
Abstract
Marine biotechnology still remains a new and emergent science, which is closely linked to marine biodiversity and to the technological capacities of investigating more atypical ecosystems. Marine microorganisms show a unique biodiversity since they have to adapt to various marine environmental conditions such as low or high temperatures, alkaline or acidic water, high pressure, and limited nutrients. Marine natural products, especially marine polysaccharides, are attracting more and more attention. Microbial polysaccharides are of growing interest for many sectors of industry, resulting in isolation of new exopolysaccharide (GlossaryTerm
EPS
)-producing bacteria. The diversity of these polysaccharides arises from the structural variations (glycosidic bonds, side branching chains, monosaccharidic content) controlled through a genetic basis. A lower molecular weight and functionalized derivatives together with the native form of the polysaccharide have been shown to possess a variety of biotechnological activities. Therefore, the biophysical and biological properties have made them useful in many pharmaceutical, food, and industrial applications. This chapter gives information on GlossaryTermEPS
-producing bacteria from the marine environment, as well as on the carbohydrate molecule they produce, including the chemical composition or structure when available, the putative pathways of biosynthesis, and the potential applications in industry with a focus on healthcare and glycosaminoglycan-like compounds.An erratum to this chapter is available at http://dx.doi.org/10.1007/978-3-642-53971-8_66
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Hyaluronic Acid
- Lactic Acid Bacterium
- Chondroitin Sulfate
- Marine Bacterium
- Geobacillus Thermodenitrificans
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 In Search of New Polysaccharides
Polysaccharides constitute the most abundant and the most diverse materials found on earth and in the oceans. Polysaccharides (GlossaryTerm
PS
s) form a class of biotechnological molecules that find applications in many industrial fields. They consist of monosaccharides (sugar units) that are linked to each other, generally in a repeated sequence order. By composition, GlossaryTermPS
s may be divided into homopolysaccharides, composed of single monosaccharide type and heteropolysaccharides with several different monosaccharides. Interest in carbohydrates was first very low due to their extreme structural complexity and the lack of understanding of their key role in different biological systems or processes. The type of glycosidic bond, the chain length, the degree and type of branching, as well as the presence of organic or chemical groups are important features of the molecular structure, which are also of great importance for the biophysical or biological properties. Except for the GlossaryTermPS
used traditionally in the food industry, the structure of most GlossaryTermPS
, as well as the structure–function relationships are mostly unknown.For biotechnological purposes, marine GlossaryTerm
PS
s have been studied in animals (chitin from crustaceans, chondroitin sulfate from fish, sulfated GlossaryTermPS
s from sponge and glycosaminoglycans from scallops and abalone), plants (ulvans, agarans, carrageenans, alginates, fucans, laminarin from seaweeds), and microorganisms such as fungi, microalgae, and bacteria. In bacteria, several kinds of GlossaryTermPS
exist, depending on their cellular localization and structure. Exopolysaccharides (GlossaryTermEPS
s) represent an important component of the biofilm matrix and serve as functional elements for adhesion, protection, and recognition. They also have chelating properties especially of heavy metals. The main advantages of microbial GlossaryTermEPS
s over plants or macroalgal and animal GlossaryTermPS
s lie on a stable supply independently of climatic or physiologic variations. Moreover, new structures have been described in bacteria, in particular in marine bacteria, giving the opportunity for the development of new applications, especially in the pharmaceutical field.Marine-derived GlossaryTerm
PS
s and their oligosaccharidic derivatives have been studied for a variety of biological activities such as antithrombotic, antitumor, antiviral, antioxidant, and immunomodulatory effects. A high specificity linked with highly beneficial properties compared with the reference molecules already existing on the market are the major conditions of the development of new molecules, especially if these molecules present a cheaper production cost.There is in particular a big diversity in the abundant microbial life in the oceans. The marine environments include coastal ecosystems, the deep ocean with hydrothermal vents, sediments, and the cold water of the poles. The extremes of temperature, pressure, and acidity met in the ocean, and the availability of the sources of energy and nutrients results in different strategies of microorganisms to survive. The combination of the diversity, the quantity, and the rate of metabolism makes the oceanic world a vital for the earth. Indeed, marine microbes are responsible for the global cycle of nutriments and other elements and are involved in the chemistry of the ocean, the composition of the atmosphere, and climate. As microorganisms quickly reproduce when conditions are favorable, the huge number of generations allows, by selection and natural evolution, the appearance of new species or other groups of the classification. The microbial genetic diversity also results in a biochemical diversity offering opportunities of biotechnological and pharmaceutical applications.
This chapter will focus on GlossaryTerm
PS
from bacterial microorganisms, presenting an overview of microbial marine biodiversity, structures of the produced polymers, some molecular aspects of their biosynthesis, as well as some of their applications driven by their features and biological properties with a focus on the human health domain.2 Marine Biodiversity
For a long time, oceans have been considered as unlimited to human beings, who have always wanted to exploit the attractive resources. However, the industrial exploitation of the oceans only began 100 years ago. Studies to understand oceans and their ecosystems with the aim to exploit molecules produced by marine organisms or to develop new drugs inspired by marine molecules have started to reveal secrets, new biodiversity, and innovative molecules of biotechnological interest.
2.1 Ecosystems in the Oceans
The oceans cover of the earth’s surface, with an average depth of . Depths reach for the abyssal plains; the deepest point is the Mariana Trench in the Pacific Ocean (estimated to be ). Smaller seas of salt water also exist; they are partly or fully enclosed by land.
Except on the surface and in temperate or tropical areas, most of seawater is characterized by a temperature lower than , low nutrients, and no sunlight to allow photosynthesis. The open ocean is a huge ecosystem but particular habitats sometimes qualified as extreme have also been discovered: underwater salt lakes, volcanos, mountains, hot smokers, cold seeps, …
In the deep sea, pressure is very high, the temperature is between and , and there is total darkness. A remarkable variety of deep-sea habitats exist: cold seeping waters, sinking particles, sediments, animal guts and surfaces, deep sediments, and hydrothermal vents [1]. These latter areas are unstable due to high volcanic activity; they are characterized by a large gradient of temperatures [2], high amounts of dissolved compounds, gas, as well as very low pH values. The first deep-sea hydrothermal vent was discovered in 1977, near the Galapagos islands, on the East Pacific Rise, and the first black smoker in 1979 on the East Pacific Rise, North. Deep-sea hydrothermal vents are mainly located on the active Mid-Atlantic Ridge, the East Pacific Rise, and back-arc basins (Fiji, Okinawa, Lau Basin). They come from the seeping of seawater down into the crust through rock cracks in the seafloor. During course of the water, several chemical processes enrich it in minerals, such as calcium, sulfates, magnesium, sodium, hydrogen sulphide, and metals, and it also becomes hotter. Jetting upwards by chimneys called black or white smokers formed by compound deposits or diffusing out of cracks, the hydrothermal fluid gives rise to the development of a luxuriant life contrasting with the apparently deserted surrounding water. Various vent fauna, bivalves, crustaceans, and worms take their energy and resources from endosymbiotic and episymbiotic chemoautotrophic microorganisms capable of oxidation of and sulfur compounds [3, 4, 5]. To survive under extreme conditions, such organisms must be adapted by appropriate metabolic pathways and protective mechanisms.
Some other volcanic areas also exist near the coast [1]. Shallow hot springs have been studied [6, 7]. Cold seeps are also characterized by hydrocarbon-rich fluids emanating from the ocean floor and are also the center of ecosystems with a large biodiversity [8, 9].
Near-shore sediments and deep-sea sediments have also been explored (the Ocean Drilling Project) and have revealed microbial life down to in the sediment under the sea floor [10].
In polar areas, the environment is perennially cold, and in some cases permanently covered with ice; therefore, microbes need to adapt to cold temperatures and fight against freezing. However, even in these extreme conditions, some bacteria have been collected at around in the Antarctic [11, 12, 13].
High saline water in inland water (the Dead Sea) together with high pH, high saltwater lakes situated underwater of seas such as the Black Sea, the Red Sea, the Mediterranean sea, or Gulf of Mexico have also been explored [14].
French Polynesia microbial mats are characterized by variable parameters such as pH values between and , salinity ranging from –, and temperatures between 20 nd . Microbial mats are laminated abundant communities of phototrophic and chemotrophic microorganisms. Pseudomonas, Alteromonas, Paracoccus, and Vibrio bacteria from these environments have been shown to produce GlossaryTerm
EPS
s [15, 16].Marine surfaces and especially marine eukaryotes are covered with microorganisms embedded in a matrix forming a biofilm. The microbial epibiotic consortia differ significantly from microorganisms living in the surrounding environment and highlight the close relationships between microbial epibionts and hosts [17].
From deep-sea vents to surface-sea ice and open ocean, nutrient sources and physic-chemical conditions may be highly fluctuant, making quick adaptive responses necessary; the adaptation is possible on the basis of genetic diversity and has led to huge biochemical diversity.
2.2 Microbial Diversity and the Limitation of Collections
A high diversity of ecosystems exists within the world’s oceans; they are different from terrestrial and freshwater environments; some endemic species have, therefore, been identified resulting in specific diversity.
Most of the biodiversity is microbial. The genetic diversity of microorganisms is far larger than that of plants and animals all together. Due to their large population size and since they grow rapidly, prokaryotes have an enormous potential to accumulate mutations, allowing them to adapt, and thus acquire genetic diversity. By weight, more than of all living organisms found in the oceans are microbial. Microorganisms are the most abundant life form in the ocean, they are the basis of the marine food web, drive energy and nutrient cycling, and are at the origin of life on the earth [18]. This genetic resource is mainly unknown and underexploited and will probably be the major source for biochemical diversity resulting in novel molecules in the future.
Isolating new species requires the ability to cultivate them. Cultivation efficiency only allows the isolation of – of microbial species present in marine ecosystems [19]. Therefore, microbial diversity is largely undiscovered. From oceanographic cruises or sampling campaigns, several academic or industrial teams all over the world have developed marine microbial collections. These they are a good source of screening for innovative biomolecules. These collections, except private ones, offer marine microorganisms and other services such as various screening. However, these collections shelter only cultivable microorganisms and, therefore, are limited. Nevertheless cultivation is a prerequisite to isolate compounds of biotechnological value such as GlossaryTerm
PS
s [20].Culture-independent molecular techniques such as metagenomics, also named environmental genomics, have been adopted to explore the actual diversity of natural assemblages of microbes. Their sensitivity allows the detection of taxa at very low abundance. They have also revealed the incredible richness of marine life.
