen quality in infertile men. In Proceedings of the 13th International Carotenoid Symposium, Honolulu, HI (pp. 180–197).
Elliot, A. M. (1934). Morphology and life history of Haematococcus pluvialis. Archiv für Protistenkunde, 82, 250–272.
Eui, M., Il, H., Seok, H., Hwang, S., & Joon, Y. (2018). Rapid selection of astaxanthin-hyperproducing Haematococcus mutant via azide-based colorimetric assay combined with oil-based astaxanthin extraction. Bioresource Technology, 267, 175–181.
Evens, T. J., Niedz, R. P., & Kirkpatrick, G. J. (2008). Temperature and irradiance impacts on the growth, pigmentation and photosystem II quantum yields of Haematococcus pluvialis (Chlorophyceae). Journal of Applied Phycology, 20, 411–422.
Fábregas, J., Dominguez, A., & Regueiro, M. (2000). Optimization of culture medium for the continuous cultivation of the microalga Haematococcus pluvialis. Applied Microbiology and Biotechnology, 53, 530–535.
Fábregas, J., Otero, A., Maseda, A., & Domínguez, A. (2001). Two-stage cultures for the production of Astaxanthin from Haematococcus pluvialis. Journal of Biotechnology, 89, 65–71.
Fassett, R. G., & Coombes, J. S. (2012). Astaxanthin in cardiovascular health and disease. Molecules, 17, 2030–2048.
Feng, L. H., Xiaonan, L., Xuecheng, Z., Bangxiang, Z., & Yating, H. (2017). Cloning and transcription analysis of the nitrate reductase gene from Haematococcus pluvialis. Biotechnology Letters, 39, 589–597.
de la Fuente, J. L., Rodríguez-Sáiz, M., Schleissner, C., Díez, B., Peiro, E., & Barredo, J. L. (2010). High-titer production of astaxanthin by the semi-industrial fermentation of Xanthophyllomyces dendrorhous. Journal of Biotechnology, 148, 144–146.
Fujii, K., Imazato, E., Nakashima, H., Ooi, O., & Saeki, A. (2006). Isolation of the non-fastidious microalga with astaxanthin-accumulating property and its potential for application to aquaculture. Aquaculture, 261, 285–293.
Gacheva, G., Dimitrova, P., & Pilarski, P. (2015). New strain Haematococcus cf. pluvialis Rozhen-12 - Growth, biochemical characteristics and future perspectives. Genetics and Plant Physiology, 5, 29–38.
Galarza, J. I., Gimpel, J. A., Rojas, V., & Arredondo-vega, B. O. (2018). Over-accumulation of astaxanthin in Haematococcus pluvialis through chloroplast genetic engineering. Algal Research, 31, 291–297.
Gao, Z., Meng, C., Zhang, X., Xu, D., Miao, X., Wang, Y., Yang, L., Lv, H., Chen, L., & Ye, N. (2012). Induction of salicylic acid (SA) on transcriptional expression of eight carotenoid genes and astaxanthin accumulation in Haematococcus pluvialis. Enzyme and Microbial Technology, 51, 225–230.
García-Malea López, M. C., Del Río Sánchez, E., Casas López, J. L., Acién Fernández, F. G., Fernández Sevilla, J. M., Rivas, J., Guerrero, M. G., & Molina Grima, E. (2006). Comparative analysis of the outdoor culture of Haematococcus pluvialis in tubular and bubble column photobioreactors. Journal of Biotechnology, 123, 329–342.
García-Malea, M. C., Brindley, C., Del Río, E., Acién, F. G., Fernández, J. M., & Molina, E. (2005). Modelling of growth and accumulation of carotenoids in Haematococcus pluvialis as a function of irradiance and nutrients supply. Biochemical Engineering Journal, 26, 107–114.
García-Malea, M. C., Acién, F. G., Fernández, J. M., Cerón, M. C., & Molina, E. (2006). Continuous production of green cells of Haematococcus pluvialis: Modeling of the irradiance effect. Enzyme and Microbial Technology, 38, 981–989.
García-Malea, M. C., Acién, F. G., Del Río, E., Fernández, J. M., Cerón, M. C., Guerrero, M. G., & Molina-Grima, E. (2009). Production of astaxanthin by Haematococcus pluvialis: Taking the one-step system outdoors. Biotechnology and Bioengineering, 102, 651–657.
Gassel, S., Schewe, H., Schmidt, I., Schrader, J., & Sandmann, G. (2013). Multiple improvement of astaxanthin biosynthesis in Xanthophyllomyces dendrorhous by a combination of conventional mutagenesis and metabolic pathway engineering. Biotechnology Letters, 35, 565–569.
Giannelli, L., Yamada, H., Katsuda, T., & Yamaji, H. (2015). Effects of temperature on the astaxanthin productivity and light harvesting characteristics of the green alga Haematococcus pluvialis. Journal of Bioscience and Bioengineering, 119, 345–350.
Göksan, T., Ak, I., & Gökpinar, Ş. (2010). An alternative approach to the traditional mixotrophic cultures of Haematococcus pluvialis flotow (Chlorophyceae). Journal of Microbiology and Biotechnology, 20, 1276–1282.
Gómez, P. I., Inostroza, I., Pizarro, M., & Pérez, J. (2013). From genetic improvement to commercial-scale mass culture of a Chilean strain of the green microalga Haematococcus pluvialis with enhanced productivity of the red ketocarotenoid astaxanthin. AoB Plants, 5, 1–7.
Gong, X., & Chen, F. (1997). Rapid detection of heterotrophic growth of Haematococcus pluvialis using indirect conductimetry. Biotechnology Techniques, 11, 841–844.
González, M. A., Cifuentes, A. S., & Gómez, P. I. (2009). Growth and total carotenoid content in four Chilean strains of Haematococcus pluvialis flotow, under laboratory conditions. Gayana Botánica, 66, 58–70.
Granata, T. (2017). Dependency of microalgal production on biomass and the relationship to yield and bioreactor scale-up for biofuels: A statistical analysis of 60+ years of algal bioreactor data. Bioenergy Research, 10, 267–287.
Greenwell, H. C., Laurens, L. M. L., Lovitt, R. W., Flynn, K. J., & Shields, R. J. (2010). Placing microalgae on the biofuels priority list: A review of the technological challenges. Journal of The Royal Society Interface, 7, 703–726.
