
Abstract
Long-tailed macaques commonly live near human settlements in Southeast Asia and Singapore is one example of such an interface. In 2011 and 2012, we conducted a census for Singapore’s National Parks Board (NParks), during which we collected behavioral, demographic, and ranging data. We used these data to examine how the presence of humans and access to human food related to changes in the macaques’ time budget, ranging behavior, and group size. We found that human presence was associated with decreased traveling rates, decreased arboreality, increased terrestriality, and increased use of human-made substrates. In particular, access to human food was associated with larger macaque group sizes, decreased arboreality, and increased use of human-made structures. Our results demonstrate how living near humans in an intensely urban habitat impacts macaques. Perhaps with better knowledge of how humans affect urban macaques, we can better plan management strategies to mitigate conflict. We discuss some nonlethal strategies for managing Singapore’s human–macaque interface that could potentially reduce human–macaque conflict. Specifically, we recommend consistent enforcement of an existing feeding ban, the employment of security guards to mitigate conflict in particularly problematic areas, and the expansion of existing education programs for local people and tourists.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Ethnoprimatology
- Human–macaque interface
- Human–wildlife conflict
- Long-tailed macaques
- Macaca fascicularis
- Macaque management
- Provisioning
- Singapore
- Southeast Asia
- Urban wildlife
Introduction
There are currently 23 recognized species in the genus Macaca , the most widely distributed primate genus following Homo (Li et al. 2015; Thierry 2007). Their wide geographic range is a testament to their adaptability to different environments, including human-altered ones. While not all macaques are adapted to anthropogenic landscapes (i.e., lion-tailed macaques, see Singh et al. 2002), species such as long-tailed macaques ( M. fascicularis ), rhesus macaques (M. mulatta), toque macaques (M. sinica), and bonnet macaques (M. radiata) live in urban areas (Richard et al. 1989) and are often referred to as "weed macaques." As human populations and economic development continue to increase, more natural habitats will be converted into anthropogenic landscapes, causing more macaques to face the challenges associated with urban settings. In this chapter, we briefly summarize the human–macaque urban interface and focus on the time budgets, ranging, and group size of long-tailed macaques living in Singapore. We then discuss our perspective on nonlethal management strategies for urban macaques.
The Urbanization of Macaques
Most macaque species reside in South Asia, which is growing and developing rapidly. For instance, the percentage of the human population living in urban areas in Cambodia is expected to increase from 17 % in the year 2000 to 44 % in 2050 (Sheng 2012). Similarly, Singapore is projected to grow from 5.5 to 6.9 million people by 2030, a 25 % increase (National Population and Talent Division 2013). As this growth occurs, it is reasonable to think that the frequency of human and macaque encounters will increase. To better prepare for this increase, it is important to grasp the manner in which humans and macaques regard each other in urban settings. Farmland areas (Peterson and Riley 2013; Riley and Priston 2010), temple complexes (Fuentes 2013; Mallapur 2013; Wheatley 1999), and cities (Sha et al. 2009) are only a few of the areas where the human–macaque interface has been studied. The results of these studies suggest that humans impact macaques in several ways. For example, humans can transmit infectious agents to macaques (Epstein and Price 2009; Muehlenbein et al. 2010). Additionally, people can physically harm macaques. Intentional killings may include culling as a management strategy, pest control near crops, or hunting for food (Nahallage et al. 2008; Nahallage and Huffman 2013). Humans can also unintentionally affect macaque population levels by hitting them with cars or allowing pet dogs to kill them (Gumert et al. 2013; Riley et al. 2015a).
Studies have also found that the anthropogenic landscape alters several macaque behaviors including those associated with substrate use (Aggimarangsee 1992; Seth et al. 2001), ranging patterns (Berman et al. 2014; Klegarth in press), activity budgets (Chauhan and Pirta 2010; Chopra et al. 1992; Jaman and Huffman 2013), social structures (Chapman and Rothman 2009; Kamilar and Baden 2014; Lane-deGraaf et al. 2014; Sinha 2005; Sinha et al. 2005), aggression levels (Southwick 1972), and diets (Gumert et al. 2013; Klegarth in press; Singh et al. 2001; Sinha and Mukhopadhyay 2013). Additionally, obesity has been found to threaten the health of urban macaques (Aggimarangsee 1992; Knight 2011; Lane et al. 2010).
Many of the changes listed above are due to differences in food availability near humans when compared to “wild” habitats, as urban areas tend to be food rich. For example, people provision macaques for recreational pleasure, bait them out for viewing, or indirectly provide them with discarded refuse. Direct provisioning is especially common from tourists at monkey parks (Berman et al. 2014; Fa 1992; Knight 2011) and temples (Aggimarangsee 1992; Wheatley and Putra 1994a). Provisioning is generally encouraged at these sites so that food vendors can gain profits (Gumert 2011; Wheatley and Putra 1994b). Direct provisioning can also occur in other urban areas, but is generally less organized than at tourist sites. Provisioning in this manner may decrease reliance on natural food sources. In Shimla, India for example, rhesus macaques derived only 9 % of their diet from natural sources, with scavenging and human feeding accounting for 77 % of their diet, snatching/stealing for 9 %, and crop raiding for 5 % (Chauhan and Pirta 2010). Humans can also indirectly affect food availability and diets with food that macaques scavenge from garbage bins, take from homes, or snatch from passersby. Such food snatching is especially common where macaques are highly habituated and do not fear people.