For several years, J. Craig Venter’s Sorcerer II (Venter’s Global Ocean Sampling Expeditions) sailed all around the sea world to collect samples of near-surface sea water every miles. After filtration and freezing to collect and store microorganisms in the samples, GlossaryTerm
DNA
was extracted in the J. Craig Venter Institute laboratory and sequenced. Results revealed the power of metagenomic approaches to obtain a good estimation of microbial diversity and biochemical diversity; they also revealed a high proportion of new bacterial and viral sequences, which suggested that until now marine microorganisms have not been well isolated, especially viruses [21]. Many of these sequences must have potential as sources of new drugs and they are also studied for protein evolution, discovery of new enzymes, and microbial ecology concerns.Tara Oceans expeditions [22] started on 2004 to study the impact of climate change on marine ecosystems, especially marine plankton including microorganisms such as viruses and bacteria, and other organisms such as medusas. It sailed all over the seas to collect samples and data and provide them to the world’s scientific community. In years, expeditions have been organized with the aim to establish a time zero of marine ecosystems, to discover new organisms in each size class, to obtain a good estimation of oceanic biodiversity and its distribution, and to develop new models to estimate this biodiversity.
Deep-sea microbial diversity has been explored by metagenomic [23] or cultivation approaches [24, 25]. Sea floor sediments as well as hydrothermal vents with black or white smokers have been sampled. Studies on the genomic diversity of water columns [26] or sea floor sediments [27] have also been carried out as well as on microbial communities as associations with marine invertebrates, sponges [28] or with corals [29, 30].
In all cases, sequences related to fungi, bacteria, archaea, viruses and phages have been detected in variable ratios in a site-specific manner or isolated on culture media. The orders Thermococcales, Archaeoglobales, and Methanococcales are widely encountered in extremophilic archaea. On the other hand, the vast majority of bacterial sequences are mainly assigned to Thermotogales: Proteobacteria, Firmicutes, Cyanobacteria, and Actinobacteria, and – of marine bacteria are Gram-negative rather than Gram-positive.
Global Ocean Sampling Expeditions revealed that the archaea are nearly absent from the list of dominant organisms in the near-surface samples. Based on 16S ribonucleic acid (GlossaryTerm
RNA
) data, the most abundant phyla or classes are Proteobacteria, Bacteroidetes, Cyanobacteria, Firmicutes, Actinobacteria, and Planctomycetes [21]. The sequences mainly cluster within Alphaproteobacteria and Gammaproteobacteria. However distribution of these ribotypes is not homogeneous all over the ocean and different microbial communities can be distinguished.The diversity of seawater habitats results in diversity of the organisms, and subsequently of the metabolites they produce. Thus marine genomics provide sequence data for microbial ecology but also biodiscovery [31]. This renders marine ecosystems really interesting to discover innovative molecules for biotechnological purposes. Extremophiles will provide new molecules or pathways for novel biotechnological processes but are also models to investigate how biomolecules are stabilized under extreme conditions. Several studies on marine biodiversity for biotechnological applications have been carried out on particular habitats such as deep-sea hydrothermal vents for extremophiles and thermostable enzymes, polar areas for psychrophilic ones, for exopolysaccharides [1, 32, 33, 34, 35, 36], microbial mats from French Polynesia [16].
These bacteria may produce new enzymes and bioactive compounds, including exopolysaccharides (GlossaryTerm
EPS
s) with innovative structures (Table 39.1). Many known marine bacteria can produce molecules of biotechnological interest, some of which have unique properties, and the search for new microorganisms of biotechnological value is still promising.3 Bacterial Polysaccharides
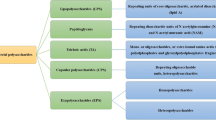
Different types of GlossaryTerm
PS
s exist within or around the bacterial cell depending on their location:-
i
Polysaccharides found in the cytoplasm that serve as carbon and energy reserve,
-
ii
Cell wall GlossaryTerm
PS
s including teichoic acids and peptidoglycans, -
iii
Extracellular GlossaryTerm
PS
s which are released outside the cell: exopolysaccharides (GlossaryTermEPS
s) or capsular polysaccharides (GlossaryTermCPS
).
GlossaryTerm
CPS
s are extracellular but remain attached to the cells through covalent bonds to other outer surface polymers thus forming the capsule.Intracellular GlossaryTerm
PS
s are produced when bacteria are grown in excess of sugar and may serve as reserve, since the cell also produces enzymes to degrade them. They usually appear as granules of glycogen or amylopectin type GlossaryTermPS
s and may be related to sporulation of Clostridium sp., for example. However, bacteria synthesize only very few intracellular GlossaryTermPS
s but they produce many diverse extracellular GlossaryTermPS
s.The cell wall is complex and rich in carbohydrate compounds. Among these, peptidoglycans are linear macromolecules consisting of GlossaryTerm
PS
s cross-linked by peptide moieties. The GlossaryTermPS
part is composed of alternating N-acetylmuramic acid (GlossaryTermMurNAc
) and N-acetylglucosamine (GlossaryTermGlcNAc
) linked by β-1,4-glycosidic linkages. Carboxylic groups from N-acetylmuramic acids are involved in the linkage to peptidic chains. Teichoic acids, which are only found in Gram-positive bacteria, may be glycerol phosphate units, or ribitol phosphate units linked by phosphodiester bonds. Glycerol and ribitol may also be associated with glucose, galactose, or N-acetylglucosamine. Teichoic acids are closely connected to the peptidoglycan network and make it more cohesive. Gram-negative bacteria also produce lipopolysaccharides (GlossaryTermLPS
) whose lipidic part (lipid A) is set inside the external membrane. They are composed of two carbohydrate parts, the O-specific chain, and the core [81]. The O-specific chain is highly variable, especially in pathogens such as Vibrio cholerae [81]. Lipid A is composed of aliphatic chains linked to glucosamine residue. GlossaryTermLPS
is also called endotoxin.The following paragraph deals specifically with extracellular GlossaryTerm
PS
s and GlossaryTermEPS
.3.1 Bacterial Exopolysaccharides
From different carbon sources, bacterial cells synthesize extracellular GlossaryTerm
PS
s; sugar units are synthesized within the cytoplasm before being excreted outside and polymerized. Extracellular GlossaryTermPS
s may remain attached to the cell membrane, forming a capsule or a slime, while the other part is released into the surrounding environment.Depending on the enzymatic machinery, GlossaryTerm
EPS
are usually constituted of a regular repeating unit. They may be linear or branched. Homopolysaccharides are composed of a unique type of sugar residue type, while heteropolysaccharides contain different ones [82].In heteropolysaccharides, glucose, galactose, and glucuronic or glacturonic acids are common. However, other neutral or acid sugars, hexosamines or N-acetyl-hexosamines, and some rarer sugars are also found. The presence or absence of a repeating unit containing up to or monomers [42], linear or branched, the presence of organic (lactate, pyruvate, N and O-acetates) or chemical (sulfate, phosphate) groups give a very large structural diversity of the molecules with consequently very diverse physicochemical or biological properties [83].
Depending on both sugar chain and osidic linkages, the behavior of the homopolysaccharides in solution may be different [84, 85]. Scleroglucan or pullulan are soluble and highly viscous, while curdlan lacking side-chains is poorly soluble and forms a gel; bacterial cellulose is insoluble; however, these three GlossaryTerm
PS
s are only composed of glucose. The presence of side-chains renders the polymer insoluble; acyl and non-sugar substituents may affect the physical features as well as uronic acids. β-1,4-linkages are rigid, while α-1,4-bonds found in dextrans confer flexibility of the structure [85].GlossaryTerm
EPS
chain length may vary from one species to another, but also within a species; the molecular weight may range from to over several million g mol.3.2 EPS from Marine Bacteria
A number of species of marine microorganisms can produce GlossaryTerm
EPS
s: cyanobacteria, bacteria, and archaea [86]. These strains are screened for mucoid phenotypes on a solidified medium supplemented with a carbohydrate source, for a high viscosity in liquid medium, by staining specifically GlossaryTermEPS
s.Chitin, the main marine polymer composed of β-1,4-linked N-acetylglucosamine residues, is one of the most abundant GlossaryTerm
PS
s in nature after cellulose and is usually prepared from the shells of crabs and shrimps [87, 88]. Until now, unlike for cellulose [89, 90], no bacterium has been shown to produce chitin. Nevertheless, many marine bacteria species exhibit chitinolytic activity and also produce chitosan from chitin, the N-deacetylated product [91]; bacteria are, therefore, involved in the turnover of the GlossaryTermPS
in the marine environment [87]. Some non marine bacteria species still produce polyglucosamine exopolymers with a structural homology with chitosan [92].Alginates, another kind of the most common marine GlossaryTerm
PS
s, are currently mainly obtained through brown seaweed harvesting; the non-marine bacteria (Pseudomonas aeruginosa and Azotobacter vinelandii) are also capable of producing acetylated alginates that bind some divalent ions but fail to form gels in the manner of their algal analogs.Polysaccharide-producing bacteria have been isolated from the Gulf of Naples [7], from microbial mats [16, 58, 93], the Antarctic [11], and deep-sea hydrothermal vents [43, 94, 95, 96, 97, 98]. Psychrotolerant Pseudoalteromonas, Shewanella, Polaribacter, or Flavobacterium sp. have been isolated from Antarctic water [56] and are able to produce high molecular weight GlossaryTerm
EPS
.Some GlossaryTerm
EPS
-producing marine bacteria are listed in Table 39.1. The vast majority of marine bacteria producing exopolysaccharides are Alteromonas or the Pseudoalteromonas, Pseudomonas, Shewanella, and Vibrio species. In contrast only a few extremophilic bacteria or archaea have been studied for GlossaryTermEPS
s [36, 55, 64].GlossaryTerm
EPS
production by microorganisms has the advantages of a rapid production (a few days compared to the – months in the case of plants), of a production in bioreactors on different substrates of possibly hydrocarbon residues and of easier extraction since they are exuded in the extracellular environment.3.3 Benefits for the Bacterial Cell
Many marine bacteria produce GlossaryTerm
EPS
as a strategy for growth and survival, adhesion to surfaces, and resistance to adverse conditions. GlossaryTermEPS
s have also been involved in virulence and host contamination. The physiologic roles of GlossaryTermEPS
for bacteria have not been completely identified, but a protective or adhesive role has frequently been proposed. Most of the time, GlossaryTermEPS
s are not degraded by the bacteria producing them and thus do not represent a carbon source [99]. In a variable nutrient environment such as the ocean, bacteria need greater adaptability in the pathways involved in the detection of nutrients and in metabolism to utilize them. This could explain the diversity of already found polysaccharidic structures.Therefore, GlossaryTerm
EPS
s have an effect on the interactions between the cells and the surrounding environment [86]. GlossaryTermCPS
or slime may help bacterial cells to overcome the various stresses encountered in the environment or to adhere to the surfaces, providing survival advantage. In aquatic environments, the majority of bacteria adhere in a selective manner or not to inert surfaces or living organisms. In biofilms, microbial cells aggregate within a matrix of extracellular polymeric substances composed of GlossaryTermPS
s, proteins, nucleic acids, peptidoglycan and lipids. Extracellular GlossaryTermPS
s are generally considered to be the main part of the biofilm matrix [100] and are involved in the flexibility of the biofilm shape under shear force as well as in some of the functional properties of the matrix [101].Moreover, anionic GlossaryTerm
EPS
s slimes or from capsules allow the aggregation and storage of the soluble nutriments necessary for the growth of the cell or other particles and may chelate metals and ions [11]. Bacterial symbionts have been involved in the resistance of polychaetes to the high metal levels around deep-sea hydrothermal vents [3] and in environments anthropogenically polluted with metals [102]. In other respects, several anionic GlossaryTermEPS
s from marine bacteria have been shown to bind toxic compounds [13, 37, 62, 94]. The presence of bacteria on marine worms may protect them against toxic metals.3.4 Putative Pathways of Biosynthesis
Oceans contain huge bioresources. To exploit them in a manner respectful to the environment, researchers are trying to decipher their production mechanisms. The first particular mechanism of biosynthesis was encountered for homopolysaccharides such as dextran, glucan, levan, inulin, and fructan; it involves an extracellular enzyme (glycan-sucrase) secreted at the cell surface of Gram-positive bacteria such as Leuconostoc and other lactic acid bacteria; it has not been yet described in Gram-negative bacteria [103, 84]. A specific glycosyltransferase (GlossaryTerm
GT
) transfers the sugar residue to the nascent polymer from a disaccharide, usually saccharose.The other kinds of biosynthesis concern homo and heteropolysaccharides with repeating units. This biosynthesis usually involves three steps: sugar precursor production, repeating unit synthesis, export and polymerization (Fig. 39.1). The whole process is not fully understood, in particular for polymer exportation and synthesis regulations.