Groben, R. (2007). Phytoplankton and nutrient analysis of a nuclear fuel-storage pond at Sellafield.
Grünewald, K., Hagen, C., & Braune, W. (1997). Secondary carotenoid accumulation in flagellates of the green alga Haematococcus lacustris. European Journal of Phycology, 32, 387–392.
Grunewald, K., Hirschberg, J., & Hagen, C. (2001). Ketocarotenoid biosynthesis outside of plastids in the unicellular green alga Haematococcus pluvialis. The Journal of Biological Chemistry, 276, 6023–6029.
Gu, W., Li, H., Zhao, P., Yu, R., Pan, G., Gao, S., Xie, X., Huang, A., He, L., & Wang, G. (2014). Quantitative proteomic analysis of thylakoid from two microalgae (Haematococcus pluvialis and Dunaliella salina) reveals two different high light-responsive strategies. Scientific Reports, 4, 6661.
Guedes, A. C., Amaro, H. M., & Malcata, F. X. (2011). Microalgae as sources of carotenoids. Marine Drugs, 9, 625–644.
Guerin, M., Huntley, M. E., & Olaizola, M. (2003). Haematococcus astaxanthin: Applications for human health and nutrition. Trends in Biotechnology, 21, 210–216.
Guiry, M. D. (2010). AlgaeBase. Retrieved June 12, 2015, from http://www.algaebase.org.
Gutiérrez, C. L., Gimpel, J., Escobar, C., Marshall, S. H., & Henríquez, V. (2012). Chloroplast genetic tool for the green microalgae Haematococcus pluvialis (chlorophyceae, volvocales). Journal of Phycology, 48, 976–983.
Gutman, J., Zarka, A., & Boussiba, S. (2009). The host-range of Paraphysoderma sedebokerensis, a chytrid that infects Haematococcus pluvialis. European Journal of Phycology, 44, 509–514.
Gutman, J., Zarka, A., & Boussiba, S. (2011). Evidence for the involvement of surface carbohydrates in the recognition of Haematococcus pluvialis by the parasitic blastoclad Paraphysoderma sedebokerensis. Fungal Biology, 115, 803–811.
Gwak, Y., Hwang, Y.-S., Wang, B., Kim, M., Jeong, J., Lee, C.-G., Hu, Q., Han, D., & Jin, E. (2014). Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus pluvialis. Journal of Experimental Botany, 65, 4317–4334.
Ha, P. T., Hoang, N. H., Lien, N. T. K., Phuong, N. T. D., Huy, N. Q., & Thoa, N. K. (2018). Selection of bacterial strains belonging to the astaxanthin producing Paracoccus genus. Vietnam Journal of Biotechnology, 16(3), 565–572.
Hagen, C., Grünewald, K., Schmidt, S., & Müller, J. (2000). Accumulation of secondary carotenoids in flagellates of Haematococcus pluvialis (Chlorophyta) is accompanied by an increase in per unit chlorophyll productivity of photosynthesis. European Journal of Phycology, 35, 75–82.
Hagen, C., Grünewald, K., Xyländer, M., & Rothe, E. (2001). Effect of cultivation parameters on growth and pigment biosynthesis in flagellated cells of Haematococcus pluvialis. Journal of Applied Phycology, 13, 79–87.
Hagen, C., Siegmund, S., & Braune, W. (2002). Ultrastructural and chemical changes in the cell wall of Haematococcus pluvialis (Volvocales, Chlorophyta) during aplanospore formation. European Journal of Phycology, 37, 217.
Han, D., Wang, J., Sommerfeld, M., & Hu, Q. (2012). Susceptibility and protective mechanisms of motile and non motile cells of Haematococcus pluvialis (Chlorophyceae) to photooxidative stress1. Journal of Phycology, 48, 693–705.
Han, D., Li, Y., & Hu, Q. (2013a). Astaxanthin in microalgae: Pathways, functions and biotechnological implications. Algae, 28, 131–147.
Han, D., Li, Y., & Hu, Q. (2013b). Biology and commercial aspects of Haematococcus pluvialis. In Handbook of microalgal culture: Applied phycology and biotechnology (2nd ed., pp. 388–405). Oxford: Wiley-Blackwell.
Harker, M., & Tsavalos, A. J. (1996). Autotrophic growth and carotenoid production of Haematococcus pluvialis in a autotrophic growth and carotenoid production of Haematococcus pluvialis in a 30 liter air-lift photobioreactor. Journal of Fermentation and Bioengineering, 82, 113.
Harker, M., Tsavalos, A. J., & Young, A. J. (1996). Factors responsible for astaxanthin formation in the chlorophyte Haematococcus pluvialis. Bioresource Technology, 55, 207–214.
Hartung, F., Schiemann, J., & Quedlinburg, D. (2014). Precise plant breeding using new genome editing techniques: Opportunities, safety and regulation in the EU. The Plant Journal, 78, 742–752.
Hashimoto, H., Arai, K., Hayashi, S., Okamoto, H., Takahashi, J., Chikuda, M., & Obara, Y. (2014). Effects of astaxanthin on antioxidation in human aqueous humor. Journal of Clinical Biochemistry and Nutrition, 54, 86–89.
Hata, N., Ogbonna, J. C., Hasegawa, Y., Taroda, H., & Tanaka, H. (2001). Production of astaxanthin by Haematococcus pluvialis in a sequential heterotrophic-photoautotrophic culture. Journal of Applied Phycology, 13, 395–402.
He, B., Hou, L., Dong, M., Shi, J., Huang, X., Ding, Y., Cong, X., Zhang, F., Zhang, X., & Zang, X. (2018). Transcriptome analysis in Haematococcus pluvialis: Astaxanthin induction by high light with acetate. International Journal of Molecular Sciences, 19, 175–193.
Higuera-Ciapara, I., Félix-Valenzuela, L., & Goycoolea, F. M. (2006). Astaxanthin: A review of its chemistry and applications. Critical Reviews in Food Science and Nutrition, 46, 185–196.
Hoffman, Y., Aflalo, C., Zarka, A., Gutman, J., James, T. Y., & Boussiba, S. (2008). Isolation and characterization of a novel chytrid species (phylum Blastocladiomycota), parasitic on the green alga Haematococcus. Mycological Research, 112, 70–81.
