
Abstract
In the last years, there is a growing awareness regarding the role of the microbiota in various chronic inflammatory disorders including cancers. The intestinal microbiome is not only regulating the spontaneous course of malignancies of the digestive tract but also acts at a distant site on extraintestinal neoplasia, to play a beneficial or a detrimental role, depending on the context. By providing pathogen and damage-associated molecular patterns and by activating local and distant antigen-presenting cells, our commensals modulate not only the gut, the systemic inflammatory, and immune tonus but also the tumor microenvironment and anticancer-specific adaptive immune responses. Additionally, therapeutic interventions alter the delicate balance between the epithelium, the microbial community, and the lamina propria immunity, influencing the clinical outcome. This chapter will focus on the impact of the intestinal composition in the bioactivity and therapeutic efficacy of distinct compounds (alkylating agents, platinum salts, radiotherapy, conditioning regimen for stem cell transplantation, immunotherapies, and immune checkpoint blockers). This era of research opens novel therapeutic avenues that aim to restore gut eubiosis for a better clinical outcome in cancer patients.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

1 Introduction
Only recently, the scientific community gained insights on the importance of the intestinal resident microflora for the host’s health and pathophysiology [1,2,3].
The gut microflora exerts a variety of different fundamental functions, such as the degradation of nutrients to provide energy source, the elimination of xenobiotics, the education of the immune system, the growth and differentiation of epithelial cells of the barrier, the intestinal peristaltism, and the production of antimicrobial peptides to eradicate pathogens and ensure colonization resistance [4]. The intestine represents the largest compartment of the immune system. It is exposed to food and commensal antigens that our body needs to tolerate and is the portal of entry for many pathogens. The numbers of bacteria increase going down the gastrointestinal tract, with up to 1012 per ml in the colon. Aerobic species are prevalent in the upper small intestine, whereas anaerobic bacteria dominate in the colon. Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria are the most prevalent, together with members of the Archaea kingdom [4].
Gut microbiota plays a crucial role in modulating innate and acquired immune responses and thus interferes with the delicate balance between inflammation and tolerance. Recent advances in sequencing technologies allowed the deep characterization of the human microbiota, thus greatly improving our knowledge on the role of the microbiome in human health and disease. Human microbiome project consortium studies (MetaHIT and HMP) demonstrated that healthy individuals have not only a high degree of bacterial diversity, dependent on their habitat (intestine, oral cavity, skin, or vagina), but that there is also a remarkable interindividual variability at the level of species [5, 6]. Despite the species diversity found at various locations, there is a certain constancy that preserves both the function and the bacterial gene profiling associated to specific tissue sites. Hence, the anaerobic Firmicutes/Bacteroidetes spp. dominate the intestine, whereas Actinobacteria and Proteobacteria spp. are highly abundant in the skin [7]. Defining the “normal microbiome” is a difficult task, considering that the same microbe may behave as commensal or as pathogen depending on the dietary components, the nutritional milieu, the genetic background of its host, and a potential coinfection. Nonetheless, “commensal” bacteria are expected to contribute to immune homeostasis, whereas “pathobionts” are associated with immune reactivities against themselves [8]. Perhaps paradoxically, it is possible to find in serum of healthy subjects antibodies specific to commensals as well as circulating T cells able to react against nonpathogenic bacteria [9,10,11].
Gut colonization at birth is indispensable to trigger the maturation of the mucosal immune system. At this phase, the intestinal microbiota plays a crucial role in setting up innate and acquired immune responses “imprinted” for life, interfering in the balance between inflammation, infection, and tolerance. Germ-free mice have hypoplastic Peyer’s patches, lack isolated lymphoid follicles in the intestine, and have low levels of IgM natural antibodies in the serum, this phenotype being reverted by gut colonization [12]. Undoubtedly, colonization of the distal small intestine by segmented filamentous bacteria (SFB) is crucial for the development of resident lamina propria dendritic cells secreting IL-6 and IL-22 and the concomitant differentiation of Th17-T regulatory (Treg) cells in the newborn gut [13]. The discovery of a link between defined members of the microbiota (such as Bacteroides fragilis and TLR2) and the induction of Treg cells by Clostridiaceae generated a huge interest. However, the homeostasis of the colonic Treg compartment relies more on the synergistic effect of different bacterial strains and an interkingdom ecosystem creating a balanced microenvironment able to sustain the generation and maintenance of the anti-inflammatory milieu [14,15,16].
2 Evaluating Gut Dysbiosis in Cancer Bearers
Our knowledge of the genetic and functional diversity in gut microbes is far from being complete. Old methods consisting in qPCR or FISH analyses were neither comprehensive, specific, nor sensitive enough. The burst of recent knowledge emerged from the targeted 16S rRNA gene pyrosequencing followed by metagenomic shotgun sequencing, a costly method with heavy data mining. The catalog of reference genes in the human gut microbiome was reported, gathering data from MetaHIT, Human Microbiome Project (HMP), and a large Chinese diabetes study, as well as >500 sequenced genomes of gut-related bacteria and archaea. This nonredundant reference catalog of 9,879,896 genes is freely accessible through the website (http://meta.genomics.cn), and the data are deposited in the GigaScience Database. We may have reached saturated coverage of core gene content and functions. Of note, the number of genes present in more than 50% of the subjects remained below 300,000, pointing out the dominance of individual-specific genes. The individual-specific genes are enriched in the categories cell wall/membrane/envelope biogenesis and DNA replication, recombination, and repair. The common genes are enriched in functions such as signal transduction mechanism, energy production, carbohydrate transport and metabolism, and amino acid transport and metabolism [17].
Several groups investigated the gut microbiota in healthy individuals and in cancer bearers to establish microbe-cancer relationships. For example, Goedert et al. analyzed gut microbiota using 16S rRNA sequencing in postmenopausal women. The authors observed a lower alpha diversity (number of different taxa) and altered beta diversity (between-subject community composition) in subjects suffering from a breast cancer, when compared to healthy individuals.
Dejea et al. analyzed colorectal mucosa from subjects bearing or not colorectal cancer (CRC), using FISH and 16S rRNA sequencing. They demonstrated that bacterial biofilm presence, but no consistent bacterial genus, was associated with colorectal cancers [18]. Other studies, using 16S rRNA or metagenomic sequencing, revealed case-control discrepancies in the gut microbiota composition, showing that feces from CRC subjects display higher prevalence and levels of Fusobacterium and Porphyromonas compared to controls [19, 20]. However, across studies many differences were observed in taxa that diverged between cases and controls. This could be related to characteristics of the population studied (diet, medication) as well as to setting parameters (experimental methods or case-control matching). Furthermore, in complex ecosystems, current metagenomic studies are unable to detect bacteria at concentrations below 105 bacteria/gram. Moreover, only 15% of identified species were concomitantly detected by the culturomics and the metagenomics, highlighting the complementarity between culture-dependent and culture-independent methods. The diversification of culture conditions coupled with the identification of the bacterial repertoire by matrix-assisted laser desorption ionization-time-of-flight mass spectrometry [MALDI-TOF MS] enabled significant progresses in taxonogenomics. These efforts will help defining “cancer-associated gut fingerprints” in the near future in different cohorts of cancer types to establish the impact of cancer topography on gut dysbiosis.
