
Abstract
The way ecologists think about canopy biology as a scientific discipline could lead them to overlook different communities of spatially fixed organisms that may have properties usefully compared to or contrasted with forest canopies. This chapter represents a series of discussions and reviews on the possible nature and limits of canopy biology and introduces the prospect of a general comparative science of biological canopies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Architecture
- Biofilm
- Biodiversity
- Biomechanics
- Coral reef• Epiphyte
- Kelp
- Periphyton
- Rhizosphere
- Stratification
1 Summary
-
The way ecologists think about canopy biology as a scientific discipline could lead them to overlook different communities of spatially fixed organisms that may have properties usefully compared to or contrasted with forest canopies. This chapter represents a series of discussions and reviews on the possible nature and limits of canopy biology and introduces the prospect of a general comparative science of biological canopies.
-
Rather than restricting canopy biology to plants in terrestrial systems, I argue that canopy can be defined in terms of the parts of any community of sessile organisms that emerge from a substratum, the structural products derived from them included. This opens the field to diverse communities that could share many properties with forest (or plant) canopies. I overview the canopy literature on kelp forests, algal turfs, periphyton, bacterial films, and coral reefs. The word canopy has already been applied to each of these ecosystems. Periphyton and biofilms in particular have great potential as model systems for studying assembly rules for the physical structure and dynamics of canopies.
-
Among the similarities and differences between these types of canopy are the distribution of resources, such as light and nutrient gradients; factors affecting these distributions, such as the flow of air or water; and the resulting disposition of the species living within this matrix. Prioritizing a search for general principles must be a primary goal of comparative canopy biology because the microhabitats generated by stratification and other complex distribution patterns within communities are critical to sustaining global biodiversity.
-
I consider five rationales for the practice, the norm prior to this article, of limiting discussions of canopy biology to forests: if people have unique interactions with or concerns about forest canopies, if a substantive basis exists for treating trees as a distinct category of plant, if a substantive basis exists for treating trees as a distinct category in forests, if a substantive basis exists for treating forests as a distinct category of terrestrial community, or if attributes of the tree-crown residents and ground-rooted forest plants prove distinct. In no case is there unequivocal evidence for the usefulness of separating the study of forest canopies from the study of the aerial parts of other terrestrial plant communities.
-
In mainstream ecology, the organisms of a community are typically studied in two dimensions, or as isolated points on the earth. While canopy biology encompasses all aspects of the study of the portion of a community that projects into a medium, the discipline can in large part be distinguished as the science of treating plants (or other sessile hosts) as three dimensional.
-
In many studies of canopy species, the inhabiting organisms’ relationship to the canopy is treated as incidental. “Putting the canopy into canopy biology”—that is, contributing fundamentally to canopy biology as an independent field of study—requires that the attributes of the canopy form an integral part of the research. This would include issues of community ecospace; properties emerging from community residents in aggregate, such as stratification of microclimate; host distributions; host architectures; properties of a canopy’s structural elements; and characteristics of the open spaces within a community.
-
Applying the principles of canopy biology to communities as a whole, including those parts of sessile organisms and their associated species living on or in the substratum, suggests the value of developing a more comprehensive science, referred to here as structural ecology.
Words compartmentalize information, and that has many repercussions (Lakoff 1987). This affects how we identify subjects for academic study (Bates 1960; Hull 1988). The word canopy has been applied to vegetation in varied (Moffett 2000) and often inconsistent (Moffett 2002) ways, leading to varied interpretations of the domain of canopy biology. To select one example, if by canopy we mean the highest plant surfaces in a forest, as many authors do (e.g., Kricher 1997; the “outer canopy”), it is likely that, through habit, canopy biologists will develop a search image effective at picking out information only on the highest parts of the forest. In what philosopher W.T. Jones (in Bateson 1972) describes as the “topography of ignorance,” our knowledge of parallel and potentially useful studies on, say, shrubs and herbs, whether in forests or shorter ecosystems, or studies on the lower parts of forest trees, is likely to be marginalized. Such intellectual fragmentation is borne out by the literature. For example, citations on stratification in herbaceous communities, such as Monteith (1975/1976) or Liira et al. (2002), tend to be scarce in forest-canopy publications. One consequence can be independent discoveries by different academics, such as those working in agriculture, economic entomology, or landscape ecology, along parallel research tracks, thus wasting time and effort.
How, then, to delineate a discipline? While any “conceptual framing” can be arbitrary and subjective (Bohr 1955), the most compelling criteria are those of widest general importance to the experts. Consider the shifts in content and perspective between a biochemist’s focus on molecules and a cell biologist’s concern with structural integration, or a psychologist’s focus on mind and a sociologist’s interest in communities. In this respect, it makes little sense to grant canopy biology an independent status as a discipline if by canopy, we specifically mean “outer canopy.” After all, most aspects of the biology of the outer canopy change gradually from those features found among the plant organs lying beneath the topmost foliage. This is not to deny that some biologists must concentrate on the outer canopy because of its relatively pronounced features, such as the disproportionate significance of upper leaves to forest productivity.
What is the most fruitful basis for delineating canopy biology as a discipline? I will show that there is no clear evidence, for example, for trees or forests being distinct from other terrestrial vegetation. Then I consider modifications of the definition of canopy that would encourage canopy biologists to compare environments previously ignored by forest ecologists, such as coral reefs and bacterial films. Following this, I present some themes for an expanded canopy biology and consider the utility of establishing a field of study more comprehensive than canopy biology—one that applies the principles of canopy biology to communities of sessile organisms treated as a whole, including those with a portion of their anatomies positioned on or beneath a substratum. I conclude by addressing the central role of comparative canopy biology, as well as addressing this more expansive discipline, referred to here as “structural ecology,” as a means of interpreting patterns of biodiversity at a global level.
2 Seeing the Forest for the Herbs
The word canopy is often applied to the upper parts of forest ecosystems. Moffett (2001) argued for incorporating the literature on all aerial parts of any terrestrial plant community as canopy biology. Because attributes of this vegetation and the species residing within it are likely to change gradually, in a relatively continuous way from one community to the next, and within a community from one height to the next, I propose that we adhere to this approach except when dealing with “concepts or situations necessarily restricted to trees” (Moffett 2000). In fact, no one has specified any rationale for the common practice of restricting the scope of canopy discussions to trees or to forests (as in the useful distinction made between “tree canopies” and “forest canopies” by Shaw 1996). Consider five criteria by which forest canopies could merit this kind of separate attention (Fig. 3.1):

Forest cross section revealed where the bank of the Rio Napo has fallen away. One can identify a number of strata, including an emergent tree towering above its neighbors (at left); the overstory trees in direct sunlight; the understory trees in the shade of the overstory; and a shrub layer below those. Between the overstory and understory is visible a dark open space commonly used as a flyway by birds and bats. Because plant growth is dynamic with the height of each plant dependent on its age, species, health, and situation, including the distance and density of plant crowns overhead, each stratum in a community is unlikely to occur as exact layers at consistent heights, but instead will be “a product of localized conditions (e.g., varying jaggedly in a patchwork of vegetation types or successional stages), and, even in a uniform environment, it need not exist at one height above ground but rather may occur relative to the distance from the outer canopy” (Moffett 2000) (Photograph by Mark W. Moffett, Minden Pictures. All rights reserved)
-
1.
Humans have unique interactions with and concerns about forest canopies. Many people have grave concerns about the extinction of canopy species and great interest in the value of canopy products to societies past, present, and future. Because conserving the uppermost level of a forest is not possible without conserving its bottom, I would argue that conservation issues are most sensibly considered not specifically as canopy biology, but under the general rubric of ecology, which encompasses numerous aspects of economics, ethnography, and environmentalism.
-
2.
There is a substantive basis for treating trees as a distinct category. The diverse suite of characteristics associated with trees serves “as an example of the molding of the entire phenotype by selection pressures” that have come about convergently in numerous lineages (Niklas 1997). The ecological impetus discussed most often in reference to plant height is competition, especially for light (Tilman 1982; Givnish 1995; Leigh 1999; Sterck and Schieving 2007). The fundamental basis of “treeness” per se, however, may be biomechanical (Niklas and Kerchner 1984). Any self-supporting terrestrial plant growing beyond a certain height appears to be developmentally channeled into evolving a main vertical trunk built of the stiffest available structural elements (especially around its perimeter), surmounted by a branched crown—in short, it becomes a tree. The transformation seems to occur in a similar way under diverse environmental conditions, a result of gradual shifts from small herbaceous structural designs to an architecture that allows large upright plants to cope with bending or torsion (Niklas 2000). If many of these changes occur at some critical juncture during the evolution of increasing mass or stature (e.g., from 3 to 5 m in height: Givnish 1983), the tree-growth form could be sufficiently distinct to treat the study of the canopies in ecosystems in which the uppermost stratum is dominated by trees (i.e., forests) as an independent research discipline. Although this transformation process appears central to our very conception of “tree,” it remains poorly understood (Givnish 1984). Consider that trees allocate a large portion of their photosynthate to supportive tissues, and that, if they have high crowns, they pay a high price in aerodynamic drag, friction during fluid transport, and increased potential for structural failure (see Vogel 1996). Because of such costs, we would expect that plants would readily lose the tree-growth form when doing so would increase fitness, especially given that the character of woodiness, and, apparently, treeness, has been labile in plant evolution (Judd et al. 1994; Dodd et al. 1999; Kim et al. 2004). Nevertheless, trees in deserts, savannas, and other open ecosystems grow relatively very tall, even though they occur widely separated from their neighbors and so by ecological criteria seem conspicuously overbuilt. Tree species adapted to these habitats may be large because they use their trunks to store water (Holbrook 1995); depend on height to avoid herbivory, as arborescent cacti do from tortoises (Dawson 1966) and acacias from giraffes (Brooks and Owen-Smith 1994); are maximizing reproductive dispersal (Richards 1986); are avoiding damage from fast-moving ground fires (Givnish 1995); or are shading out grasses that compete with them for water (Walter 1973). Yet such factors appear neither pervasive nor severe enough to explain the almost ubiquitous occurrence of widely-spaced trees radically taller than the other plants in their communities. For example, giraffes always forage below 5 m, while savanna acacia trees often exceed 20 m in height (Young and Isbell 1991, T.P. Young, pers. comm.). Plant evolutionary mechanics could hold the key to this apparent mystery; further investigations in this area might shed light on the nature of “treeness” itself.
-
3.
There is a substantive basis for treating trees as a distinct category within forests. Although adult trees are usually assigned to strata separate from other plants in a forest, the question of whether mature specimens are distinct as a group or are part of a continuum that contains other, smaller forest plants has not been clearly resolved, in part because of inconsistencies among the research approaches to stratification (Parker and Brown 2000). In a frequency distribution of the size of mature vascular plant individuals, excluding vines, in a forest, is there a distinct peak (or peaks) corresponding with trees? Size-frequency distributions are common in studies of animal diversity, while, apparently, are absent from studies of complete plant communities (although some size-frequency distributions have been included in studies of only the trees: Poorter et al. 2008). This is presumably because modular construction and indeterminate growth can make plant size difficult to assess (e.g., Sterck et al. 2003).
-
4.
There is a substantive basis for treating forests as a distinct category. Forests could be considered a distinct category of vegetation if, by some parameter of community physiognomy, they can be separated from other terrestrial ecosystems in a nonarbitrary way. Suppose we graph some measure of overall community height—say, for each major community type in a classification of ecosystems, the modal height reached by the vegetation averaged over randomly chosen points on the ground. Suppose the distribution indeed turns out to be bi- or multimodal, such that forest systems represent a distinct peak. This would suggest that forests are more than an arbitrary construct that humans have split off from a continuum of natural communities. Perhaps on that basis, forest canopies can be distinguished as a separate research discipline. But so far, no information seems to exist on patterns of overall height across communities. If the available classifications are biased with respect to height (e.g., if ecosystem taxonomists have been “splitters” with respect to forest communities), or if community categories are largely artificial, at best representing opportunistic associations of species (Brown 1995), that could make attaining such information difficult.
-
5.
Attributes of tree-crown residents or ground-rooted plants in a forest prove distinct. If future studies demonstrate that forests harbor canopy communities distinct in some fundamental and reasonably abrupt way from the organisms dwelling on progressively lower kinds of vegetable communities, that might be taken as evidence for distinguishing forest canopies as a distinct biological entity. This seems unlikely, however, given that most resident canopy organisms respond not to height but to environmental factors that happen to correlate with height (Moffett 2000). Thus, epiphytes seemingly associated with high forest-canopy situations occur closer to the ground when conditions allow (McCune 1993; Benzing 2012). Biodiversity in forest canopies can be extreme, but since most inventories of biodiversity to date have been made in tree crowns (e.g., Stork et al. 1997; Foottit and Adler 2009), the relation between species diversity and community scaling is open to question. For any given latitude, how much of the high diversity of forest-canopy-dwelling species can be attributed to the canopies offering a relatively large overall mass, surface area, productivity, or microhabitat richness? There is also little basis to date for asserting that the organizational principles manifested by ground-rooted plants in forests (such as in the way the trees distribute horizontally or vertically) may be distinct from those operating in other communities, beyond matters of scaling that might be expected to vary in a reasonably continuous manner with successively shorter vegetative types (Moffett 2000).