Common biosynthetic mechanism of PSs with repeating units (the biosynthesis begins within the cytoplasm with the synthesis of activated precursors. The repeating unit is then assembled by GT on a carrier located on the membrane. The repeating unit is then translocated across the membrane, polymerized on the nascent polymer within the periplasm or outside the cell, and then released in the surrounding environment or slime)
The synthesis of heteropolysaccharides has been widely studied in Gram-negative bacteria (Escherichia coli [104]) as well as lactic acid bacteria and appears to involve a common mechanism. Several enzymes are involved in the assemblage of the repeating unit; they are encoded by genes located within large GlossaryTerm
EPS
gene clusters ranging up to –. They are sometimes located on plasmids, especially when considering lactic acid bacteria [105]. The biosynthetic machinery involves activated sugar precursors such as nucleoside sugar diphosphate (GlossaryTermUDP-sugar
), which are produced in the central metabolic pathways. GlossaryTermEPS
biosynthesis from cytoplasmic activated GlossaryTermUDP-sugar
precursors is common. However, alternative activated sugar donors are encountered: nucleoside sugar monophosphates, lipid sugar phosphates or pyrophosphates, or unsubstituted sugar phosphate [106]. GlossaryTermEPS
s are usually synthesized inside the cytoplasm in the form of repeating units by glycosyltransferases before being excreted [104]. The repeated unit is assembled on a carrier molecule, anchored in the cytoplasmic membrane. Different molecular acceptors on the membrane for the carbohydrate in formation have been proposed; in particular, a lipidic transporter such as undecaprenyl phosphate or isoprenoid pyrophosphate is used in GlossaryTermEPS
biosynthesis [95]. The first monomer is linked to the lipid-carrier by the priming-glycosyltransferase or phosphoglycosyltransferase. The following sugar monomers are linked by appropriate GlossaryTermGT
s to the growing repeating unit. After completion, the repeating unit is exported outside the cell and subsequently polymerized by addition to the reducing end of the growing GlossaryTermEPS
chain on the outer face of the cell membrane (Fig. 39.1). Regulations are also involved but not well understood. In particular, a chain length determination factor regulates the extension of the GlossaryTermPS
molecule; the efficiency of this regulation may depend on whether or not the chain length feature is important for the cell. Various exportation mechanisms have been described; some of them involve GlossaryTermATP
(adenosine triphosphate)-binding-cassette (GlossaryTermABC
)-transporters and others an externalization of the lipid carrier by a Wzx flippase [104, 107].A specific biosynthesis mechanism involves a bifunctional glycosyltransferase called synthase. Cellulose [108] and hyaluronan (GlossaryTerm
HA
) [109, 110] are examples of the molecular pathway of that biosynthesis. The GlossaryTermEPS
chain is built by repetitive, non-processive addition of monosaccharides to the nascent polymer. Therefore, it seems that this mechanism can only exist for simple heteropolysaccharides (up to two sugar residues in the repeating unit). The GlossaryTermEPS
is subsequently exported out of the cell by an GlossaryTermABC
transporter; synthase has also been described to be able to form a pore across the cell membrane, allowing the polymer to be exported while synthesized [107, 110].Alginate is a linear, anionic heteropolysaccharide consisting of β-d-mannuronic acid (GlossaryTerm
ManA
) and α-d-guluronic acid (GlossaryTermGulA
). The primary structure of alginate relies on GlossaryTermManA
-blocks containing homogeneous (GlossaryTermManA
), GlossaryTermGulA
-blocks or (GlossaryTermGulA
), and alternating GlossaryTermManA
–GlossaryTermGulA
-blocks. The relative amount of GlossaryTermManA
and GlossaryTermGulA
as well as the length and distribution of the blocks are dependent on the producing species but are fundamental to the swelling and gelling features, as well as the solubility [111]. The bacterial alginate biosynthesis mechanism is particular since the polymer is not constituted of a repeating unit [112].Except for xanthan production (up to in the culture broth) or pullulan production, production yields upon fermentation seem low (up to usually). However, an improved activity or a new one resulting in new applications may have a good chance of success for industrial applications.
Engineering of microbial production as well as of genetic material of bacteria, together with the exploration of biodiversity, are different promising approaches to find new GlossaryTerm
PS
s with innovative properties.4 Applications of EPSs
Several GlossaryTerm
PS
s are well established in different industrial fields. The large range of these existing applications together with the potential ones reflects the many proposed functions in nature as well as their biochemical biodiversity.Polysaccharides as texture agents of thickening or stabilizing additives are already widely used, such as xanthan, gellan, or some algal GlossaryTerm
PS
s (agars, alginates, carrageenans). Therefore, a large number of applications exploit the rheological properties of GlossaryTermEPS
s. Marine GlossaryTermEPS
s having particular functional properties render them more resistant to extreme temperatures, pH, or salinity are of great value for industry [113].Alginate GlossaryTerm
PS
s form hydrogels and have found applications in explosives as gelling agents, in the paper industry and textile printing as water-holding agents, in antifoams and lattices because of their emulsifying properties, and in cleaners and ceramics as stabilizers. Alginates are also used in the controlled release of active ingredients that are entrapped in calcium alginate beads, and as new biomaterials for cell immobilization and tissue engineering [111]. Research on chitin and chitosan have resulted in applications in various industrial and biomedical fields [88].In plant and animal cells, GlossaryTerm
PS
s participate in many central biological processes through the interaction with key proteins such as chemokines, cytokines, growth factors, enzymes, and adhesion molecules involved in cell development, cell signaling, and cell integrity. Therefore, glycobiology, which studies these interactions, and impact on human health have resulted in an increased demand for GlossaryTermPS
s for therapeutic purposes in cancer, inflammatory diseases, pathogen infections, and thromboembolic disease [114, 115]. The potential in this domain of bacterial GlossaryTermPS
s is now well recognized [116, 117].GlossaryTerm
EPS
from the bacteria Xanthomonas campestris (xanthan), Sphingomonas paucimobilis, or Sphingomonas (formerly Pseudomonas) elodea (gellan), Acetobacter xylinum now called Gluconacetobacter xylinus (cellulose), Rhizobium sp. (succinoglycan), Agrobacterium sp. (previously Alcaligenes faecalis) (curdlan), and from the fungi Sclerotium rolfsii (scleroglucan) and Aureobasidium pullulans (pullulan), as well as dextrans from various bacteria have been commercialized and are the most utilized GlossaryTermPS
s [118, 84]. Microbial hyaluronic acid (Streptococcus sp.) is also at commercial level and is mainly used in cosmetics, in ophthalmology, and in wound healing [119].4.1 Food Products
Polysaccharides are used as ingredients in food products, as carbohydrate and dietary fibre source but also as hydrocolloid to influence rheology and texture. Therefore, in addition to the sensory benefits of GlossaryTerm
EPS
in food products, they also impact human health and nutrition [120]. GlossaryTermEPS
s are also produced in situ by bacteria involved in food processing, such as probiotic lactic acid bacteria, and act as functional or prebiotic ingredients. Alginates and various vegetal gums are thickeners, gelling agents, and emulsifiers in food products [121].4.2 Environment
The biotechnologies applied to the domain of the environment are being developed and concern essentially the depollution and the rehabilitation of ground, water, and effluents by means of microbial techniques (bioremediation, biodetoxification). Thus, research projects relative to the applications of the bacterial biopolymers in the problems of environmental protection have been conducted for a few years. These concerns are mainly due to the biosorbent properties of the GlossaryTerm
PS
s to remove toxic metal pollutants [113].4.3 Cosmetics
Cosmetics is a fast growing branch of industry that responds to a societal demand. The lifetime of new molecules ranges between –, leading to a constant search for new molecules. Polysaccharides act in feeding, regenerating, and maintaining the skin. Among the most popular products are molecules with moisturizing, smoothing, and/or antiwrinkle activity, as well as those active in the cellular repair or the regeneration of dermic cells and GlossaryTerm
UV
protection. Polysaccharides represent more than of the molecules used in the cosmetic field (hyaluronic acid, chitosan, and β-glucane).The GlossaryTerm
PS
deepsane is produced by Alteromonas macleodii subsp. fijiensis biovar deepsane [43] and is used in cosmetics. This GlossaryTermEPS
shows a protective action of keratinocytes, which are the main cells of the skin, from a proinflammatory agent. Protective effects were also found for cells sensitive to ultraviolet attacks and involved in the cutaneous immune defense system. This GlossaryTermEPS
would, therefore, contribute to the repair of the skin [122].4.4 Medical Applications
The most expected contribution from biotechnologies in the healthcare field concerns the development of new molecules with new biological activities or a better profit–risk ratio. This search inevitably passes by a phase of bioprospecting, and the marine environment constitutes a privileged field of investigations. Therefore, bacterial GlossaryTerm
EPS
s offer new and innovative approaches to handle a large number of diseases and to replace certain drugs. Moreover, bacterial GlossaryTermEPS
s are not dependent on climatic or physiologic variation, since they are produced in a controlled environment in bioreactors.The first kind of medical applications derives from the functional properties of the GlossaryTerm
PS
molecule (rheology, water-holding, or gelification) [123]. As an example, dextran, even widely used as chromatographic supports for purifications of molecules, is also developed in the therapeutic field as artificial plasma. Some bacterial GlossaryTermEPS
s may also serve as encapsulating agents for a controlled release of drugs near the target, in medical textiles for wound care, in artificial skin and tissue engineering scaffolds [121]. Streptococcus zooepidermicus produces a hyaluronate with a structure identical to the mammalian one. This GlossaryTermEPS
is used in ophthalmic surgery. Some other uses are under study, such as various biomaterials and prosthetic surgery to improve biocompatibility. Hydrogels based on natural polymers are close to living tissues and can help during the healing process. Most of these GlossaryTermPS
s are high molecular weight GlossaryTermPS
s.Biological properties constitute the basis for the second type of applications. Some pathogenic bacteria exhibit GlossaryTerm
PS
s on their surface; this is the principal factor for immune response. Consequently, they are potential vaccines. Examples of GlossaryTermPS
vaccines are those against pneumonia (Streptococcus pneumoniae) and pneumococcal disease (Neisseria meningitidis serogroups A, C, Y, and W135).Some GlossaryTerm
EPS
s have been studied for their antiviral effects [124, 125, 126], e. g., microbial β-1,3-d-glucans, including levan as well as sulfated dextran [127, 128]. Other bacterial GlossaryTermPS
s exhibit diverse bioactivities, which could also find interest in the biomedical field as therapeutics. Curdlan [127] shows antitumor activity. The marine Geobacillus thermodenitrificans produces an GlossaryTermEPS
having immunomodulatory and antiviral effects [49]. Some GlossaryTermPS
s may exhibit innovative structure similar to heparin, a mammalian GlossaryTermPS
with biological activities such as anticoagulant, antitumor or antiviral. Indeed within the structural diversity, some homologies of new bacterial GlossaryTermEPS
s have been identified with molecules from animal and bacterial molecules might advantageously replace animal ones. GlossaryTermCPS
s from Escherichia coli are usually antigenic but the Escherichia coli K5 strain produces an extracellular GlossaryTermPS
, called K5, whose structure, [4-d-GlcA β-1,4-d-GlossaryTermGlcNAc
α-1,], is similar to a heparin precursor, N-acetylheparosan [129]. Because of this structural relationship, chemically and/or enzymatically modified K5 GlossaryTermPS
is of considerable interest for biomedical applications. Such a project is currently being undertaken by several research teams. This concern is developed in the following paragraph with a focus on marine GlossaryTermEPS
s.5 Marine EPSs as Glycosaminoglycans (GAGs)
Glycosaminoglycans (GlossaryTerm
GAG
s) constitute a class of glycans found ubiquitously in mammalian tissues as components of proteoglycans that have various cellular and intercellular matrix functions. Vertebrates utilize GlossaryTermGAG
s in structural, recognition, adhesion, and signaling roles. GlossaryTermGAG
chains are negatively charged by the presence of uronic acids and sulfate groups. They are made up of hexuronic acid and N-acetylhexosamine in alternating linear sequence. These GlossaryTermPS
s exhibit polydisperse high molecular weights in the range of several thousands to millions g mol. GlossaryTermGAG
structural diversity comes from isomers differing in sulfate presence and position and uronic acid epimerization (Table 39.2). Among them, heparin and heparan sulfate (GlossaryTermHS
) represent the most heterogeneous structural group because of different sulfation degrees and position as well as various extents of uronic acid epimerization. Postpolymerization modifications of the GlossaryTermPS
backbone varies depending on the tissue and developmental stage [130]. GlossaryTermHA
is the only non-sulfated GlossaryTermGAG
.Heparin is widely used in the clinical treatment of thrombosis as an intravenous anticoagulant with produced annually through extraction from porcine intestinal mucous membranes.
