
Abstract
This chapter describes the basic principle of force spectroscopy based on atomic force microscopy, with particular attention to instrumental and applications aspects more strictly related to the study of single biomolecules and cell membrane.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


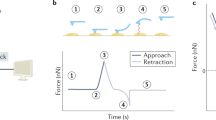
1 Introduction
1.1 AFM (Atomic Force Microscopy)
Invented by IBM scientists in 1982, Atomic force microscopy (AFM) was experimental implemented by Binning, Quate and Gerber in 1986. It is a type of scanning probe microscopy (SPM), with demonstrated resolution on the order of nanometers, more than 1000 times better than the optical diffraction limit. The AFM consists of a cantilever with a sharp tip at its end that is used to scan the sample surface. The cantilevers is typically made up of silicon or silicon nitride with the tip apex radius on the order of nanometers. When the tip was brought into proximity of the sample surface, force between the tip apex and the sample surface lead to a deflection of the cantilever. And the deflection was monitored by the quadrant photodiode then “tell” the piezo scanner to up or down to follow the topography of the sample surface. Normally, three different operation modes were used to scan the surface of the sample, according to the motion of the tip: contact mode; intermittent mode (AC mode or tapping mode); non-contact mode. Let’s follow the history to introduce these modes. When AFM was invented, contact mode was used to scan sample. In contact mode, the tip is “dragged” across the surface of the sample and the contours of the surface is measured either using the deflection of the cantilever directly or, more commonly, using the feedback signal which used to keep the piezo scanner follow the surface. In order to decrease the damage and get enough vertical deflection of the tip, low spring constant cantilevers are used for contact mode. As the application area expends, people found that the contact mode will created large later force and damage to the soft sample, like bio-sample. Then intermittent mode (AC, tapping) and non-contact mode was invented to scan the soft sample surface. Due to very hard operation of non-contact mode exceptionally in ambient conditions, intermittent mode is normally used to scan the surface. Because in ambient conditions, most samples develop a liquid meniscus layer, it is very hard to keep the tip above the surface without “snap” into the surface and scan the surface. AC mode can bypass this problem by intermittent contact the surface with oscillation near its resonance frequency. The oscillation of the tip was achieved with small piezo element in the Z scanner, but other possibilities include an AC magnetic field or periodic heating with a modulated laser beam. The amplitude of the oscillation usually varies from below one nanometer to several hundred nanometers. In AC mode, frequency and amplitude of the driving signal are kept constant, leading to constant amplitude of the cantilever oscillation as long as there is no drift or interaction between tip and sample surface. The interaction of forces acting on the cantilever when the tip approach to the surface and decrease the amplitude and change the phase of the oscillation of the tip. This amplitude is used as the parameter that goes into the electronic servo that control the height to maintain a set cantilever oscillation amplitude as the cantilever is scanned over the sample surface. Although the peak force applied during the contacting part of the oscillation can be much higher than typically during contact mode, AC mode generally lessens the damage done to the sample surface and the tip compare done in contact mode. But AC mode is gentle enough ever for the lipid bilayers or adsorbed single polymer molecules under liquid medium. Beside the image, the phase of the cantilever’s oscillation with respect to the driving signal can be recorded and analyzed. The phase signal contains information about the energy dissipated by the cantilever in each oscillation cycle. As the development of the life science technology, another image scanning technics (QI™) was invented to get the topography and stiffness of sample at the same time. QI™ (Quantitative imaging, JPK Instrument, AG) is one of the best AFM imaging technics used for life bio-samples. With QI- a force curve based image mode- the user has full control over the tip-sample interacting force at every pixel of the image. There is no longer for setpoint or gain adjustment while scanning anymore, and every raw point data of image are saved for further analysis.
AFM was introduced into three major application areas: force measurement, imaging and manipulation. In force measurement, AFM was used to detect the force between the tip and the sample surface as a function of their mutual separation according to Hooke’s law. For imaging, keeping the force between the tip and sample as constant, the probe can follow the topography and give the three dimensional shape with high resolution. In manipulation, the forces between tip and sample can be used to change the properties of the sample in a controlled way, which means that the features of the sample can be removed or changed as we want. Although, image scanning is the most wide use of AFM. Molecular interactions is the most important subject people use AFM to work on.