In summary, there seems no unequivocal basis for the common practice of treating forest canopies independent of the study of the canopies of other terrestrial plant communities. Further general investigations into the nature of trees and forests, of course, may prove me wrong. As I have written elsewhere (Moffett 2000):
Furthermore, I encourage the application of “canopy” to all flora, reserving the phrase “forest canopy” to concepts or situations necessarily limited to trees. ...Broadening our perspective on canopies encourages us to pursue the reasonable hypothesis that most or all ecological processes scale up from a meadow to a redwood grove, so that problems considered intractable in the latter can be addressed by looking at shorter [and faster growing] systems.
3 More to Pond Scum than Meets the Eye
The Essence of knowledge is generalization. —
Hans Reichenbach (1951)
Incorporating into canopy science all the studies of all aboveground (aerial) plant organs and their occupants within any community (Moffett 2000) is a given, reflecting a history of the use of the word canopy with both cultivated and natural forb and grass communities (e.g., Monteith 1975–1976, Campbell and Norman 1989, Burrows 1990, Roxburgh et al. 1993, Hirose and Werger 1995). But this represents only a first step in the development of a truly comparative discipline. Indeed, though widely unappreciated by terrestrial “macrobiologists,” the word canopy is used by aquatic and microbial scientists to describe ecosystems that share many properties with terrestrial plant canopies. In contrast to terrestrial systems, where the hosts are usually vascular plants, these sessile organisms are typically algal species, zooxanthellae-bearing animals, or erect animal species such as hydroids and bryozoans. For an exception, see the information on seagrass communities, including issues bearing on canopy stratification, architecture, and biodiversity by Heck and Wetstone (1977), Kikuchi and Peres (1977), Williams and Heck (2001), and Pogoreutz et al. (2012).
The existence of plant-dominated aquatic systems and the remarkable parallels between these and other aquatic systems and terrestrial plant communities (described below) are among the reasons we should consider the common focus of canopy biology on terrestrial systems to be arbitrary and antiquated. To encompass these various kinds of hosts, canopy can reasonably be redefined as the parts of any community of sessile organisms that emerge from a substratum. Canopy biology (or canopy science) is by this criterion the study of the canopy of a community, including the organs of the sessile individuals and any affixed products of those organisms, and anything in, on, or between those organs and products. The affixed “products” can range from dead organisms, such as tree snags, to the secreted skeletons of corals or algal mucilage.
In this analysis, sessile describes an organism that emerges from or adheres to a substratum at positions fixed over a large part of its life history. Substratum refers to any surface or structural matrix that provides points of attachment for a sessile species, fixing the location of individuals or colonies. (I prefer this word to substrate, which is confusing, especially for microbial ecosystems, because it is also used to describe enzyme reactions.) The substratum thereby establishes the spatial relations between sessile individuals, including, to some degree, the organs that project into the fluid medium (the “canopy structure”). In turn, the sessile communities variously transform and stabilize the substratum (e.g., Stevenson 1996). (Many authors have applied the words structure and architecture to communities less literally than I have here—e.g., simply to denote biodiversity or nonphysical attributes of organization, such as niche spaces and trophic hierarchies; e.g., Connell 1975.) The substratum typically is a solid, but the air–water interface may give some level of stability to the relative position of organisms, such as in floating algal mats (metaphyton). I exclude from the canopy communities, or portions of communities, distributed entirely within the substratum matrix, as in the terrestrial soil community or its aquatic equivalent, the epipelic or bottom-sediment community (for instance, the microphytobenthos; MacIntyre et al. 1996). In some situations, the canopies of “different” ecosystems can be studied as one. In forest shade, stream-dwelling algae may show some of the same physiological adaptations as understory terrestrial plants (Robinson and Minshall 1986; Hill 1996).
The following sections summarize the canopy biology of diverse ecosystems.
Kelp Canopies. Describing kelp communities off South America, Darwin (1839) wrote, “I can only compare those great aquatic forests of the southern hemisphere with the terrestrial ones in the inter-tropical regions.” The term “kelp forest” has been common in the literature ever since. Application of the term “canopy” to kelp began with Jack Kitching, who, using a milk can with a window made from an old glass cookie box, was the first scientist to successfully dive into this ecosystem (Kitching et al. 1934). In these and other algal communities, there is a relationship between canopy height and algal growth form (Neushul 1972; Hay 1986; Steneck and Dethier 1994). All kelp forests convergently accommodate guilds of species that fall into five distinct “canopies,” or strata (including coralline crust as a stratum; Dayton 1985) (Fig. 3.2), the same number typically described for tropical forests (e.g., Richards 1996). The composition of the understory community depends on the distribution and diversity of the species above them (Wernberg et al. 2005; Irving and Connell 2006). The largest and most complex canopies occur in shallow, productive sites (Vadas and Steneck 1988), as might be predicted given that water attenuates light sharply as compared to air in terrestrial communities.

The two upper strata in a kelp forest off the Aleutian Islands of Alaska. An understory of Cymathera triplicata kelp grows under an overstory of another kelp species, Alaria fistulosa (Photograph by David Duggins. All rights reserved. Reproduced with permission)
Many findings from kelp forests parallel those for terrestrial communities, such as light attenuation through strata in relation to frond coverage (Gerard 1984), which at the benthos beneath kelp communities often declines to ca. 1 % of surface light, similar to tropical rainforests (Richards 1996); the importance of disturbance and gaps to succession (Neushul 1971; Foster 1975; Dayton 1975a; Hurby 1976; Pearse and Hines 1979; Reed and Foster 1984; Dayton et al. 1999); the value of sunflecks to understory growth and survivorship (Wing et al. 1993); differential survivorship and growth rates that result from the shading of benthic plants (Kastendiek 1982; Santelices and Ojeda 1984; Dean et al. 1989; Miller et al. 2011) and phytoplankton (Borchers and Field 1981); and other diverse competitive effects leading to specialized shade-tolerant (understory) communities (Dayton 1975b; Dayton et al. 1999). Unless predation on them is severe, sessile animals may outcompete kelp in low-light conditions in deeper parts of the benthos (Foster 1975), a pattern that holds to some extent in understory shade in shallower waters, though low-light adapted algae also occur there. Dayton (1971) distinguishes competition for space on the substratum (“primary space”) from competition within the volume above that surface, that is, within the canopy (“secondary space”), a concept worthy of widespread application. Because of their flexible tissues, kelp and other (Carpenter 1986) algal communities could in some ways be more ecologically comparable to grassland than to terrestrial forest (but see Holbrook et al. 1991), even though kelp can rise 50 m or more in height because of their reliance on the opportunities for flotation offered by water. The capacity for upward growth in kelp is by this means greatly enhanced over nonwoody terrestrial plants.
There have been general studies on the relation between canopy residents and kelp forest structure. Many fish stratify in a kelp forests, although this generally becomes less pronounced as the fish mature (Anderson 1994), and structural features of the forests influence fish diversity (Russell 1977). Shading by the kelp overstory can reduce algal growth rates in lower strata, thereby altering the abundance of some fish relative to canopy gaps (Carr 1989; Schmitt and Holbrook 1990; Jones 1992). Manipulations of physical structure are common in the study of kelp communities, showing, for example, that simplifying canopy structure can increase fish mortality by removing refuges (Anderson 2001). Predators can be so efficient at feasting on prey that pass through kelp canopies that recruitment of these prey species to ecosystems closer to shore is strikingly reduced (Gaines and Roughgarden 1987).
In macroalgal mats on boulders and within tide pools, canopy-resident diversity relates to algal architectural complexity (Dean and Connell 1987; Hacker and Steneck 1990; for a successional study of this kind of ecosystem, see Sousa 1979). Williams and Seed (1992) review the positive and negative effects of epiphytic animals on large algae.
Periphyton and Algal Turf Canopies. Periphyton (aufwuchs) constitute a “complex community of microbiota (algae, bacteria, fungi, animals, inorganic and organic detritus) that is attached to substrata” (Wetzel 1983; for further terms, see Weitzel 1979)—the microbial equivalent of an epiphyte mat. Indeed, periphyton can be epiphytic (Ruinen 1961, 1975; Morris et al. 1997; Claflin 1968; Whipps et al. 2008; for the rhizosphere equivalent, typically involving more bacteria than algae, see Pearce et al. 1995), albeit “periphyton” also applies to growth on nonliving or deceased substrata, such as submerged rock or leaf litter. Periphyton “have extensive vertical development on a small scale, and cells within the community matrix are tightly packed” (Boston and Hill 1991). They show a repeatable pattern of succession (Lowe et al. 1996; Sekar et al. 2004; Passy and Larson 2011), a pattern that can be disrupted by a high disturbance regime as occurs with communities on the surface of sand grains (Miller et al. 1987) except when algal mucilage binds the grains together, allowing further community development (Hoagland et al. 1982). Succession proceeds from a monolayer community to a stratification of species and chemistry within a matrix of cells and their secretions (Jørgensen et al. 1979; Jørgensen and Revsbech 1983; Kuenen et al. 1986; Lassen et al. 1994; Johnson et al. 1997; Fierer et al. 2010). Stalked microalgae can contribute to the greater depth of late-successional communities, resulting in “an upperstory of growth perhaps functionally analogous to the canopies characteristic of terrestrial forests” (Hoagland et al. 1982). Mature periphyton communities often have a dense understory of small cells overgrown by filamentous algae, though some interesting variants on this are shown in Fig. 3.3. The upper stratum provides attachment points for diatoms that live as epiphytes (Roos 1979; Roemer et al. 1984; Marks and Power 2001), described as “dependent organisms” or pseudoperiphyton, and treated as part of the same community as their hosts (Sládecková 1962). In addition, there appear to be algae that are the functional equivalent of vines (or more accurately, hemiepiphytes; Fig. 3.3b). While some sessile algae cannot adhere to other algae and so require a direct connection with the substratum, other species may preferentially attach to algae in the layer below and thereby epiphytically form a canopy stratum of their own (Fig. 3.3a). This strategy is unknown and probably biomechanically impossible within terrestrial canopies. Vines positioning their foliage uniformly above that of their hosts (Putz 1995) may approach it, although being rooted to the ground, these canopy plants are not epiphytes.

Multilayered periphyton attached to a screen in a freshwater aquarium (a) and to a screen in an Algal Turf Scrubber (b). Typically the layer directly attached to the screen (the hard rock or carbonate substratum in the wild) is dominated by either blue green (cyanobacteria) or a diminutive green (Stigeoclonium) or diatom algae (W. Adey, pers. com.). In (a), the upper layers are attached to the layer below them and not the substratum, and so these entire strata are epiphytic on lower strata. The middle layer consists of diatoms and small filamentous algae suspended in mucilage generated by the community. In contrast, when the overstory is made up of large, typically branched filaments (b), these are also attached to the substratum, along with the blue-green or small green and red “understory” algae. Much like vines (especially nomadic ones: Moffett 2000), the Spirogyra and Dichotomosiphon in 3b spiral around their supporting hosts like vines, apparently moving along them by rotation and circumnutation sensu Darwin (Yeh and Gibor 1970); in monoculture, the Spirogyra will even coil around one another like plant tendrils with no alternative place to go. These two illustrations show what I would call “monodominant periphyton,” because only one species is affixed to the substratum, but more biodiverse communities clearly exist (Both illustrations are from Adey and Loveland (2007). Reproduced with the permission of the authors. All rights reserved)
Canopy complexity of periphyton may be greater at sites with higher light intensities (Hudon and Bourget 1983). Furthermore, the internal physiognomy depends on local flow regime, in part because greater turbulence increases the penetration of nutrients and light (Peterson 1996). Wetzel (1993) argues, however, that such penetration is difficult (unless aided by grazers: Hillebrand et al. 2008) and that the high productivity of periphyton is a result of efficient recycling of nutrients within their canopies. Growth of the outer layer can shade the strata below (Johnson et al. 1997; Dodds et al. 1999) and block nutrient inputs to the understory (McCormick and Stevenson 1991; Peterson and Grimm 1992). Shading can lead to understory deterioration (Stock and Ward 1991) in time causing a community to slough from its substratum (Meulemans and Roos 1985). Substrata are colonized both by these detached communities (detrital microcosms; Korte and Blinn 1983) and by isolated cells in suspension (Stevenson 1983). Sloughing can be reduced where understory algae species are able to produce more photopigments or become increasingly heterotrophic in dim conditions (Tuchman 1996; Peterson 1996), in which case the periphyton can last longer and achieve higher biomasses. Because of the cycle of growth, death, and sloughing, the community that establishes after a site is grazed by herbivores may depend on the prior successional status at the site (Peterson 1996), among other factors (Tuchman and Stevenson 1991).