3 Microbiome and Cancer Incidence
Commensal microbial communities inhabiting the intestine, as well as other places in the body, appear to play an unappreciated role in intestinal and extraintestinal carcinogenesis by providing yet to be characterized environmental signals [21]. Pioneering studies performed in germ-free, gnotobiotic, or antibiotic-treated rodents revealed an unsuspected role for commensals in tumorigenesis—irrespective of the role of inflammation. In the genesis of colon cancer or hepatocarcinoma, microbes can be direct transforming agents [22, 23] by providing a toxic metabolite, an oncogenic product, or by inducing an inflammatory milieu which will culminate in genomic instability and/or DNA damage response and/or immune escape [24,25,26]. Commensals can also form cooperative biofilms that facilitate cross-feeding or cross-metabolism, redefining the cancer landscape [18, 27]. Recently, the development of extraintestinal (breast and ovarian) neoplasia was linked to TLR5-mediated, IL-6- or IL-17-driven systemic inflammation provoked by intestinal microbes [28]. In contrast, other observations support a beneficial role for bacteria in combatting cancer. Prolonged antibiotic treatment with a combination of metronidazole and ciprofloxacine subsequently tripled breast cancer (BC) incidence in proto-oncogene HER2/neu-driven transgenic mice [29]. In humans, epidemiological studies suggested a dose-dependent association between antibiotic use and risk of BC [30]. Antibiotic use disrupted the intestinal microfloral metabolism of estrogens [31, 32]. Knekt et al. first reported an epidemiologic study on the association of antibiotic uptake and BC incidence. Women <50 who self-reported previous and/or present antibiotic use for urinary tract infections but did not harbor a bacteriuria had an elevated risk of breast cancer, compared with women without this usage [33].
By extension, associations between antibiotic exposure and cancer risk were reported in various cancers [34,35,36]. In contrast, the use of antivirals or antifungals did not affect the cancer risk [37]. Multiple studies reported an association between an altered composition of local microbiomes at portals of entree and cancer occurrence (such as oral in head and neck cancers [38], bronchial in NSCLC [39], intestinal and colorectal malignancies [40] and vaginal in cancer of the cervix [41]).
These studies are often flawed or biased by (a) the lack of rigorous methodologies (old methods of PCR versus 16S pyrosequencing analyses versus deep sequencing (metagenomics) versus culturomics), (b) overlooking lifestyle factors (such as social milieu, sedentary way of life, malnutrition, alcoholic or smoking behavior, etc.) that may modify the microbiome and increase cancer risk independently from each other, (c) the lack of a validation cohort (according to the REMARK criteria), and (d) the lack of cause-effect relationship between microbial dominance and the underlying pathophysiology. The current momentum for the impact of the gut microbiome in cancer will reinvigorate clinicians to launch prospective studies evaluating the impact of ATB uptake, as well as nutrition, genetic, and environmental factors in driving or influencing the course of oncogenesis processes.
4 Radiotherapy and Microbiota
The effects of radiotherapy on the microbiome and cancer immunotherapy have not been studied extensively. Most studies to date address radiation-induced bowel injury and its modulation by the host microbiome.
Crawford et al. reported first that the intestinal microbiome is an important modifier of radiation-induced intestinal injury, showing that germ-free animals are more resistant to radiation enteritis [42]. Microbial composition in mice was significantly altered by radiation. Modulation of the intestinal microbiome by fecal microbial transplantation (FMT) after whole-body irradiation increased survival and partly restored intestinal epithelial integrity through stimulation of angiogenesis [43, 44]. TLR signaling may play an important role in intestinal radioprotection. Ciorba et al. showed that Lactobacillus administration in a preclinical model of radiation-induced intestinal injury exerted protective effects through TLR2 and migration of Cox2-expressing mesenchymal stem cells [45]. Several clinical studies reported the modification of the intestinal microbiota post-radiation-induced intestinal injury [46,47,48]. Wang et al. showed that pelvic irradiation induced the release of inflammatory markers in the intestine of patients, associated with asthenia. Low microbial intestinal diversity also predicted radiation-induced colitis development [49]. Demers et al. showed that the administration of Bifidobacterium/Lactobacillus significantly lowered diarrhea [50]. However, no single bacterial strain was consistently shown to be beneficial in radiation-induced intestinal injury.
The beneficial role of the microbiome in the efficacy of radiotherapy remains to be demonstrated. There is good rationale to believe that disruption of the mucosal barrier facilitating the translocation of mucosal microbiota may activate the innate arm of immunity [51]. This was first reported by Paulos et al. showing that total body irradiation promoted a LPS-TLR4-dependent activation of antigen-presenting cells facilitating the efficacy of adoptive T cell transfer [52]. Radiation-induced intestinal injuries promoted the translocation of commensals and the release of LPS resulting in the priming of dendritic cells and activation of adoptively transferred T cells. This was correlated with the treatment efficacy and with the long-term cure of mice harboring melanoma B16F10 tumors. Thereby, modulating the homeostatic balance between the gut microbiota and the immune system could enhance cell-based tumor immunotherapy. Conversely, a subsequent study by Espinosa-Carrasco showed that LPS translocation only partly accounts for the effect seen by Paulos et al. [53]. To date, there is no data supporting the role of microbiome components as response modifiers in radio-immunotherapy.