1.2 The Importance of Biomolecular Interactions
Because of biomolecular interactions playing important roles in many biological and physiological processes, it has been recognized that the analysis of single biomolecules spectroscopy is of critical importance in fundamental genetic research, clinical analysis, and industrial biotechnological development [1, 2]. Interaction processes between macromolecules on biological surfaces are essential for diverse cellular functions including embryonic development, signal transduction, immune response, cell adhesion, and tissue assembly [3]. Atomic force microscopy (AFM) can provide force-distance curves while scanning the surface of samples. According to the force-distance relationship, we can get data of adhesion and bonding force of mechanical properties [4]. AFM takes unique advantages in isolation of single biomolecules [5] for following reasons: First, force spectroscopy ensures a dynamic analytical method to measurement of interaction forces between single molecules superior to conventional ensemble measurements [6]. Second, AFM has potential to distinguish the target molecules from a complex sample because of evaluating the morphology, mechanical properties, charge, and even chemistry of molecular species by various AFM imaging modes. Moreover, isolation of biomolecules can be done on a surface in ambient conditions by AFM. It enables us to improve the performance in solutions where contaminations are likely to be introduced.
As the base of many biochemical processes, specific molecular interactions involve several types of noncovalent bonds including Van der Waals forces, hydrogen bonds, hydrophobic and electrostatic forces. The highly specific interactions between a ligand and its receptor can be recorded by atomic force microscopy force spectroscopy with a functionalized AFM tip to avoid the detection of non-specific events during the retraction of the tip from the sample surface. These experiments are so called Atomic Force Microscopy-based Single Molecule Force Spectroscopy (AFM-SMFS), since they allow measuring specific interaction forces between even one molecule on the tip and another molecule at the surface of the sample [7].
The unique advantages of AFM including piconewton sensitivity, working under near physiological environments and lateral atomic resolution, making it an excellent approach for biomolecules, especially for soft cell surfaces at single molecule level [8]. For the biological applications, AFM-SMFS is mainly used for measuring protein and glycan on the surface of cells [9]. This chapter mainly reviews single molecule force spectroscopy for biological application, especially on cell membrane. Also, some works on the most advanced front of application will be discussed.
2 Force Spectroscopy of Proteins on Cell Surface
Membrane proteins, mostly serve as membrane receptor in many cell functions [10]. As the barrier of massive pharmacological therapies, the target cell membranes can be studied on delivery system of drugs. Peetla et al. mentioned AFM as a benchmark approach to screen drug candidates [11]. To increase the efficiency of therapeutic discovery and delivery, one should understand the therapeutic-effector interactions and their cell and tissue responses at the molecular level [12].
As the first identified aquaporin of membrane water channels found in all forms of life, Aquaporin-1 (AQP1) is studied for novel drug targets by AFM. Fotiadis D. et al. reveals that AQP1 attributes to a possible secondary function as a cyclic guanosine monophosphate (cGMP) gated ion channel [13]. A striking degree of sequence homology suggests the existence of a Ca2+-binding proteins belonging to Ca2+-binding site at the C terminus of AQP1 instead of the putative cGMP-binding site reported previously. To locate on the acquired surface topographies, 2D crystals AQP1 were treated with carboxypeptidase Y, which cleaves off the intracellular C terminus. Difference maps of AFM topography between the native AQP1 and the carboxypeptidase Y treated AQP1s showed the closed four-fold symmetry axis of the tetramer at carboxylic tail. The unique ability of AFM to provide structural information about molecules on the single molecule level enables thorough study of their behavior at simulated physiological conditions.