Holtin, K., Kuehnle, M., Rehbein, J., Schuler, P., Nicholson, G., & Albert, K. (2009). Determination of astaxanthin and astaxanthin esters in the microalgae Haematococcus pluvialis by LC-(APCI)MS and characterization of predominant carotenoid isomers by NMR spectroscopy. Analytical and Bioanalytical Chemistry, 395, 1613–1622.
Hossain, A. K. M. M., Brennan, M. A., Mason, S. L., Guo, X., Zeng, X. A., & Brennan, C. S. (2017). The effect of astaxanthin-rich microalgae “Haematococcus pluvialis” and wholemeal flours incorporation in improving the physical and functional properties of cookies. Food, 6, E57.
Hu, Z., Li, Y., Sommerfeld, M., Chen, F., & Hu, Q. (2008). Enhanced protection against oxidative stress in an astaxanthin-overproduction Haematococcus mutant (Chlorophyceae). European Journal of Phycology, 43, 365–376.
Ip, P. F., Wong, K. H., & Chen, F. (2004). Enhanced production of astaxanthin by the green microalga Chlorella zofingiensis in mixotrophic culture. Process Biochemistry, 39, 1761–1766.
Issarapayup, K., Powtongsook, S., & Pavasant, P. (2011). Economical review of Haematococcus pluvialis culture in flat-panel airlift photobioreactors. Aquacultural Engineering, 44, 65–71.
Johnson, E., & Schroeder, W. (1995). Microbial carotenoids. Advances in Biochemical Engineering/Biotechnology, 53, 119–178.
Kadam, S. U., Tiwari, B. K., & O’donnell, C. P. (2013). Application of novel extraction technologies for bioactives from marine algae. Journal of Agricultural and Food Chemistry, 61, 4667–4675.
Kaewpintong, K., Shotipruk, A., Powtongsook, S., & Pavasant, P. (2007). Photoautotrophic high-density cultivation of vegetative cells of Haematococcus pluvialis in airlift bioreactor. Bioresource Technology, 98, 288–295.
Kamath, S., Sarada, R. (2007). Biotechnological production of microalgal carotenoids with reference to astaxanthin and evaluation of its biological activity. Doctoral Dissertation, University of Mysore.
Kang, H., & Kim, H. (2017). Astaxanthin and β-carotene in Helicobacter pylori -induced gastric inflammation: A mini-review on action mechanisms. Journal of Cance R Prevention, 22, 57–61.
Kang, C. D., Lee, J. S., Park, T. H., & Sim, S. J. (2005). Comparison of heterotrophic and photoautotrophic induction on astaxanthin production by Haematococcus pluvialis. Applied Microbiology and Biotechnology, 68, 237–241.
Kang, C. D., Lee, J. S., Park, T. H., & Sim, S. J. (2007). Complementary limiting factors of astaxanthin synthesis during photoautotrophic induction of Haematococcus pluvialis: C/N ratio and light intensity. Applied Microbiology and Biotechnology, 74, 987–994.
Kang, C. D., Han, S. J., Choi, S. P., & Sim, S. J. (2010). Fed-batch culture of astaxanthin-rich Haematococcus pluvialis by exponential nutrient feeding and stepwise light supplementation. Bioprocess and Biosystems Engineering, 33, 133–139.
Kapoore, R. V., Butler, T. O., Pandhal, J., & Vaidyanathan, S. (2018). Microwave-assisted extraction for microalgae: From biofuels to biorefinery. Biology, 7(1), 18.
Katagiri, M., Satoh, A., Tsuji, S., & Shirasawa, T. (2012). Effects of astaxanthin-rich Haematococcus pluvialis extract on cognitive function: A randomised, double-blind, placebo-controlled study. Journal of Clinical Biochemistry and Nutrition, 51, 102–107.
Kathiresan, S., Chandrashekar, A., Ravishankar, G. A., & Sarada, R. (2009). Agrobacterium-mediated transformation in the green alga Haematococcus pluvialis (chlorophyceae, volvocales). Journal of Phycology, 45, 642–649.
Katsuda, T., Lababpour, A., Shimahara, K., & Katoh, S. (2004). Astaxanthin production by Haematococcus pluvialis under illumination with LEDs. Enzyme and Microbial Technology, 35, 81–86.
Khalid, N., & Barrow, C. J. (2018). Critical review of encapsulation methods for stabilization and delivery of astaxanthin. Journal of Food Bioactives, 1, 104–115.
Kim, G., Klochkova, T. A., Won Han, J., Kang, S. H., Gu Choi, H., Wha Chung, K., & Ja Kim, S. (2011). Freshwater and terrestrial algae from Ny-Ålesund and Blomstrandhalvøya island (svalbard). Arctic, 64, 25–31.
Kim, J. H., Affan, M. A., Jang, J., Kang, M. H., Ko, A. R., Jeon, S. M., Oh, C., Heo, S. J., Lee, Y. H., Ju, S. J., & Kang, D. H. (2015). Morphological, molecular, and biochemical characterization of astaxanthin-producing green microalga Haematococcus sp. KORDI03 (Haematococcaceae, chlorophyta) isolated from Korea. Journal of Microbiology and Biotechnology, 25, 238–246.
Kim, D. Y., Vijayan, D., Praveenkumar, R., Han, J. I., Lee, K., Park, J. Y., Chang, W. S., Lee, J. S., & Oh, Y. K. (2016). Cell-wall disruption and lipid/astaxanthin extraction from microalgae: Chlorella and Haematococcus. Bioresource Technology, 199, 300–310.
Kiperstok, A. C., Sebestyén, P., Podola, B., & Melkonian, M. (2017). Biofilm cultivation of Haematococcus pluvialis enables a highly productive one-phase process for astaxanthin production using high light intensities. Algal Research, 21, 213–222.
Klochkova, T. A., Kwak, M. S., Han, J. W., Motomura, T., Nagasato, C., & Kim, G. H. (2013). Cold-tolerant strain of Haematococcus pluvialis (Haematococcaceae, Chlorophyta) from Blomstrandhalvøya (Svalbard). Algae, 28, 185–192.
Kobayashi, M., Kakizono, T., & Nagai, S. (1991). Astaxanthin production by a green alga, Haematococcus pluvialis accompanied with morphological changes in acetate media. Journal of Fermentation and Bioengineering, 71, 335–339.
Kobayashi, M., Kakizono, T., & Nagai, S. (1993). Enhanced carotenoid biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular alga, Haematococcus pluvialis. Applied and Environmental Microbiology, 59, 867–873.