5 Cyclophosphamide and Gut Microbiota
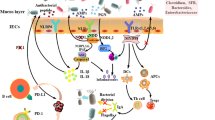
It is well known that chemotherapy can induce mucositis as well as neutropenia, two major side effects that could lead to the use of antibiotics and result in dysbiosis [54, 55]. The question of the impact of the gut microbiota on the efficacy on chemotherapeutic agents, and especially those capable of affecting the intestinal homeostasis, was addressed [56, 57]. The efficacy of cyclophosphamide (CTX), a DNA-alkylating agent belonging to the family of nitrogen with immunomodulatory and anti-angiogenesis properties currently used in clinics for the treatment of breast cancer, sarcoma, and pediatric malignancies [58], depends on its ability to induce the translocation of selective Gram-positive bacteria niching in the small intestine such as Enterococcus hirae or Lactobacillus johnsonii into secondary lymphoid organs. Indeed, CTX is responsible for disrupting the gut barrier integrity as well as intestinal homeostasis (both epithelial and immune compartments). Patients treated with CTX have a weaker intestinal barrier which breaks the tolerance toward the intestinal microbiota and leads to its immunization against some bacterial strains. This immunization is composed of CD4+ effector lymphocytes called “pathogenic Th17” producing IFNγ and IL-17, which helps tumor-infiltrating Th1 lymphocytes to control the tumor growth in mice. Interestingly, broad-spectrum antibiotics as well as vancomycin (which mainly kills Gram-positive bacteria) and colistin (which mainly eliminates Gram-negative bacteria) all compromised the polarization of pTh17 in the spleen and the full-blown anticancer activity of CTX in vivo in P815 mastocytoma- and MCA205 sarcoma-bearing mice, supporting the notion that the efficacy of cyclophosphamide was microbiota related [59]. Further study showed that among the translocating Gram-positive bacteria, E. hirae induces Th17 and Th1 CD4+ T lymphocytes and stimulates tumor-specific CD8+ T cells, as it reduces immunosuppressive intratumoral Tregs and IL-17-producing γδ T cells (Fig. 27.1). Mono-association of antibiotic-treated mice with E. hirae greatly improved tumor growth reduction by CTX, and this effect was blocked by antibodies depleting CD8+ T cells or neutralizing IFNγ, indicating that it requires an anticancer immune response. In parallel, we have demonstrated that the antitumor efficacy of CTX was greatly ameliorated in mice presenting a defect in NOD2 expression in intestinal epithelial cells, demonstrating the role of NOD2 as a gatekeeper and a gut immune checkpoint restricting the immunogenicity of E. hirae. The characterization of the gut microbiota in NOD2 animals highlighted that Gram-negative bacteria, from the Porphyromonadaceae family, dominated the colon, genus Barnesiella. Mice that were mono-associated with Barnesiella intestinihominis developed high numbers of polyfunctional CD4+, CD8+, or γδT cells, in the spleen and the tumor bed. B. intestinihominis-fed mice contained more abundant IFNγ-producing γδ T cells at the expense of immunosuppressive IL-17-producing γδ T cells compared with nonfed (or E. hirae-fed) animals. Moreover, CTX combined with B. intestinihominis reduced the growth of transplantable cancers (RET melanoma, MCA205 sarcoma, MC38 colon cancers) in mice through a pathway requiring CD8+ T cells and IFNγ, but not IL-17. Finally, E. hirae and B. intestinihominis-specific memory Th1 cell immune responses selectively predicted longer progression-free survival in advanced lung and ovarian cancer patients treated with chemo-immunotherapy. At present, it is not known whether E. hirae can synergize with B. intestinihominis with respect to its tumor growth-blocking activity [60].

The immunogenicity of E. hirae. E. hirae resides in the jejunum and colon in about 25% individuals. It is very dominant in mouse guts at Gustave Roussy Villejuif. This Enterococcus is not pathogenic and benefits from cyclophosphamide-mediated increased gut permeability to translocate in secondary lymphoid organs, prime specific pathogenic CXCR3 + CCR3+ T cells that traffic to tumor lesions, and reprogram the tumor microenvironment, i.e., decreasing regulatory T cells and γδT17 cells and enhancing cancer-specific CTL responses. E. hirae also acts on the intestinal epithelial cells, modulating apoptosis and IL-18 secretion in a NOD2-dependent manner. E. hirae is recognized by bone marrow dendritic cells to induce IL-12 and IL-27 production in a NOD2-dependent fashion. All these properties participate to its adjuvant effects to boost cyclophosphamide-mediated immunostimulatory capacities and anticancer efficacy
Therefore, part of the immunomodulatory effects of the most well-known immunomodulator (CTX) require a functional microbiome, at least a Gram-positive small intestine residing Enterococcus and a colon residing Barnesiella.
6 Platinum Salts and Gut Microbiota
The demonstration of the importance of the gut microbiota on the efficacy of anticancer agents was also reported by the Dr. Giorgio Trinchieri’s group. Iida et al. demonstrated that bacteria accounted for the ROS-mediated antitumor effects of tumor-infiltrating myeloid cells during platinum-based anticancer therapies [61]. Indeed, the authors showed that the efficacy of platinum salts (oxaliplatin, cisplatin) relied on the production of reactive oxygen species (ROS) by intratumoral myeloid cells (myeloid suppressor cells, monocytic and immature dendritic cells) through a mechanism involving MyD88. ROS induce DNA damage culminating in DNA adduct formation and apoptosis of tumor cells and account for part of the tumoricidal activity of oxaliplatin against MC38 or EL4 tumor-bearing mice. Interestingly, the efficacy of chemotherapy was abrogated in germ-free or ATB-treated mice compared to SPF animals. Next, the authors investigated the impact of ATB treatment on oxaliplatin-mediated ROS production. The expression of Cybb encoding reactive oxygen species (ROS)-generating nicotinamide adenine dinucleotide phosphate oxidase (NOX2) was attenuated by the ATBs [61]. Furthermore, microbial compensation of ATB-treated mice with Lactobacillus acidophilus restored efficacy of cisplatin-based chemotherapy and reestablished inflammatory gene expression related to cisplatin functions [62]. Altogether, these results support the notion that the gut microbiota impacts on the redox equilibrium of the tumor microenvironment, affecting the therapeutic effects of platinum salts currently used in clinic.
7 Gut Bacteria and the Efficacy of Immuno-oncological Compounds
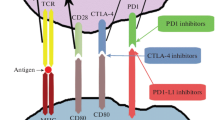
Oncologists no longer consider to treat cancer by only targeting tumor cell clones but also by mobilizing the immune system and eliciting long-term memory T cell responses protecting against the minimal residual disease. Immune checkpoint blockade (ICB) became the backbone of cancer treatment modalities, but primary resistance to ICB concerns about 70% of all comers, suggesting that T cell responses culminating in T cell infiltration of tumor lesions required for tumoricidal effects are quite a rare event. The immunity of a person is influenced by a complex set of factors, i.e., host, tumor, and environmental cues as well as prior histories of distinct therapies, that govern the threshold and timing of anticancer immune responses. Several lines of evidence point out to the critical role of the microbiome in dictating the “cancer immune set point” of a person, i.e., the threshold beyond which an immune response will ensure. Several groups indeed highlighted the role of distinct commensals in determining the antitumoral efficacy of various types of immunotherapeutics.
After administration of anti-interleukin-10 receptor (IL-10R) plus cytosine-phosphate-guanosine oligodeoxynucleotides (CpG-ODN) TLR9 agonists, Alistipes shahii was found overrepresented in the feces of colon cancer-bearing mice. Upon mono-association of previously sterilized mice with A. shahii, the immunotherapeutic response of subcutaneous colon cancers was improved, as compared to sterile mice. In this model, A. shahii led to an increase of TNFα production by intratumoral myeloid cells, and neutralization of TNFα abolished the therapeutic effect. Thus, A. shahii impact innate immune effectors of the myeloid lineage, reshaping the tumor microenvironment to improve the outcome of immunotherapy [61].