Human aquaporin-1 (AQP1) AFM imaging. (a) deglycosylated hAQP1 2D crystals. (b) carboxypeptidase Y treated AQP1 2D crystals. (c) The symmetrized correlation average of the deglycosylated hAQP1, windmill-shaped peripheral (1) and central protrusions (2) on the other side. (d) the symmetrized correlation average of the deglycosylated and decarboxylated hAQP1. On the lower side, the central protrusion, initially present, has disappeared while the peripheral one (1) does not seem to be affected by the digestion. (e) Differences in this difference map between the undigested and digested hAQP1 topographs are evident. (Figure reproduced from [13])
Established AFM-SMFS is widely applied to characterize the molecule affinity between antibody and antigen [14]. Hinterdorfer, P et al. studied on the interaction between protein antibody and antigen, and improve the fixation method of protein by connecting protein with a chain of PEG polymer, as an approach to distinguish the specific interaction between antibody and substrate on force-distance curves [15]. They further modified the tip with antibody to probe the sample, and developed a recognition imaging technology that combines the AFM measuring force mode and the scanning imaging, which ensures AFM a certain ability to identify the types and components of the imaging molecules [16].
Macromolecules on cell surface play an important role as the interface link between the cytoplasm and the extracellular region, such as receptors and ion channels. Vascular endothelial growth factor receptor (VEGFR) is an essential transmembrane receptor protein expressed on vascular endothelial cell surfaces [17]. N. Almqvist et al. initially measured the adhesion forces between isolated VEGFR and an AFM tip modified with its antibody [18]. In this research, the maps of interaction forces between antibody conjugated atomic force microscope tips, an anti-Flk-1-antibody-functionalized tip and a specific receptor, a vascular endothelial growth factor (VEGF) receptor reveals the effect of plasma membrane receptor clustering on local cell mechanics in semiquantitative measurements of binding (or unbinding) forces. Depending on the unique advantages of AFM-FS, distribution and density of VEGFRs were mapped in the cell plasma membrane, which shows that antibody and ligand induced real-time clustering and redistribution of VEGFRs. Later, Lee et al. developed a two-step procedure for VEGFRs surface force analysis: first locating the VEGFRs by dynamic recognition imaging, then conducting AFM-FS on the recognition sites to measure the rupture force and unbinding times [19].

Endothelial cells in real-time atomic force maps. (a–c) Specific interaction between an endothelial cell surface and a Si3N4 tip functionalized with anti-Flk-1. (a) Force map at beginning. (b) Force curves taken at various points on the cell surface from the map shown in (a). The curves are offset with respect to zero force. (c) The unbinding forces probability histogram of the force curves from the force map, fitted with a Gaussian and the corresponding maxima and s is indicated in the figure. The dominant unbinding force (60–70 pN) suggests breakage of single receptor-antibody bonds. (d–f) Competitive inhibition probed with the antibody tip at 10 min after adding antibody in the recording medium. The panels correspond to (a–c). The characteristic unbinding force is suppressed and the measured forces are shifted toward higher values. (g–i) Corresponds to (a–c), but at 45 min after adding antibody. The single brighter spots in (g) are identified as clusters of receptor. Numbers 1–4 refer to a few of these clusters. (Figure reproduced from [18])
Due to the tractability of genetic and biochemical manipulation, the fission yeast Schizosaccharomyces pombe (S. pombe) has become a popular tool for analyzing heterologous G protein coupled receptors (GPCRs) [20]. P-factor is a peptide pheromone and Mam2 is a fission yeast pheromone receptor belonging to GPCRs on the cell surface of the fission yeast. Their interaction was examined by AFM in Sasuga et al.’s work [21]. The AFM tips were modified with P-factor derivatives and truncated P-factor derivative lacking specific binding sites to measure the specific interaction forces, which were recorded to be around 120 pN at a probe speed of 1.74 μm/s. As expected, the truncated P-factor derivative modified tips do not show a force jump in the force curves. This yeast based AFM method shows that this approach has the potential for the signaling study of heterologous GPCRs and screening of their ligands.