Turfs are filamentous periphyton communities, typically a few millimeters high (Fig. 3.4) that occur widely on coral reef surfaces and produce the bulk of reef primary productivity (Adey and Steneck 1985). Disturbances (say, by herbivores or abrasion from the movements of taller algae overhead; Russell 2007) reduce turf height and increase light penetration and turbulence through their canopy (Carpenter 1986; Williams and Carpenter 1990; Carpenter and Williams 1993; Cheroske et al. 2000). Stratification can occur within turfs, but is limited (Hackney et al. 1989; R.C. Carpenter, pers. comm.).

Algal turf from St. Croix dominated by Herposiphonia. Width of the image is about 1 mm (SEM photography by Robert S. Steneck. All rights reserved. Reproduced with permission)
Bacterial Films. Almost all bacteria live packed within surface-bound multispecies communities called biofilms (Watnick and Kolter 2000). Until the 1980s, bacteria were studied mainly by traditional sampling and culture methods. Extrapolations from monospecies planktonic laboratory cultures led to serious misunderstandings about bacterial ecosystems (Costerton et al. 1995). Especially in light-exposed habitats, cyanobacteria and their accumulated remains can induce thick perennial accretions called microbial mats or stromatolites (Stal 2000), but non-photosynthetic mats thrive in dark habitats where there are few grazers (Teske and Stahl 2001).
Biofilms often intergrade with eukaryote-dominated periphyton, and increasingly the term has been used broadly to include periphyton and even fungi (Reynolds and Fink 2001; Ramage et al. 2009). In the algal-dominated systems discussed in the previous section, for example, either bacteria or algae can colonize early in succession (Hoagland et al. 1982; Roeselers et al. 2007), and pioneer species of bacteria, in combination with certain diatoms and fungi, may “precondition” the substratum for adherence of subsequent algae, or provide the foundation they require for attachment (Korte and Blinn 1983; Burkholder and Wetzel 1989; Hodoki 2005). In a developed periphyton community, the bacteria can be nutritionally dependent on excreted algal products (Haack and McFeters 1982; Sobczak 1996), but a reverse dependency can arise when nutrients are scarce (Scott et al. 2008), suggesting facultative mutualisms can develop (cf. Carr et al. 2005). The remainder of this section will focus on communities dominated by bacteria, which have become the most extensively studied canopy system.
For macroscopic canopies, interest typically falls into two arenas: studies of the substratum-bound species themselves (the hosts) and studies on species that live within the canopy generated by the hosts (canopy residents). This distinction is apparently of limited utility for bacterial biofilms and mats, which are formed of cells en masse, with a very strong coupling of structure and function. Light penetration and chemical gradients are critical to the makeup of such communities, where the concentration of any nutrient consumed in the biofilm typically decreases with depth (Jørgensen and Revsbech 1983; Kühl et al. 1996; Wimpenny and Kinniment 1995; Stal 2000; Ward et al. 2006; Stewart and Franklin 2008), such that anaerobic or anoxic species often occupy the depths of a film (Jørgensen et al. 1986; Sagan and Margulis 1988, p. 53–54; Ramsing et al. 1993; van Gemerden 1993). More complex aspects of spatial heterogeneity in these communities are becoming evident (Kühl and Polerecky 2008). The internal organization of photosynthetic biofilms and mats often includes stratification in relation to light levels (Kühl and Fenchel 2000; Ward et al. 2006; Ramsing et al. 2000; Al-Najjar et al. 2012).
While most biofilm residents are sessile or at least relatively immobile within the cell matrix (Costerton et al. 1995), the integrity of biofilms may be as much a product of a secreted matrix of polymers, which are complexly structured (Lawrence et al. 2007) and essential to biofilm formation, as of the fixed location of organisms (Flemming and Wingender 2010). Thus, while some species are held in space by intercellular connections (Schaudinn et al. 2007), motile species can shift position within the film—for example, moving within the matrix to different “microzones” (strata)—in response to light or chemical cycles (Doemel and Brock 1977; Garcia-Pichel et al. 1994; Stal 2000); this is also true of some periphyton-dwelling diatoms (Johnson et al. 1997). Such movements can bring about distinct biofilm morphologies (Klausen et al. 2003). Biofilm residents are phenotypically distinct from conspecific planktonic forms, which function as a dispersal stage. The sloughing of bacteria from films, adhesion of the colonists to a surface or within the matrix, and other aspects of establishment and development of films have been documented or modeled in three dimensions (Bryers 2000; Kierek-Pearson and Karatan 2005), as has the ecological succession of biofilm residents through time (Lawrence et al. 1995; Korber et al. 1995; Jackson 2003; Fierer et al. 2010).
The matrix of bacteria in a biofilm was originally described as developing channels or pores through which nutrients enter and wastes leave the community (Costerton et al. 1994; Massol-Deyá et al. 1995; Stoodley et al. 1999a). The structure of many biofilms is so partitioned into independent units by these “waterways” that on close inspection, they do not resemble a “film” at all but rather a landscape of cone-shaped and mushroom-shaped bodies called “microcolonies” distributed intriguingly like trees in a forest (Fig. 3.5). When mature microcolonies grow into contact, they do not fuse but rather they wave past each other when shear forces operate (Stoodley et al. 1999a; J.W. Costerton pers. com.). Microcolony formation is widespread, but not universal (Wimpenny and Colasanti 1997), in natural and artificial systems, both for monocultures and for mixed-species communities, and is presumed to result from nutrient limitation and niche exploitation (Costerton et al. 1995). The distribution of microcolonies (including both the cells and the exopolymeric materials they secrete) and the channels and other spaces between them is referred to as biofilm architecture (Lawrence et al. 1991). Both the species composition and the nature of the substratum affect this architecture (Costerton et al. 1995; Wimpenny and Colasanti 1997). So does water flow around the films (Beyenal and Lewandowski 2002), which may cause entire microcolonies to drift across a surface (Stoodley et al. 1999b; Rupp et al. 2005; Venugopalan et al. 2005), straining the definitions of “sessile” and “canopy.” There is a close coupling between such structural patterns, water flow, and the chemical landscape within such biofilm canopies (Staal et al. 2011).

Bacterial films. The schematic drawing of a generalized biofilm shows microcolonies separated by channels. Streamers develop as water flow increases, with the arrow indicating the water flow penetrating the film (drawing by Peg Dirckx, reproduced with permission). Below is a confocal scanning laser microscopic image of microcolonies composed of the single bacterial species Pseudomonas aeruginosa (E.S. Lauchnor and B. Pitts, Center for Biofilm Engineering, Montana State University. Reproduced with permission)
The structure within and between microcolonies “provides very diverse habitats on a small scale, favoring biodiversity” (Flemming and Wingender 2010). Microbiologists have described biofilms as coordinated communities with “primitive homeostasis, a primitive circulatory system and metabolic cooperativity” that can “resemble the tissues formed by eukaryotic cells” (Costerton et al. 1995; Costerton and Lappin-Scott 1995). As Foster (2010) points out, such analogies can be overdone in that there is still little evidence of coordination in biofilms, particularly between species. Nevertheless, there appears to be a stronger interdependency between different taxa in a biofilm than is typically described between most plants in a forest, such that certain bacteria depend on the metabolic products of other microbes (Kühl et al. 1996; van Gemerden 1993; Molin et al. 2000; Paerl et al. 2000) and show other forms of cooperation (Crespi 2001; Mitri et al. 2011) and communication (Davies et al. 1998; Irie and Parsek 2008). Coordination can arise through rapid evolution (e.g., of parasitisms; Hansen et al. 2007) in part through gene transfer within the films (Molin and Tolker-Nielsen 2003; Madsen et al. 2012). In addition to these mutualistic “consortia,” biofilms and microbial mats are home to diverse competitive and predator–prey interactions (Lawrence et al. 1995; Allison et al. 2000; Foster and Bell 2012; van Gemerden 1993). A question wide open for investigation is how selective forces and the potential for intense competition among different strains and species can lead to the seemingly cohesive systems that appear stable, functionally integrated, and structurally convergent (Nadell et al. 2008).
Stolzenbach (1989) and subsequent authors have applied the word “canopy” to biofilms, and microbiologists have been increasingly interested in employing more integrative approaches (e.g., from landscape ecology: Battin et al. 2007). At the same time, new technological advances allow combined mapping of structure, composition, and function in biofilms (Kühl and Polerecky 2008). Indeed, microbial communities can be easily manipulated for studies of canopy structure: Various combinations of microbial species or strains can be mixed, centrifuged, and allowed to form biofilms to investigate assembly rules for canopies under specified nutrient or substratum regimes. While most biofilm studies rely on well-defined communities of one or a few species, formation of biodiverse microbial mats can be induced by treatment of natural sediments (Kühl et al. 2003). All canopy biologists can benefit from knowing about biofilm research (e.g., Guerrero et al. 2002). However, the central role of a polymer matrix and the frequency of facultative symbioses in bacterial communities suggest that some of the algae-based periphyton could be more straightforward microbial analogs of plant communities.
Coral Reefs as Canopies. Dahl (1973) writes of coral reefs that “organisms often occur in many layers and the substratum itself is organism generated.” Not surprisingly, overarching corals have been described as producing a “canopy” overtopping an “understory” community (Baird and Hughes 2000). While a coral’s supportive structure is not living, it is an immediate by-product of living things and therefore can be treated as a part of a canopy in much the same way as snags are treated as part of a terrestrial canopy. Actually, there is a veneer of living tissue on live coral, much as there is a small zone of living phloem surrounding the mostly “dead” xylem of trees, so that in fact in both ecosystems, a large part of the structural foundation of the canopy is dead.
Somewhat like trees (Horn 1971), the architectures of photosynthetic corals change with light regime (Porter 1976). Death or suppression of the growth of corals from shading by other corals results in an understory that can include shade-tolerant phototropic species (Stimson 1985; Anthony and Hoegh-Guldberg 2003) and that incorporates abundant sessile heterotrophs (Karlson 1999; Baird and Hughes 2000). Other shade-tolerant (or, in the case of sessile animals, shade-indifferent) species can densely occupy the undersurfaces of the corals themselves (Jackson et al. 1971; Maida et al. 1994). Colonization of these habitats can depend on active larval choice for “cryptic” (shaded) microsites (Maida et al. 1994; Mundy and Babcock 1998). The change of species composition with shading can parallel community changes resulting from light falloff with depth in the water column, such that understory shade permits certain deepwater species to extend their distribution into shallow water (an effect for which there is no terrestrial equivalent). In general, however, these understory corals are distinct from reef-building corals, as the latter, when adapted to dim conditions, tend to occur in deeper or more turbid water. Nutrient input to the understory is strongly dependent on wave action (Lowe et al. 2008). Other species prosper in the gaps formed by the death of overstory coral colonies, yielding a habitat mosaic (Stimson 1985). The coral reef community includes algae (among them the turfs discussed previously) that show a complex pattern of competition depending on their heights and interfrond densities (Steneck 1997). The structural complexity of coral reefs (including that of the algae within them: Levin and Hay 1996; see review by Graham and Nash 2012) provides for a high abundance and diversity of resident organisms, such as fishes (Dahl 1973; Steele 1999; Holbrook et al. 2003), clinging invertebrates (Vytopil and Willis 2001; Fraser and Sedberry 2008), and zooplankton (Porter 1974). Species richness declines when this complexity degrades (e.g., as a result of environmental disturbances: Wilson et al. 2007b; Graham et al. 2009; Alvarez-Filip et al. 2009, 2011).
Other Canopies. Various other sessile animals form dense aggregations that could be studied as canopies including both clonal (e.g., anemones, hydroids, bryozoans, ascidians) and nonclonal species such as mussels and barnacles (Paine and Suchanek 1983), for example, in fouling communities (Boyle et al. 2007). Further, if we allow that communities of these organisms have canopies, we can apply the idea of an extended phenotype (i.e., extending the definition of phenotype to include nonliving products of an organism such as nests or retreats: Dawkins 1982; Turner 2000) to enlarge the concept of canopy almost indefinitely, depending on our interests. Least controversial would be static structures like coral skeletons whose architectures and spatial relations are produced directly by living things that serve as “ecosystem engineers” (Jones et al. 1997). Tubes of polychaetes (Bell and Coen 1982) and stream fly larvae (Pringle 1985) attract assemblages of plants and animals. These structures recolonize rapidly after defaunation, forming communities organized around tube architecture (Bell and Coen 1982). On land, patches of earthworm castings (Maraun et al. 1999; Aira et al. 2009) or of fungal fruiting bodies (O’Connell and Bolger 1997) are possible analogs of plant canopies.