Other studies showed that the efficacy of immune checkpoint blockers (ICB), aimed at reactivating tumor-infiltrating T cells, is also dependent on the gut microbiota. Ipilimumab, a monoclonal antibody against cytotoxic-T-lymphocyte-associated protein 4 (CTL4-4), induced considerable improvement in the overall survival of patients with metastatic melanoma with up to 18% long-term control of the disease [63, 64]. The antitumoral efficacy of ipilimumab was abrogated in ATB-treated SPF mice or in germ-free animals bearing many types of transplantable tumors. Vétizou et al. showed that non-enterotoxin-producing strains of B. fragilis have anticancer properties in the context of immunotherapy with CTLA4 blockade. After neutralization of this immune checkpoint with specific monoclonal antibodies, Bacteroidales representation in the ileums and feces was affected, and B. fragilis spp. could take over, associated with DC migration and activation in the mesenteric lymph nodes and class II-restricted T cell memory responses against B. fragilis antigens. B. fragilis-mediated immune responses post-CTLA4 blockade were IL-12 dependent. The defect in preclinical response of GF tumor-bearing mice to anti-CTLA4 Ab was overcome by mono-association with B. fragilis as well as by adoptive transfer of CD4+ T cells that were previously primed with B. fragilis-pulsed dendritic cells (DCs) that contributed to the antitumor immune rejection. Interestingly, mice that were mono-associated with B. fragilis exhibited a more mature DC phenotype in tumor beds than controls with respect to the expression of MHC class II as well as the co-stimulatory CD80 and CD86 molecules. It has been documented that B. fragilis cell walls contain the immunostimulatory polysaccharide A (PSA) that can act on DCs. However, it remains to be determined whether PSA alone would be as efficient as live B. fragilis with respect to its antineoplastic activity. Finally, we were able to confirm that these findings were of clinical relevance by analyzing the gut microbiota of metastatic melanoma patients before and after ipilimumab. The 16S pyrosequencing analyses of feces contents in 25 stage IV melanoma patients revealed three major enterotypes based on the abundance or relative representativity of distinct spp. of Bacteroides and Prevotella genera. By performing fecal microbial transplantation (FMT) of feces representative of each enterotype into tumor-bearing GF mice subsequently treated with anti-CTLA4 Ab, we demonstrated that the microbial composition of enterotype C, enriched in immunogenic Bacteroides species, was able to allow the niching or colonization of B. fragilis (but not Bacteroides uniformis, vulgatus, or distasonis) and to restore the efficacy of anti-CTLA4 Ab, otherwise lost in GF mice, while clusters A and B failed to do both [65]. Of note, melanoma patients tended to exhibit an enterotype C while being treated with ipilimumab.
In parallel, Gajewski’s group showed that the antitumor efficacy of anti-PD-L1 Ab was influenced by the colon content in Bifidobacterium species (Bifidobacterium breve and Bifidobacterium longum). In this study, Sivan and colleagues compared relative antitumor CTL responses against a candidate tumor antigen in genetically similar C57BL/6 tumor bearers derived from two different mouse facilities (bought from two distinct vendors) differing in terms of microbiome composition. Contrasting mice from the Jackson Laboratory and from Taconic Farms, they revealed significant differences in the growth kinetics of subcutaneously implanted melanomas, with more aggressive tumors in Taconic Farms derived- mice attributable to lower dendritic cell maturation and IFN signatures associated with poor intratumoral cancer antigen-specific TIL accumulation. Interestingly, the tumor growth in Taconic Farms mice was reduced following FMT from mice originating from the Jackson Laboratory or cohoused with these littermates. Pyrosequencing of gene amplicons of Jackson and Taconic mice feces revealed a high content in Bifidobacteriales spp. in the colony that exhibited reduced growth of transplantable melanomas and improved CTL-mediated immunosurveillance [4]. Selective transfer of B. breve or B. longum into mice that are normally devoid of these strains was sufficient to reduce melanoma growth and restored anti-melanoma CTL responses. B. breve and B. longum stimulated the maturation of DC both in vitro and in vivo. As a consequence, the frequency of tumor-specific CTL accumulating in melanomas increased in mice carrying B. breve or B. longum, and such CTL-infiltrated tumors responded more vigorously to immunotherapy with an antibody targeting PD-L1 than did melanomas evolving on sterile mice or mice bearing a gut microbiome devoid of immunostimulatory Bifidobacteriales [66].
Hence, various strains of commensals were associated with the immunostimulatory effects of distinct I-O strategies. The future will tell us whether most of these commensals would mediate their bioactivity on innate or cognate immune responses regardless of the therapeutic compound or whether they would act within the mode of action and scope of the defined compound.
8 Role of Intestinal Microbes in Graft-Versus-Host Disease (GVHD)
Graft-versus-host disease occurs after allogeneic hematopoietic stem cell transplantation (allo-HSCT) and often limits the success of the therapy. GVHD results from the attack of the host cells by the transplanted donor immune cells. Numerous studies demonstrated that GVHD depends on several factors such as age, conditioning regimen, graft source, etc. Moreover, increasing evidences indicate that the gut microbiota plays a significant role in the pathogenesis of GVHD and could be associated to the relapse of hematologic malignancies after allo-HSCT. Previous studies performed in mice showed that the severity of GVHD is attenuated in GF mice or mice treated with antibiotics [67, 68] or in humans [69]. Characterization of the gut microbiota of GVHD patients revealed a significant decrease of diversity which was associated with an increase of Lactobacillales and decrease of Clostridiales, which in turn can modulate intestinal inflammation [70]. In a later study, Jenq et al. anticorrelated the abundance of Blautia, belonging to the Clostridia class, with GVDH-related mortality [71]. While bacterial species known for their health-promoting properties such as Faecalibacterium or Ruminococcus were deeply decreased, the abundance of Enterococci was markedly enhanced in GVHD patients, compared with patients without GVHD [72]. Indeed, an increase of Enterococci after transplantation in adult patients with severe GVHD was noticed. This was also observed in ten pediatric patients undergoing allo-HSCT. Interestingly, patients undergoing GVHD displayed lower amounts of Bacteroides and Parabacteroides, whose abundance positively correlated with the levels of SCFAs, especially propionate, before allo-HSCT [73]. Lately, Simms-Waldrip et al. demonstrated that the use of antibiotics targeting anaerobic bacteria in the course of allo-HSCT correlated with a significant decline of anti-inflammatory Clostridia and the development of GVHD [74]. As a result, manipulating the gut microbiota through the use of probiotics or prebiotics could be considered as a therapeutic approach aiming at decreasing the risk of GVHD. Preliminary data indicated that administration of Lactobacillus rhamnosus before and after allo-HSCT promoted less severe GVHD and improved survival of mice [75]. Other studies observed that administration of Lactobacillus johnsonii reduced GVHD severity through prevention of Enterococci accumulation post allo-HSCT [76]. Moreover, FMT was used in a clinical pilot study to treat GVHD of the gut occurring after allo-HSCT [77]. The abundance of Lactobacilli increased post-FMT in most of the patients, and the gut microbiota shifted toward an anti-inflammatory state. Other studies showed that FMT was efficient in treating Clostridium difficile infection post allo-HSCT [78, 79]. Butyrate-producing Clostridia strains reduced GVHD of the gut that is induced by allogeneic BMT. This effect was correlated with improved junctional integrity and reduced apoptosis of intestinal epithelial cells [80].