Als3 protein is an adhesin involved in adhesion during host invasion, while fluorescence image of Als3 proteins in the cell wall lacks spatial resolution and does not show the biophysical property information, such as elasticity and extension. Beaussart et al. therefore used AFM with anti-Als3 antibodies functionalized tips to get Als3 cell surface force spectroscopy [22]. The morphogenesis of C. albicans from yeast to hyphae was observed. In the nongerminating yeast form, adhesion maps show low rates of Als3. While Als3 Proteins on Hyphae show differences including a higher detection frequency and more specifically on the germ tube.

AFM single molecular mapping of Als3 protein on cell membrane. (a) Schematic representation of interactions between proteins and anti-Als3 antibody modified AFM tip. (b) Adhesion force histogram (1 μm × 1 μm, color scale: 300 pN) of Als3 proteins on nongerminating yeast cell. (c) Adhesion force histogram on germinating yeast corresponds to (b). (d) Adhesion force histogram of a germ tube. (Figure reproduced from [22])
Formosa et al. used AFM as an imaging tool to visualize and localize adhesins nanodomains at the surface of living wild-type Candida albicans cells [23]. They observed that the molecules at the origin of these adhesions could aggregate into nanodomains. Further by the use of functionalized with dendrimers presenting CHO functions able to covalently link with NH2 functions of proteins, they determined that the less adhesive nanodomains were formed by mannoproteins that can interact specifically with Concanavalin A, and force measurements showed that Als proteins (probably among others) were participating to these nanodomains. This study suggested the ability of AFM force spectroscopy to measure the nanoscale size domains at the surface of living cells.
In a very recent research of Santos’s group, AFM has been used as a tool to evaluate the risk of cardiovascular diseases in patients [24]. After pretreatment, the AFM tips were functionalized with human purified fibrinogen to detect the fibrinogen-erythrocyte binding forces. Chronic heart failure (CHF) patients showed altered cell stiffness. Compared with non-ischaemic patients, the fibrinogen-erythrocyte binding forces of ischaemic patients also increased. This experiment showed the potential of AFM-single molecule force spectroscopy for identifying patients with increased risk for cardiovascular diseases.

AFM-based force spectroscopy set-up of fibrinogen-erythrocyte binding. (a) Schematic representation of erythrocytes deposited on a glass slide and fibrinogen molecules mounted on an AFM tip. (b) Example of a force mapping height image of erythrocytes (20 μm × 20 μm, height color scale:1.2 μm). The symbols indicate the surface locations chosen on each cell to perform force spectroscopy measurements. (Figure reproduced from [24])
3 Protein AFM–SMFS with DNA Aptamer Tips
AFM-based single molecule force spectroscopy has the potential for drug design due to its molecular recognition between DNA aptamers and drug molecules. Compared to antibodies, aptammers show higher chemical stability and are easier to manipulate [25].
One of the typical examples is to measue single molecule binding via rupture forces of a split aptamer. Nguyen et al. used a split aptamer by AFM to detect adenosine monophosphate (AMP) [26]. For their atomic force spectroscopy, the sequence of aptamer was split into two parts, oligo a and oligo b, immobilized on tip and gold substrate respectively. In this way, they avoided the target analyte interacting with surface. They found that the rupture force between two oligos were increased in the presence AMP in comparison with the one measured in absence of AMP. Thus, changes in the rupture force could be directly attributed to the specific binding of AMP to the split aptamer.
Protein tyrosine kinase 7 (PTK7), as a protein receptor in tyrosine kinase family, is a surface biomarker member with potential clinical utility in cancer [27]. Aptamers could be a useful bio-probes to detect specific biomarkers on various cell membranes with functionalized AFM tips. To figure out whether synthetic aptamers bind as robustly as natural antibodies, O’Donoghue et al. measured the rupture force between PTK7 and both a natural antibody and a synthetic DNA aptamer by single-molecule AFM technique [28]. The rupture force on live cell membranes between the aptamer and PTK7 was 46 ± 26 pN, while the force with the antibody was 68 ± 33 pN, suggesting that the binding forces are very similar. This study showed that DNA aptamer has the potential for targeted drug delivery to tumors.