5.1 Biological Activity and Structure–Activity Relationship
The high negative charge of GlossaryTerm
GAG
molecules allows binding to many cellular compounds (receptors, growth factors, …), which results in various bioactivities [131]. Medical applications for GlossaryTermGAG
s are manifold. They are used as surgical aids, moisturizers, for drug delivery, in tissue engineering, as anticoagulants, and for their anticancer activities [123, 130, 132, 133]. These biological activities are dependent on the key structural features. Advances in the understanding of the relationships between biological activity and structure are important for the development of safer bioactive analogs.Sulfation of carbohydrate compounds enhances their biological activity such as their antiviral [125, 126] or anticoagulant activity [115]. Although the mechanisms are not clear, it has been proposed that sulfates contribute to a molecular conformation that is essential for the activity which, in the case of spirulan, is dependent on calcium ion chelation by sulfates [125].
GlossaryTerm
HA
, heparin, and chondroitin sulfate (GlossaryTermCS
) are currently used in various medical applications. Linear GlossaryTermHA
is mainly used in cosmetics to reduce wrinkles, in drug delivery, in ophthalmology, and in wound healing for its hydratation capability and viscoelasticity [119]. Derivatized GlossaryTermHA
and cross-linked GlossaryTermHA
are also used in healthcare as non-surgical dermal tissue fillers and in the treatment of osteoarthritis and in tissue engineering [119].The key pentasaccharide of heparin has been well studied and shown to be involved in the interaction with antithrombin, inhibiting blood coagulation [134]. In addition, some trisaccharides have been identified to be responsible for binding with fibroblast growth factors having an important role in cell proliferation, differentiation, and migration, as well as angiogenesis and, therefore, can inhibit cancer tumor development. The production of low molecular weight heparinoid would allow us to obtain more efficient preparations with lower side effects by targeting the structural features involved and consequently to obtain more specific preparations [131]. In addition, low molecular weight compounds are easier to inject as therapeutics drugs. The use of GlossaryTerm
EPS
s as active ingredients does, indeed, concern mainly low molecular weight polymers with bioactivity in cardiovascular and cancer domains.5.2 Modifications to Create GAG-Like Molecules
When an active drug is extracted from animals (heparin from porcine intestinal mucosa, GlossaryTerm
HA
from rooster comb, GlossaryTermCS
from bovine trachea or shark cartilage) or marine invertebrates (scallop, whelk, crustacean, sea squirt), a risk of allergic response to contaminating compounds, or of contamination by non-conventional agents, e. g., prions or viruses [130], exists and some new molecules are searched for. In actual fact, GlossaryTermGAG
s also exhibit undesirable side effects such as hemorrhagic risk; therefore, new analog drugs are being developed [132].For future applications, GlossaryTerm
GAG
s derived from bacteria seem to be safer materials with higher purity. Molecules similar to GlossaryTermGAG
s have been found in microbial sources [130]. As has already been presented, Streptococci as well as Pasteurella produce an GlossaryTermEPS
identical to GlossaryTermHA
acid [119]. Recently, the recombinant production of GlossaryTermHA
has been achieved in Bacillus subtilis [135] and Lactococcus lactis [136]. Both GlossaryTermHA
produced by extraction from rooster combs or by microbial fermentation are structurally identical and it can, therefore, be used for its physical properties such as viscosification in ophthalmology, orthopaedic surgery, rheumatology, dermatology, plastic surgery, and wound healing without any modifications of the structure [137].The marine bacterium Vibrio diabolicus isolated on the Pompei worm from deep-sea hydrothermal vents has been shown to produce the HE800 polymer constituted of a tetrasaccharidic repeating unit [3-d-GlossaryTerm
GlcNAc
-β-(1,4)-d-GlcA-β-(1,4)-d-GlcA-β-(1,4)-d-GalNAc-α-(1] [66, 67]. This GlossaryTermEPS
shows structural similarities to GlossaryTermHA
. Bioactivity in bone repair has also been described [138]. The HE800 GlossaryTermEPS
in its native form has found application in skin and cartilage therapy [133]. Microbial-derived GlossaryTermHA
has now been commercialized and even if research on marine bacteria may produce analogs, research is also directed at producing heparin via microbial fermentation [139].Beside natural the GlossaryTerm
PS
s available, some GlossaryTermPS
derivatives exhibiting enhanced bioactivity or features can be generated by chemical, biochemical, and genetic methods, or a combination thereof to modify the GlossaryTermPS
structure through depolymerization and regioselective functionalization. The current challenge, in addition to improving the productivity of GlossaryTermEPS
s from bacteria, is to produce GlossaryTermEPS
s of a structure and size that allow the desired activity. In particular, the sulfation degree and pattern must be controlled since the biological activity of heparinoid relies greatly on the specific modification pattern (N-deacetylation, N and O-sulfation, and epimerization of glucuronic acid to iduronic acid). Tailoring the structure is currently performed in chemical ways, but enzymatic ways are also under study.Pullulan was modified by chemical sulfation to produce new anticoagulant drugs as a potential heparin substituent [140]. Some analogs of GlossaryTerm
GAG
sugar backbones have also been found in bacteria such as Escherichia coli K4 and K5, as well as in marine bacteria. E. coli K4 and K5 have been shown to produce GlossaryTermPS
s with a basic structural similarity to chondroitin sulfate and heparin, respectively [139, 141]. The E. coli K5 polysaccharidic antigen is similar to a precursor of heparin (desulfoheparin). Much in vitro chemoenzymatic production of heparin, in which the GlossaryTermEPS
backbone of E. coli K5 is subjected to chemical and/or enzymatic modification, have been described [110, 139, 142, 143, 144, 145].Alteromonas infernus, a deep-sea marine bacterium has been isolated from seawater samples collected around a dense population of giant worms Riftia pachyptila [38]. It produces an anionic complex heteropolysaccharide with a high molecular weight and around sulfate content [39]. Some other sulfated GlossaryTerm
EPS
s from marine bacteria have been described in Alteromonas macleodii strains and Pseudoalteromonas sp. [42, 60]. There are very few bacterial sulfated GlossaryTermPS
s whose structure has been determined (Table 39.1), but they provide the basis for future applications and basic research on GlossaryTermGAG
-like molecules from marine environments. On the other hand, l-iduronic acid resulting from 5-epimerization of glucuronic acid has been described in one bacterial GlossaryTermPS
[146]. Since sulfated GlossaryTermPS
s and l-iduronic acid exist in bacteria, one can expect the isolation of enzymes involved in their biosynthesis in the near future. These enzymes would then be very useful tools to produce engineered GlossaryTermGAG
-like GlossaryTermPS
s.The A. infernus GlossaryTerm
EPS
repeating unit is a non-saccharide composed of glucuronic acid, galacturonic acid, and neutral sugars (Gal and Glc), as well as one sulfate group on a galacturonic acid residue (Table 39.1). This GlossaryTermPS
prepared in a hydrogel exhibited interesting activity in cartilage tissue engineering applications [147]. Chemically oversulfated low molecular weight derivatives have been synthesized by two processes that differ in the order of the same steps (radical depolymerization and sulfation). Surprisingly, the two kinds of derivatives did not exhibit the same biological features: one was able to stimulate the proliferation of mature endothelial cells, whereas the other was not [148], which suggested that the modification pattern catalyzed by both processes is not the same; therefore, the sulfation pattern appears to be critical for bioactivity. However, lteromonas infernus GlossaryTermEPS
derivatives presented anticoagulant activity similar to that of heparin with a lower hemorrhagic risk [149]. These derivatives have also shown interesting properties for cell therapy and tissue engineering [133], and also in cartilage regeneration [150].These studies have highlighted the crucial roles of molecular weight and the the sulfation pattern [151]. Chemical reactions may be selective to some extent but are not specific enough for such molecules. In particular, it is very difficult to obtain compounds with specific sulfated positions [151, 152]. Therefore, the use of enzymatic methods in the modification process leading to bioactive oligosaccharidic derivatives may have clear advantages, such as a better control of reaction selectivity and an easier reaction in a single step (without functional group protection and de-protection steps) under mild conditions. Most enzymes involved in GlossaryTerm
GAG
modification are very specific, especially sulfotransferases [114, 153, 154]. Moreover, the GlossaryTermPAPS
(3-phosphoadenosine 5-phosphosulfate) molecule required for the sulfation reaction as the sulfate donor is expensive. Therefore, approaches to regenerate this cofactor have been developed [153, 155]. However, finding active enzymes on each GlossaryTermPS
backbone is likely to be the main bottleneck of such a strategy; therefore, enzyme design and optimization by molecular techniques will surely need to be envisaged.In the future, one can also expect that genetic engineering will be performed to tailor carbohydrate biomolecules in vivo. This will depend on a deep understanding of biosynthetic mechanisms of GlossaryTerm
EPS
s to be able to engineer them and of how the structure of GlossaryTermEPS
s determines the final activity. The development of improved characterization methods is also required, especially to determine the distribution of functional groups as well as new methods to modify the GlossaryTermPS
backbone in a regiospecific manner.6 Conclusion
Life emerged from the ocean; nowadays, oceans shelter large biomasses that are mainly microscopic. Among these, marine bacterial diversity, associated with the original ecosystem and biochemical metabolim, is a huge reservoir for the identification of new species and the extraction of new molecules of biotechnological interest, resulting in both fundamental and applied research. It is pointed out that the ocean provides renewable resources for different industrial fields.