In a recent study, Peled et al. reported that intestinal abundance of Eubacterium limosum was associated with a reduced risk of relapse/progression of hematologic malignancies after allo-HSCT. Interestingly, the strongest association between E. limosum and a lower risk of disease relapse was observed in patients receiving T cell replete grafts. Donor cell-mediated graft-versus-tumor (GVT) effect was virtually higher in these patients, when compared to T cell-depleted graft recipients, suggesting that gut microbiota might play a role in GVT activity [81].
Altogether, these studies highlighted the dominant role of gut microbiota in dictating the efficacy and toxicity of allo-HSCT and open up new avenues of interventions on GVHD/GVT by harnessing the gut microbiome.
9 Bacteria for Therapy of Cancer: Probiotics and Others
The development of anticancer agents based on live microbial agents traditionally focused on local or systemic parenteral routes. William Coley was the first who partially succeeded in obtaining anticancer effect using microbes, by injecting intratumorally a mixture of Streptococcus pyogenes and Serratia marcescens [82]. Many efforts were then made to develop microbial-based anticancer treatment. Mycobacterium bovis Bacille Calmette-Guérin (BCG) was approved by FDA and EMA in 1990 for superficial bladder cancer treatment. BCG efficacy relies on the induction of a local immune response against residual cancer cells, reducing the probability of relapse [83, 84]. Since then, no other bacteria obtained marketing authorization despite Phase I clinical trials utilizing various strategies outlined in Table 27.1.
Investigators recently raised an alternative approach that consists in administering live bacteria (probiotics) per os to colonize the gut and consequently obtain anticancer activities.
Probiotics are defined as “live microorganisms which, when administered in adequate amounts, confer health benefits on the host” [85]. They are known to reinforce natural defenses, protect against gastrointestinal disorders and pathogens, and modulate innate and/or adaptive immunity.
Lactobacilli are substantial probiotics, which belong to the group of lactic acid bacteria. Many studies demonstrated that various isolates of Lactobacillus casei, Lactobacillus plantarum, Lactobacillus rhamnosus GG, and/or L. acidophilus mediate anticancer effects through different mechanisms, including activation of NK cells, maturation of DCs, or release of probiotic-derived ferrichrome [87,88,89,90,91,92,93,94,95,95].
Prohep, a probiotic mixture of Lactobacillus rhamnosus GG (LGG), Escherichia coli Nissle 1917, and heat-inactivated VSL#3, delivered orally to mice bearing subcutaneous hepatocellular carcinoma, prevented tumor progression, and shifted the gut microbial community toward Prevotella and Oscillibacter. These later are known to produce anti-inflammatory metabolites, which in turn decrease the Th17 polarization and enhance the differentiation of anti-inflammatory Treg/Tr1 cells in the gut [96].
Whereas these probiotics taken individually may elicit antitumor activities, it is not established if when optimally combined they can create an ecosystem with extensive antitumor properties.
As mentioned previously, some intestinal commensals modulate the antitumor immune responses generated by anticancer compounds as well as the efficacy of the treatment (CTX and E. hirae/B. intestinihominis [60], CpG-ODN + anti-IL10 and Alistipes shahii [61], anti-PDL1 and Bifidobacterium longum and breve [66], anti-CTLA4 and B. fragilis/Burkholderia cepacia [65]).
The mechanisms of action of the microorganisms described above are not fully deciphered at the molecular level. The identification of bacterial structures directly involved in anticancer activity or immunosurveillance promotion will raise favorable circumstances to (a) improve antitumor effect via probiotic engineering or (b) to develop compounds that mimic their pharmacological activity.
10 Genetically Modified Bacteria
Din et al. demonstrated that synthetic engineering of bacteria improved the antitumor effects of the antimetabolite 5-FU in a mice model of liver metastases of colorectal cancers. Pulsatile delivery cycles of 5-FU by the bacteria were allowed via a synchronized lysis cycle of E. coli based on quorum sensing feedback loops [110]. In necrotic tumor characterized by low vascularization, chemotherapy has limited efficacy due to restricted accessibility. This can be partially restored using anaerobic bacteria as tumor-targeting vectors. Bacteria engineering increased 5-FU efficacy in liver metastasis, but this strategy was fully successful when combined with I-O or other anticancer drugs [110]. This unique way to deliver compounds in avascular tumors needs to be further assessed in patients resistant to traditional therapies.
11 Microbial Products with Cancer-Modulating Properties
Microbial agents can synthetize a wide range of molecules that affect either antitumor immunosurveillance or growth/survival of cancer cells. One distinguishes (a) toxins, (b) ligands of pattern recognition receptors (PRRs), and (c) metabolites (short-chain fatty acids, polyamines, vitamins, secondary bile products, AhR ligands). While toxins display direct cytotoxic properties, activation of PRRs stimulates immune response, and metabolites affect the host metabolism. However, certain metabolites can also behave as PRR ligands, as it was shown for N-acetylglucosamine (a sugar subunit of bacterial peptidoglycan) acting on hexokinase to activate the NLRP3 inflammasome [111] or for tryptophan derivatives acting on the aryl hydrocarbon receptor (AhR) [112]. These approaches have been detailed in a previous report [113]. We will only recapitulate the most prominent ones that can be harnessed for oncological purposes.
Bacterial toxins are usually peptides with amphipathic alpha-helices containing cationic charges that cause the lysis of non-protected bacterial membranes. Structural analogs of toxins were developed in order to kill cancer cells. LTX-315, a synthetic peptide developed for intratumoral therapy, targets mitochondria. It also triggers necrotic cell death with immunogenic properties. This means its direct tumoricidal effects are prolonged as a consequence of danger signal emission that activates specific antineoplastic immune responses [114].
Several PRR ligands are FDA and EMA approved. Monophosphoryl lipid A (MPL), a derivative of Salmonella minnesota LPS, is used as an adjuvant in a peptide-based vaccine for the prevention of cervical carcinoma-associated strains of human papillomavirus [115]. Imiquimod, a synthetic agonist of TLR7, is topically delivered for the treatment of actinic keratosis.
Short-chain fatty acids (SCFAs) such as acetate, butyrate, and propionate are synthetized from dietary fibers and polysaccharides by the clostridial clusters IV and XIVa of Firmicutes. While acetate supports the expansion of various human cancer types [117,118,118], propionate and butyrate may have anticancer properties through distinct mechanisms. Propionate and butyrate could repair gut dysfunction, by promoting regulatory T cell (Treg) differentiation and/or accumulation and mediating anti-inflammatory activities [120,121,121] or inhibiting histone deacetylases (HDACs), conferring to these metabolites anticancer properties. Butyrate also induced apoptosis in colorectal cancer and lymphoma cells, suggesting direct tumoricidal activities [122, 123]. As a result, a way to prevent or treat cancer would consist in developing dietary regimen that increases intestinal butyrate over acetate production.