Systems that are not canopies by any definition could be useful models for examining certain features of canopy life. Suspended bacteria can stratify under conditions of low turbulence (Guerrero and Mas 1989). Studies of the planktonic cells show that “increased productivity produces a physical scaffold to support biological heterogeneity (as, for example, in the spatial complexity of forest canopies) on which other species can build” (Morin 2000).
4 The Geometry of Canopy Biology
If we expand our definition of canopy beyond what I suggested in Moffett (2000), to encompass all parts of any community of sessile organisms that project into a medium, on what basis might the discipline of canopy science rest on firmer (and more fruitful) ground?
Ecologists traditionally treat plants (or other sessile organisms) in two dimensions, or as points on the earth. While such topics as orientation in canopy ants, the conservation of orchids, and the foliage uptake of pollutants bear on canopy biology (i.e., sensu Moffett 2000), much of the research on these topics falls squarely within this tradition. For example, most studies of orientation in canopy ants ignore 3-D spatial issues bearing on plant topographies, as well as other issues that could potentially be unique to canopy substrata, such as the properties of pheromone diffusion from bark as opposed to that from leaves. In a typical experiment, ants are not studied in their canopy environment, as when a species normally found on foliage is housed on a flat laboratory surface. While we can learn a great deal about canopy organisms with this approach, none of it has to do specifically with their origins in the canopy.
By contrast, the core of canopy biology as an independent discipline can be characterized in large part as the science of treating communities of plants (or other sessile organisms) as three dimensional. This is becoming increasingly tractable even for whole forests (Shugart et al. 2010). More generally, creating a robust canopy science requires us to “put the canopy into canopy biology” through research that directly contributes to understanding the aspects of life associated with sessile organisms. This can be achieved by introducing the “z” axis or other canopy attributes (as has been done in the studies on ant orientation by Jander and Voss (1963), Beugnon and Fourcassie (1988), Fourcassie and Beugnon (1988), Jander (1990), Klotz and Reid (1992), and Wohlgemuth et al. (2001)). To take one example, the book Geometry of Ecological Interactions, by Dieckmann et al. (2000), conforms to the 2-D “mainstream tradition” except for some material on three-dimensional gap structure, which is the only part of their coverage of ecological geometry that, by my criterion, represents canopy research. The same philosophy applies to aquatic systems, which offer unique experimental opportunities: Consider the value of artificial reefs of varied architecture in understanding the productivity and diversity of canopy residents (Carr and Hixon 1997).
Moffett (1999) outlined attributes falling into six general categories that put the canopy into canopy biology, calling these the discipline’s core issues. By framing questions with respect to one or more of the categories (reviewed below), researchers can fundamentally contribute to canopy biology as a discipline. To keep my treatment brief, I have chosen examples from the forest literature alone, although information on each category is available for other kinds of canopies as well. While most scientific results can be partitioned along these categories, they clearly are not independent issues; many studies make important contributions when looking at two or more of the six. Ultimately, much of canopy science may codify according to how these attributes have contributed, over ecological and evolutionary time, to the diversity of both the host organisms and their occupants.
-
1.
Community Ecospace. For any canopy, the quantity and quality of space available to the canopy dwellers depend on the host structure. How much so is a matter of conjecture. Any small, nimble bird or agile climbing animal, such as a gibbon, seems to experience canopies as a volume, although even here, not all points in the volume may be accessible: Vegetation may be too dense for a flying bird (Cuthill and Guilford 1990) or too widely spaced for primates to cross (Cant 1992; Cannon and Leighton 1994). Small, flightless arthropods, such as ants and earthworms, are unlikely to register community ecospace in this way. Ants are restricted to mere millimeters of every surface within their territory, such that—despite models to the contrary (Hölldobler and Lumsden 1980)—ants experience canopies as something between a 2-D and 3-D space (Moffett 1994, 2010). In essence, a canopy represents for them a highly warped surface. Like a science-fiction ship using a wormhole to bridge points normally experienced as quite distant from one another, weaver ants create shortcuts through this space by linking their bodies into chains, thus enabling them to quickly access new branches (Hölldobler and Wilson 1977). In this way, they can bridge entire tree crowns that otherwise could be reached only by way of a long march down to, across, and up from the ground.
-
2.
Aggregate Properties of the Community. Here, I include the nonuniform vertical distribution of canopy species and their architectural parts (Fig. 3.1) and the concomitant stratification of other elements of the canopy environment, such as microclimate (other gradients can exist, such as radially from the interior to the exterior of a tree crown [Freiberg 1997] or a bacterial microcolony [Stewart and Franklin 2008]). Parker and Brown (2000) (cf. McElhinny et al. 2005) criticize studies of stratification for their lack of reproducibility, inconsistent terminology, and other weaknesses. Regardless of the difficulties, understanding stratification is at the core canopy science, and to accommodate a variety of research interests, the word is best applied flexibly (Moffett 2000). While many studies of terrestrial nutrient interception treat the canopy as a single “black box” with overall inputs and outputs (e.g., Coxson and Nadkarni 1995), some researchers have uncovered complex internal patterns within canopies that act as atmospheric filters (Wiman et al. 1985; Meyers et al. 1989; Lovett and Lindberg 1992) and nutrient transfer systems (Pike 1978; Reiners and Olson 1984; Coxson et al. 1992; Lindo and Whiteley 2011). Defining strata or gradients requires broad spatial averaging, while explaining emerging patterns necessitates research at a finer spatial scale. For example, bark pH can vary with height in the trees (Hyvärinen et al. 1992). If the height distribution of an epiphyte corresponds generally to that of a certain bark pH, we could propose that these plants prefer that pH. Testing this hypothesis requires determining the detailed distribution of bark pH at the locations occupied by the epiphytes (e.g., Gauslaa 1995) and then manipulating the pH in the field or a laboratory (e.g., Hallingbäck 1990). In short, as I wrote about stratification elsewhere (Moffett 2000): A common finding is that short distances traversed vertically in canopies are equivalent in effect to changes likely to occur over much greater horizontal distances (e.g., Geiger 1965; Russell et al. 1989), demonstrating the critical importance for the height dimension as an environmental determinant. The vertical richness in microhabitat may be the primary reason so much diversity packs into structurally complex ecosystems. This could explain the high alpha diversity and low beta diversity of epiphytes in relation to other plants (McCune and Antos 1981), intimating the utility of quantifying a vertical component to beta diversity (DeVries et al. 2012).
-
3.
Host Distribution. Many canopy species may be specialists at residing on one or a few plant taxa (e.g., Erwin 1982) suggesting that hosts can be considered islands over evolutionary time (Janzen 1968, 1973) (Fig. 3.6), which could help in modeling the processes that occur within and between plants that generate patterns of species distribution and diversity. Rather than treating hosts as islands in a uniform ocean to conform to the perspective of MacArthur and Wilson (1967), “patchwork” biogeography models could treat communities as a continuum of host islands of varied acceptability as sources of food, retreats, or transit routes for each canopy-dwelling species. Diversity is further organized on scales both smaller and larger than that of a host. A distinct community can develop on any stable canopy element that qualifies as an island sensu Haila (1990), such as a flower (Seifert 1975), a phytotelmatum (Jenkins and Kitching 1990; Richardson 1999), a leaf (to a microbe: Andrews 2006), or even certain microclimatic features (Herwitz and Slye 1992). Even ant territories could form habitat islands within tropical canopies (Moffett 2010, p. 132–133). The territories of different ant species are distributed as a mosaic that overlays, but are partially independent of, the mosaic of tree crowns (e.g., Dejean et al. 1999). Ants scour their territories to drive off intruders and kill prey while promoting the survival of species-specific assemblages of associates (Hölldobler and Wilson 1990). As with other island-like canopy features, the persistent domination of different aggressive ants with large colonies across the canopy volume may add to the potential for other species to pack into vegetable space.
Fig. 3.6 
The patchwork of different tree species in a forest canopy is shown here by the distribution of gaudy flowering trees (Vochysia ferruginea) in the Osa Peninsula of Costa Rica (Mark W. Moffett/Minden Pictures. All rights reserved)
-
4.
Host Architecture. All canopies, from redwood forests to biofilms (Lawrence et al. 1991), have varied architectural parts. In forest ecology, a burgeoning literature on this topic covers the size, angles, distributions, development, and spatial relations of aerial plant parts. Classically, the models of Hallé et al. (1978) have been used to describe the architecture of trees (Fig. 3.7). One key practitioner considers the “Hallé-Oldeman architectural model” classification “comparable to the development of the binary system of nomenclature by Linnaeus” (Tomlinson 1983). Nonetheless, the system has been little used by non-morphologists, arguably in part because of the overall neglect of the potential importance of substratum architecture on canopy organisms (but see Hallé 1990). There are studies of the effects of simple architectural attributes, especially branch angle and width, on the growth of epiphytes (e.g., Rasmussen 1975; Ingram and Nadkarni 1993; Moe and Botnen 1997; Zotz 2006) and on animal locomotion, particularly in reptiles (Losos and Greene 2009; Byrnes and Jayne 2012) and primates (Demes et al. 1995; Povinelli and Cant 1995; Dagosto and Yamashita 1998; Hamrick 1998; McGraw 1998; Young 2009, just to mention a few citations on positional behavior). Yet few broadly scaled studies of how community-level aspects of plant architecture influence canopy life have been done, excluding some examples for insects (e.g., Lawton 1983; Sinoquet et al. 2009). Consider that many tropical canopies have well-beaten vertebrate highways extending from tree to tree, which can be detected by the epiphytes that spread to each side of a branch like hair from a part (Perry 1978; Sillett et al. 1995). Perry (1978) found evidence of multispecies use and active pruning of these trails. But as yet, no one has mapped such a trail in relation to the tree architectures available locally, or documented how the trail originates, how long it lasts, or how usage shifts with changes in canopy structure and resource availability.
Fig. 3.7 
The architecture of canopy species is exemplified by an immature Ceiba pentandra tree from Cameroon, showing the “bicycle spoke” branching pattern of Massart’s architectural model (Mark W. Moffett/Minden Pictures. All rights reserved)
-
5.
Open Space. Not even biofilms are a continuous matrix of organisms: All canopies consist of a framework occupying a dynamic fluid matrix that has open communication and exchange with the adjacent atmosphere or hydrosphere, which typically includes the bulk of canopy volume (Chiarucci et al. 2002). Free space (air or water) within canopies merits special consideration, because of its potential effects on microclimate and on the locomotion or dispersal of organisms. Many aspects of this topic remain virtually ignored. Space between forest trees is commonly distinguished through the use of two categories: gaps, the result of plant death, and the openings resulting from shyness, which is often the result of diminished plant growth and reconfiguration—that is, plant foraging (Hutchings and de Kroon 1994), although physical abrasion can also be involved in shyness patterns (Franco 1986) (Fig. 3.8). Gaps are commonly studied spaces, because of their role in forest succession and species-diversity patterns (Lieberman et al. 1989). Spaces represent barriers to some species and pathways (flyways) or resource (light) access and growth opportunities to others (e.g., Aluja et al. 1989; Brady et al. 1989; Cuthill and Guilford 1990; Cannon and Leighton 1994; Brigham et al. 1997; Aylor 1999; Svenning 2000; Montgomery and Chazdon 2001; Dial et al. 2004; Randlkofer et al. 2010; Byrnes and Jayne 2012). Little information exists, however, on how the distribution of open space might be involved in structuring the populations of canopy residents. There is a tendency to think in terms of canopy structures such as trunks and branches when the space between structures could be the resource used, as might be the case among gliding animals (Moffett 2000), though competition for space per se is seldom likely (Wilson et al. 2007a). Within the open spaces, boundary layers—the regions of lowered fluid velocity that exist around any surface in a turbulent medium—are a general feature of attached communities. Their presence partially isolates canopies from the surrounding medium and thereby can increase community reliance on efficient and potentially autogenically controlled internal (within-canopy) nutrient cycling. This isolation may be particularly important in flowing water (Mulholland 1996), where canopy physiognomy can substantially ameliorate the downstream displacement of chemicals or of any organisms that are moving within a canopy or that have a poorly developed capacity to attach to a substratum.