When macaques snatch food, people sometimes respond with aggression, such as chasing or attempting to strike the macaques, or with fear, such as fleeing or screaming. Thus, food snatching is typically a negative experience for people. For example, at Shou-Shan Nature Park in Taiwan, Formosan macaques (M. cyclopis) displayed aggression towards tourists nearly five times more frequently during food provisioning than during times of non-provisioning (Hsu et al. 2009). In Dehradun, India, humans engaged in aggression towards macaques six times more frequently than the macaques were aggressive towards humans, and aggression was often triggered in response to macaques attempting to steal food. Furthermore, macaque aggression was usually counter to the human-initiated aggression (Beisner et al. 2014).
Macaque Impacts on Humans
Although humans have many impacts on macaques, the human–macaque interface is bidirectional and macaques have harmful and beneficial impacts on people too. For example, macaques can transmit infectious agents to humans (Jones-Engel et al. 2005, 2006), damage people’s property, raid crops, and cause other economic losses (Riley and Priston 2010). Macaques can also physically harm humans, typically during conflicts over food access (Fa 1992; Fuentes 2006; Fuentes and Gamerl 2005; Imam and Yahya 2001). Very rarely, macaques have killed people (BBC News 2010; Southwick and Siddiqi 2011; Zhao and Deng 1992).
Macaques can also benefit humans . For example, macaques provide substantial economic gains in the form of revenues from tourism (Fuentes 2006, 2010; Gumert 2011; Knight 2011) and are sacred symbols for millions of people (Aggimarangsee 1992; Fuentes 2006; Fuentes et al. 2005; Wheatley 1999). Macaques may also benefit the local environment as seed dispersers, helping to maintain the health of ecosystems (Dudgeon and Corlett 1994). Additionally, some people simply enjoy watching macaques (Malaivijitnond and Hamada 2008). In Singapore specifically, macaques add to urban biodiversity because they are among the only medium sized mammals that live in the urban environment (Lucas and Corlett 1998).
The Human–Macaque Interface in Singapore
Singapore is a densely populated city-state located at the southern tip of peninsular Malaysia. The 718-km2 island is home to 5.5 million people (Singapore Department of Statistics 2015) and approximately 1900 macaques, according to a census in 2012 (Riley et al. 2015b). The majority of the macaque population is concentrated in the protected areas of the Central Catchment Nature Reserves and adjacent Bukit Timah Nature Reserve, which constitutes 4 % of Singapore’s land area (National Parks Board of Singapore 2009, Riley et al. 2015b). The human–macaque interface in Singapore is relatively mild compared to other countries with human–macaque conflict problems (Fuentes et al. 2008; Sha et al. 2009). For example, direct physical contact between humans and macaques is less frequent than elsewhere in Southeast Asia (Fuentes et al. 2008), in part because Singapore does not have monkey temples and has strict regulations about macaque feeding. The mild nature of conflict can be partially attributed to proactive strategies of Singaporean government agencies. For example, they periodically conduct public service campaigns to educate people regarding how to live and act near macaques and they actively manage portions of the macaque population through culling when they reach high levels of overlap and conflict (Lee and Chan 2011). Despite these programs, Singapore still faces considerable challenges in managing their macaque population and limiting human–macaque conflict. Macaques take food, raid garbage bins, damage property, and injure people, albeit rarely (Feng 2011; Sha et al. 2009). Humans also continue to feed macaques in spite of the feeding ban, threaten or chase macaques, and unintentionally kill macaques with their cars.
Attitudes toward macaques in Singapore vary. Visitors to recreation parks tended to view macaques more positively than residents who live near recreation parks, as 37 % of visitors reported a positive attitude toward macaques compared to 21 % of residents (Sha et al. 2009). Resident attitudes are also reflected in frequent media coverage of macaques. Sha and colleagues (2009) found 47 macaque-related headlines in Singaporean newspapers between 2004 and 2008, showing that the issue is frequently discussed in the public. Wildlife authorities have hotlines for receiving complaints about macaques, and complaints about macaques are frequently received (Ee 2013; Feng 2013) with high levels of complaints associated with culling. In 2013, 570 macaques were culled (Khew 2014) after receiving 1860 complaints (Feng 2015). This is approximately 30 % of the estimated macaque population in Singapore (Riley et al. 2015b). Several years prior to the culling, a small survey found high levels of support for conserving macaques and only a small number of people supported culling efforts (Sha et al. 2009).
For this chapter, we analyzed data collected during a nationwide census of the long-tailed population of macaques in Singapore. In particular, we focused on how overlap with humans affects group size, substrate use, and behavioral time budgets . We used two variables as indicators of human overlap: presence of humans during observation and whether or not a group had ever been observed accessing human food. Based on prior research, we expected groups with greater exposure to humans and human food to be larger. We also expected macaques with such exposure to be more terrestrial, less arboreal, and have altered time budgets.
Methods
A nationwide census of Singapore’s macaques was conducted between October 2011 and November 2012 (Riley et al. 2015b). We studied 67 groups and spent an average of 7 h and 45 min with each group of macaques. During our study, positional and behavioral data were collected. Upon encountering a macaque group, macaques were counted every 30 min until we were confident in the accuracy of our count. For additional details on how groups were determined, see Riley and colleagues (2015b). While with macaques, our GPS device was set to automatically generate track points at 30-s intervals (Fig. 1). We conducted scan samples (Altmann 1974) of each visible individual’s behavior at 30-min intervals to obtain data on general time budgets. Each subject was scored into one of six behavioral categories : resting, socializing, traveling, natural feeding, human feeding, and human–macaque interaction (other than feeding). For feeding scans, we scored the type of food being consumed into one of 17 categories (e.g., fruits, insects). For social scans, we scored the type of social behavior as aggression, grooming, mounting/being mounted, or playing. We also scored the type of substrate (i.e., arboreal, terrestrial, or human-made structure) and distance to the nearest human other than the researcher(s) (no humans visible, less than 1 m, 1–3 m, 3–5 m, 5–10 m, or greater than 10 m). Although the presence of the researcher(s) likely did have some effect on the macaques, we attempted to mitigate those effects by maintaining a reasonable distance and avoiding direct eye contact, loud noises, and sudden movement.