12 Future Prospects
Over the past decade, microbiome raised a major interest, since it appears that these commensal communities influence the development and outcome of a wide range of disease, including cancer. Microbiota might affect tumor formation and progression. It may also determine anticancer treatment responses. Either indirect effects such as immunosurveillance or direct effects of microbial compounds (i.e., carcinogens, cytotoxic agents, and metabolites) may affect tumor cells via diverse processes (mutagenesis, epigenetic modulation, stimulation of receptors on host cells, effect on anabolic and catabolic pathways).
Given that the cancer immune set point may be largely controlled by the health of our gut microbiome, diagnosis tools to evaluate a patient’s gut dysbiosis are urgently needed to be able to guide the indications of specific therapies and anticancer probiotics. Technologies based on culturomics, metagenomics, or PCR or mass spectrometry will lead to diagnosis tools for cancer-associated dysbiosis and predictors of primary resistance to ICB.
Increasing knowledge of the functional exploration of patient-derived microbiomes, coupled with relevant preclinical models, will allow the development of four alternative anticancer interventions: (a) orally administrable microorganisms (probiotics), (b) specific dietary or drug-based interventions that favor the expansion of beneficial microorganisms, (c) drugs that specifically target microbial enzymes that generate harmful toxins and metabolites, and (d) microbial products with anticancer properties. Each of these therapies could be used alone or in combination with conventional anticancer treatment. However, live microbes, especially when genetically modified, raise safety concerns with regard to their escape into the environment, their potential pathogenicity, and the acquisition of antibiotic or enzymatic chemoresistance. Hence, live microbial agents are subjected to regulatory and intellectual property-related unsolved issues. An alternative to eventually overcome such limitations would be the development of small molecules or chemically defined macromolecules that influence the natural gut microbiome in a favorable fashion or mediate beneficial effects on their own.
References
de Vos WM, de Vos EA. Role of the intestinal microbiome in health and disease: from correlation to causation. Nutr Rev. 2012;70:S45–56.
Karlsson FH, et al. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature. 2013;498:99–103.
Le Chatelier E, et al. Richness of human gut microbiome correlates with metabolic markers. Nature. 2013;500:541–6.
Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. 2014;14:667–85.
Arumugam M, et al. Enterotypes of the human gut microbiome. Nature. 2011;473:174–80.
Cotillard A, et al. Dietary intervention impact on gut microbial gene richness. Nature. 2013;500:585–8.
Segre JA. MICROBIOME. Microbial growth dynamics and human disease. Science. 2015;349:1058–9.
Hornef M. Pathogens, commensal symbionts, and pathobionts: discovery and functional effects on the host. ILAR J. 2015;56:159–62.
Ergin A, et al. Impaired peripheral Th1 CD4+ T cell response to Escherichia coli proteins in patients with Crohn’s disease and ankylosing spondylitis. J Clin Immunol. 2011;31:998–1009.
Macpherson A, Khoo UY, Forgacs I, Philpott-Howard J, Bjarnason I. Mucosal antibodies in inflammatory bowel disease are directed against intestinal bacteria. Gut. 1996;38:365–75.
Palm NW, et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell. 2014;158:1000–10.
Rhee K-J, Sethupathi P, Driks A, Lanning DK, Knight KL. Role of commensal bacteria in development of gut-associated lymphoid tissues and preimmune antibody repertoire. J Immunol. 2004;172:1118–24.
Ivanov II, et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139:485–98.
Faith JJ, Ahern PP, Ridaura VK, Cheng J, Gordon JI. Identifying gut microbe-host phenotype relationships using combinatorial communities in gnotobiotic mice. Sci Transl Med. 2014;6:220ra11.
Atarashi K, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331:337–41.
Atarashi K, et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature. 2013;500:232–6.
Qin J, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464:59–65.
Dejea CM, et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc Natl Acad Sci USA. 2014;111:18321–6.
Zackular JP, Rogers MAM, Ruffin MT, Schloss PD. The human gut microbiome as a screening tool for colorectal cancer. Cancer Prev Res Phila. 2014;7:1112–21.
Zeller G, et al. Potential of fecal microbiota for early-stage detection of colorectal cancer. Mol Syst Biol. 2014;10:766.
Zitvogel L, et al. Cancer and the gut microbiota: an unexpected link. Sci Transl Med. 2015;7:271ps1.
Abreu MT, Peek RM. Gastrointestinal malignancy and the microbiome. Gastroenterology. 2014;146:1534–1546.e3.
Sears CL, Garrett WS. Microbes, microbiota, and colon cancer. Cell Host Microbe. 2014;15:317–28.
Garrett WS. Cancer and the microbiota. Science. 2015;348:80–6.
Gur C, et al. Binding of the Fap 2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity. 2015;42:344–55.
Louis P, Hold GL, Flint HJ. The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol. 2014;12:661–72.
Bongers G, et al. Interplay of host microbiota, genetic perturbations, and inflammation promotes local development of intestinal neoplasms in mice. J Exp Med. 2014;211:457–72.
Rutkowski MR, et al. Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation. Cancer Cell. 2015;27:27–40.
Rossini A, et al. Influence of antibiotic treatment on breast carcinoma development in proto-neu transgenic mice. Cancer Res. 2006;66:6219–24.
Blaser M. Antibiotic overuse: stop the killing of beneficial bacteria. Nature. 2011;476:393–4.
Tikkanen MJ, Adlercreutz H, Pulkkinen MO. Effects of antibiotics on oestrogen metabolism. Br Med J. 1973;2:369.
Adlercreutz H, et al. Intestinal metabolism of estrogens. J Clin Endocrinol Metab. 1976;43:497–505.
Knekt P, et al. Does antibacterial treatment for urinary tract infection contribute to the risk of breast cancer? Br J Cancer. 2000;82:1107–10.
Kilkkinen A, et al. Antibiotic use predicts an increased risk of cancer. Int J Cancer. 2008;123:2152–5.
Zhang H, García Rodríguez LA, Hernández-Díaz S. Antibiotic use and the risk of lung cancer. Cancer Epidemiol Biomark Prev. 2008;17:1308–15.
Tamim HM, Hajeer AH, Boivin J-F, Collet J-P. Association between antibiotic use and risk of prostate cancer. Int J Cancer. 2010;127(4):952–60. doi:10.1002/ijc.25139.
Boursi B, Mamtani R, Haynes K, Yang Y-X. Recurrent antibiotic exposure may promote cancer formation – another step in understanding the role of the human microbiota? Eur J Cancer. 2015;51:2655–64.
Shiga K, et al. Presence of Streptococcus infection in extra-oropharyngeal head and neck squamous cell carcinoma and its implication in carcinogenesis. Oncol Rep. 2001;8:245–8.
Yu G, et al. Characterizing human lung tissue microbiota and its relationship to epidemiological and clinical features. Genome Biol. 2016;17:163.