Fig. 3.8 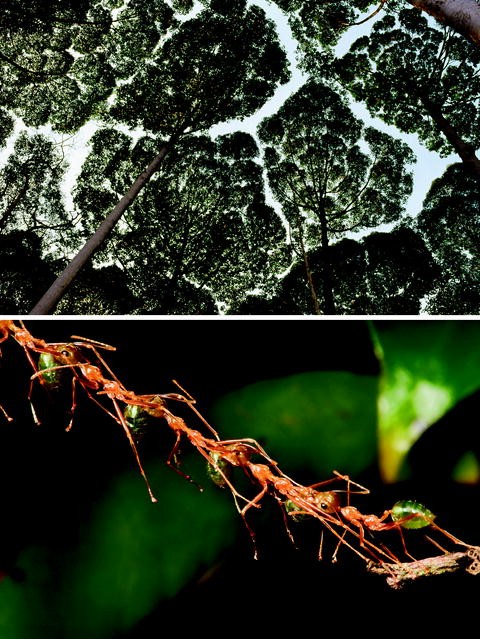
Open spaces within forests include shyness between tree crowns, as shown in a grove of Dryobalanops lanceolata trees in Peninsular Malaysia (above). Tree spacing is a challenge for the mobility of canopy-dwelling species, including for weaver ants (Oecophylla smaragdina), which bridge gaps by forming living chains of workers (below) (Both images are from Mark W. Moffett/Minden Pictures. All rights reserved)
-
6.
Physical and Chemical Properties of Structural Elements. In all canopies, the sessile hosts present associated species with a variety of surfaces, both between host individuals or species and within each host (such as wood versus leaves in a tree). These structural elements vary in their properties; examples include the capacity for insulation or water absorption, tendency to leach nutrients, efficiency at transmitting vibration, and their texture (Fig. 3.9), stability, density, hardness, compliance, stiffness, strength, pH, and so on. How do such variables affect life on or in a host? One of the oldest areas of canopy investigation in terrestrial biology is the question of substratum choice by epiphytes, especially cryptogams (e.g., Barkman 1958). Another area of intensive study has been herbivory as it relates to secondary compounds, nutrient content, and the mechanical difficulties of feeding (Schowalter 2011). Outside of these focal points, the literature is widely scattered, leaving many potential research avenues of enormous prospects.
Fig. 3.9 
The physical and structural properties of trees are important to canopy residents. Here a jararaca snake (Bothrops jararaca) is able to climb the trunk of an Araucaria tree in Brazil by gripping irregularities the rough bark (Mark W. Moffett/Minden Pictures. All rights reserved)




5 Getting to the Root of the Matter
Much of the language and thinking of forest canopy biology has been predicated on notions of plants as supports for the organisms residing in them (epiphytes, vines, and so on). Structural support between individuals occurs as well in the rhizosphere (Moffett 2001), reminding us that distinctions between above- and belowground plant parts can be arbitrary. In many ways, it would be logical to define words such as “epiphyte” so that they apply to the host in its entirety (e.g., a tree from root to crown: in marine biology, a more inclusive term, epibiont, describes any nonparasitic species living on any part of the surface of another organism; Wahl 1997). For many research purposes, this idea suggests the value of expanding beyond the realm of canopy biology to fashion a comprehensive science of sessile communities, from crowns to roots (or their equivalents). This nascent subject, which I refer to as structural ecology, would apply the principles of canopy biology to communities as a whole.
This would be no big leap for communities structured around sessile taxa lacking elaborate organs that penetrate the substrate, such as algae or corals. In such species, nutrient inputs tend to be greatest in the upper (outer) canopy rather than at or in the substratum, such that traits adaptive for light and nutrient procurement function almost entirely in synchrony (McCormick 1996). I focus instead on plants, with their extensive root systems. It is true that roots evolved from shoots prior to the evolution of leaves (Barlow 1994; Langdale 2008) and have remained developmentally distinct from leaves. In separating canopy from rhizosphere, however, it is more significant that roots and shoots are not necessarily distinguishable in either function or location relative to the ground surface: Shoots can absorb nutrients and water (Parker 1983; Schaefer and Reiners 1989) and can occur belowground, where they are referred to as rhizomes, and roots can occur aboveground, where they can be photosynthetic (Benzing 1991) or have ventilation and aeration functions, as in mangroves. The distinction made by botanists between belowground and aboveground (and often between root and shoot) has been largely methodological, a matter of choice between using a climbing rope or a shovel, for instance. The resulting academic fragmentation can be transcended, as in research on root stratification (or lack of clear strata) by several groups of researchers (Mommer et al. 2010; Frank et al. 2010; Kesanakurti et al. 2011; de Kroon et al. 2012). Of the published studies in recent years on the rhizosphere, efforts of this kind most closely follow the paradigm for canopy studies developed in this review.
The field of structural ecology will by necessity emerge in slow increments. While it is true that ecology took a long time to enter the treetops (Moffett and Lowman 1995), in many ways, it is not the aerial world but the subterranean one that remains most alien to us today, given the rhizosphere’s complexity (compare Beare et al. 1995 with Freiberg 1997), inaccessibility barring in most cases wholesale destructive intrusion (Smit et al. 2000), and difficulties in identifying plants from their roots (Mommer et al. 2011). Root systems and the species associated with their rhizosphere therefore could merit consideration equal to the rainforest canopy as the last—though assuredly not the highest (sensu Moffett 1994)—biotic frontier (André et al. 1994, 2002; Decaëns 2010). Compared with their crown architecture, the architecture of plant root systems in natural settings is poorly known (but see, e.g., Jeník 1978; Atger and Edelin 1993). This includes the links between the above- and belowground communities, which thus far have largely been framed in terms either of lone plants or of the above- versus belowground ecologies as gross-level compartments, rather than in terms of detailed architecture and stratification (Coleman et al. 1983; van der Putten et al. 2001; Moore et al. 2003; de Deyn and van der Putten 2005; Wardle et al. 2004; Poorter et al. 2012). Despite confounding effects from competition (e.g., Mahall and Callaway 1992; Burgess et al. 1998; Schenk 2006), root systems in nature are seldom isolated: Consider the drawings in Weaver and Clements (1929) of herb and grass communities (the distribution of tree roots is not known to this detail, but see Chilvers (1972), Lyford (1975), and the figures in Külla and Lõhmus (1999)) (Fig. 3.10). Still, 3-D studies of roots that extend beyond simple depth measurements of the kind reviewed by Jackson et al. (1996) and Schenk and Jackson (2002) are scarce (Tsegaye et al. 1995 a, b; Lynch et al. 1997; Ge et al. 2000; Pages 2000; Pages et al. 2000; Danjon et al. 2008; Fang et al. 2012), and most examples have been considered at the level of a single plant rather than of a community, although see the descriptions of the architecture and stratification of root systems by Caldwell and Richards (1986), Mommer et al. (2010), and Postma and Lynch (2012).

Rooting pattern of a group of Norway spruce trees, where root growth and distribution reflects competitive interactions (Gebauer and Martinková 2005) that are most intense near the soil surface, as they are for most plants (Photo by Roman Gebauer. All rights reserved. Reproduced with permission)
How would parasitology stand as a coherent discipline if its practitioners chose to look at the head of their subject while ignoring what its feet were doing? By amalgamating the findings from canopy biology with those from soil sciences, terrestrial biologists could fashion a comprehensive science of plant associates.
6 Conclusions Vis-à-vis Biodiversity
There are species from every kingdom of life that attach to or grow from substrata, and, when aggregated, these organisms form “a three-dimensional complex of structures,” as Spies (1998) described forests, whose parts that emerge from the substratum in aggregate can be (and often have been) described as canopies. Often, terrestrial studies appear to be categorized as “canopy biology” either on the basis of inaccessibility, as when specialized gear is required to access trees (Moffett and Lowman 1995), or on the basis of the communities’ cover-like properties. (In the latter case, canopy is treated as synonymous with overstory or used more broadly as any stratum in which sessile host organs are distributed so as to appreciably shade the layers below, as the word can be used in marine science; Dayton 1975a; Baird and Hughes 2000.) Because such criteria are arbitrary and of limited general import, I apply the word canopy to all above-substratum parts of sessile communities (of plants, when I originally described this idea in Moffett 2000). The common feature distinguishing studies as canopy biology is the treatment of sessile communities in three spatial dimensions, along with certain other attributes that are unique to life within this “canopy space.”
Why are the three-dimensional attributes of communities important enough to serve as the basis for a field of study? Taken in aggregate, the architecture of affixed life forms such as plants serves as scaffolding for much of the biological richness of this planet. By projecting into fluid media, canopies can augment productivity through increasing the biotic mass and the live surface area available for capturing and processing nutrients and energy and can enhance α-diversity through transforming in a heterogeneous way the climatic and chemical properties of the space they occupy, potentially multiplying the available niche space associated with a given surface area of the earth (e.g., Morin 2000; Moffett 2000; Hill and Hill 2001; Walla et al. 2004; DeVries et al. 2012). Moreover, canopies can provide retreats from predators, competitors, and adverse conditions, as well as additional surface area to attract or accumulate new species and the nutrients required to support a rich community (e.g., Crowder and Cooper 1982; Dean and Connell 1987; Lovett and Lindberg 1992; Jones et al. 1997; Johansen et al. 2008). We have yet to explain the differences and even more intriguing similarities in the physical structure and dynamics of canopies that develop in air as contrasted with water, and across orders of magnitude in host size: For example, compare Edred Corner’s views on trees with Timothy Allen’s on microalgae (Corner 1967; Allen 1977). Surely these size extremes are of special fascination; that may be the primary reason (beyond fulfilling our sense of adventure) that many of us climb trees to collect our data. Will scaling functions prove sufficiently linear to permit straightforward extrapolations across the full expanse of the different ecosystems?
Finally, canopy biology will have to be integrated into a more coherent understanding of sessile communities as a whole—that is, by taking similar account of the role of within-substratum organs such as roots and holdfasts. A central challenge of this holistic approach to communities, which I have called structural ecology, will be to determine the rules by which ecosystems, including both the sessile species and the residents of those species, assemble in all their three-dimensional glory, thereby creating opportunities for increased production and diversity—an idea that can be traced back to MacArthur and MacArthur (1961).