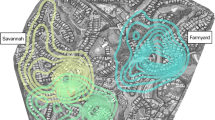
This map shows the track points (n = 64,666) denoting the ranging patterns of Singapore’s macaque groups (n = 69). Track points were collected at 30-s intervals during macaque follows
Statistical Analyses
We coded macaques regarding whether they were in a group observed accessing human food at least one time or in a group never observed accessing human food. We examined differences in activity budgets and substrate use depending on human food access and presence or absence of humans other than the researcher(s). For count data, omnibus analyses were done using Pearson goodness-of-fit tests if sufficient observations were in each cell to meet assumptions (i.e., at least 80 % of cells have expected frequencies less than 5). If this assumption was not met, we instead report the likelihood ratio test. For post hoc analyses, the variable level of interest (e.g., arboreal) was separated from the others (e.g., structures and terrestrial) by coding the level of interest as a one and the other levels as zeroes. This process was repeated so that each level of the variable was designated as the level of interest and then compared with the other levels of the variable. Post hoc analyses used Pearson goodness-of-fit tests (or likelihood ratio tests if too many expected values were less than 5) unless analyses were of 2 × 2 tables, in which case we used Fisher’s exact test. Alpha was set to p < 0.05 for all tests other than post hoc tests, which used Bonferroni-corrected alpha values to control for multiple comparisons. Throughout, any other statistical tests are explicitly mentioned. Statistical analyses were conducted in IBM SPSS Statistics version 22.
Spatial Analysis
In addition to behavioral data, we used ranging data to examine the extent of human–macaque overlap in Singapore. Spatial joins were conducted on each track point to determine the percentage of time macaques spent ranging within the boundaries of designated nature reserves and near anthropogenic areas such as residences or businesses. Five-meter buffers were placed around all roads to determine the amount of time macaques ranged on or within five meters of a road. Spatial analyses were conducted in ArcGIS version 10.3.1.
Results
Group Size
We studied 67 groups and found an average of 24.2 individuals per group (SD = 9.85, range 2–48). We also observed two lone males. Groups observed accessing human food (M = 28.27, SD = 11.50, 95 % CI: 23.17–33.37) were larger than those not observed accessing human food (M = 21.36, SD = 9.28, 95 % CI: 20.29–25.12), t (31.35) = 2.04, p = .050, d = 0.57 (using Welch two sample t-test due to unequal variances; Bartlett’s K-squared = 4.03, df = 1, p = 0.04). For additional details on population characteristics (age-sex breakdown, population density by region, spatial distribution of groups), see Riley and colleagues (2015b).
Distance to Humans
Macaques spent 0.4 % of their time less than 1 m from a human, 1.7 % between 1 and 3 m, 4.4 % between 3 and 5 m, 4.8 % between 5 and 10 m, 31.5 % greater than 10 m from a human, and 57.2 % with no humans visible. Considering only whether humans were present or absent, 57.2 % of observations (n = 6392) occurred without a human present and 42.8 % of observations occurred in the presence of other humans. Given the low counts at some distances, and the fact that much of the variability in distance to humans can be accounted for by recoding distance as humans present or absent, subsequent analyses of activity budgets and substrate usage evaluated presence-versus-absence of humans rather than the distance to the nearest human.
Activity Budgets
Activity budgets differed for macaques in groups that had been observed accessing human food compared to those from groups never observed accessing food, χ 2 (3, N = 6301) = 9.36, p = .025 (Fig. 2). Observations of human–macaque interactions were excluded because there were too few observations for chi-square analysis. Post hoc analyses confirmed that the effect for food access held for traveling (p = .001), but not eating natural food (p = .713), resting (p = .034), or socializing (p = .389). Macaques from groups observed accessing human food traveled less.

Activity budgets of macaques with and without access to human food. An asterisk indicates a significant difference
Whether people were present or not (Fig. 3) was also related to activity budgets, χ 2 (4, N = 6379) = 90.79, p < .001. Post hoc analyses indicated this held for eating human food (p < .001), and traveling (p < .001), but not eating natural foods (p = .496), resting (p = .488), or socializing (p = .947). Thus, when humans were present the macaques were more likely to be eating human food and less likely to be traveling , but equally likely to be eating natural food, resting, or socializing.

Activity budget of macaques when humans were present versus absent. An asterisk indicates a significant difference
Substrate Use
Whether an individual was in a group that had been observed accessing human food was related to the likelihood of using the three substrates χ 2 (2, N = 6434) = 136.42, p < .001. Post hoc analyses indicated differences for time spent arboreally (p < .001) and on structures (p < .001), but not terrestrially (p = .790). The pattern of results (Fig. 4) indicates that individuals in groups that had been observed accessing human food were more likely to be on structures and less likely to be arboreal, as compared to individuals from groups never observed accessing human foods.

Substrate use by macaques with and without access to human food. An asterisk indicates a significant difference
Human presence also had an effect on substrate use, χ 2 (2, N = 6392) = 77.49, p < .001. Post hoc analyses indicated that human presence was important for arboreal, structures, and terrestrial usage (ps < .001). When humans were present, the macaques were less likely to be in the trees and more likely to be terrestrial or on structures (Fig. 5).