Recently, Li et al. demonstrate a simple force-based label-free strategy for the highly sensitive sensing of adenosine [29]. By binding an adenosine ssDNA aptamer onto an AFM probe, this sensor took advantage of the specific molecular recognition between adenosine and an appropriate DNA aptamer, and the recognition resulted in the formation of a folded, hairpin-like DNA structure. This adenosine/aptamer complex showed a variation of the adsorption force at the graphite/water interface. Because of various aptamers reported and methods for discovering new aptamers, this AFM-based single molecule force spectroscopy is simple but promising for the detection of numerous analytes.
4 AFM-SMFS of Cell Surface Glycans
Glycans can be found attached to proteins in glycoproteins and proteoglycans. In general, they are found on the exterior surface of cells. Cell surface glycans, involved in various biological events including immune recognition, cell adhesion, cell migration, bacterial infections, inflammation and cancer, have been well studied in the field of cellular physiology and pathology [30]. Lectins are carbohydrate-binding proteins of macromolecules that are highly specific for sugar moieties. With a specific affinity to glycans, their interactions have been applied into detecting specific glycans on cell surfaces. Several works have been focused on the interactions between lectins and glycans by using AFM-FS. In AFM-based single molecule force spectroscopy, lectins act as functional tips with specificity and high efficiency.
For example, Sletmoen et al. used soybean agglutinin (SBA) as a molecular probe in the structural analysis of glycoproteins in their research [31]. In this study, dynamic force spectroscopy is used to measure the unbinding forces between SBA on a mica surface and a modified porcine submaxillary mucin (Tn-PSM) on AFM tip. A number of force jumps, which consistent with the structure of the carbohydrate chains of mucin, demonstrate that unbinding events or rebinding events were observed up to a distance equal to the length of the mucin chain. These results combined with the long lifetime of the SBA-TnPSM complex can fit with a binding model in which lectin molecules tend to “bind and jump” from a glycan residue to another along the polypeptide chain of mucin before dissociating. In another report, Ricinus communis agglutinin-120 (RCA120) was used as a probe to directly measure the interaction forces between RCA120 and galactosyl residues on living HeLa cell surfaces in Li et al.’s research [32]. RCA120, a lectin from Ricinus communis, possesses a peculiar affinity for terminal β-D-galactosyl residues, hence it has been developed as a functionalized tip to detect the unbinding force, which was observed to be 43 pN at loading rate of 0.4 Nn·s−1. This research successfully applied molecular recognition force spectroscopy to study the specificity of the lectin-carbohydrate interactions.
In a very recent research, Gunning et al. used AFM to unravel the mechanism driving mucus-binding protein(MUB) mediated adhesion to mucins [33]. In this study, MUB was immobilized on the functionalized tips and mucin was immobilized on the glass slide. By single molecule force spectroscopy, they showed a nanospring-like adhesion model between MUB and mucin mediated by unfolding of the multiple repeats constituting the adhesin. The direct evidence for MUB self-interaction was in marked contrast with the mucin adhesion behavior presented by Galectin-3, a mammalian lectin characterized by a single carbohydrate binding domain.
5 Perspective
As shown, AFM-SMFS could be applied in vast cell surface measurements. Its high resolution at the nanoscale size and ability to measure forces in the pN range make it a valuable tool for analysis in biological applications [34]. On the cell surfaces, the AFM-based single molecule force spectroscopy can not only measure interaction forces between various biomolecules, but also mapping recognition sites on cell surfaces directly. Knowledge of these forces contribute to a better understanding of the molecular basis of molecular recognition events. Published data shows that AFM-SMFS could be applied into various areas such as bimolecular recognitions, action mechanisms detection, disease evaluation and drug design [35,36,37]. The amount of newly published work with AFM-SMFS is astonishing and heralds a bright future for AFM research in this area.
References
Hinterdorfer P, Dufrene YF. Detection and localization of single molecular recognition events using atomic force microscopy. Nat Methods. 2006;3:347–55.