References
Adey WH, Loveland K (2007) Dynamic aquaria: building living ecosystems, 3rd edn. Academic, New York
Adey WH, Steneck RS (1985) Highly productive eastern Caribbean reefs: synergistic effects of biological, chemical, physical, and geological factors. In: Reaka ML (ed) The ecology of coral reefs, 3. Office of Undersea Research, Rockville
Aira M, McNamara NP, Piearce TG, Domínguez J (2009) Microbial communities of Lumbricus terrestris L. middens: structure, activity, and changes through time in relation to earthworm presence. J Soils Sediments 9:54–61
Allen TFH (1977) Scale in microscopic algal ecology: a neglected dimension. Phycologia 16:253–257
Allison DG, Gilbert P, Lappin-Scott HM, Wilson M (eds) (2000) Community structure and co–operation in biofilms. Cambridge University Press, Cambridge, UK
Al-Najjar MAA, de Beer D, Kühl M, Polerecky L (2012) Light utilization efficiency in photosynthetic microbial mats. Environ Microbiol 14:982–992
Aluja M, Prokopy RJ, Elkinton JS, Laurence WF (1989) Novel approach for tracking and quantifying the movement patterns of insects in three dimensions under seminatural conditions. Environ Entomol 18:1–7
Alvarez-Filip L, Dulvy NK, Gill JA, Côté IM, Watkinson AR (2009) Flattening of Caribbean coral reefs: region–wide declines in architectural complexity. Proc R Soc Lond B Biol 276:3019–3025
Alvarez-Filip L, Dulvy NK, Gill JA, Côté IM, Watkinson AR, Gill JA (2011) Coral identity underpins architectural complexity on Caribbean reefs. Ecol App. 21:2223–2231
Anderson TW (1994) Role of macroalgal structure in the distribution and abundance of a temperate reef fish. Mar Ecol Prog Ser 113:279–290
Anderson TW (2001) Predator responses, prey refuges, and density–dependent mortality of a marine fish. Ecology 82:245–257
André HM, Noti M-I, Lebrun P (1994) The soil fauna: the other last biotic frontier. Biodivers Conserv 3:45–56
André HM, Ducarme X, Lebrun P (2002) Soil biodiversity: myth, reality, or conning? Oikos 96:3–24
Andrews JH (2006) Population growth and the landscape ecology of microbes on leaf surfaces. In: Bailey MJ, Lilley AK, Timms-Wilson TM, Spencer-Phillips PTN (eds) Microbial ecology of aerial plant surfaces. CAB International, Wallingford
Anthony KR, Hoegh-Guldberg O (2003) Variation in coral photosynthesis, respiration and growth characteristics in contrasting light microhabitats: an analogue to plants in forest gaps and understoreys? Funct Ecol 17:246–259
Atger C, Edelin C (1993) Premières données sur l’architecture comparée des systèmes racinaires et caulinaires. Can J Bot 72:963–975
Aylor DE (1999) Biophysical scaling and the passive dispersal of fungus spores: relationship to integrated pest management strategies. Agric For Meteorol 97:275–292
Baird AH, Hughes TP (2000) Competitive dominance by tabular corals: an experimental analysis of recruitment and survival of understorey assemblages. J Exp Mar Biol Ecol 251:117–132
Barkman JJ (1958) Phytosociology and ecology of cryptogamic epiphytes. Van Gorcum, Assen
Barlow PW (1994) Rhythm, periodicity and polarity as bases for morphogenesis in plants. Biol Rev 69:475–525
Bates M (1960) The forest and the sea. Random House, New York
Bateson MC (1972) Our own metaphor. Knopf, New York
Battin TJ, Sloan WT, Kjelleberg S, Daims H, Head IM, Curtis TP, Eberl L (2007) Microbial landscapes: new paths to biofilm research. Nat Rev Microbiol 5:76–81
Beare MH, Coleman DC, Crossley DA Jr, Hendrix PF, Odum EP (1995) A hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling. Plant Soil 170:5–22
Bell SS, Coen LD (1982) Investigations on epibenthic meiofauna. I. Abundances on and repopulation of the tube–caps of Diopatra cuprea (Polychaeta: Onuphidae). Mar Biol 67:303–309
Benzing DH (1991) Aerial roots and their environments. In: Waisel Y, Eshel A, Kafkafi U (eds) Plant roots: the hidden half. Marcel Dekker, New York
Benzing DH (2012) Air plants: epiphytes and aerial gardens. Cornell University Press, Ithaca
Beugnon G, Fourcassié V (1988) How do red wood ants orient when foraging in a three dimensional system? II. Field experiments. Ins Soc 35:106–124
Beyenal H, Lewandowski Z (2002) Internal and external mass transfer in biofilms grown at various flow velocities. Biotechnol Prog 18:55–61
Bohr N (1955) Science and the unity of Knowledge. In: Leary L (ed) The unity of knowledge. Doubleday, Garden City
Borchers P, Field JG (1981) The effect of kelp shading on phytoplankton production. Bot Mar 24:89–91
Boston HL, Hill WR (1991) Photosynthesis–light relations of stream periphyton communities. Limnol Oceanogr 36:644–656
Boyle M, Janiak D, Craig S (2007) Succession in a Humboldt Bay marine fouling community: the role of exotic species, larval settlement and winter storms. In: Proceedings 2004 Humboldt Bay symposium, California Sea Grant, San Diego
Brady J, Gibson G, Packer MJ (1989) Odour movement, wind direction, and the problem of host–finding by tsetse flies. Physiol Entomol 14:369–380
Brigham RM, Grindal SD, Firman MC, Morissette JL (1997) The influence of structural clutter on activity patterns of insectivorous bats. Can J Zool 75:131–136
Brooks R, Owen-Smith N (1994) Plant defenses against mammalian herbivores: are juvenile Acacia more heavily defended than mature trees? Bothalia 24:211–215
Brown JH (1995) Macroecology. University of Chicago Press, Chicago
Bryers JD (2000) Biofilm formation and persistence. In: Bryers JD (ed) Biofilms II: process and applications. Wiley-Liss, New York
Burgess SSO, Adams MA, Turner NC, Ong CK (1998) The redistribution of soil water by tree root systems. Oecologia 115:306–311
Burkholder JM, Wetzel RG (1989) Epiphytic microalgae on a natural substratum in a phosphorus–limited hardwater lake: seasonal dynamics of community structure, biomass and ATP content. Arch Hydrobiol Suppl 83:1–56
Burrows CJ (1990) Processes of vegetation change. Unwin Hyman, London
Byrnes G, Jayne BC (2012) The effects of three-dimensional gap orientation on bridging performance and behavior of brown tree snakes (Boiga irregularis). J Exp Biol 215:2611–2620
Caldwell MM, Richards JM (1986) Competing root systems: morphology and models of absorption. In: Givnish TJ (ed) On the economy of plant form and function. Cambridge University Press, Cambridge, UK
Campbell GS, Norman JM (1989) The description and measurement of plant canopy structure. In: Russell G, Marshall B, Jarvis PG (eds) Plant canopies: their growth, form and function. Cambridge University Press, Cambridge, UK
Cannon CH, Leighton M (1994) Comparative locomotor ecology of gibbons and macaques: selection of canopy elements for crossing gaps. Am J Phys Anthropol 93:505–524
Cant JGH (1992) Positional behavior and body size of arboreal primates: a theoretical framework for field studies and an illustration of its application. Am J Phys Anthropol 88:273–283
Carpenter RC (1986) Partitioning herbivory and its effects on coral reef algal communities. Ecol Monogr 56:345–363
Carpenter RC, Williams SL (1993) Effects of algal turf canopy height and microscale substratum topography on profiles of flow speed in a coral forereef environment. Limnol Oceanogr 38:687–694
Carr MH (1989) Effects of macroalgal assemblages on the recruitment of temperate zone reef fishes. J Exp Mar Biol Ecol 126:59–76
Carr MH, Hixon MA (1997) Artificial reefs: the importance of comparisons with natural reefs. Fisheries 22(4):28–33
Carr GM, Morin A, Chambers PA (2005) Bacteria and algae in stream periphyton along a nutrient gradient. Freshwater Biol 50:1337–1350
Cheroske AG, Williams SL, Carpenter RC (2000) Effects of physical and biological disturbances on algal turfs in Kaneohe Bay, Hawaii. J Exp Mar Biol Ecol 248:1–34
Chiarucci A, Mistral M, Bonini I, Anderson BJ, Wilson JB (2002) Canopy occupancy: how much of the space in plant communities is filled? Folia Geobot 37:333–338
Chilvers GA (1972) Tree root pattern in a mixed eucalypt forest. Aust J Bot 20:229–234
Claflin TO (1968) Reservoir aufwuchs on inundated trees. Trans Am Microsc Soc 87:97–104
Coleman DC, Reid CPP, Cole CV (1983) Biological strategies of nutrient cycling in soil systems. Adv Ecol Res 13:1–55
Connell JH (1975) Some mechanisms producing structure in natural communities: a model and evidence from field experiments. In: Cody ML, Diamond JM (eds) Ecology and evolution of communities. Harvard University Press, Cambridge, MA
Corner EJH (1967) On thinking big. Phytomorphology 17:24–28
Costerton JW, Lappin-Scott HM (1995) Introduction to microbial biofilms. In: Lappin-Scott HM, Costerton JW (eds) Microbial biofilms. Cambridge University Press, Cambridge, UK
Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM (1995) Microbial biofilms. Ann Rev Microbiol 49:711–745
Costerton JW, Lewandowski Z, DeBeer D, Caldwell D, Korber D, James G (1994) Biofilms: the customized microniche. J Bacteriol 176:2137–2142
Coxson DS, Nadkarni NM (1995) Ecological roles of epiphytes in nutrient cycles of forest ecosystems. In: Lowman MD, Nadkarni NM (eds) Forest canopies. Academic, New York
Coxson DS, McIntyre DD, Vogel HJ (1992) Pulse release of sugars and polyols from canopy bryophytes in tropical montane rain forest (Guadeloupe, French West Indies). Biotropica 24:121–133
Crespi BJ (2001) The evolution of social behavior in microorganisms. Trends Ecol Evol 16:178–183
Crowder LB, Cooper WE (1982) Habitat structural complexity and the interaction between bluegills and their prey. Ecology 63:1802–1813
Cuthill I, Guilford T (1990) Perceived risk and obstacle avoidance in flying birds. Anim Behav 40:188–190
Dagosto M, Yamashita N (1998) Effect of habitat structure on positional behavior and support use in three species of lemur. Primates 39:459–472
Dahl AL (1973) Surface area in ecological analysis: quantification of benthic coral–reef algae. Mar Biol 23:239–249
Danjon F, Barker DH, Drexhage M, Stokes A (2008) Using three–dimensional plant root architecture in models of shallow–slope stability. Ann Bot 101:1281–1293
Darwin C (1839) Journal of researches into the geology and natural history of the various countries visited by H.M.S. Beagle. Colburn, London
Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP (1998) The involvement of cell–to–cell signals in the development of a bacterial biofilm. Science 280:295–298
Dawkins R (1982) The extended phenotype. Oxford University Press, Oxford, UK
Dawson EY (1966) Cacti in the Galapagos Islands with special reference to their relations with tortoises. In: Bowman RI (ed) The Galapagos, proceedings of the symposium of the California international scientific project, University of California Press, Los Angeles
Dayton PK (1971) Competition, disturbance, and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecol Monogr 41:351–389
Dayton PK (1975a) Experimental studies of algal canopy interactions in a sea otter–dominated kelp community at Amchitka Island, Alaska. Fish Bull 73:230–237
Dayton PK (1975b) Experimental evaluation of ecological dominance in a rocky intertidal algal community. Ecol Monogr 45:137–159
Dayton PK (1985) Ecology of kelp communities. Ann Rev Ecol Syst 16:215–245
Dayton PK, Tegner MJ, Edwards PB, Riser KL (1999) Temporal and spatial scales of kelp demography: the role of oceanographic climate. Ecol Mongr 69:219–250
De Deyn GB, Van der Putten WH (2005) Linking aboveground and belowground diversity. Trends Ecol Evol 20:625–633
de Kroon H, Hendriks M, van Ruijven J, Ravenek J, Padilla FM, Jongejans E, Visser EJW, Mommer L (2012) Root responses to nutrients and soil biota: drivers of species coexistence and ecosystem productivity. J Ecol 100:6–15
DeVries PJ, Alexander LG, Chacon IA, Fordyce JA (2012) Similarity and difference among rainforest fruit–feeding butterfly communities in Central and South America. J Anim Ecol 81:472–482
Dean RL, Connell JH (1987) Marine invertebrates in an algal succession. III. Mechanisms linking habitat complexity with diversity. J Exp Mar Biol Ecol 109:249–273
Dean TA, Thies K, Lagos SL (1989) Survival of juvenile giant kelp: the effects of demographic factors, competitors, and grazers. Ecology 70:483–495
Decaëns T (2010) Macroecological patterns in soil communities. Global Ecol Beogeogr 19:287–302
Dejean A, Corbara B, Orival J (1999) The arboreal ant mosaic in two Atlantic rain forests. Selbyana 20:133–145
Demes B, Jungers WL, Gross TS, Fleagle JG (1995) Kinetics of leaping primates: influence of substrate orientation and compliance. Am J Phys Anthropol 96:419–429
Dial R, Bloodworth B, Lee A, Boyne P, Heys J (2004) The distribution of free space and its relation to canopy composition at six forest sites. Forest Sci 50:312–325