Substrate use by macaques when humans were present versus absent. An asterisk indicates a significant difference
Spatial Distribution
Our analysis showed that macaques spent much of their time within the boundaries of nature reserves. Of 64,666 track points that marked the position of the researcher every 30 s when following macaques, 43.2 % (n = 27,910) occurred within the boundaries of a nature reserve. Of the 69 social units (67 groups and two lone males) observed during the census, over half (n = 35) ranged in an anthropogenic area (near a road, residence, or business) at some point during our observations. Another 15.7 % (n = 11) ranged within 100 m of an anthropogenic area. Of the track points that marked macaque ranging, 17.5 % (n = 11,337) occurred on or within five meters of a road. A large portion of roadside track points, 14.6 % of observations (n = 1658), were accounted for by one road (Old Upper Thompson Road) which passed through home ranges of four groups of macaques.
Discussion
Our study found that for macaques in Singapore, human presence is associated with decreased traveling rates, decreased arboreality, increased terrestriality, and increased use of human-made substrates. A related variable, access to human food, correlates with larger macaque group sizes, decreased arboreality, and increased use of human-made structures. We also found that Singapore’s macaques spent most of their time within the boundaries of the nature reserves.
These results highlight the differences related to exposure to humans and human food . Given that Singapore is highly urbanized and all macaque groups can reasonably be considered urban, substantial differences nonetheless exist among individuals and groups in the degree to which they overlap with humans and human food. Individuals in groups that accessed human food spent more time on human-made structures, consistent with results from Aggimarangsee (1992), less time in trees, consistent with Fooden (2000), and less time traveling. Surprisingly, individuals in groups that accessed human food did not spend significantly less time foraging for natural foods, which has been observed in other populations (Chauhan and Pirta 2010). Overall, macaque substrate use differed depending upon whether the group was observed accessing human food and whether or not a person was present. A clear pattern emerges across these analyses: human influence seems to bring the macaques out of the trees and onto the land and structures.
In the highly urbanized habitat of Singapore, it is perhaps surprising that macaques managed to avoid areas heavily populated by humans to some degree. Overall, 34 of 69 social units did not range in an anthropogenic habitat during our observations, and macaques spent 43 % of their time within the boundaries of nature reserves, even though protected parkland accounts for less than 5 % of Singapore’s land area. Furthermore, 43 groups never accessed human food during our observations. However, simply because a group was not observed accessing human food or ranging in an anthropogenic area does not mean that those groups always avoid human food and human-inhabited areas. The finding that macaques spent 43 % of their time in nature reserves could be misinterpreted to mean that macaques spend a large proportion of their time away from humans. However, nature reserves in Singapore are heavily used for recreational purposes, so they are common areas of interface for macaques and humans, although they are typically less intense interfaces than anthropogenic areas such as residential complexes or roads.
Effects of Food Access
The macaque groups in our study differed on variables typically associated with urban versus nonurban living (including substrate use, behavioral time budgets, and group size), even though all the groups in our study could reasonably be considered urban. The reasons for such differences are not easy to determine. Jaman and Huffman (2013) found that human food access was responsible for the differences in activity budgets between urban and nonurban groups in their study. It seems likely that access to human food underlies activity budget differences in our study as well. Human food access may also result in larger groups, or larger groups may simply outcompete smaller groups for access to human food and to areas with human food access (Lane-deGraaf et al. 2014). If this is indeed the case, then human food access may be a feedback loop in which large groups outcompete smaller groups for access to human food; access to human food increases reproductive rates and groups get larger; and larger groups outcompete other groups for access to human food.
Management of Urban Macaques
In order to effectively manage human–macaque interfaces, officials need to address three key dimensions: the macaque dimension, the human dimension, and the environmental dimension (Jones-Engel et al. 2011). The macaque dimension focuses on understanding and changing the composition, distribution, and/or behavior of the macaque population. The human dimension involves understanding the changing composition, distribution, and/or behavior of the human population. Lastly, the environmental dimension deals with understanding, developing, and modifying the local conditions where humans and macaques interface, so as to reduce overlap and contact between humans and macaques. Although some conflict management approaches attend to all three of these dimensions, we briefly discuss the macaque dimension (to discourage culling) and then focus our recommendations on the human dimension, as human behavior is typically the main driver of human–macaque conflict (Lee and Chan 2011; Sha et al. 2009). Given our finding that humans can influence macaques by their presence and by the availability of food, it seems that management efforts should take greater effort to mitigate the influences that humans have on macaques. Below we provide a brief summary of several management ideas. For a more thorough review of management recommendations for long-tailed macaques, see Jones-Engel and colleagues (2011).
Focus on Preventive Measures Instead of Culling
Interventions to manage human–macaque conflict are often made based on two related assumptions. First, overpopulation of macaques drives conflict. Second, reducing the overcrowded population will cause conflict to subside. However, little to no research has examined these assumptions. The effects of culling on the structure of populations in Southeast Asia are poorly understood. Widespread culling on Ngeaur Island, Republic of Palau failed to achieve the desired result of eradicating the population (Wheatley 2011). In addition, macaque populations are a natural part of their habitat and serve important functions such as seed dispersal (Dudgeon and Corlett 1994). This is especially important in places like Singapore where large vertebrates have become scarce (Lucas and Corlett 1998). Nevertheless, Singapore has heavily employed culling as a macaque management strategy (Khew 2014). We recommend that culling should become better informed through research and there should be a managerial focus on nonlethal techniques, as described in the subsequent sections. If culling continues to be used as a management technique, we recommend careful data collection to determine whether culling achieves the intended goal of reducing human–macaque conflict. For example, Lee and Chan (2011) reported that macaques are trapped and culled in response to nuisance complaints from the public. However, little effort is made to ensure that the individual caught in the trap is in fact the individual responsible for the complaints. Thus, a juvenile female could be caught and culled in response to complaints about an aggressive adult male. This will not resolve the source of the complaints and it may sensitize the macaques to the trap resulting in greater difficulty catching the target macaque in the future. Officials investigating nuisance complaints should gather sufficient information from the public, including photographs when possible, to identify a target macaque before culling. A studbook of identification sheets (which contain photos and identifying marks) for 596 individual macaques was created as part of the 2012 macaque census (Riley et al. 2013), and could aid with identification of specific macaques.