Warren RL, et al. Co-occurrence of anaerobic bacteria in colorectal carcinomas. Microbiome. 2013;1:16.
Chase D, Goulder A, Zenhausern F, Monk B, Herbst-Kralovetz M. The vaginal and gastrointestinal microbiomes in gynecologic cancers: a review of applications in etiology, symptoms and treatment. Gynecol Oncol. 2015;138:190–200.
Crawford PA, Gordon JI. Microbial regulation of intestinal radiosensitivity. Proc Natl Acad Sci U S A. 2005;102:13254–9.
Cui M, et al. Faecal microbiota transplantation protects against radiation-induced toxicity. EMBO Mol Med. 2017;9(4):448–61. doi:10.15252/emmm.201606932.
Nam Y-D, Kim HJ, Seo J-G, Kang SW, Bae J-W. Impact of pelvic radiotherapy on gut microbiota of gynecological cancer patients revealed by massive pyrosequencing. PLoS One. 2013;8:e82659.
Ciorba MA, et al. Lactobacillus probiotic protects intestinal epithelium from radiation injury in a TLR-2/cyclo-oxygenase-2-dependent manner. Gut. 2012;61:829–38.
Salminen E, Elomaa I, Minkkinen J, Vapaatalo H, Salminen S. Preservation of intestinal integrity during radiotherapy using live Lactobacillus acidophilus cultures. Clin Radiol. 1988;39:435–7.
Delia P, et al. Use of probiotics for prevention of radiation-induced diarrhea. World J Gastroenterol. 2007;13:912–5.
Chitapanarux I, et al. Randomized controlled trial of live lactobacillus acidophilus plus bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat Oncol. 2010;5:31.
Wang A, et al. Gut microbial dysbiosis may predict diarrhea and fatigue in patients undergoing pelvic cancer radiotherapy: a pilot study. PLoS One. 2015;10:e0126312.
Ciorba MA, Hallemeier CL, Stenson WF, Parikh PJ. Probiotics to prevent gastrointestinal toxicity from cancer therapy: an interpretive review and call to action. Curr Opin Support Palliat Care. 2015;9:157–62.
Nejdfors P, Ekelund M, Weström BR, Willén R, Jeppsson B. Intestinal permeability in humans is increased after radiation therapy. Dis Colon Rectum. 2000;43:1582–1587.; discussion 1587–1588.
Paulos CM, et al. Microbial translocation augments the function of adoptively transferred self/tumor-specific CD8+ T cells via TLR4 signaling. J Clin Invest. 2007;117:2197–204.
Espinosa-Carrasco G, et al. Systemic LPS translocation activates cross-presenting dendritic cells but is dispensable for the breakdown of CD8+ T cell peripheral tolerance in irradiated mice. PLoS One. 2015;10:e0130041.
Ubeda C, et al. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J Clin Invest. 2010;120:4332–41.
van Vliet MJ, Harmsen HJM, de Bont ESJM, Tissing WJE. The role of intestinal microbiota in the development and severity of chemotherapy-induced mucositis. PLoS Pathog. 2010;6:e1000879.
Stringer AM, et al. Irinotecan-induced mucositis manifesting as diarrhoea corresponds with an amended intestinal flora and mucin profile. Int J Exp Pathol. 2009;90:489–99.
Yang J, Liu K, Qu J, Wang X. The changes induced by cyclophosphamide in intestinal barrier and microflora in mice. Eur J Pharmacol. 2013;714:120–4.
Sistigu A, et al. Immunomodulatory effects of cyclophosphamide and implementations for vaccine design. Semin Immunopathol. 2011;33:369–83.
Viaud S, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science. 2013;342:971–6.
Daillère R, et al. Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity. 2016;45:931–43.
Iida N, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342:967–70.
Gui Q-F, Lu H-F, Zhang C-X, Xu Z-R, Yang Y-H. Well-balanced commensal microbiota contributes to anti-cancer response in a lung cancer mouse model. Genet Mol Res. 2015;14:5642–51.
Hodi FS, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010;363:711–23.
Robert C, Mateus C. Anti-CTLA-4 monoclonal antibody: a major step in the treatment of metastatic melanoma. Med Sci. 2011;27:850–8.
Vétizou M, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350:1079–84.
Sivan A, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015;350:1084–9.
Jones JM, Wilson R, Bealmear PM. Mortality and gross pathology of secondary disease in germfree mouse radiation chimeras. Radiat Res. 1971;45:577.
van Bekkum DW, Roodenburg J, Heidt PJ, van der Waaij D. Mitigation of secondary disease of allogeneic mouse radiation chimeras by modification of the intestinal microflora. J Natl Cancer Inst. 1974;52:401–4.
Vossen JM, et al. Complete suppression of the gut microbiome prevents acute graft-versus-host disease following allogeneic bone marrow transplantation. PLoS One. 2014;9:e105706.
Taur Y, et al. The effects of intestinal tract bacterial diversity on mortality following allogeneic hematopoietic stem cell transplantation. Blood. 2014;124:1174–82.
Jenq RR, et al. Intestinal Blautia is associated with reduced death from graft-versus-host disease. Biol Blood Marrow Transplant. 2015;21:1373–83.
Holler E, et al. Metagenomic analysis of the stool microbiome in patients receiving allogeneic stem cell transplantation: loss of diversity is associated with use of systemic antibiotics and more pronounced in gastrointestinal graft-versus-host disease. Biol Blood Marrow Transplant. 2014;20:640–5.
Biagi E, et al. Gut microbiota trajectory in pediatric patients undergoing hematopoietic SCT. Bone Marrow Transplant. 2015;50:992–8.
Simms-Waldrip TR, et al. Antibiotic-induced depletion of anti-inflammatory Clostridia is associated with the development of graft-versus-host disease in pediatric stem cell transplantation patients. Biol Blood Marrow Transplant. 2017;23(5):820–9. doi:10.1016/j.bbmt.2017.02.004.
Gerbitz A, et al. Probiotic effects on experimental graft-versus-host disease: let them eat yogurt. Blood. 2004;103:4365–7.
Jenq RR, et al. Regulation of intestinal inflammation by microbiota following allogeneic bone marrow transplantation. J Exp Med. 2012;209:903–11.
Kakihana K, et al. Fecal microbiota transplantation for patients with steroid-resistant acute graft-versus-host disease of the gut. Blood. 2016;128:2083–8.
Neemann K, et al. Fecal microbiota transplantation for fulminant Clostridium difficile infection in an allogeneic stem cell transplant patient. Transpl Infect Dis. 2012;14:E161–5.
de Castro CG, Ganc AJ, Ganc RL, Petrolli MS, Hamerschlack N. Fecal microbiota transplant after hematopoietic SCT: report of a successful case. Bone Marrow Transplant. 2015;50:145.
Mathewson ND, et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat Immunol. 2016;17:505–13.
Peled JU, et al. Intestinal microbiota and relapse after hematopoietic-cell transplantation. J Clin Oncol Off J Am Soc Clin Oncol. 2017;35(15):1650–9. doi:10.1200/JCO.2016.70.3348.