The putative or demonstrated benefits of some new GlossaryTerm
PS
s and their derivatives are still being studied. Crucial knowledge has been gained on how the structural composition influences the functional properties and on how these features can be applied in biomedical research. In vitro and in vivo structural design of GlossaryTermPS
s to tailor targeted GlossaryTermPS
derivatives might produce important bioactive materials in the future. The growing demand will probably lead to the establishment of an adequate and efficient microbial production process.Only a few marine products have reached commercial production. Indeed, the search for new molecules for healthcare rarely encounters success, and it is always a long time before they are released on the market. The number of marine molecules used as drugs is very low; anticancer, antiviral, and pain-fighting molecules are available on the market, but none are carbohydrates. However, oceans are not only a source of molecules but also a source of inspiration for the synthesis of new drugs [156].
In biotechnology, the development of new marine molecules is linked to the progress in the sustainable production process. Moreover, one will have to take into consideration the evolution of legislation regarding the use of natural products, intellectual property rights, the availability of the resource, the ease of implementation, and naturally the production costs.
Abbreviations
- ABC:
-
ATP-binding cassette
- ATCC:
-
American Type Culture Collection
- ATP:
-
adenosine triphosphate
- CPS:
-
capsular polysaccharide
- CS:
-
chondroitin sulfate
- DNA:
-
deoxyribonucleic acid
- EPS:
-
exopolysaccharide
- GAG:
-
glycosaminoglycan
- GT:
-
glycosyltransferase
- GlcNAc:
-
N-acetylglucosamine
- GulA:
-
α-d-guluronic acid
- HA:
-
hyaluronic acid
- HS:
-
heparan sulfate
- LPS:
-
lipopolysaccharide
- ManA:
-
β-d-mannuronic acid
- MurNAc:
-
N-acetylmuramic acid
- PAPS:
-
3-phosphoadenosine 5-phosphosulfate
- PS:
-
polysaccharide
- RNA:
-
ribonucleic acid
- UDP-sugar:
-
nucleoside sugar diphosphate
- UV:
-
ultraviolet
References
J.W. Deming: Deep ocean environmental biotechnology, Curr. Opin. Biotechnol. 9, 283–287 (1998)
N. Le Bris, F. Gaill: How does the annelid Alvinella pompejana deal with an extreme hydrothermal environment?, Rev. Environ. Sci. Biotechnol. 6(1–3), 197–221 (2007)
D. Desbruyères, P. Chevaldonné, A.M. Alayse, D. Jollivet, F.H. Lallier, C. Jouin-Toulmond, F. Zal, P.M. Sarradin, R. Cosson, J.C. Caprais, C. Arndt, J. O'Brien, J. Guezennec, S. Hourdez, R. Riso, F. Gaill, L. Laubier, A. Toulmond: Biology and ecology of the Pompeii worm (Alvinella pompejana Desbruyères and Laubier), a normal dweller of an extreme deep-sea environment: A synthesis of current knowledge and recent developments, Deep Sea Res. Part II Top. Stud. Oceanogr. 45, 383–422 (1998)
K. Alain, M. Olagnon, D. Desbruyères, A. Pagé, G. Barbier, S.K. Juniper, J. Quérellou, M.-A. Cambon-Bonavita: Phylogenetic characterization of the bacterial assemblage associated with mucous secretions of the hydrothermal vent polychaete Paralvinella palmiformis, FEMS Microbiol. Ecol. 42(3), 463–476 (2002)
S. Nakagawa, K. Takai: Deep-sea vent chemoautotrophs: Diversity, biochemistry and ecological significance, FEMS Microbiol. Ecol. 65(1), 1–14 (2008)
T.L. Maugeri, C. Gugliandolo, D. Caccamo, A. Panico, L. Lama, A. Gambacorta, B. Nicolaus: A halophilic thermotolerant Bacillus isolated from a marine hot spring able to produce a new exopolysaccharide, Biotechnol. Lett. 24, 515–519 (2002)
B. Nicolaus, L. Lama, A. Panico, V.S. Moriello, I. Romano, A. Gambacorta: Production and characterization of exopolysaccharides excreted by thermophilic bacteria from shallow, marine hydrothermal vents of flegrean ares (Italy), Syst. Appl. Microbiol. 25(3), 319–325 (2002)
B.B. Jorgensen, A. Boetius: Feast and famine – micobial life in the deep-sea bed, Nat. Rev. Microbiol. 5, 770–781 (2007)
C.L. Van Dover, C.R. German, K.G. Speer, L.M. Parson, R.C. Vrijenhoek: Evolution and biogeography of deep-sea vent and seep invertebrates, Science 295(5558), 1253–1257 (2002)
E.G. Roussel, M.-A.C. Bonavita, J. Querellou, B.A. Cragg, G. Webster, D. Prieur, R.J. Parkes: Extending the sub-sea-floor biosphere, Science 320(5879), 1046 (2008)
C.S. Hassler, E. Alasonati, C.A. Mancuso Nichols, V.I. Slaveykova: Exopolysaccharides produced by bacteria isolated from the pelagic Southern Ocean – Role in Fe binding, chemical reactivity, and bioavailability, Mar. Chem. 123, 88–98 (2011)
C.A. Nichols, J. Guezennec, J.P. Bowman: Bacterial exopolysaccharides from extreme marine environments with special consideration of the southern ocean, sea ice, and deep-sea hydrothermal vents: A review, Mar. Biotechnol. 7(4), 253–271 (2005)
C.A. Mancuso Nichols, S. Garon, J.P. Bowman, G. Raguenes, J. Guezennec: Production of exopolysaccharides by Antarctic marine bacterial isolates, J. Appl. Microbiol. 96, 1057–1066 (2004)
J. Quérellou, J. Guézennec: Biotechnologie des extrêmophiles, Tech. Ing. BIO 580, 1–13 (2010)
L. Richert, S. Golubic, R. Le Guédès, J. Ratiskol, C. Payri, J. Guézennec: Characterization of exopolysaccharides produced by cyanobacteria isolated from Polynesian microbial mats, Curr. Microbiol. 51, 379–384 (2005)
J. Guézennec, X. Moppert, G. Raguénès, L. Richert, B. Costa, C. Simon-Colin: Microbial mats in French Polynesia and their biotechnological applications, Process Biochem. 46(1), 16–22 (2011)
A. Penesyan, S. Kjelleberg, S. Egan: Development of novel drugs from marine surface associated microorganisms, Mar. Drugs 8(3), 438–459 (2010)
W.B. Whitman, D.C. Coleman, W.J. Wiebe: Prokaryotes: The unseen majority, Proc. Natl. Acad. Sci. USA 95(12), 6578–6583 (1998)
J.T. Staley, A. Konopka: Measurement of in situ activities of non photosynthetic microorganisms in aquatic and terrestrial habitats, Annu. Rev. Microbiol. 39(1), 321–346 (1985)
I. Joint, M. Mühling, J. Querellou: Minireview: Culturing marine bacteria – an essential prerequisite for biodiscovery, Microb. Biotechnol. 3(5), 564–575 (2010)
Oceanic metagenomics collection: PLoS Biology 5(3), http://plosbiology.org (2007)
Tara Oceans expeditions website: http://oceans.taraexpeditions.org (2009)
M.L. Sogin, H.G. Morrison, J.A. Huber, D.M. Welch, S.M. Huse, P.R. Neal, J.M. Arrieta, G.J. Herndl: Microbial diversity in the deep sea and the underexplored rare biosphere, Proc. Natl. Acad. Sci. USA 103(32), 12115–12120 (2006)
G. Raguenes, J.R. Meunier, E. Antoine, A. Godfroy, J.C. Caprais, F. Lesongeur, J. Guezennec, G. Barbier: Biodiversity of hyperthermophilic archaea from hydrothermal vents of the East Pacific Rise, Comptes Rendus Acad. Sci. Ser. III, Sci. Vie. 318(3), 395–402 (1995)
A. Postec, F. Lesongeur, P. Pignet, B. Ollivier, J. Querellou, A. Godfroy: Continuous enrichment cultures: Insights into prokaryotic diversity and metabolic interactions in deep-sea vent chimneys, Extremophiles 11(6), 747–757 (2007)
E.F. DeLong, C.M. Preston, T. Mincer, V. Rich, S.J. Hallam, N.-U. Frigaard, A. Martinez, M.B. Sullivan, R. Edwards, B.R. Brito, S.W. Chisholm, D.M. Karl: Community genomics among stratified microbial assemblages in the ocean's interior, Science 311(5760), 496–503 (2006)
J.F. Biddle, S. Fitz-Gibbon, S.C. Schuster, J.E. Brenchley, C.H. House: Metagenomic signatures of the Peru Margin subseafloor biosphere show a genetically distinct environment, Proc. Natl. Acad. Sci. USA 105(30), 10583–10588 (2008)
N.S. Webster, A.P. Negri, M.M.H.G. Munro, C.N. Battershill: Diverse microbial communities inhabit Antarctic sponges, Environ. Microbiol. 6(3), 288–300 (2004)
N.S. Webster, D. Bourne: Bacterial community structure associated with the Antarctic soft coral, Alcyonium antarcticum, FEMS Microbiol. Ecol. 59(1), 81–94 (2007)
L. Wegley, R. Edwards, B. Rodriguez-Brito, H. Liu, F. Rohwer: Metagenomic analysis of the microbial community associated with the coral Porites astreoides, Environ. Microbiol. 9(11), 2707–2719 (2007)
K.B. Heidelberg, J.A. Gilbert, I. Joint: Minireview: Marine genomics: At the interface of marine microbial ecology and biodiscovery, Microb. Biotechnol. 3(5), 531–543 (2010)
B. van den Burg: Extremophiles as a source for novel enzymes, Curr. Opin. Microbiol. 6(3), 213–218 (2003)
C.A. Mancuso Nichols, J. Guezennec, J.P. Bowman: Bacterial exopolysaccharides from exteme marine environments with special consideration of the Southern Ocean, sea ice, and deep-sea hydrothermal vents: A review, Mar. Biotechnol. 7, 253–271 (2005)