Enforce the Feeding Ban
The undesirable effects associated with macaques accessing human food—especially aggression—suggest that preventing macaques from eating human food is critical to any management program. In fact, preventing macaques from eating human food could be the single most important step in reducing human–macaque conflict. Benefits include preventing aggression from macaques to humans, decreasing macaque population growth rates, reducing disease transmission, and maintaining a healthier macaque population. Singapore has already taken an important step by banning macaque feeding and imposing large fines. In 2008, NParks doubled the monkey feeding fine to SG$500 (about US$380; NParks 2008) and enforcement also increased during that time (Lee and Chan 2011). However, monkey feeding still frequently occurs without intervention by officials. By stepping up enforcement, Singapore could reduce undesirable macaque behavior through removing access to human food sources. Yet another benefit is that the revenue from macaque feeding fines could be used to fund other macaque management initiatives.
In addition to stopping direct feeding of macaques by people, it is critical that management also curb indirect access to foods. At residences near nature reserves, most residents receive monkey-proof trash bins or use bungee cords to secure the bins against monkeys. When trash bins are improperly secured, macaques gain access to the refuse. Imposing fines on residents that improperly secure their garbage could make residential areas less attractive to macaques, reducing the time macaques spend there and the messes they make while scavenging, thus alleviating some forms of human–macaque conflict. Effective litter control in areas that macaques inhabit is also important.
Employ “Macaque Security Guards ” to Mitigate Conflict
“Macaque security guards” could be useful in Singapore for monitoring and directly managing human–macaque interfaces. Hong Kong has successfully used guards to patrol their areas of human–macaque interface and intervene in situations of potential conflict (Shek 2011). Because conflict often occurs near private residential condominiums, which typically employ private security guards, it would be practical and efficient for those guards to be trained in the mitigation and prevention of human–macaque conflict by appropriate wildlife authorities and experts. With some basic training, guards could assist people by helping them to navigate safely in areas with macaques, for example passing near macaques without having their grocery bags or food snatched. The guards would also be able to herd macaques away from residences, businesses, and busy streets. Furthermore, they could inform people in human–macaque interface zones how to best avoid interaction with macaques and what behaviors reduce the likelihood of conflict escalating. For example, guards can help ensure people keep food secure and out of sight, while also maintaining a safe distance from macaques. More professional guards associated with wildlife authorities could also be employed that could issue citations to those who engage in macaque feeding and the feeding fines could pay some of the salary for guards. If they were not empowered to issue citations, they could still remind violators that feeding macaques is illegal and call proper authorities. The nongovernmental organization Animal Concerns Research and Education Society (ACRES) has made some efforts to use macaque guards, but efforts could be increased (Khew 2014). Security guards could be especially helpful in urban areas, where human–macaque interfaces are more concentrated and thus easier to monitor than in rural areas, where interfaces are spread over larger areas.
Focus Efforts on Conflict Hotspots
Almost 15 % of our observations near roads occurred along a single road. This suggests the potential benefit of targeting macaque management efforts at particular hotspots that are especially likely to attract both macaques and people. Sha and colleagues (2009) identified six human–macaque interface hotspots in Singapore: Bukit Timah Nature Reserve, Upper Pierce Reservoir Park, Lower Pierce Reservoir Park, MacRitchie Reservoir Park, Upper Seletar Reservoir Park, and Rifle Range Road. This strategy would be most successful when paired with an ongoing macaque-monitoring program that can assist in identifying conflict areas.
Continue and Expand Current Educational Programs
NParks and other local agencies organize programs to educate the public about how to peacefully coexist with macaques (Lee and Chan 2011). Those programs include Monkey Walks, which are tours of nature parks that are led by a primatologist who explains macaque behavior and social structure to people while observing macaques (Ang 2010). Educational programs are valuable tools, but tailoring some programs specifically to residents who live near nature reserves could be useful, as these residents regularly engage with the local macaque population and are more likely to have negative attitudes toward macaques than park visitors (Sha et al. 2009).
Conclusion
Overall, Singapore has relatively mild human–macaque conflict when compared to human–macaque interfaces in other locations, especially considering that humans and macaques extensively overlap within a small, highly urbanized area. This relatively benign interface may be due in part to the fact that, compared to most other macaque habitat countries, Singapore is relatively affluent and has resources available for managing human–macaque conflict. In addition, the macaque feeding ban, although not one hundred percent effective, does reduce feeding to some degree and thus likely cuts down on conflicts and other issues (e.g., health problems such as obesity and disease transmission) that could arise from unregulated food access.
Our research showed that long-tailed macaques that access food in Singapore have larger group sizes, travel less, and use substrates differently from macaques that do not access human food. We made recommendations for decreasing macaques’ access to human food and more effectively managing the urban interface between humans and macaques. Our management recommendations apply specifically to Singapore, but they could potentially be useful for the management of other urban human–macaque interfaces. However, the most successful management strategies will be tailored specifically to the species, the human population with which they interface, and the local environment.