Kobayashi, M., Kakizono, T., Nishio, N., Nagai, S., Kurimura, Y., & Tsuji, Y. (1997a). Antioxidant role of astaxanthin in the green alga Haematococcus pluvialis. Applied Microbiology and Biotechnology, 48, 351–356.
Kobayashi, M., Kurimura, Y., & Tsuji, Y. (1997b). Light-independent, astaxanthin production by the green microalga Haematococcus pluvialis under salt stress. Biotechnology Letters, 19, 507–509.
Koller, M., Muhr, A., & Braunegg, G. (2014). Microalgae as versatile cellular factories for valued products. Algal Research, 6, 52–63.
Kwan, T. A., Kwan, S. E., Peccia, J., & Zimmerman, J. B. (2018). Selectively biorefining astaxanthin and triacylglycerol co-products from microalgae with supercritical carbon dioxide extraction. Bioresource Technology, 269, 81–88.
Lababpour, A., Hada, K., Shimahara, K., Katsuda, T., & Katoh, S. (2004). Effects of nutrient supply methods and illumination with blue light emitting diodes (LEDs) on astaxanthin production by Haematococcus pluvialis. Journal of Bioscience and Bioengineering, 98, 452–456.
Lao, Y. M., Jin, H., Zhou, J., Zhang, H. J., & Cai, Z. H. (2017). Functional characterization of a missing branch component in Haematococcus pluvialis for control of algal carotenoid biosynthesis. Frontiers in Plant Science, 8, 1–15.
Lee, C., Woo, J., Jin, A., Kim, B., Young, J., & Yoon, K. (2018). Comparative transcriptome analysis of Haematococcus pluvialis on astaxanthin biosynthesis in response to irradiation with red or blue LED wavelength. World Journal of Microbiology and Biotechnology, 34, 1–14.
Leite, M. F., De Lima, A., Massuyama, M. M., & Otton, R. (2010). In vivo astaxanthin treatment partially prevents antioxidant alterations in dental pulp from alloxan-induced diabetic rats. International Endodontic Journal, 43, 959–967.
Lemoine, Y., & Schoefs, B. (2010). Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynthesis Research, 106, 155–177.
Li, Y., Sommerfeld, M., Chen, F., & Hu, Q. (2008). Consumption of oxygen by astaxanthin biosynthesis: A protective mechanism against oxidative stress in Haematococcus pluvialis (Chlorophyceae). Journal of Plant Physiology, 165, 1783–1797.
Li, Y., Sommerfeld, M., Chen, F., & Hu, Q. (2010). Effect of photon flux densities on regulation of carotenogenesis and cell viability of Haematococcus pluvialis (Chlorophyceae). Journal of Applied Phycology, 22, 253–263.
Li, J., Zhu, D., Niu, J., Shen, S., & Wang, G. (2011). An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnology Advances, 29, 568–574.
Li, M., Wu, W., Zhou, P., Xie, F., Zhou, Q., & Mai, K. (2014). Comparison effect of dietary astaxanthin and Haematococcus pluvialis on growth performance, antioxidant status and immune response of large yellow croaker Pseudosciaena crocea. Aquaculture, 434, 227–232.
Li, K., Cheng, J., Ye, Q., He, Y., Zhou, J., & Cen, K. (2017). In vivo kinetics of lipids and astaxanthin evolution in Haematococcus pluvialis mutant under 15% CO2 using Raman microspectroscopy. Bioresource Technology, 244, 1439–1444.
Liu, J., & Huang, Q. (2016). Screening of astaxanthin-hyperproducing Haematococcus pluvialis using fourier transform infrared (FT-IR) and Raman microspectroscopy. Applied Spectroscopy, 70, 1639–1648.
Liu, Z., Liu, C., Hou, Y., Chen, S., Xiao, D., Zhang, J., & Chen, F. (2013). Isolation and characterization of a marine microalga for biofuel production with astaxanthin as a co-product. Energies, 6, 2759–2772.
Liu, J., Gerken, H., & Li, Y. (2014a). Single-tube colony PCR for DNA amplification and transformant screening of oleaginous microalgae. Journal of Applied Phycology, 26, 1719–1726.
Liu, J., Sun, Z., Gerken, H., Liu, Z., Jiang, Y., & Chen, F. (2014b). Chlorella zofingiensis as an alternative microalgal producer of astaxanthin: Biology and industrial potential. Marine Drugs, 12, 3487–3515.
Liu, Z., Zeng, X., & Cheng, J. (2018). The efficiency and comparison of novel techniques for cell wall disruption in astaxanthin extraction from Haematococcus pluvialis. International Journal of Food Science & Technology, 53, 2212–2219.
Lorenz, R. T. (1999). A technical review of Haematococcus algae (NatuRose™ technical bulletin #060) (pp. 1–12). Kailua-Kona: Cyanotech Corporation.
Lorenz, R. T., & Cysewski, G. R. (2000). Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends in Biotechnology, 18, 160–167.
Łukomska-Kowalczyk, M., Karnkowska, A., Krupska, M., Milanowski, R., & Zakryś, B. (2016). DNA barcoding in autotrophic euglenids: Evaluation of coi and 18s rDNA. Journal of Phycology, 52, 951–960.
Ma, R. Y., & Chen, F. (2001). Enhanced production of free trans -astaxanthin by oxidative stress in the cultures of the green microalga Chlorococcum sp. Process Biochemistry, 36, 1175–1179.
Ma, R., Thomas-Hall, S. R., Chua, E. T., Alsenani, F., Eltanahy, E., Netzel, M. E., Netzel, G., Lu, Y., & Schenk, P. M. (2018). Gene expression profiling of astaxanthin and fatty acid pathways in Haematococcus pluvialis in response to different LED lighting conditions. Bioresource Technology, 250, 591–602.
Manabe, Y., Komatsu, T., Seki, S., & Sugawara, T. (2018). Dietary astaxanthin can accumulate in the brain of rats. Bioscience, Biotechnology, and Biochemistry, 8451, 1–4.
Mann, V., Harker, M., Pecker, I., & Hirschberg, J. (2000). Metabolic engineering of astaxanthin production in tobacco flowers. Nature Biotechnology, 18, 888–892.
Margalith, P. Z. (1999). Production of ketocarotenoids by microalgae. Applied Microbiology and Biotechnology, 51, 431–438.