J. Kennedy, B. Flemer, S.A. Jackson, D.P.H. Lejon, J.P. Morrissey, F. O'Gara, A.D.W. Dobson: Marine metagenomics: New tools for the study and exploitation of marine microbial metabolism, Mar. Drugs 8(3), 608–628 (2010)
A. Trincone: Marine biocatalysts: Enzymatic features and applications, Mar. Drugs 9(4), 478–499 (2011)
B. Nicolaus, M. Kambourova, E.T. Oner: Exopolysaccharides from extremophiles: From fundamentals to biotechnology, Environ. Technology 31(10), 1145–1158 (2010)
J.A. Mata, V. Béjar, P. Bressollier, R. Tallon, M.C. Urdaci, E. Quesada, I. Llamas: Characterization of exopolysaccharides produced by three moderately halophilic bacteria belonging to the family Alteromonadaceae, J. Appl. Microbiol. 105(2), 521–528 (2008)
G.H. Raguenes, A. Peres, R. Ruimy, P. Pignet, R. Christen, M. Loaec, H. Rougeaux, G. Barbier, J.G. Guezennec: Alteromonas infernus sp. nov., a new polysaccharide-producing bacterium isolated from a deep-sea hydrothermal vent, J. Appl. Microbiol. 82(4), 422–430 (1997)
O. Roger, N. Kervarec, J. Ratiskol, S. Colliec-Jouault, L. Chevolot: Structural studies of the main exopolysaccharide produced by the deep-sea bacterium Alteromonas infernus, Carbohydr. Res. 339(14), 2371–2380 (2004)
G. Raguenes, P. Pignet, G. Gauthier, A. Peres, R. Christen, H. Rougeaux, G. Barbier, J. Guezennec: Description of a new polymer-secreting bacterium from a deep-sea hydrothermal vent, Alteromonas macleodii subsp. fijiensis, and preliminary characterization of the polymer, Appl. Environ. Microbiol. 62(1), 67–73 (1996)
H. Rougeaux, P. Talaga, R.W. Carlson, J. Guezennec: Structural studies of an exopolysaccharide produced by Alteromonas macleodii subsp. fijiensis originating from a deep-sea hydrothermal vent, Carbohydr. Res. 312(1–2), 53–59 (1998)
T. Le Costaouec, S. Cérantola, B. Ropartz, J. Ratiskol, C. Sinquin, S. Colliec-Jouault, C. Boisset: Structural data on a bacterial exopolysaccharide produced by a deep-sea Alteromonas macleodii strain, Carbohydr. Polym. 90, 49–59 (2012)
M.A. Cambon-Bonavita, G. Raguenes, J. Jean, P. Vincent, J. Guezennec: A novel polymer produced by a bacterium isolated from a deep-sea hydrothermal vent polychaete annelid, J. Appl. Microbiol. 93(2), 310–315 (2002)
G. Dubreucq, B. Domon, B. Fournet: Structure determination of a novel uronic acid residue isolated from the exopolysaccharide produced by a bacterium originating from deep sea hydrothermal vents, Carbohydr. Res. 290(2), 175–181 (1996)
A. Arena, T.L. Maugeri, B. Pavone, D. Iannello, C. Gugliandolo, G. Bisignano: Antiviral and immunoregulatory effect of a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis, Int. Immunopharmacol. 6(1), 8–13 (2006)
A. Poli, G. Anzelmo, B. Nicolaus: Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities, Mar. Drugs 8(6), 1779–1802 (2010)
M.C. Manca, L. Lama, R. Improta, E. Esposito, A. Gambacorta, B. Nicolaus: Chemical composition of two exopolysaccharides from Bacillus thermoantarcticus, Appl. Environ. Microbiol. 62(9), 3265–3269 (1996)
M. Kambourova, R. Mandeva, D. Dimova, A. Poli, B. Nicolaus, G. Tommonaro: Production and characterization of a microbial glucan, synthesized by Geobacillus tepidamans V264 isolated from Bulgarian hot spring, Carbohydr. Polym. 77(2), 338–343 (2009)
A. Arena, C. Gugliandolo, G. Stassi, B. Pavone, D. Iannello, G. Bisignano, T.L. Maugeri: An exopolysaccharide produced by Geobacillus thermodenitrificans strain B3-72: Antiviral activity on immunocompetent cells, Immunol. Lett. 123(2), 132–137 (2009)
V. Schiano Moriello, L. Lama, A. Poli, C. Gugliandolo, T.L. Maugeri, A. Gambacorta, Nicolaus: Production of exopolysaccharides from a thermophilic microorganism isolated from a marine hot spring in flegrean areas, J. Ind. Microbiol. Biotechnol. 30, 95–101 (2003)
H.K. Lee, J. Chun, E.Y. Moon, S.H. Ko, D.S. Lee, H.S. Lee, K.S. Bae: Hahella chejuensis gen. nov., sp. nov., an extracellular-polysaccharide-producing marine bacterium, Int. J. Syst. Evol. Microbiol. 51, 661–666 (2001)
A. Poli, H. Kazak, B. Gürleyendağ, G. Tommonaro, G. Pieretti, E.T. Öner, B. Nicolaus: High level synthesis of levan by a novel Halomonas species growing on defined media, Carbohydr. Polym. 78(4), 651–657 (2009)
I. Llamas, H. Amjres, J.A. Mata, E. Quesada, V. Béjar: The potential biotechnological applications of the exopolysaccharide produced by the halophilic bacterium Halomonas almeriensis, Molecules 17(6), 7103–7120 (2012)
J.A. Mata, V. Béjar, I. Llamas, S. Arias, P. Bressollier, R. Tallon, M.C. Urdaci, E. Quesada: Exopolysaccharides produced by the recently described halophilic bacteria Halomonas ventosae and Halomonas anticariensis., Res. Microbiol. 157(9), 827–835 (2006)
S. Arias, A. del Moral, M.R. Ferrer, R. Tallon, E. Quesada, V. Bejar: Mauran, an exopolysaccharide produced by the halophilic bacterium Halomonas maura, with a novel composition and interesting properties for biotechnology, Extremophiles 7(4), 319–326 (2003)
C. Mancuso Nichols, S. Garon Lardière, J.P. Bowman, P.D. Nichols, J.A.E. Gibson, J. Guézennec: Chemical characterization of exopolysaccharides from Antarctic marine bacteria, Microb. Ecol. 49, 578–589 (2005)
C.M. Nichols, J.P. Bowman, J. Guezennec: Olleya marilimosa gen. nov., sp. nov., an exopolysaccharide-producing marine bacterium from the family Flavobacteriaceae, isolated from the Southern Ocean, Int. J. Syst. Evol, Microbiol. 55(4), 1557–1561 (2005)
G. Raguenes, X. Moppert, L. Richert, J. Ratiskol, C. Payri, B. Costa, J. Guezennec: A novel exopolymer-producing bacterium, Paracoccus zeaxanthinifaciens subsp. payriae, isolated from a Kopara mat located in Rangiroa, an atoll of French Polynesia, Curr. Microbiol. 49, 145–151 (2004)
E.L. Nazarenko, N.A. Komandrova, R.P. Gorshkova, S.V. Tomshich, V.A. Zubkov, M. Kilcoyne, A.V. Savage: Structures of polysaccharides and oligosaccharides of some Gram-negative marine Proteobacteria, Carbohydr. Res. 338(23), 2449–2457 (2003)
H. Rougeaux, J. Guezennec, R.W. Carlson, N. Kervarec, R. Pichon, P. Talaga: Structural determination of the exopolysaccharide of Pseudoalteromonas strain HYD 721 isolated from a deep-sea hydrothermal vent, Carbohydr. Res. 315(3–4), 273–285 (1999)
P. Saravanan, S. Jayachandran: Preliminary characterization of exopolysaccharides produced by a marine biofilm-forming bacterium Pseudoalteromonas ruthenica (SBT 033), Lett. Appl. Microbiol. 46(1), 1–6 (2008)
G. Qin, L. Zhu, X. Chen, P.G. Wang, Y. Zhang: Structural characterization and ecological roles of a novel exopolysaccharide from the deep-sea psychrotolerant bacterium Pseudoalteromonas sp., SM9913, Microbiology 153, 1566–1572 (2007)
D.D. Sledjeski, R.M. Weiner: Production and characterization of monoclonal antibodies specific for Shewanella colwelliana exopolysaccharide, Appl. Environ. Microbiol. 59(5), 1565–1572 (1993)
K.D. Rinker, R.M. Kelly: Effect of carbon and nitrogen sources on growth dynamics and exopolysaccharide production for the hyperthermophilic archaeon Thermococcus litoralis and bacterium Thermotoga maritima, Biotechnol. Bioeng. 69(5), 537–547 (2000)
J. Muralidharan, S. Jayachandran: Physicochemical analyses of the exopolysaccharides produced by a marine biofouling bacterium, Vibrio alginolyticus, Process Biochem. 38(6), 841–847 (2003)
G. Raguenes, R. Christen, J. Guezennec, P. Pignet, G. Barbier: Vibrio diabolicus sp. nov., a new polysaccharide-secreting organism isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana, Int. J. Syst. Bacteriol. 47(4), 989–995 (1997)
H. Rougeaux, N. Kervarec, R. Pichon, J. Guezennec: Structure of the exopolysaccharide of Vibrio diabolicus isolated from a deep-sea hydrothermal vent, Carbohydr. Res. 322(1–2), 40–45 (1999)
P.V. Bramhachari, P.B. Kavi Kishor, R. Ramadevi, R. Kumar, B. Rama Rao, S.K. Dubey: Isolation and characterization of mucous exopolysaccharide (EPS) produced by Vibrio furnissii strain VB0S3, J. Microbiol. Biotechnol. 17(1), 44–51 (2007)
P.V. Bramhachari, S.K. Dubey: Isolation and characterization of exopolysaccharide produced by Vibrio harveyi strain VB23, Lett. Appl. Microbiol. 43(5), 571–577 (2006)
K. Kavita, A. Mishra, B. Jha: Isolation and physico-chemical characterisation of extracellular polymeric substances produced by the marine bacterium Vibrio parahaemolyticus, Biofouling 27, 309–317 (2011)
G.A. Morris, P. Li, M. Puaud, Z. Liu, J.R. Mitchell, S.E. Harding: Hydrodynamic characterisation of the exopolysaccharide from the halophilic cyanobacterium Aphanothece halophytica GR02: A comparison with xanthan Original Research Article, Carbohydr. Polym. 44(3), 261–268 (2001)
W.F. Zheng, C.F. Chen, Q.P. Cheng, Y.Q. Wang, C.C. Chu: Oral administration of exopolysaccharide from Aphanothece halophytica (Chroococcales) significantly inhibits influenza virus (H1N1)-induced pneumonia in mice, Int. Immunopharmacol. 6(7), 1093–1099 (2006)