References
Aggimarangsee, N. (1992). Survey for semi-tame colonies of macaques in Thailand. Natural History Bulletin of the Siam Society, 40, 103–166.
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227–267.
Ang, Y. (2010, October 21). Don’t turn tail, get to know them instead; Guided walks to help residents, visitors cope with monkeys at Bukit Timah Nature Reserve. Straits Times. Retrieved from http://www.straitstimes.com
BBC News. (2010, October 7). Malaysian baby killed by macaque monkey. BBC News (London). Retrieved from http://www.bbc.co.uk/news/world-asia-pacific-11490575.
Beisner, B. A., Heagerty, A., Seil, S. K., Balasubramaniam, K. N., Atwill, E. R., Gupta, B. K., et al. (2014). Human-wildlife conflict: Proximate predictors of aggression between humans and rhesus macaques in India. American Journal of Physical Anthropology, 156(2), 286–294.
Berman, C. M., Matheson, M. D., Li, J.-H., Ogawa, H., & Ionica, C. S. (2014). Tourism, infant mortality and stress indicators among Tibetan macaques at Huangshan, China. In A. E. Russon & J. Wallis (Eds.), Primate tourism: A tool for conservation? (pp. 21–43). Cambridge, England: Cambridge University Press.
Chapman, C. A., & Rothman, J. M. (2009). Within-species differences in primate social structure: Evolution of plasticity and phylogenetic constraints. Primates, 50(1), 12–22.
Chauhan, A., & Pirta, R. S. (2010). Socio-ecology of two species of non-human primates, rhesus monkey (Macaca mulatta) and Hanuman langur (Semnopithecus entellus), in Shimla, Himachal Pradesh. Journal of Human Ecology, 30(3), 171–177.
Chopra, P. K., Seth, S., & Seth, P. K. (1992). Behavioural profile of free-ranging rhesus monkeys. Primate Report, 32, 75–105.
Dudgeon, D., & Corlett, R. (1994). Hills and streams: An ecology of Hong Kong. Hong Kong, China: Hong Kong University Press.
Ee, D. (2013, October 9). No monkeying around with ACRES rescue team; It seeks to help residents keep macaques away without culling them. Straits Times. Retrieved from http://www.straitstimes.com.
Epstein, J. H., & Price, J. T. (2009). The significant but understudied impact of pathogen transmission from humans to animals. The Mount Sinai Journal of Medicine, 76(5), 448–455.
Fa, J. E. (1992). Visitor-directed aggression among the Gibraltar macaques. Zoo Biology, 11(1), 43–52.
Feng, Z. (2011, October 7). Forest walk at HortPark: Attacks spark hunt for monkey. Straits Times. Retrieved from http://www.straitstimes.com.
Feng, Z. (2013, March 11). More reports on animal cases to AVA; Spike of about 20% from 2011 to last year comes as 24-hour hotline starts. Straits Times. Retrieved from http://www.straitstimes.com.
Feng, Z. (2015, April 6). Only a minority of Singaporeans want stray animals to be culled: Survey. Straits Times. Retrieved from http://www.straitstimes.com.
Fooden, J. (2000). Systematic review of the rhesus macaque, Macaca mulatta (Zimmermann, 1780). Zoology New Series, 96, i–180.
Fuentes, A. (2006). Human culture and monkey behavior: Assessing the contexts of potential pathogen transmission between macaques and humans. American Journal of Primatology, 68, 880–896.
Fuentes, A. (2010). Naturalcultural encounters in Bali: Monkeys, temples, tourists, and ethnoprimatology. Cultural Anthropology, 25(4), 600–624.
Fuentes, A. (2013). Pets, property, and partners: Macaques as commodities in the human-other primate interface. In S. Radhakrishna, M. A. Huffman, & A. Sinha (Eds.), The macaque connection: Cooperation and conflict between humans and macaques (pp. 149–166). New York: Springer.
Fuentes, A., & Gamerl, S. (2005). Disproportionate participation by age/sex classes in aggressive interactions between long-tailed macaques (Macaca fascicularis) and human tourists at Padangtegal Monkey Forest, Bali, Indonesia. American Journal of Primatology, 66(2), 197–204.
Fuentes, A., Kalchik, S., Gettler, L., Kwiatt, A., Konecki, M., & Jones-Engel, L. (2008). Characterizing human-macaque interactions in Singapore. American Journal of Primatology, 70(9), 879–883.
Fuentes, A., Southern, M., & Suaryana, K. G. (2005). Monkey forests and human landscapes: Is extensive sympatry sustainable for Homo sapiens and Macaca fascicularis in Bali? In J. Patterson & J. Wallis (Eds.), Commensalism and conflict: The primate-human interface (pp. 168–195). Norman, OK: American Society of Primatology.
Gumert, M. D. (2011). The common monkey of Southeast Asia: Long-tailed macaque populations, ethnophoresy, and their occurrence in human environments. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 3–44). Cambridge, England: Cambridge University Press.
Gumert, M. D., Hamada, Y., & Malaivijitnond, S. (2013). Human activity negatively affects stone tool-using Burmese long-tailed macaques Macaca fascicularis aurea in Laem Son National Park, Thailand. Oryx, 47(04), 535–543.
Hsu, M. J., Kao, C.-C., & Agoramoorthy, G. (2009). Interactions between visitors and Formosan macaques (Macaca cyclopis) at Shou-Shan Nature Park, Taiwan. American Journal of Primatology, 71(3), 214–222.