Market Watch. (2019). Astaxanthin market projected to witness vigorous growth of 800 million by 2024. Retrieved May 10, 2019, from https://www.marketwatch.com/press-release/astaxanthin-market-projected-to-witness-vigorous-growth-of-800-million-by-2024-2019-04-24.
Martínez-delgado, A. A., Khandual, S., Josefina, S., & Rodríguez, V. (2017). Chemical stability of astaxanthin integrated into a food matrix: Effects of food processing and methods for preservation. Food Chemistry, 225, 23–30.
Mazumdar, N., Gopalakrishnan, K. K., Visnovsky, G., & Novis, M. (2018). A novel alpine species of Haematococcus (Chlamydomonadales: Chlorophyta) from New Zealand. New Zealand Journal of Botany, 56, 216.
Mendes-Pinto, M. M., Raposo, M. F. J., Bowen, J., Young, A. J., & Morais, R. (2001). Evaluation of different cell disruption processes on encysted cells of Haematococcus pluvialis: Effects on astaxanthin recovery and implications for bio-availability. Journal of Applied Phycology, 13, 19–24.
Michalak, I., & Chojnacka, K. (2014). Algal extracts: Technology and advances. Engineering in Life Sciences, 14, 581–591.
Miki, W. (1991). Biological functions and activities of animal carotenoids. Pure and Applied Chemistry, 63, 141–146.
Minatelli, J. (2008). Astaxanthin and eye health: The newest carotenoid dietary supplement solution. Nutraceutical Business & Technology, 18–19.
Molino, A., Rimauro, J., Casella, P., Cerbone, A., Larocca, V., Chianese, S., Karatza, D., Mehariya, S., & Ferraro, A. (2018). Extraction of astaxanthin from microalga Haematococcus pluvialis in red phase by using generally recognized as safe solvents and accelerated extraction. Journal of Biotechnology, 283, 51–61.
Mont, L., Juc, A., Pedrosa, C., Seabra, J., Federal, U., Grande, R., & Cordeiro, G. G. (2010). Astaxanthin: Structural and e funcionais. Revista de Nutrição, 23, 1041–1050.
Montsant, A., Zarka, A., & Boussiba, S. (2001). Presence of a nonhydrolyzable biopolymer in the cell wall of vegetative cells and astaxanthin-rich cysts of Haematococcus pluvialis (chlorophyceae). Marine Biotechnology, 3, 515–521.
Mostafa, N., Omar, H., Tan, S. G., & Napis, S. (2011). Studies on the genetic variation of the green unicellular alga Haematococcus pluvialis (chlorophyceae) obtained from different geographical locations using ISSR and RAPD molecular marker. Molecules, 16, 2599–2608.
Nahidian, B., Ghanati, F., Shahbazi, M., & Soltani, N. (2018). Effect of nutrients on the growth and physiological features of newly isolated Haematococcus pluvialis TMU1. Bioresource Technology, 255, 229–237.
Nakada, T., & Ota, S. (2016). What is the correct name for the type of Haematococcus Flot. Taxon, 65, 343–348.
Nawrocki, W. J., Tourasse, N. J., Taly, A., Rappaport, F., & Wollman, F.-A. (2015). The plastid terminal oxidase: Its elusive function points to multiple contributions to plastid physiology. Annual Review of Plant Biology, 66, 49–74.
Newsome, R. (1986). Food colors. Food Technology, 40, 49–56.
Nguyen, K. D. (2013). Astaxanthin: A comparative case of synthetic vs. natural production. Chemical and Biomolecular Engineering Publications and Other Works, 1, 1–9.
Nishida, Y., Yamashita, E., & Miki, W. (2007, September). Comparison of Astaxanthin’s singlet oxygen quenching activity with common fat and water soluble antioxidants. Paper presented at the 21st Annual Meeting on Carotenoid Research held at Osaka, Japan (Vol. 6).
Nobre, B., Marcelo, F., Passos, R., Beirão, L., Palavra, A., Gouveia, L., & Mendes, R. (2006). Supercritical carbon dioxide extraction of astaxanthin and other carotenoids from the microalga Haematococcus pluvialis. European Food Research and Technology, 223, 787–790.
Oilalgae. (2015). Astaxanthin. Retrieved March 30, 2015, from http://www.oilgae.com/non_fuel_products/astaxanthin.html.
Olaizola, M. (2000). Commercial production of astaxanthin from Haematococcus pluvialis using 25,000-liter outdoor photobioreactors. Journal of Applied Phycology, 12, 499–506.
Olaizola, M. (2003). Commercial development of microalgal biotechnology: From the test tube to the marketplace. Biomolecular Engineering, 20, 459.
Olaizola, M., & Huntley, M. E. (2003). Recent advances in commercial production of astaxanthin from microalgae. In Recent advances in marine biotechnology. Volume 9. Biomaterials and bioprocessing (Vol. 9, pp. 143–164). New Hampshire: Science Publishers.
Oncel, S. S., Imamoglu, E., Gunerken, E., & Sukan, F. V. (2011). Comparison of different cultivation modes and light intensities using mono-cultures and co-cultures of Haematococcus pluvialis and Chlorella zofingiensis. Journal of Chemical Technology and Biotechnology, 86, 414–420.
Orosa, M., Valero, J. F., Herrero, C., & Abalde, J. (2001). Comparison of the accumulation of astaxanthin in Haematococcus pluvialis and other green microalgae under N -starvation and high light conditions. Biotechnology Letters, 23, 1079–1085.
Overton, K., Samsing, F., Oppedal, F., Stien, H., & Dempster, T. (2018). Lowering treatment temperature reduces salmon mortality: A new way to treat with hydrogen peroxide in aquaculture. Pest Management Science, 74, 535.
Pan, J. L., Wang, H. M., Chen, C. Y., & Chang, J. S. (2012). Extraction of astaxanthin from Haematococcus pluvialis by supercritical carbon dioxide fluid with ethanol modifier. Engineering in Life Sciences, 12, 638–647.
Pang, N., & Chen, S. (2017). Effects of C5 organic carbon and light on growth and cell activity of Haematococcus pluvialis under mixotrophic conditions. Algal Research, 21, 227–235.
Panis, G., & Rosales, C. J. (2016). Commercial astaxanthin production derived by green alga Haematococcus pluvialis: A microalgae process model and a techno-economic assessment all through production line. Algal Research, 18, 175–190.