Z. Chi, C.D. Su, W.D. Lu: A new exopolysaccharide produced by marine Cyanothece sp., 113, Bioresour. Technol. 98(6), 1329–1332 (2007)
V. Shah, A. Ray, N. Garg, D. Madamwar: Characterization of the extracellular polysaccharide produced by a marine cyanobacterium, Cyanothece sp. ATCC51142, and its exploitation toward metal removal from solutions, Curr. Microbiol. 40, 274–278 (2000)
J.-B. Lee, T. Hayashi, K. Hayashi, U. Sankawa: Structural analysis of calcium spirulan (Ca-SP)-derived oligosaccharides using electrospray ionization mass spectrometry, J. Nat. Prod. 63(1), 136–138 (2000)
B. Nicolaus, A. Panico, L. Lama, I. Romano, M.C. Manca, A. De Giulio, A. Gambacorta: Chemical composition and production of exopolysaccharides from representative members of heterocystous and non-heterocystous cyanobacteria, Phytochemistry 52(4), 639–647 (1999)
L.A.S. Parolis, H. Parolis, N.A. Paramonov, I.F. Boán, J. Antón, F. Rodríguez-Valera: Structural studies on the acidic exopolysaccharide from Haloferax denitrificans ATCC 35960, Carbohydr. Res. 319(1–4), 133–140 (1999)
N.A. Paramonov, L.A.S. Parolis, H. Parolis, I.F. Boán, J. Antón, F. Rodrìguez-Valera: The structure of the exocellular polysaccharide produced by the Archaeon Haloferax gibbonsii (ATCC 33959), Carbohydr. Res. 309(1), 89–94 (1998)
H. Parolis, L.A.S. Parolis, I.F. Boan, F. Rodriguez-Valera, G. Widmalm, M.C. Manca, P.-E. Jansson, I.W. Sutherland: The structure of the exopolysaccharide produced by the halophilic Archaeon Haloferax mediterranei strain R4 (ATCC 33500), Carbohydr. Res. 295(0), 147–156 (1996)
B. Nicolaus, M.C. Manca, I. Ramano, L. Lama: Production of an exopolysaccharide from two thermophilic archaea belonging to the genus Sulfolobus, FEMS Microbiol. Lett. 109(2–3), 203–206 (1993)
R.E. Reyes, A.A. Andrade, R.C. Jiménez, C.R. Gonzalez, M.O. Herrera: Mechanisms of o-antigen structural variation of bacterial lipopolysaccharide (LPS). In: The Complex World of Polysaccharides, ed. by D.N. Karunaratne (InTech, Rijeka 2012) pp. 71–98
I.W. Sutherland, A.H. Rose, D.W. Tempest: Bacterial exopolysaccharides, Adv. Microb. Physiol. 8, 143–213 (1972)
I.W. Sutherland: Bacterial exopolysaccharides. In: Comprehensive Glycoscience, from Chemistry to Systems Biology, ed. by J.P. Kamerling (Elsevier, Amsterdam 2007) pp. 521–558
I.W. Sutherland: Microbial polysaccharides from Gram-negative bacteria, Int. Dairy J. 11(9), 663–674 (2001)
A. Cesàro, B. Bellich, M. Borgogna: Biophysical functionality in polysaccharides: From Lego-blocks to nano-particles, Eur. Biophys. J. 41(4), 379–395 (2012)
P. Laurienzo: Marine Polysaccharides in pharmaceutical applications: An overview, Mar. Drugs 8(9), 2435–2465 (2010)
N.O. Keyhani, S. Roseman: Physiological aspects of chitin catabolism in marine bacteria, Biochim. Biophys. Acta General Subj. 1473(1), 108–122 (1999)
P.K. Dutta, J. Dutta, V.S. Tripathi: Chitin and chitosan: Chemistry, properties and applications, J. Sci. Ind. Res. 63, 20–31 (2004)
S. Bielecki, A. Krystynowicz, M. Turkiewicz, H. Kalinowska: Bacterial cellulose. In: Biopolymers Online, ed. by A. Steinbüchel (Wiley, Weinheim 2005) pp. 37–90
C. Castro, R. Zuluaga, J.-L. Putaux, G. Caro, I. Mondragon, P. Ganan: Structural characterization of bacterial cellulose produced by Gluconacetobacter swingsii sp., from Colombian agroindustrial wastes, Carbohydr. Polym. 84(1), 96–102 (2011)
K. Kaur, V. Dattajirao, V. Shrivastava, U. Bhardwaj: Isolation and Characterization of Chitosan-Producing Bacteria from Beaches of Chennai, India, Enzym. Res. 2012, 1–6 (2012)
L.-S. Kim, S.-J. Hong, M.-K. Son, Y.-H. Lee: Polymeric and Compositional Properties of Novel Extracellular Microbial Polyglucosamine Biopolymer from New Strain of Citrobacter sp. BL-4, Biotechnol. Lett. 28(4), 241–245 (2006)
H. Rougeaux, M. Guezennec, L. Mao Che, C. Payri, E. Deslandes, J. Guezennec: Microbial communities and exopolysaccharides from Polynesian mats, Mar. Biotechnol. 3, 181–187 (2001)
P. Vincent, P. Pignet, F. Talmont, L. Bozzi, B. Fournet, J. Guezennec: Production and characterization of an exopolysaccharide excreted by a deep-sea hydrothermal vent bacterium isolated from the polychaete annelid Alvinella pompejana, Appl. Environ. Microb. 60, 4134–4141 (1994)
L. De Vuyst, F. De Vin, F. Vaningelgem, B. Degeest: Recent developments in the biosynthesis and applications of heteropolysaccharides from lactic acid bacteria, Int. Dairy J. 11(9), 687–707 (2001)
J.G. Guezennec, P. Pignet, G. Raguenes, E. Deslandes, Y. Lijour, E. Gentric: Preliminary chemical characterization of unusual eubacterial exopolysaccharides of deep-sea origin, Carbohydr. Polym. 24(4), 287–294 (1994)
H. Rougeaux, R. Pichon, N. Kervarec, G.H.C. Raguenes, J.G. Guezennec: Novel bacterial exopolysaccharides from deep-sea hydrothermal vents, Carbohydr. Polym. 31(4), 237–242 (1996)
G. Raguenes, M.A. Cambon-Bonavita, J.F. Lohier, C. Boisset, J. Guezennec: A novel, highly viscous polysaccharide excreted by an Alteromonas isolated from a deep-sea hydrothermal vent shrimp, Curr. Microbiol. 46, 448–452 (2003)
I.C. Boels, R.V. Kranenburg, J. Hugenholtz, M. Kleerebezem, W.M. de Vos: Sugar catabolism and its impact on the biosynthesis and engineering of exopolysaccharide production in lactic acid bacteria, Int. Dairy J. 11(9), 723–732 (2001)
F.H. Yildiz, K.L. Visick: Vibrio biofilms: So much the same yet so different, Trends Microbiol. 17(3), 109–118 (2009)
P. Lembre, C. Lorentz, P. Di Martino: Exopolysaccharides of the biofilm matrix: A complex biophysical world. In: The Complex World of Polysaccharides, ed. by D.N. Karunaratne (InTech, Rijeka 2012) pp. 371–392
M. Neave, C. Streten-Joyce, C. Glasby, K. McGuinness, D. Parry, K. Gibb: The bacterial community associated with the marine polychaete Ophelina sp.1 (Annelida: Opheliidae) is altered by copper and zinc contamination in sediments, Microb. Ecol. 63(3), 639–650 (2012)
P. Monsan, S. Bozonnet, C. Albenne, G. Joucla, R.-M. Willemot, M. Remaud-Simeon: Homopolysaccharides from lactic acid bacteria, Int. Dairy J. 11(9), 675–685 (2001)
C. Whitfield: Biosynthesis and assembly of capsular polysaccharides in Escherichia coli, Annu. Rev. Biochem. 75, 39–68 (2006)
R.V. Kranenburg, J.D. Marugg, I.I. Van Swam, N.J. Willem, W.M. De Vos: Molecular characterization of the plasmid-encoded eps gene cluster essential for exopolysaccharide biosynthesis in Lactococcus lactis, Mol. Microbiol. 24(2), 387–397 (1997)
W.W. Wakarchuk: Application of bacterial glycosyltransferases in the synthesis of bioactive glycans. In: Bacterial Glycomics, Current Research, Technology and Applications, ed. by C.W. Reid, S.M. Twine, A.N. Reid (Caister Academic, Norfolk 2012) pp. 213–228
L. Cuthbertson, V. Kos, C. Whitfield: ABC transporters involved in export of cell surface glycoconjugates, Microbiol. Mol. Biol. Rev. 74(3), 341–362 (2010)
H.C. Wong, A.L. Fear, R.D. Calhoon, G.H. Eichinger, R. Mayer, D. Amikam, M. Benziman, D.H. Gelfand, J.H. Meade, A.W. Emerick: Genetic organization of the cellulose synthase operon in Acetobacter xylinum, Proc. Natl. Acad. Sci. USA 87(20), 8130–8134 (1990)
P.L. DeAngelis, W. Jing, R.R. Drake, A.M. Achyuthan: Identification and Molecular Cloning of a Unique Hyaluronan Synthase from Pasteurella multocida, J. Biol. Chem. 273(14), 8454–8458 (1998)
D. Cimini, M. De Rosa, C. Schiraldi: Production of glucuronic acid-based polysaccharides by microbial fermentation for biomedical applications, Biotechnol. J. 7(2), 237–250 (2011)
G. Gomez d'Ayala, M. Maklinconico, P. Laurienzo: Marine derived polysaccharides for biomedical applications: Chemical modification approaches, Molecules 13, 2069–2106 (2008)
B.H.A. Rehm, S. Valla: Bacterial alginates: Biosynthesis and applications, Appl. Microbiol. Biotechnol. 48(3), 281–288 (1997)
S.K. Satpute, I.M. Banat, P.K. Dhakephalkar, A.G. Banpurkar, B.A. Chopade: Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms, Biotechnol. Adv. 28(4), 436–450 (2010)
U. Bhaskar, E. Sterner, A. Hickey, A. Onishi, F. Zhang, J. Dordick, R. Linhardt: Engineering of routes to heparin and related polysaccharides, Appl. Microbiol. Biotechnol. 93(1), 1–16 (2012)
B. Mulloy: The specificity of interactions between proteins and sulfated polysaccharides, An. Acad. Bras. Cienc. 77, 651–664 (2005)
Z. Shriver, S. Raguram, R. Sasisekharan: Glycomics: A pathway to a class of new and improved therapeutics, Nat. Rev. 3(10), 863–873 (2004)
J.E. Turnbull, R.A. Field: Emerging glycomics technologies, Nat. Chem. Biol. 3(2), 74–77 (2007)