Imam, E., & Yahya, H. S. A. (2001). Management of monkey problem in Aligarh Muslim University Campus, Uttar Pradesh. Zoos’ Print Journal, 17(1), 685–687.
Jaman, M. F., & Huffman, M. A. (2013). The effect of urban and rural habitats and resource type on activity budgets of commensal rhesus macaques (Macaca mulatta) in Bangladesh. Primates, 54, 49–59.
Jones-Engel, L., Engel, G. A., Gumert, M. D., & Fuentes, A. (2011). Developing sustainable human-macaque communities. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 295–327). Cambridge, England: Cambridge University Press.
Jones-Engel, L., Engel, G. A., Heidrich, J., Chalise, M., Poudel, N., Viscidi, R., et al. (2006). Temple monkeys and health implications of commensalism, Kathmandu, Nepal. Emerging Infectious Diseases, 12(6), 900–906.
Jones-Engel, L., Engel, G. A., Schillaci, M. A., Rompis, A., Putra, A., Suaryana, K. G., et al. (2005). Primate-to-human retroviral transmission in Asia. Emerging Infectious Diseases, 11(7), 1028–1035.
Kamilar, J. M., & Baden, A. L. (2014). What drives flexibility in primate social organization? Behavioral Ecology and Sociobiology, 68, 1677–1692.
Khew, C (2014, October 23) Monkey see, monkey do as the guard says? Straits Times. Retrieved from http://www.straitstimes.com.
Klegarth, A. R. (in press). Measuring movement: How remote telemetry facilitates our understanding of the human-macaque interface. In K. Dore, E. Riley, & A. Fuentes (Eds.), Ethnoprimatology: A practical guide to research on the human-nonhuman primate interface (Chapter 8). Cambridge, England: Cambridge University Press.
Knight, J. (2011). Herding monkeys to paradise: How macaque troops are managed for tourism in Japan. Portland, OR: Brill Academic.
Lane, K. E., Lute, M., Rompis, A., Wandia, I. N., Putra, I. G. A. A., Hollocher, H., & Fuentes, A. (2010). Pests, pestilence, and people: The long-tailed macaque and its role in the cultural complexities of Bali. In S. Gursky-Doyen & J. Supriatna (Eds.), Indonesian primates (pp. 235–248). New York: Springer.
Lane-deGraaf, K. E., Fuentes, A., & Hollocher, H. (2014). Landscape genetics reveal fine-scale boundaries in island populations of Indonesian long-tailed macaques. Landscape Ecology, 29(9), 1505–1519.
Lee, B. P. Y.-H., & Chan, S. (2011). Lessons and challenges in the management of long-tailed macaques in urban Singapore. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 307–313). Cambridge, England: Cambridge University Press.
Li, C., Zhao, C., & Fan, P.-F. (2015). White-cheeked macaque (Macaca leucogenys): A new macaque species from Modog, southeastern Tibet. American Journal of Primatology, 77(7), 753–766.
Lucas, P. W., & Corlett, R. T. (1998). Seed dispersal by long-tailed macaques. American Journal of Primatology, 45(1), 29–44.
Malaivijitnond, S., & Hamada, Y. (2008). Current situation and status of long-tailed macaques (Macaca fascicularis) in Thailand. Natural History Journal of Chulalongkorn University, 8(2), 185–204.
Mallapur, A. (2013). Macaque tourism: Implications for their management and conservation. In S. Radhakrishna, M. A. Huffman, & A. Sinha (Eds.), The macaque connection: Cooperation and conflict between humans and macaques (pp. 149–166). New York: Springer.
Muehlenbein, M. P., Martinez, L. A., Lemke, A. A., Ambu, L., Nathan, S., Alsisto, S., & Sakong, R. (2010). Unhealthy travelers present challenges to sustainable primate ecotourism. Travel Medicine and Infectious Disease, 8(3), 169–175.
Nahallage, C. A. D., & Huffman, M. A. (2013). Macaque-human interactions in past and present-day Sri Lanka. In S. Radhakrishna, M. A. Huffman, & A. Sinha (Eds.), The macaque connection: Cooperation and Conflict between humans and macaques (pp. 135–148). New York: Springer.
Nahallage, C. A. D., Huffman, M. A., Kuruppu, N., & Weerasingha, T. (2008). Diurnal primates in Sri Lanka and people’s perception of them. Primate Conservation, 23(1), 81–87.
National Parks Board, Singapore. (2008). Tougher measure against monkey feeding. Retrieved May 16, 2015, from https://www.nparks.giv.sg/cms/index.php?option=com_news&task=view&id=55&Itemid=50.
National Parks Board, Singapore. (2009). Central Catchment Nature Reserve. Retrieved May 16, 2015, from https://www.nparks.gov.sg/gardens-parks-and-nature/parks-and-nature-reserves/central-catchment-nature-reserve.
National Population and Talent Division. (2013). A sustainable population for a dynamic Singapore: Population white paper. http://population.sg/whitepaper/downloads/population-white-paper.pdf.
Peterson, J. V., & Riley, E. P. (2013). Monyet yang dihargai, monyet yang dibenci: The human-macaque interface in Indonesia. In S. Radhakrishna, M. A. Huffman, & A. Sinha (Eds.), The macaque connection: Cooperation and Conflict between humans and macaques (pp. 149–166). New York: Springer.
Richard, A. F., Goldstein, S. J., & Dewar, R. E. (1989). Weed macaques: The evolutionary implications of macaque feeding ecology. International Journal of Primatology, 10(6), 569–594.
Riley, C. M., Jayasri, S. L., & Gumert, M. D. (2013). Singapore macaque listing: 2012. Report to the National Parks Board, Singapore, Singapore.