Dieckmann U, Law R, Metz JAJ (eds) (2000) The geometry of ecological interactions: simplifying spatial complexity. Cambridge University Press, Cambridge, UK
Dodd ME, Silvertown J, Chase MW (1999) Phylogenetic analysis of trait evolution and species diversity variation among angiosperm families. Evolution 53:732–744
Dodds WK, Biggs BJF, Lowe RL (1999) Photosynthesis–irradiance patterns in benthic microalgae: variations as a function of assemblage thickness and community structure. J Phycol 35:42–53
Doemel WN, Brock TD (1977) Structure, growth, and decomposition of laminated algal–bacterial mats in alkaline hot springs. Appl Environ Microbiol 34:433–452
Erwin TL (1982) Tropical forests: their richness in Coleoptera and other arthropod species. Coleopt Bull 36:74–75
Fang S, Clark R, Liao H (2012) 3D quantification of plant root architecture in situ. In: Mancuso S (ed) Measuring roots. Springer, New York
Fierer N, Nemergut D, Knight R, Craine JM (2010) Changes through time: integrating microorganisms into the study of succession. Res Microbiol 161:635–642
Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8:623–633
Foottit RG, Adler PH (2009) Insect biodiversity: science and society. Wiley–Blackwell, Oxford, UK
Foster MS (1975) Regulation of algal community development in a Macrocystis pyrifera forest. Mar Biol 32:331–342
Foster K (2010) Social behaviour in microorganisms. In: Székely T, Komdeur J, Moore AJ (eds) Social behaviour: genes, ecology and evolution. Cambridge University Press, Cambridge, UK
Foster K, Bell T (2012) Competition, not cooperation, dominates interactions among culturable microbial species. Curr Biol 22:1845–1850
Fourcassie V, Beugnon G (1988) How do red wood ants orient when foraging in a three dimensional system? I. Laboratory experiments. Insect Soc 35:92–105
Franco M (1986) The influence of neighbours on the growth of modular organisms with an example from trees. Phil Trans R Soc Lond B 313:209–225
Frank DA, Pontes AW, Maine EM, Caruana J, Raina R, Raina S, Fridley JD (2010) Grassland root communities: species distributions and how they are linked to aboveground abundance. Ecology 91:3201–3209
Fraser SB, Sedberry GR (2008) Reef morphology and invertebrate distribution at continental shelf edge reefs in the south Atlantic bight. Southeast Nat 7:191–206
Freiberg M (1997) Spatial and temporal pattern of temperature and humidity of a tropical premontane rain forest tree in Costa Rica. Selbyana 18:77–84
Gaines SD, Roughgarden J (1987) Fish in offshore kelp forests affect recruitment to intertidal barnacle populations. Science 235:479–481
Garcia-Pichel F, Mechling M, Castenholz RW (1994) Diel migrations of microorganisms within a benthic, hypersaline mat community. Appl Environ Microbiol 60:1500–1511
Gauslaa Y (1995) The Lobarion, an epiphytic community of ancient forests threatened by acid rain. Lichenologist 27:59–76
Ge Z, Rubio G, Lynch JP (2000) The importance of root gravitropism for inter-root competition and phosphorus acquisition efficiency: results from a geometric simulation model. Plant Soil 218:159–171
Gebauer R, Martinková M (2005) Structure and functions of the types of Norway spruce (Picea abies [L.] Karst.) roots. J Forest Sci 51:305–311
Geiger R (1965) The climate near the ground. Harvard University Press, Cambridge, MA
Gerard VA (1984) The light environment in a giant kelp forest: influence of Macrocystis pyrifera on spatial and temporal variability. Mar Biol 84:189–194
Givnish TJ (1983) Convergent evolution of crown form in woody plants of southwestern Australia and New Caledonia. Am Phil Soc Yearb 1983:136
Givnish TJ (1984) Leaf and canopy adaptations in tropical forests. In: Medina E, Mooney HA, Vásquez-Yánes C (eds) Physiological ecology of plants of the wet tropics. Dr. Junk, The Hague
Givnish TJ (1995) Plant stems: biomechanical adaptation for energy capture and influence on species distributions. In: Gartner BL (ed) Plant stems: physiology and functional morphology. Chapman and Hall, New York
Graham NAJ, Nash KL (2012) The importance of structural complexity in coral reef ecosystems. Coral Reefs. doi:10.1007/s00338-012-0984-y
Graham NAJ, Wilson SK, Pratchett MS, Polunin NVC, Spalding MD (2009) Coral mortality versus structural collapse as drivers of corallivorous butterflyfish decline. Biodivers Conserv 18:3325–3336
Guerrero R, Mas J (1989) Multilayered microbial communities in aquatic ecosystems: growth and loss factors. In: Cohen Y, Rosenberg E (eds) Microbial mats: physiological ecology of benthic microbial communities. American Society for Microbiology, Washington, DC
Guerrero R, Piqueras M, Berlanga M (2002) Microbial mats and the search for minimal ecosystems. Int Microbiol 5:177–188
Haack TK, McFeters GA (1982) Nutritional relationships among microorganisms in an epilithic biofilm community. Microb Ecol 8:115–126
Hacker SD, Steneck RS (1990) Habitat architecture and the abundance and body-size-dependent habitat selection of a phytal amphipod. Ecology 71:2269–2285
Hackney JM, Carpenter RC, Adey WH (1989) Characteristic adaptations to grazing among algal turfs on a Caribbean coral reef. Phycologia 28:109–119
Haila Y (1990) Toward an ecological definition of an island: a northwest European perspective. J Biogeogr 17:561–568
Hallé F (1990) Tropical rain forests: structure and growth dynamics relative to utilization by birds. In: Keast A (ed) Biogeography and ecology of forest bird communities. SPB Academic, The Hague
Hallé F, Oldeman RAA, Tomlinson PB (1978) Tropical trees and forests: an architectural analysis. Springer, Berlin
Hallingbäck T (1990) Transplanting Lobaria pulmonaria to new localities and a review on the transplanting of lichens. Windahlia 18:57–64
Hamrick MW (1998) Functional and adaptive significance of primate pads and claws: evidence from New World anthropoids. Am J Phys Anthropol 106:113–127
Hansen SK, Rainey PB, Haagensen JA, Molin S (2007) Evolution of species interactions in a biofilm community. Nature 445:533–536
Hay ME (1986) Functional geometry of seaweeds: ecological consequences of thallus layering and shape in contrasting light environments. In: Givnish TJ (ed) On the economy of plant form and function. Cambridge University Press, Cambridge, UK
Heck KL Jr, Wetstone GS (1977) Habitat complexity and invertebrate species richness and abundance in tropical seagrass meadows. J Biogeogr 4:135–142
Herwitz SR, Slye RE (1992) Spatial variability in the interception of inclined rainfall by a tropical rainforest canopy. Selbyana 13:62–71
Hill WR (1996) Effects of light. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, New York
Hill JL, Hill RA (2001) Why are tropical rain forests so species rich? Classifying, reviewing and evaluating theories. Prog Phys Geogr 25:326–354
Hillebrand H, Frost P, Liess A (2008) Ecological stoichiometry of indirect grazer effects on periphyton nutrient content. Oecologia 155:619–630
Hirose T, Werger MJA (1995) Canopy structure and photon flux partitioning among species in a herbaceous plant community. Ecology 76:466–474
Hoagland KD, Roemer SC, Rosowski JR (1982) Colonization and community structure of two periphyton assemblages, with emphasis on the diatoms (Bacillariophyceae). Am J Bot 69:188–213
Hodoki Y (2005) Bacteria biofilm encourages algal immigration onto substrata in lotic systems. Hydrobiologia 539:27–34
Holbrook NM (1995) Stem water storage. In: Gartner BL (ed) Plant stems: physiology and functional morphology. Chapman and Hall, New York
Holbrook SJ, Denny MW, Koehl MAR (1991) Intertidal “trees”:consequences of aggregation on the mechanical and photosynthetic properties of sea-palms Postelsia palmaeformis Ruprecht. J Exp Mar Biol Ecol 146:39–67
Holbrook SJ, Brooks AJ, Schmitt RJ (2003) Variation in structural attributes of patch–forming corals and in patterns of abundance of associated fishes. Mar Freshw Res 53:1045–1053
Hölldobler B, Lumsden CJ (1980) Territorial strategies in ants. Science 210:732–739
Hölldobler B, Wilson EO (1977) Weaver ants: social establishment and maintenance of territory. Science 195:900–902
Hölldobler B, Wilson EO (1990) The ants. Harvard University Press, Cambridge, MA
Horn H (1971) The adaptive geometry of trees. Princeton University Press, Princeton
Hudon C, Bourget E (1983) The effect of light on the vertical structure of epibenthic diatom communities. Bot Mar 26:317–330
Hull DL (1988) Science as a process. University of Chicago Press, Chicago
Hurby T (1976) Observations of algal zonation resulting from competition. Estuar Coast Mar Sci 4:231–233
Hutchings MJ, De Kroon H (1994) Foraging in plants: the role of morphological plasticity in resource acquisition. Adv Ecol Res 25:159–238
Hyvärinen M, Halonen P, Kauppi M (1992) Influence of stand age and structure on the epiphytic lichen vegetation in the middle–boreal forests of Finland. Lichenologist 24:165–180
Ingram SW, Nadkarni NM (1993) Composition and distribution of epiphytic organic matter in a neotropical cloud forest, Costa Rica. Biotropica 25:370–383
Irie Y, Parsek MR (2008) Quorum sensing and microbial biofilms. Curr Top Microbiol Immunol 322:67–84
Irving AD, Connell SD (2006) Predicting understorey structure from the presence and composition of canopies: an assembly rule for marine algae. Oecologia 148:491–502
Jackson CR (2003) Changes in community properties during microbial succession. Oikos 101:444–448
Jackson JBC, Goreau TF, Hartman WD (1971) Recent brachiopod-coralline sponge communities and their paleoecological significance. Science 173:623–625
Jackson RB, Canadell J, Ehleringer JR, Mooney HA, Sala OE, Schulze ED (1996) A global analysis of root distributions for terrestrial biomes. Oecologia 108:389–411
Jander R (1990) Arboreal search in ants: search on branches (Hymenoptera: Formicidae). J Insect Behav 3:515–527
Jander R, Voss C (1963) Die Bedeutung von Streifenmustern fuer das Formensehen der Roten Waldameise (Formica rufa L.). Z Tierpsychol 20:1–9
Janzen DH (1968) Host plants as islands in evolutionary and contemporary time. Am Nat 102:592–595
Janzen DH (1973) Host plants as islands. II. Competition in evolutionary and contemporary time. Am Nat 107:786–790
Jeník J (1978) Roots and root systems in tropical trees. In: Tomlinson PB, Zimmerman MH (eds) Tropical trees as living systems. Cambridge University Press, Cambridge, UK
Jenkins B, Kitching RL (1990) The ecology of water-filled treeholes in Australian rainforests: food web reassembly as a measure of community recovery. Aust J Ecol 15:199–205
Johansen JL, Bellwood DR, Fulton CJ (2008) Coral reef fishes exploit flow refuges in high–flow habitats. Mar Ecol Prog Ser 360:219–226
Johnson RE, Tuchman NC, Peterson CG (1997) Changes in the vertical microdistribution of diatoms within a developing periphyton mat. J N Am Benthol Soc 16:503–519
Jones GP (1992) Interactions between herbivorous fishes and macro–algae on a temperate rocky reef. J Exp Mar Biol Ecol 159:217–235
Jones CG, Lawton JH, Shachak M (1997) Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946–1957
Jørgensen BB, Revsbech NP, Cohen Y (1983) Photosynthesis and structure of benthic microbial mats: microelectrode and SEM studies of four cyanobacterial communities. Limnol Oceanogr 28:1075–1093
Jørgensen BB, Revsbech NP, Blackburn TH, Cohen Y (1979) Diurnal cycle of oxygen and sulfide microgradients and microbial photosynthesis in a cyanobacterial mat sediment. Appl Environ Microbiol 38:46–58
Jørgensen BB, Cohen Y, Revsbech NP (1986) Transition from anoxygenic to oxygenic photosynthesis in a Microcoleus chthonoplastes cyanobacterial mat. Appl Environ Microbiol 51:408–417
Judd WS, Sanders RW, Donoghue MWJ (1994) Angiosperm family pairs: preliminary phylogenetic analyses. Harv Pap Bot 5:1–51
Karlson RH (1999) Dynamics of coral communities. Kluwer Academic, Dordrecht
Kastendiek J (1982) Competitor-mediated coexistence: interactions among three species of benthic macroalgae. J Exp Mar Biol Ecol 62:201–210
Kesanakurti PR, Fazekas AJ, Burgess KS, Percy DM, Newmaster SG, Graham SW, Barrett SCH, Hajibabaei M, Husband BC (2011) Spatial patterns of plant diversity below-ground as revealed by DNA barcoding. Mol Ecol 20:1289–1302
Kierek-Pearson K, Karatan E (2005) Biofilm development in bacteria. Adv Appl Microbiol 57:79–111
Kikuchi T, Peres JM (1977) Consumer ecology of seagrass beds. In: McRoy CP, Helfferich C (eds) Seagrass ecosystems. Marcel Dekker, New York
Kim S, Soltis DE, Soltis PS, Zanis MJ, Suh Y (2004) Phylogenetic relationships among early-diverging eudicots based on four genes: were the eudicots ancestrally woody? Mol Phylogenet Evol 31:16–30