Nauts HC, Swift WE, Coley BL. The treatment of malignant tumors by bacterial toxins as developed by the late William B. Coley, M.D., reviewed in the light of modern research. Cancer Res. 1946;6:205–16.
Böhle A, Brandau S. Immune mechanisms in bacillus Calmette-Guerin immunotherapy for superficial bladder cancer. J Urol. 2003;170:964–9.
Zbar B, Bernstein I, Tanaka T, Rapp HJ. Tumor immunity produced by the intradermal inoculation of living tumor cells and living Mycobacterium bovis (strain BCG). Science. 1970;170:1217–8.
Morelli L, Capurso L. FAO/WHO guidelines on probiotics: 10 years later. J Clin Gastroenterol. 2012;46(Suppl):S1–2.
Aragón F, Carino S, Perdigón G, de Moreno de LeBlanc A. Inhibition of growth and metastasis of breast cancer in mice by milk fermented with lactobacillus casei CRL 431. J Immunother. 2015;1997(38):185–96.
Hu J, et al. Anti-tumour immune effect of oral administration of Lactobacillus plantarum to CT26 tumour-bearing mice. J Biosci. 2015;40:269–79.
Kato I, Endo K, Yokokura T. Effects of oral administration of Lactobacillus casei on antitumor responses induced by tumor resection in mice. Int J Immunopharmacol. 1994;16:29–36.
Cai S, et al. Lactobacillus rhamnosus GG activation of dendritic cells and neutrophils depends on the dose and time of exposure. J Immunol Res. 2016;2016:7402760.
Khazaie K, et al. Abating colon cancer polyposis by Lactobacillus acidophilus deficient in lipoteichoic acid. Proc Natl Acad Sci U S A. 2012;109:10462–7.
Lenoir M, et al. Lactobacillus casei BL23 regulates Treg and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J Gastroenterol. 2016;51:862–73.
Lee J-W, et al. Immunomodulatory and antitumor effects in vivo by the cytoplasmic fraction of Lactobacillus casei and Bifidobacterium longum. J Vet Sci. 2004;5:41–8.
Baldwin C, et al. Probiotic Lactobacillus acidophilus and L. casei mix sensitize colorectal tumoral cells to 5-fluorouracil-induced apoptosis. Nutr Cancer. 2010;62:371–8.
Konishi H, et al. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat Commun. 2016;7:12365.
Takagi A, et al. Relationship between the in vitro response of dendritic cells to Lactobacillus and prevention of tumorigenesis in the mouse. J Gastroenterol. 2008;43:661–9.
Li J, et al. Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. Proc Natl Acad Sci USA. 2016;113:E1306–15.
Martínez-Piñeiro JA, et al. Bacillus Calmette-Guerin versus doxorubicin versus thiotepa: a randomized prospective study in 202 patients with superficial bladder cancer. J Urol. 1990;143:502–6.
Aso Y, Akazan H. Prophylactic effect of a Lactobacillus casei preparation on the recurrence of superficial bladder cancer BLP study group. Urol Int. 1992;49:125–9.
Ohashi Y, et al. Habitual intake of lactic acid bacteria and risk reduction of bladder cancer. Urol Int. 2002;68:273–80.
Hoesl CE, Altwein JE. The probiotic approach: an alternative treatment option in urology. Eur Urol. 2005;47:288–96.
Stebbing J, et al. An intra-patient placebo-controlled phase I trial to evaluate the safety and tolerability of intradermal IMM-101 in melanoma. Ann Oncol. 2012;23:1314–9.
Dalgleish AG, et al. Randomised, open-label, phase II study of gemcitabine with and without IMM-101 for advanced pancreatic cancer. Br J Cancer. 2016;115:789–96.
Le DT, et al. Safety and survival with GVAX pancreas prime and Listeria monocytogenes-expressing mesothelin (CRS-207) boost vaccines for metastatic pancreatic cancer. J Clin Oncol Off J Am Soc Clin Oncol. 2015;33:1325–33.
Liu-Chittenden Y, et al. Phase I trial of systemic intravenous infusion of interleukin-13-Pseudomonas exotoxin in patients with metastatic adrenocortical carcinoma. Cancer Med. 2015;4:1060–8.
Weber F, et al. Safety, tolerability, and tumor response of IL4-Pseudomonas exotoxin (NBI-3001) in patients with recurrent malignant glioma. J Neuro-Oncol. 2003;64:125–37.
Toso JF, et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J Clin Oncol Off J Am Soc Clin Oncol. 2002;20:142–52.
Heimann DM, Rosenberg SA. Continuous intravenous administration of live genetically modified salmonella typhimurium in patients with metastatic melanoma. J. Immunother. 2003;1997(26):179–80.
Nemunaitis J, et al. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther. 2003;10:737–44.
Laske DW, Youle RJ, Oldfield EH. Tumor regression with regional distribution of the targeted toxin TF-CRM107 in patients with malignant brain tumors. Nat Med. 1997;3:1362–8.
Din MO, et al. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016;536:81–5.
Wolf AJ, et al. Hexokinase is an innate immune receptor for the detection of bacterial peptidoglycan. Cell. 2016;166:624–36.
Moura-Alves P, et al. AhR sensing of bacterial pigments regulates antibacterial defence. Nature. 2014;512:387–92.
Zitvogel L, et al. Anticancer effects of the microbiome and its products. Nat Rev Microbiol. 2017;15:465–78.
Ellerby HM, et al. Anti-cancer activity of targeted pro-apoptotic peptides. Nat Med. 1999;5:1032–8.
Paavonen J, et al. Efficacy of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine against cervical infection and precancer caused by oncogenic HPV types (PATRICIA): final analysis of a double-blind, randomised study in young women. Lancet. 2009;374:301–14.
Comerford SA, et al. Acetate dependence of tumors. Cell. 2014;159:1591–602.
Mashimo T, et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 2014;159:1603–14.
Schug ZT, et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 2015;27:57–71.
Arpaia N, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 2013;504:451–5.
Furusawa Y, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 2013;504:446–50.
Smith PM, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. 2013;341:569–73.
Jan G, et al. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002;9:179–88.
Wei W, Sun W, Yu S, Yang Y, Ai L. Butyrate production from high-fiber diet protects against lymphoma tumor. Leuk Lymphoma. 2016;57:2401–8.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Bonvalet, M., Daillère, R., Roberti, M.P., Rauber, C., Zitvogel, L. (2018). The Impact of the Intestinal Microbiota in Therapeutic Responses Against Cancer. In: Zitvogel, L., Kroemer, G. (eds) Oncoimmunology. Springer, Cham. https://doi.org/10.1007/978-3-319-62431-0_27
Download citation
DOI: https://doi.org/10.1007/978-3-319-62431-0_27
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-62430-3
Online ISBN: 978-3-319-62431-0
eBook Packages: MedicineMedicine (R0)