Pan-utai, W. (2017). Effect of inducing agents on growth and astaxanthin production in Haematococcus pluvialis: Organic and inorganic. Biocatalysis and Agricultural Biotechnology, 12, 152–158.
Park, J. S., Chyun, J. H., Kim, Y. K., Line, L. L., & Chew, B. P. (2010). Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutrition & Metabolism (London), 7, 18.
Park, J. C., Choi, S. P., Hong, M. E., & Sim, S. J. (2014). Enhanced astaxanthin production from microalga, Haematococcus pluvialis by two-stage perfusion culture with stepwise light irradiation. Bioprocess and Biosystems Engineering, 37, 2039–2047.
Peebles, F. (1909). The life history of Sphaerella lacustris (Haematococcus pluvialis) with reference to the nature and behavior of the zoospores. Zentralblatt fuer Bakteriologie Jena Abt II, 2(24), 511–521.
Peled, E., Leu, S., Zarka, A., Weiss, M., Pick, U., Khozin-Goldberg, I., & Boussiba, S. (2011). Isolation of a novel oil globule protein from the green alga Haematococcus pluvialis (chlorophyceae). Lipids, 46, 851–861.
Peled, E., Pick, U., Zarka, A., Shimoni, E., Leu, S., & Boussiba, S. (2012). Light-induced oil globule migration in Haematococcus pluvialis (Chlorophyceae). Journal of Phycology, 48, 1209–1219.
Pérez-López, P., González-García, S., Jeffryes, C., Agathos, S. N., McHugh, E., Walsh, D., Murray, P., Moane, S., Feijoo, G., & Moreira, M. T. (2014). Life cycle assessment of the production of the red antioxidant carotenoid astaxanthin by microalgae: From lab to pilot scale. Journal of Cleaner Production, 64, 332–344.
Phasex Corporation. (2015). Phasex Corporation. Retrieved October 9, 2015, from http://www.phasex4scf.com/.
Pick, U., Zarka, A., Boussiba, S., & Davidi, L. (2019). A hypothesis about the origin of carotenoid lipid droplets in the green algae Dunaliella and Haematococcus. Planta, 249, 31–47.
Piermarocchi, S., Saviano, S., Parisi, V., Tedeschi, M., Panozzo, G., Scarpa, G., Boschi, G., & Lo Giudice, G. (2012). Carotenoids in Age-related maculopathy Italian study (CARMIS): Two-year results of a randomized study. European Journal of Ophthalmology, 22, 216–225.
Poonkum, W., Powtongsook, S., & Pavasant, P. (2015). Astaxanthin induction in microalga H. pluvialis with flat panel airlift photobioreactors under indoor and outdoor conditions. Preparative Biochemistry & Biotechnology, 45, 1.
Prabhakaran, M., Elumalai, S., Santhose, B. I., & Kanna, G. R. (2014). Collection, isolation and identification of Haematococcus pluvialis Flotow from high altitude region of Pithoragarh district, Uttarakhand, India. Golden Research Thoughts, 3, 1–7.
Praveenkumar, R., Lee, K., Lee, J., & Oh, Y. (2015). Breaking dormancy: An energy-efficient means of recovering astaxanthin from microalgae. Green Chemistry, 17, 1226–1234.
Proctor, V. W. (1957a). Some controlling factors in the distribution of Haematococcus pluvialis. Ecology, 38, 457–462.
Proctor, V. W. (1957b). Preferential assimilation of nitrate ion by Haematococcus pluvialis. American Journal of Botany, 44, 141.
Proctor, V. W. (1957c). Studies of algal antibiosis using Haematococcus and Chlamydomonas. Limnology and Oceanography, 2(2), 125–139.
Qinglin, D., Xueming, Z., Xiangying, X., Jianzhong, H., & Jixian, G. (2007). Concomitant NH4+ secretion during astaxanthin synthesis in Haematococcus pluvialis under high irradiance and nitrogen deficient conditions. Chinese Journal of Chemical Engineering, 15, 162–166.
Ralston, N. V. C., Azenkeng, A., Ralston, C. R., Blackwell, J. L., & Raymond, L. J. (2014). Selenium-health benefit values as seafood safety criteria. In Seafood science: Advances in chemistry, technology and applications (pp. 433–457). Boca Raton, FL: CRC Press.
Ranga Rao, A., Raghunath Reddy, R. L., Baskaran, V., Sarada, R., & Ravishankar, G. A. (2010). Characterization of microalgal carotenoids by mass spectrometry and their bioavailability and antioxidant properties elucidated in rat model. Journal of Agricultural and Food Chemistry, 58, 8553–8559.
Ranjbar, R., Inoue, R., Katsuda, T., Yamaji, H., & Katoh, S. (2008). High efficiency production of astaxanthin in an airlift photobioreactor. Journal of Bioscience and Bioengineering, 106, 204–207.
Rayman, M. P. (2012). Selenium and human health. Lancet, 379, 1256–1268.
Renstrøm, B., & Liaaen-Jensen, S. (1981). Fatty acid composition of some esterified carotenols. Comparative Biochemistry and Physiology B, 69, 625–627.
Riccioni, G., Speranza, L., Pesce, M., Cusenza, S., D’Orazio, N., & Glade, M. J. (2012). Novel phytonutrient contributors to antioxidant protection against cardiovascular disease. Nutrition, 28, 605–610.
Rioboo, C., González-Barreiro, Ó., Abalde, J., & Cid, Á. (2011). Flow cytometric analysis of the encystment process induced by paraquat exposure in Haematococcus pluvialis (Chlorophyceae). European Journal of Phycology, 46, 89–97.
Rodríguez-Sáiz, M., De La Fuente, J. L., & Barredo, J. L. (2010). Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Applied Microbiology and Biotechnology, 88, 645–658.
Saha, S. K., Hayes, J., Moane, S., & Murray, P. (2013a). Tagging of biomolecules with deuterated water (D2O) in commercially important microalgae. Biotechnology Letters, 35, 1067–1072.
Saha, S. K., McHugh, E., Hayes, J., Moane, S., Walsh, D., & Murray, P. (2013b). Effect of various stress-regulatory factors on biomass and lipid production in microalga Haematococcus pluvialis. Bioresource Technology, 128, 118–124.
Sarada, R., Bhattacharya, S., & Ravishankar, G. A. (2002). Optimization of culture conditions for growth of the green alga Haematococcus pluvialis. World Journal of Microbiology and Biotechnology, 18, 517–521.