Branton D, Deamer DW, Marziali A, Bayley H, Benner SA, Butler T, Di Ventra M, Garaj S, Hibbs A, Huang XH, Jovanovich SB, Krstic PS, Lindsay S, Ling XSS, Mastrangelo CH, Meller A, Oliver JS, Pershin YV, Ramsey JM, Riehn R, Soni GV, Tabard-Cossa V, Wanunu M, Wiggin M, Schloss JA. The potential and challenges of nanopore sequencing. Nat Biotechnol. 2008;26:1146–53.
Wang CZ, Yadavalli VK. Investigating biomolecular recognition at the cell surface using atomic force microscopy. Micron. 2014;60:5–17.
Green NH, Allen S, Davies MC, Roberts CJ, Tendler SJB, Williams PM. Trac-Trends Anal Chem. 2002;21:64–73.
Binnig G, Quate CF, Gerber C. Atomic Force Microscope. Phys Rev Lett. 1986;56:930–3.
Hoffmann T, Dougan L. Single molecule force spectroscopy using polyproteins. Chem Soc Rev. 2012;41:4781–96.
Pillet F, Chopinet L, Formosa C, Dague E. Atomic Force Microscopy and pharmacology: From microbiology to cancerology. Biochim Biophys Acta-Gen Subj. 2014;1840:1028–50.
Engel A, Muller DJ. Observing single biomolecules at work with the atomic force microscope. Nat Struct Biol. 2000;7:715–8.
Li Q, Zhang T, Pan YG, Ciacchi LC, Xu BQ, Wei G. AFM-based force spectroscopy for bioimaging and biosensing. RSC Adv. 2016;6:12893–912.
Dirienzo JM, Nakamura K, Inouye M. The outer membrane proteins of Gram-Negative bacteria: biosynthesis, assembly, and functions. Annu Rev Biochem. 1978;47:481–532.
Peetla C, Rao KS, Labhasetwar V. Relevance of biophysical interactions of nanoparticles with a model membrane in predicting cellular pptake: study with TAT peptide-conjugated nanoparticles. Mol Pharm. 2009;6:1311–20.
La R, Arnsdorf MF. Life Sci. 2010;86:545–62.
Fotiadis D, Suda K, Tittmann P, Jeno P, Philippsen A, Muller DJ, Gross H, Engel A. Identification and structure of a putative Ca2+-binding domain at the C terminus of AQP1. J Mol Biol. 2002;318:1381–94.
Maver U, Velnar T, Gaberscek M, Planinsek O, Finsgar M. Recent progressive use of atomic force microscopy in biomedical applications. Trac-Trends Anal Chem. 2016;80:96–111.
Hinterdorfer P, Baumgartner W, Gruber HJ, Schilcher K, Schindler H. Detection and localization of individual antibody-antigen recognition events by atomic force microscopy. Proc Natl Acad Sci U S A. 1996;93:3477–81.
Kienberger F, Ebner A, Gruber HJ, Hinterdorfer P. Molecular recognition imaging and force spectroscopy of single biomolecules. Accounts Chem Res. 2006;39:29–36.
Olsson AK, Dimberg A, Kreuger J, Claesson-Welsh L. VEGF receptor signalling ? in control of vascular function. Nat Rev Mol Cell Biol. 2006;7:359–71.
Almqvist N, Bhatia R, Primbs G, Desai N, Banerjee S, Lal R. Elasticity and adhesion force mapping reveals real-time clustering of growth factor receptors and associated changes in local cellular rheological properties. Biophys J. 2004;86:1753–62.
Lee S, Mandic J, Van Vliet KJ. Chemomechanical mapping of ligand-receptor binding kinetics on cells. Proc Natl Acad Sci U S A. 2007;104:9609–14.
Kohl JV, Atzmueller M, Fink B, Grammer K. Human pheromones: integrating neuroendocrinology and ethology. Neuroendocrinol Lett. 2001;22:309–21.
Sasuga S, Abe R, Nikaido O, Kiyosaki S, Sekiguchi H, Ikai A, Osada T. J Biomed Biotechnol. 2012:7.
Beaussart A, Alsteens D, El-Kirat-Chatel S, Lipke PN, Kucharikova S, Van Dijck P, Dufrene YF. Single-molecule imaging and functional analysis of Als Adhesins and Mannans during Candida albicans morphogenesis. ACS Nano. 2012;6:10950–64.