Riley, C. M., Jayasri, S. L., & Gumert, M. D. (2015b). Results of a nationwide census of the long-tailed macaque (Macaca fascicularis) population of Singapore. Raffles Bulletin of Zoology, 63, 503–515.
Riley, C. M., Koenig, B. L., & Gumert, M. D. (2015a). Observation of a fatal dog attack on a juvenile long-tailed macaque in a human-modified environment in Singapore. Nature in Singapore, 8, 57–63.
Riley, E. P., & Priston, N. E. C. (2010). Macaques in farms and folklore: exploring the human nonhuman primate interface in Sulawesi, Indonesia. American Journal of Primatology, 71, 848–854.
Seth, P. K., Chopra, P. K., & Seth, S. (2001). Indian rhesus macaque: Habitat, ecology and activity patterns of naturally occurring populations. Envis Bulletin: Wildlife and Protected Areas, 1(1), 68–80.
Sha, J. C. M., Gumert, M. D., Lee, B. P. Y.-H., Jones-Engel, L., Chan, S., & Fuentes, A. (2009). Macaque-human interactions and the societal perceptions of macaques in Singapore. American Journal of Primatology, 71(10), 825–839.
Shek, C.-T. (2011). Management of nuisance macaques in Hong Kong. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 297–301). Cambridge, England: Cambridge University Press.
Sheng, Y. K. (2012). The challenges of promoting productive, inclusive, and sustainable urbanization. In Y. K. Sheng & M. Thuzar (Eds.), Urbanization in Southeast Asia: Issues & impacts. Singapore, Singapore: Institute of Southeast Asian Studies.
Singapore Department of Statistics. (2015). Latest data. Singapore Department of Statistics. Retrieved April 12, 2015, from http://www.singstat.gov.sg/statistics/latest-data#14.
Singh, M., Kumara, H. N., Kumar, M. A., & Sharma, A. K. (2001). Behavioural responses of lion-tailed macaques (Macaca silenus) to a changing habitat in a tropical rain forest fragment in the Western Ghats, India. Folia Primatologica, 72, 278–291.
Singh, M., Singh, M., Kumar, M. A., Kumara, H. N., Sharma, A. K., & Kaumanns, W. (2002). Distribution, population structure, and conservation of lion-tailed macaques (Macaca silenus) in the Anaimalai Hills, Western Ghats, India. American Journal of Primatology, 57(2), 91–102.
Sinha, A. (2005). Not in their genes: Phenotypic flexibility, behavioural traits, and cultural evolution. Journal of Biosciences, 30, 51–64.
Sinha, A., & Mukhopadhyay, K. (2013). The monkey in the town’s commons, revisited: An anthropogenic history of the Indian bonnet macaque. In S. Radhakrishna, M. A. Huffman, & A. Sinha (Eds.), The macaque connection: Cooperation and conflict between humans and macaques (pp. 187–208). New York: Springer.
Sinha, A., Mukhopadhyay, K., Datta-Roy, A., & Ram, S. (2005). Ecology proposes, behaviour disposes: Ecological variability in social organization and male behavioural strategies among wild bonnet macaques. Current Science, 89(7), 1166–1179.
Southwick, C. H. (1972). Aggression among nonhuman primates. Reading, MA: Addison-Wesley.
Southwick, C. H., & Siddiqi, M. F. (2011). India’s rhesus populations: Protectionism versus conservation management. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 275–292). Cambridge, England: Cambridge University Press.
Thierry, B. (2007). Unity in diversity: Lessons from macaque societies. Evolutionary Anthropology: Issues, News, and Reviews, 16(6), 224–238.
Wheatley, B. P. (1999). The sacred monkeys of Bali. Prospect Heights, IL: Waveland Press.
Wheatley, B. P. (2011). Ethnophoresy: The exotic macaques of Ngeaur Island, Republic of Palau. In M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.), Monkeys on the edge: Ecology and management of long-tailed macaques and their interface with humans (pp. 252–272). Cambridge, England: Cambridge University Press.
Wheatley, B. P., & Putra, D. K. H. (1994a). Biting the hand that feeds you: Monkeys and tourists in Balinese monkey forests. Tropical Biodiversity, 2, 317–327.
Wheatley, B. P., & Putra, D. K. H. (1994b). The effects of tourism on conservation at the monkey forest in Ubud, Bali. Revue d’Ecologie, 49(3), 245–257.
Zhao, Q.-K., & Deng, Z.-Y. (1992). Dramatic consequences of food handouts to Macaca thibetana at Mount Emei, China. Folia Primatologica, 58, 24–31.
Acknowledgments
This research was funded by a grant from the National Parks Board of Singapore (NParks). Nanyang Technological University provided administrative, technical, and logistical support. We would especially like to thank NParks staff Tuan Wah Wong, James Gan, Claire Su Ping Ng, William Ng, and Ping Ting Chew for offering logistical support. The authors would also like to thank Agustín Fuentes and Sophie Borthwick for assistance with this project, and Michel Waller for inviting and reviewing this manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Riley, C.M., DuVall-Lash, A.S., Jayasri, S.L., Koenig, B.L., Klegarth, A.R., Gumert, M.D. (2016). How Living Near Humans Affects Singapore’s Urban Macaques. In: Waller, M. (eds) Ethnoprimatology. Developments in Primatology: Progress and Prospects. Springer, Cham. https://doi.org/10.1007/978-3-319-30469-4_16
Download citation
DOI: https://doi.org/10.1007/978-3-319-30469-4_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-30467-0
Online ISBN: 978-3-319-30469-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)