Sarada, R., Vidhyavathi, R., Usha, D., Ravishankar, G. A., Sarada, R., Vidhyavathi, R., Usha, D., & Ravishankar, G. A. (2006). An efficient method for extraction of astaxanthin from green alga Haematococcus pluvialis. Journal of Agricultural and Food Chemistry, 54, 7585–7588.
Satoh, A., Tsuji, S., Okada, Y., Murakami, N., Urami, M., Nakagawa, K., Ishikura, M., Katagiri, M., Koga, Y., & Shirasawa, T. (2009). Preliminary clinical evaluation of toxicity and efficacy of a new astaxanthin-rich Haematococcus pluvialis extract. Journal of Clinical Biochemistry and Nutrition, 44, 280–284.
Schott. (2019). Schott. Retrieved May 1, 2019, from https://pbr-blog.schott.com/2017/06/06/free-guide-the-five-main-reasons-why-you-should-focus-your-attention-on-selecting-the-right-glass-components-for-your-pbr/.
Shah, M. M. R., Liang, Y., Cheng, J. J., & Daroch, M. (2016). Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Frontiers in Plant Science, 7, 531.
Shah, M. R., Lutzu, G. A., Alam, A., Sarker, P., Kabir Chowdhury, M. A., Parsaeimehr, A., Liang, Y., & Daroch, M. (2018). Microalgae in aquafeeds for a sustainable aquaculture industry. Journal of Applied Phycology, 30, 197–213.
Sharon-Gojman, R., Maimon, E., Leu, S., Zarka, A., & Boussiba, S. (2015). Advanced methods for genetic engineering of H. pluvialis. Algal Research, 10, 8–15.
Sheikhzadeh, N., Panchah, I. K., Asadpour, R., Tayefi-Nasrabadi, H., & Mahmoudi, H. (2012). Effects of Haematococcus pluvialis in maternal diet on reproductive performance and egg quality in rainbow trout (Oncorhynchus mykiss). Animal Reproduction Science, 130, 119–123.
Sipaúba-Tavares, L. H., Berchielli-Morais, F. A., & Scardoeli-Truzzi, B. (2015). Growth of Haematococcus pluvialis flotow in alternative media. Brazilian Journal of Biology, 75, 796–803.
Smith, D. R. (2018). Haematococcus lacustris: The makings of a giant-sized chloroplast genome. AoB Plants, 10, 1–7.
Solovchenko, A., & Chekanov, K. (2014). Production of carotenoids using microalgae cultivated in photobioreactors. In Production of biomass and bioactive compounds using bioreactor technology (pp. 63–91). New York, NY: Springer.
Solovchenko, A., Aflalo, C., Lukyanov, A., & Boussiba, S. (2013). Nondestructive monitoring of carotenogenesis in Haematococcus pluvialis via whole-cell optical density spectra. Applied Microbiology and Biotechnology, 97, 4533–4541.
Sommer, T. R., Potts, W. T., & Morrissy, N. M. (1991). Utilization of microalgal astaxanthin by rainbow trout (Oncorhynchus mykiss). Aquaculture, 94(1), 79.
Spiller, G. A., & Dewell, A. (2003). Safety of an astaxanthin-rich Haematococcus pluvialis algal extract: A randomized clinical trial. Journal of Medicinal Food, 6, 51–56.
Stahl, W., & Sies, H. (2003). Antioxidant activity of carotenoids. Molecular Aspects of Medicine, 24, 345–351.
Steinbrenner, J., & Sandmann, G. (2006). Transformation of the green alga Haematococcus pluvialis with a phytoene desaturase for accelerated astaxanthin biosynthesis. Applied and Environmental Microbiology, 72, 7477–7484.
Strittmatter, M., Guerra, T., Silva, J., & Gachon, C. M. M. (2016). A new flagellated dispersion stage in Paraphysoderma sedebokerense, a pathogen of Haematococcus pluvialis. Journal of Applied Phycology, 27, 1553.
Suh, I. S., Joo, H. N., & Lee, C. G. (2006). A novel double-layered photobioreactor for simultaneous Haematococcus pluvialis cell growth and astaxanthin accumulation. Journal of Biotechnology, 125, 540–546.
Sun, Y., Liu, J., Zhang, X., & Lin, W. (2008). Strain H2-419-4 of Haematococcus pluvialis induced by ethyl methanesulphonate and ultraviolet radiation. Chinese Journal of Oceanology and Limnology, 26, 152–156.
Sun, H., Kong, Q., Geng, Z., Duan, L., Yang, M., & Guan, B. (2015). Enhancement of cell biomass and cell activity of astaxanthin-rich Haematococcus pluvialis. Bioresource Technology, 186, 67–73.
Tanaka, T., Shnimizu, M., & Moriwaki, H. (2012). Cancer chemoprevention by carotenoids. Molecules, 17, 3202–3242.
Tjahjono, A. E., Hayama, Y., Kakizono, T., Terada, Y., Nishio, N., & Nagai, S. (1994). Hyper-accumulation of astaxanthin in a green alga Haematococcus pluvialis at elevated temperatures. Biotechnology Letters, 16, 133–138.
Tocquin, P., Fratamico, A., & Franck, F. (2012). Screening for a low-cost Haematococcus pluvialis medium reveals an unexpected impact of a low N/P ratio on vegetative growth. Journal of Applied Phycology, 24, 365–373.
Tominaga, K., Hongo, N., Karato, M., & Yamashita, E. (2012). Cosmetic benefits of astaxanthin on humans subjects. Acta Biochimica Polonica, 59, 43–47.
Torres-carvajal, L. K., González, Á. D., Francisco, U., & Santander, D. P. (2017). Astaxanthin production from Haematococcus pluvialis: Effects of light wavelength and salinity. Contemporary Engineering Sciences, 10, 1739–1746.
Torzillo, G., Goksan, T., Faraloni, C., Kopecky, J., & Masojidek, J. (2003). Interplay between photochemical activities and pigment composition in an outdoor culture of Haematococcus pluvialis during the shift from the green to red stage. Journal of Applied Phycology, 15, 127–136.
Triki, A., Maillard, P., & Gudin, C. (1997). Gametogenesis in Haematococcus pluvialis Flotow (Volvocales, Chlorophyta). Phycologia, 36, 190–194.
Tripathi, U., Sarada, R., Rao, S. R., & Ravishankar, G. A. (1999). Production of astaxanthin in Haematococcus pluvialis cultured in various media. Bioresource Technology, 68, 197–199.