Formosa C, Schiavone M, Boisrame A, Richard ML, Duval RE, Dague E. Multiparametric imaging of adhesive nanodomains at the surface of Candida albicans by atomic force microscopy. Nanomed-Nanotechnol Biol Med. 2015;11:57–65.
Guedes AF, Carvalho FA, Malho I, Lousada N, Sargento L, Santos NC. Nat Nanotechnol. 2016;11 687−+
You KM, Lee SH, Im A, Lee SB. Aptamers as functional nucleic acids: In vitro selection and biotechnological applications. Biotechnol Bioprocess Eng. 2003;8:64–75.
Nguyen TH, Steinbock LJ, Butt HJ, Helm M, Berger R. Measuring single small molecule binding via rupture forces of a split aptamer. J Am Chem Soc. 2011;133:2025–7.
Lewis DB, Haines C, Ross D. Protein tyrosine kinase 7: a novel surface marker for human recent thymic emigrants with potential clinical utility. J Perinatol. 2011;31:S72–81.
O’Donoghue MB, Shi XL, Fang XH, Tan WH. Single-molecule atomic force microscopy on live cells compares aptamer and antibody rupture forces. Anal Bioanal Chem. 2012;402:3205–9.
Li J, Li Q, Ciacchi LC, Wei G. Label-free sensing of adenosine based on force variations induced by molecular recognition. Biosensors. 2015;5:85–97.
Pilobello KT, Mahal LK. Deciphering the glycocode: the complexity and analytical challenge of glycomics. Curr Opin Chem Biol. 2007;11:300–5.
Sletmoen M, Dam TK, Gerken TA, Stokke BT, Brewer CF. Single-molecule pair studies of the interactions of the α-GalNAc (Tn-antigen) form of porcine submaxillary mucin with soybean agglutinin. Biopolymers. 2009;91:719–28.
Li YJ, Wang JN, Xing CY, Wang ZX, Wang HD, Zhang BL, Tang JL. Molecular recognition force spectroscopy study of the specific Lectin and Carbohydrate interaction in a living cell. ChemPhysChem. 2011;12:909–12.
Gunning AP, Kavanaugh D, Thursby E, Etzold S, MacKenzie DA, Juge N. Int J Mol Sci. 2016;17
Carvalho FA, Santos NC. Atomic force microscopy-based force spectroscopy biological and biomedical applications. IUBMB Life. 2012;64:465–72.
Dharap SS, Wang Y, Chandna P, Khandare JJ, Qiu B, Gunaseelan S, Sinko PJ, Stein S, Farmanfarmaian A, Minko T. Tumor-specific targeting of an anticancer drug delivery system by LHRH peptide. Proc Natl Acad Sci U S A. 2005;102:12962–7.
Laidler P, Dulinska J, Lekka M, Lekki J. Expression of prostate specific membrane antigen in androgen-independent prostate cancer cell line PC-3. Arch Biochem Biophys. 2005;435:1–14.
Winiarska M, Bil J, Wilczek E, Wilczynski GM, Lekka M, Engelberts PJ, Mackus WJM, Gorska E, Bojarski L, Stoklosa T, Nowis D, Kurzaj Z, Makowski M, Glodkowska E, Issat T, Mrowka P, Lasek W, Dabrowska-Iwanicka A, Basak GW, Wasik M, Warzocha K, Sinski M, Gaciong Z, Jakobisiak M, Parren PWHI, Golab J. PLoS Med. 2008:5.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Tang, C., Fan, Y., Lü, J. (2018). Atomic Force Microscopy-Based Single Molecule Force Spectroscopy for Biological Application. In: Cai, J. (eds) Atomic Force Microscopy in Molecular and Cell Biology. Springer, Singapore. https://doi.org/10.1007/978-981-13-1510-7_2
Download citation
DOI: https://doi.org/10.1007/978-981-13-1510-7_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-13-1509-1
Online ISBN: 978-981-13-1510-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)