F. Donot, A. Fontana, J.C. Baccou, S. Schorr-Galindo: Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction, Carbohydr. Polym. 87(2), 951–962 (2012)
C. Schiraldi, A. La Gatta, M. De Rosa: Biotechnological production and application of hyaluronan. In: Biopolymers, ed. by M. Elnashar (InTech, Rijeka 2010) pp. 387–412
A.D. Welman, I.S. Maddox: Exopolysaccharides from lactic acid bacteria: Perspectives and challenges, Trends Biotechnol. 21(6), 269–274 (2003)
Z. Persin, K. Stana-Kleinschek, T.J. Foster, J.E.G. van Dam, C.G. Boeriu, P. Navard: Challenges and opportunities in polysaccharides research and technology: The EPNOE views for the next decade in the areas of materials, food and health care, Carbohydr. Polym. 84(1), 22–32 (2011)
A. Thibodeau, A. Takeoka: The applications and functions of new exopolysaccharide Deepsane from the deepest oceans, Fragr. J. 34(3), 61–68 (2006)
M. Borgogna, B. Bellich, A. Cesaro: Marine polysaccharides in microencapsulation and application to aquaculture: From sea to sea, Mar. Drugs 9, 2572–2604 (2011)
W. Wang, S.-X. Wang, H.-S. Guan: The antiviral activities and mechanisms of marine polysaccharides: An overview, Mar. Drugs 10(12), 2795–2816 (2012)
K. Hayashi, T. Hayashi, I. Kojima: A natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-Herpes simplex virus and anti-human immunodeficiency virus activities, AIDS Res. Hum. Retrovir. 12(15), 1463–1471 (1996)
O. Bagasra, P. Whittle, B. Heins, R.J. Pomerantz: Anti-human immunodeficiency virus type 1 activity of sulfated monosaccharides: Comparison with sulfated polysaccharides and other polyions, J. Infect. Dis. 164(6), 1082–1092 (1991)
G.M.T. Calazans, C.E. Lopes, R. Lima, F. de França: Antitumour activities of levans produced by Zymomonas mobilis strains, Biotechnol. Lett. 19(1), 19–21 (1997)
E. Esawy: Antiviral levans from Bacillus spp. Isolated from Honey. In: The Complex World of Polysaccharides, ed. by D.N. Karunaratne (InTech, Rijeka 2012) pp. 195–214
W.F. Vann, M.A. Schmidt, B. Jann, K. Jann: The structure of the capsular polysaccharide (K5 Antigen) of urinary-tract-infective Escherichia coli 010:K5:H4, Eur. J. Biochem. 116(2), 359–364 (1981)
P.L. deAngelis: Glycoaminoglycan polysacchcaride biosynthesis and production: Today and tomorrow, Appl. Microbiol. Biotechnol. 94, 295–305 (2012)
R. Lever, C.P. Page: Novel drug development opportunities for heparin, Nat. Rev. Drug Discov. 1(2), 140–148 (2002)
N. Afratis, C. Gialeli, D. Nikitovic, T. Tsegenidis, E. Karousou, A.D. Theocharis, M.S. Pavão, G.N. Tzanakakis, N.K. Karamanos: Glycosaminoglycans: Key players in cancer cell biology and treatment, FEBS J. 279(7), 1177–1197 (2012)
K. Senni, J. Pereira, F. Gueniche, C. Delbarre-Ladrat, C. Sinquin, J. Ratiskol, G. Godeau, A.-M. Fischer, D. Helley, S. Colliec-Jouault: Marine Polysaccharides: A source of bioactive molecules for cell therapy and tissue engineering, Mar. Drugs 9(9), 1664–1681 (2011)
J. Choay, M. Petitou, J.C. Lormeau, P. Sina$\text{\"{y}}$, B. Casu, G. Gatti: Structure-activity relationship in heparin: A synthetic pentasaccharide with high affinity for antithrombin III and eliciting high anti-factor Xa activity, Biochem. Biophys. Res. Commun. 116(2), 492–499 (1983)
B. Widner, R. Behr, S. Von Dollen, M. Tang, T. Heu, A. Sloma, D. Sternberg, P.L. DeAngelis, P.H. Weigel, S. Brown: Hyaluronic production in Bacillus subtilis, Appl. Environ. Microbiol. 71, 3747–3752 (2005)
L.-J. Chien, C.-K. Lee: Hyaluronic production by recombinant Lactococcus lactis, Appl. Microbiol. Biotechnol. 77, 339–346 (2007)
G. Kogan, L. Soltes, R. Stern, P. Gemeiner: Hyaluronic acid: A natural biopolymer with a broad range of biomedical and industrial applications, Biotechnol. Lett. 29, 17–25 (2007)
P. Zanchetta, N. Lagarde, J. Guezennec: A new bone-healing material: A hyaluronic acid-like bacterial exopolysaccharide, Calcif. Tissue Int. 72(1), 74–79 (2003)
Z. Wang, M. Ly, F. Zhang, W. Zhong, A. Suen, A.M. Hickey, J.S. Dordick, R.J. Linhardt: E. coli K5 fermentation and the preparation of heparosan, a bioengineered heparin precursor, Biotechnol. Bioeng. 107(6), 964–973 (2010)
S. Alban, A. Schauerte, G. Franz: Anticoagulant sulfated polysaccharides: Part I. Synthesis and structure-activity relationships of new pullulan sulfates, Carbohydr. Polym. 47, 267–276 (2002)
D. Cimini, O. Restaino, A. Catapano, M. De Rosa, C. Schiraldi: Production of capsular polysaccharide from Escherichia coli K4 for biotechnological applications, Appl. Microbiol. Biotechnol. 85(6), 1779–1787 (2010)
Z. Wang, B. Yang, Z. Zhang, M. Ly, M. Takieddin, S. Mousa, J. Liu, J. Dordick, R. Linhardt: Control of the heparosan N-deacetylation leads to an improved bioengineered heparin, Appl. Microbiol. Biotechnol. 91(1), 91–99 (2011)
B. Casu, G. Grazioli, N. Razi, M. Guerrini, A. Naggi, G. Torri, P. Oreste, F. Tursi, G. Zopetti, U. Lindahl: Heparin-like compounds prepared by chemical modification of capsular polysaccharide from E. coli K5, Carbohydr. Res. 263, 271–284 (1994)
A. Naggi, G. Torri, B. Casu, P. Oreste, G. Zopetti, J.P. Li, U. Lindahl: Toward a biotechnological heparin through combined chemical and enzymatic modification of the Escherichia coli K5 polysaccharide, Semin. Thromb. Hemost. 27(5), 437–443 (2001)
U. Lindahl, J.P. Li, M. Kusche-Gullberg, M. Salmivirta, S. Alaranta, T. Veromaa, J. Emeis, I. Roberts, C. Taylor, P. Oreste, G. Zopetti, A. Naggi, G. Torri, B. Casu: Generation of neoheparin from E. coli K5 capsular polysaccharide, J. Med. Chem. 48, 349–352 (2005)
O.M. Hanniffy, A.S. Shashkov, S.N. Senchenkova, S.V. Tomshich, N.A. Komandrova, L.A. Romanenko, Y. Knirel, A.V. Savage: Structure of a highly acidic O-specific polysaccharide of lipopolysaccharide of Pseudoalteromonas haloplanktis KMM 223 (44-1) containing L-iduronic acid and D-QuiNHb4NHb, Carbohydr. Res. 307, 291–298 (1998)
E. Rederstorff, A. Fatimi, C. Sinquin, J. Ratiskol, C. Merceron, C. Vinatier, P. Weiss, S. Colliec-Jouault: Sterilization of exopolysaccharides produced by deep-sea bacteria: Impact on their stability and degradation, Mar. Drugs 9(2), 224–241 (2011)
S. Matou, S. Colliec Jouault, D. Helley, J. Ratiskol, C. Sinquin, C. Boisset, J. Guezennec, A.M. Fischer: Utilisation de dérivés polysaccharidiques hautement sulfatés et de faible masse molaire pour moduler l'angiogénese, FR 2 871 379 - A1 (2004)
S. Colliec-Jouault, L. Chevolot, D. Helley, J. Ratiskol, A. Bros, C. Sinquin, O. Roger, A.M. Fischer: Characterization, chemical modifications and in vitro anticoagulant properties of an exopolysaccharide produced by Alteromonas infernus, Biochim. Biophys. Acta 1528(2–3), 141–151 (2001)
C. Merceron, S. Portron, C. Vignes-Colombeix, E. Rederstorff, M. Masson, J. Lesoeur, S. Sourice, C. Sinquin, S. Colliec Jouault, P. Weiss, C. Vinatier, J. Guicheux: Pharmacological modulation of human mesenchymal stem cell chondrogenesis by a chemically oversulfated polysaccharide of marine origin: Potential application to cartilage regenerative medecine, Stem Cells 30, 471–480 (2012)
N. Mestechkina, V. Shcherbukhin: Sulfated polysaccharides and their anticoagulant activity: A review, Appl. Biochem. Microbiol. 46(3), 267–273 (2010)
C. Mahner, M.D. Lechner, E. Nordmeier: Synthesis and characterisation of dextran and pullulan sulphate, Carbohydr. Res. 331(2), 203–208 (2001)
J. Chen, F.Y. Avci, E.M. Munoz, L.M. McDowell, M. Chen, L.C. Pedersen, L. Zhang, R.J. Linhardt, J. Liu: Enzymatic redesigning of biologically active heparan sulfate, J. Biol. Chem. 280(52), 42817–42825 (2005)
J. Chen, M.B. Duncan, K. Carrick, R.M. Pope, J. Liu: Biosynthesis of 3-O-sulfated heparan sulfate: Unique substrate specifictiy of heparan sulfate 3-O-sulfotransferase isoform 5, Glycobiology 13(11), 785–794 (2003)
M.D. Burkart, M. Izumi, E. Chapman, C.H. Lin, C.H. Wong: Regeneration of PAPS for the enzymatic synthesis of sulfated oligosaccharides, J. Organ. Chem. 65, 5565–5574 (2000)
A.L. Waters, R.T. Hill, A.R. Place, M.T. Hamann: The expanding role of marine microbes in pharmaceutical development, Curr. Opin. Biotechnol. 21(6), 780–786 (2010)
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Delbarre-Ladrat, C., Boursicot, V., Colliec-Jouault, S. (2015). Marine-Derived Exopolysaccharides. In: Kim, SK. (eds) Springer Handbook of Marine Biotechnology. Springer Handbooks. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-53971-8_39
Download citation
DOI: https://doi.org/10.1007/978-3-642-53971-8_39
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-53970-1
Online ISBN: 978-3-642-53971-8
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)