Tripathi, U., Venkateshwaran, G., Sarada, R., & Ravishankar, G. A. (2001). Studies on Haematococcus pluvialis for improved production of astaxanthin by mutagenesis. World Journal of Microbiology and Biotechnology, 17, 143–148.
Van de Ven, M., & Van de Ven, J. J. M. F. (2009). Photobioreactor with a cleaning system and method for cleaning such a reactor. PCT Patent Application WO2009/051478. Accessed 16 Nov 2019.
Visioli, F., & Artaria, C. (2017). Astaxanthin in cardiovascular health and disease: mechanisms of action, therapeutic merits, and knowledge gaps. Food & Function, 8, 39–63.
Wan, M., Hou, D., Li, Y., Fan, J., Huang, J., Liang, S., Wang, W., Pan, R., Wang, J., & Li, S. (2014a). The effective photoinduction of Haematococcus pluvialis for accumulating astaxanthin with attached cultivation. Bioresource Technology, 163, 26–32.
Wan, M., Zhang, J., Hou, D., Fan, J., Li, Y., Huang, J., & Wang, J. (2014b). The effect of temperature on cell growth and astaxanthin accumulation of Haematococcus pluvialis during a light–dark cyclic cultivation. Bioresource Technology, 167, 276–283.
Wan, M., Zhang, Z., Wang, J., Huang, J., Fan, J., Yu, A., Wang, W., & Li, Y. (2015). Sequential heterotrophy–dilution–photoinduction cultivation of Haematococcus pluvialis for efficient production of astaxanthin. Bioresource Technology, 198, 557–563.
Wang, S. B., Chen, F., Sommerfeld, M., & Hu, Q. (2005). Isolation and proteomic analysis of cell wall-deficient Haematococcus pluvialis mutants. Proteomics, 5, 4839–4851.
Wang, J., Sommerfeld, M., & Hu, Q. (2009). Occurrence and environmental stress responses of two plastid terminal oxidases in Haematococcus pluvialis (Chlorophyceae). Planta, 230, 191–203.
Wang, L., Yang, B., Yan, B., & Yao, X. (2012). Supercritical fluid extraction of astaxanthin from Haematococcus pluvialis and its antioxidant potential in sun flower oil. Innovative Food Science & Emerging Technologies, 13, 120–127.
Wang, J., Han, D., Sommerfeld, M. R., Lu, C., & Hu, Q. (2013). Effect of initial biomass density on growth and astaxanthin production of Haematococcus pluvialis in an outdoor photobioreactor. Journal of Applied Phycology, 25(1), 253.
Wang, B., Zhang, Z., Hu, Q., Sommerfeld, M., Lu, Y., & Han, D. (2014). Cellular capacities for high-light acclimation and changing lipid profiles across life cycle stages of the green alga Haematococcus pluvialis. PLoS One, 9, e106679.
Wang, F., Gao, B., Wu, M., Huang, L., & Zhang, C. (2019). A novel strategy for the hyper-production of astaxanthin from the newly isolated microalga Haematococcus pluvialis JNU35. Algal Research, 39, 101466.
Wayama, M., Ota, S., Matsuura, H., Nango, N., Hirata, A., & Kawano, S. (2013). Three-dimensional ultrastructural study of oil and astaxanthin accumulation during encystment in the green alga Haematococcus pluvialis. PLoS One, 8, e53618.
Yaakob, Z., Ali, E., Zainal, A., Mohamad, M., & Takriff, M. S. (2014). An overview: Biomolecules from microalgae for animal feed and aquaculture. Journal of Biological Research, 21, 1–10.
Yokoyama, A., Izumida, H., & Miki, W. (1994). Production of astaxanthin and 4-ketozeaxanthin by the marine bacterium, agrobacterium aurantiacum. Bioscience, Biotechnology, and Biochemistry, 58, 1842–1844.
Yoo, J. J., Choi, S. P., Kim, B. W., & Sim, S. J. (2012). Optimal design of scalable photo-bioreactor for phototropic culturing of Haematococcus pluvialis. Bioprocess and Biosystems Engineering, 35, 309–315.
Young, A. J., Pritchard, J., White, D., & Davies, S. (2017). Processing of astaxanthin-rich Haematococcus cells for dietary inclusion and optimal pigmentation in Rainbow trout, Oncorhynchus mykiss L. Aquaculture Nutrition, 23, 1304–1311.
Yuan, J.-P., Peng, J., Yin, K., & Wang, J.-H. (2011). Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Molecular Nutrition & Food Research, 55, 150–165.
Zgheib, N., Saade, R., Khallouf, R., & Takache, H. (2018). Extraction of astaxanthin from microalgae: Process design and economic feasibility study. IOP Conference Series Materials Science and Engineering, 323, 1–12.
Zhang, X. W., Gong, X. D., & Chen, F. (1999). Dynamics and stability analysis of the growth and astaxanthin production system of Haematococcus pluvialis. Journal of Industrial Microbiology and Biotechnology, 23(2), 133–137.
Zhang, B. Y., Geng, Y. H., Li, Z. K., Hu, H. J., & Li, Y. G. (2009). Production of astaxanthin from Haematococcus in open pond by two-stage growth one-step process. Aquaculture, 295, 275–281.
Zhang, W., Wang, J., Wang, J., & Liu, T. (2014). Attached cultivation of Haematococcus pluvialis for astaxanthin production. Bioresource Technology, 158, 329–335.
Zhang, C., Zhang, L., & Liu, J. (2016). The role of photorespiration during astaxanthin accumulation in Haematococcus pluvialis (Chlorophyceae). Plant Physiology and Biochemistry, 107, 75–81.
Zheng, Y., Li, Z., Tao, M., Li, J., & Hu, Z. (2017). Effects of selenite on green microalga Haematococcus pluvialis: Bioaccumulation of selenium and enhancement of astaxanthin production. Aquatic Toxicology, 183, 21–27.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Butler, T., Golan, Y. (2020). Astaxanthin Production from Microalgae. In: Alam, M., Xu, JL., Wang, Z. (eds) Microalgae Biotechnology for Food, Health and High Value Products. Springer, Singapore. https://doi.org/10.1007/978-981-15-0169-2_6
Download citation
DOI: https://doi.org/10.1007/978-981-15-0169-2_6
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-15-0168-5
Online ISBN: 978-981-15-0169-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)