Kitching JA, Macan TT, Gilson HC (1934) Studies in sublittoral ecology. I. A submarine gully in Wembury Bay, South Devon. J Mar Biol Assoc UK 19:677–705
Klausen M, Aaes-Jorgensen A, Molin S, Tolker-Nielsen T (2003) Involvement of bacterial migration in the development of complex multicellular structures in Pseudomonas aeruginosa biofilms. Mol Microbiol 50:61–68
Klotz JH, Reid BL (1992) The use of spatial cues for structural guideline orientation in Tapinoma sessile and Camponotus pennsylvanicus. J Insect Behav 5:71–82
Korber DR, Lawrence JR, Lappin-Scott HM, Costerton JW (1995) Growth of microorganisms on surfaces. In: Lappin-Scott HM, Costerton JW (eds) Microbial biofilms. Cambridge University Press, Cambridge, UK
Korte VL, Blinn DW (1983) Diatom colonization on artificial substrata in pool and riffle zones studied by light and scanning electron microscopy. J Phycol 19:332–341
Kricher J (1997) A neotropical companion. Princeton University Press, Princeton
Kuenen JG, Jørgensen BB, Revsbech NP (1986) Oxygen microprofiles of trickling filter biofilms. Water Res 20:1589–1598
Kühl M, Polerecky L (2008) Functional and structural imaging of phototrophic microbial communities and symbioses. Aquat Microb Ecol 53:99–118
Kühl M, Fenchel T (2000) Bio-optical characteristics and the vertical distribution of photosynthetic pigments and photosynthesis in an artificial cyanobacterial mat. Microb Ecol 40:94–103
Kühl M, Glud RN, Ploug H, Ramsing NB (1996) Microenvironmental control of photosynthesis and photosynthesis-coupled respiration in an epilithic cyanobacterial biofilm. J Phycol 32:799–812
Kühl M, Fenchel T, Kazmierczak J (2003) Growth, structure and calcification potential of an artificial cyanobacterial mat. In: Krumbein WE, Paterson D, Zavarzin G (eds) Fossil and recent biofilms, a natural history of life on Earth. Kluwer Academic, Dordrecht
Külla T, Lõhmus K (1999) Influence of cultivation method on root grafting in Norway spruce (Picea abies (L.) Karst). Plant Soil 217:91–100
Lakoff G (1987) Women, fire, and dangerous things: what categories reveal about the mind. University of Chicago Press, Chicago
Langdale JA (2008) Evolution of developmental mechanisms in plants. Cur Opin Genet Develop 18:368–373
Lassen C, Ploug H, Kühl M, Jørgensen BB (1994) Oxygenic photosynthesis and light distribution in marine microbial mats. In: Stal LJ, Caumette P (eds) Microbial mats: structure, development, and environmental significance. Springer, Berlin
Lawrence JR, Korber DR, Hoyle BD, Costerton JW, Caldwell DE (1991) Optical sectioning of microbial biofilms. J Bacteriol 173:6558–6567
Lawrence JR, Korber DR, Wolfaardt GM, Caldwell DE (1995) Behavioral strategies of surface–colonizing bacteria. Adv Microb Ecol 14:1–75
Lawrence JR, Swerhone GDW, Kuhlicke U, Neu TR (2007) In situ evidence for microdomains in the polymer matrix of bacterial microcolonies. Can J Microbiol 53:450–458
Lawton, JH (1983) Plant architecture and the diversity of phytophagous insects. Ann Rev Entomol 28:23–29
Leigh EG Jr (1999) Tropical forest ecology: a view from Barro Colorado Island. Oxford University Press, Oxford, UK
Levin PS, Hay ME (1996) Responses of temperate reef fishes to alterations in algal structure and species composition. Mar Ecol Prog Ser 134:37–47
Lieberman M, Lieberman D, Peralta R (1989) Forests are not just swiss cheese: canopy stereogeometry of non-gaps in tropical forests. Ecology 70:550–552
Liira J, Zobel K, Mägi R, Molenberghs G (2002) Vertical structure of herbaceous canopies: the importance of plant growth–form and species–specific traits. Plant Ecol 163:123–134
Lindo Z, Whiteley JA (2011) Old trees contribute bio–available nitrogen through canopy bryophytes. Plant Soil 342:141–148
Losos JB (2009) Lizards in an evolutionary tree: ecology and adaptive radiation of anoles. University of California Press, Berkeley
Lovett GM, Lindberg SE (1992) Concentration and deposition of particles and vapors in a vertical profile through a forest canopy. Atmos Environ 26A:1469–1476
Lowe RL, Guckert JB, Belanger SE, Davidson DH, Johnson DW (1996) An evaluation of periphyton community structure and function on tile and cobble substrata in experimental stream mesocosms. Hydrobiologia 328:135–146
Lowe RL, Shavit U, Falter JL, Koseff JR, Monismith SG (2008) Modeling flow in coral communities with and without waves: a synthesis of porous media and canopy flow approaches. Limnol Oceanogr 53:2668–2680
Lyford WH (1975) Rhizography of non-woody roots of trees in the forest floor. In: Torrey JG, Clarkson DT (eds) The development and function of roots. Academic, New York
Lynch JP, Nielsen KL, Davis RD, Jablokow AG (1997) SimRoot: modelling and visualization of root systems. Plant Soil 188:139–151
MacArthur R, MacArthur JW (1961) On bird species diversity. Ecology 42:594–598
MacArthur R, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
MacIntyre HL, Geider RJ, Miller DC (1996) Microphytobenthos: the ecological role of the “secret garden” of unvegetated, shallow–water marine habitats. I. Distribution, abundance and primary production. Estuaries 19:186–201
Madsen JS, Burmølle M, Hansen LH, Sørensen SJ (2012) The interconnection between biofilm formation and horizontal gene transfer. FEMS Immu Med Microbiol 65:183–195
Mahall BE, Callaway RM (1992) Root communication mechanisms and intracommunity distributions of two mojave desert shrubs. Ecology 73:2145–2151
Maida M, Coll JC, Sammarco PW (1994) Shedding new light on scleractinian coral recruitment. J Exp Mar Biol Ecol 180:189–202
Maraun M, Alphei J, Bonkowski M, Buryn R, Migge S, Peter M, Schaefer M, Scheu S (1999) Middens of the earthworm Lumbricus terrestris (Lumbricidae): microhabitats for micro- and mesofauna in forest soil. Pedobiologia 43:276–286
Marks JC, Power ME (2001) Nutrient induced changes in the species composition of epiphytes on Cladophora glomerata Kütz. Hydrobiologia 450:187–196
Massol-Deyá AA, Whallon J, Hickey RF, Tiedje JM (1995) Channel structures in aerobic biofilms of fixed–film reactors treating contaminated groundwater. Appl Environ Microbiol 61:769–777
McCormick PV (1996) Resource competition and species coexistence in freshwater benthic algal assemblages. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic Press, New York
McCormick PV, Stevenson RJ (1991) Grazer control of nutrient availability in the periphyton. Oecologia 86:287–291
McCune B (1993) Gradients in epiphyte biomass in three Pseudotsuga–Tsuga forests of different ages in western Oregon and Washington. Bryologist 96:405–411
McCune B, Antos JA (1981) Diversity relationships of forest layers in the Swan Valley, Montana. Bull Torrey Bot Club 108:354–361
McElhinny C, Gibbons P, Brack C, Bauhus J (2005) Forest and woodland stand structural complexity: its definition and measurement. Forest Ecol Manag 218:1–24
McGraw WS (1998) Posture and support use of Old World monkeys (Cercopithecidae):the influence of foraging strategies, activity patterns, and the spatial distribution of preferred food items. Am J Primatol 46:229–250
Meulemans JT, Roos PJ (1985) Structure and architecture of the periphytic community on dead reed stems in Lake Maarsseveen. Arch Hydrobiol 102:487–502
Meyers TP, Huebert BJ, Hicks BB (1989) HNO3 deposition to a deciduous forest. Boundary-Layer Meteorol 49:395–410
Miller AR, Lowe RL, Rotenberry JT (1987) Succession of diatom communities on sand grains. J Ecol 75:693–709
Miller RJ, Reed DC, Brzezinski MA (2011) Partitioning of primary production among giant kelp (Macrocystis pyrifera), understory macroalgae, and phytoplankton on a temperate reef. Limnol Oceanogr 56:119–132
Mitri S, Xavier JB, Foster KR (2011) Social evolution in multispecies biofilms. Proc Nat Acad Sci 108:10839–10846
Moe B, Botnen A (1997) A quantitative study of the epiphytic vegetation on pollarded trunks of Fraxinus excelsior at Havrå, Osterøy, western Norway. Plant Ecol 129:157–177
Moffett MW (1994) The high frontier: exploring the tropical rainforest canopy. Harvard University Press, Cambridge, MA
Moffett MW (1999) Life on vegetation: a framework for canopy biology. In: Hallé F (ed) Biologie d’une canopée de forêt équatoriale –– IV. Pro–Natura and Opération Canopée, Montpellier, France
Moffett MW (2000) What’s “up?” A critical look at the basic terms of canopy biology. Biotropica 32:569–596
Moffett MW (2001) The nature and limits of canopy biology. Selbyana 22:155–179
Moffett MW (2002) The highs and lows of tropical forest canopies. J Biogeogr 29:1264–1265
Moffett MW (2010) Adventures among ants. University of California Press, Berkeley
Moffett MW, Lowman MD (1995) Canopy access techniques. In: Lowman MD, Nadkarni NM (eds) Forest canopies. Academic, San Diego
Molin S, Tolker-Nielsen T (2003) Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr Opin Biotechnol 14:255–261
Molin S, Nielsen AT, Christensen BB, Andersen JB, Licht TR, Tolker-Nielsen T, Sternberg C, Hansen MC, Ramos C, Givskov M (2000) Molecular ecology of biofilms. In: Bryers JD (ed) Biofilms II: process analysis and applications. Wiley-Liss, New York
Mommer L, van Ruijven J, de Caluwe H, Smit-Tiekstra AE, Wagemaker CAM, Ouborg NJ, Bogemann GM, van der Weerden GM, Berendse F, de Kroon H (2010) Unveiling below–ground species abundance in a biodiversity experiment: a test of vertical niche differentiation among grassland species. J Ecol 98:1117–1127
Mommer L, Dumbrell AJ, Wagemaker CAM, Ouborg NJ (2011) Belowground DNA–based techniques: untangling the network of plant root interactions. Plant Soil 348:115–121
Monteith JL (1975/1976) Vegetation and the atmosphere, vol 1 and 2. Academic, London
Montgomery RA, Chazdon RL (2001) Forest structure, canopy architecture, and light transmittance in tropical wet forests. Ecology 82:2707–2718
Moore JC, McCann K, Setälä H, de Ruiter PC (2003) Top-down is bottom-up: does predation in the rhizosphere regulate aboveground dynamics? Ecology 84:846–857
Morin PJ (2000) Biodiversity’s ups and downs. Nature 406:463–464
Morris CE, Monier JM, Jacques MA (1997) Methods for observing microbial biofilms directly on leaf surfaces and recovering them for isolation of culturable microorganisms. Appl Environ Microbiol 63:1570–1576
Mulholland PJ (1996) Role in nutrient cycling in streams. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, New York
Mundy CN, Babcock RC (1998) Role of light intensity and spectral quality in coral settlement: implications for depth-dependent settlement? J Exp Mar Biol Ecol 223:235–255
Nadell CD, Xavier JB, Levin SA, Foster KR (2008) The evolution of quorum sensing in bacterial biofilms. PLoS Biol 6:e14
Neushul M (1971) Submarine illumination in Macrocystis beds. Beihefte zur Nova Hedwigia 32:241–254
Neushul M (1972) Functional interpretation of benthic marine algal morphology. In: Abbott IA, Kurogi M (eds) Contributions to the systematics of benthic marine algae of the North Pacific. Japanese Society of Phycology, Kobe
Niklas KJ (1997) Evolutionary biology of plants. University of Chicago Press, Chicago
Niklas KJ (2000) The mechanical stability of vertical stems. In: Kurmann MH, Hemsley AR (eds) The evolution of plant architecture. Royal Botanical Gardens, Kew
Niklas KJ, Kerchner V (1984) Mechanical and photosynthetic constraints on the evolution of plant shape. Paleobiology 10:79–101
O’ Connell T, Bolger T (1997) Fungal fruiting bodies and the structure of fungus-micro-arthropod assemblages. Proc R Irish Acad Sect B 97:249–262
Paerl HW, Pinckney JL, Steppe TF (2000) Cyanobacterial–bacterial mat consortia: examining the functional unit of microbial survival and growth in extreme environments. Environ Microbiol 2:11–26
Pagès L (2000) How to include organ interactions in models of the root system architecture? The concept of endogenous environment. Ann For Sci 57:535–541
Pagès L, Doussan C, Vercambre G (2000) An introduction on below–ground environment and resource acquisition, with special reference on trees: simulation models should include plant structure and function. Ann For Sci 57:513–520
Paine RT, Suchanek TH (1983) Convergence of ecological processes between independently evolved competitive dominants: a tunicate–mussel comparison. Evolution 37:821–831
Parker GG (1983) Throughfall and stemflow in the forest nutrient cycle. Adv Ecol Res 13:57–133
Parker GG, Brown MJ (2000) Forest canopy stratification –– is it useful? Am Nat 155:473–484
Passy SI, Larson CA (2011) Succession in stream biofilms is an environmentally driven gradient of stress tolerance. Microb Ecol 62:414–424
Pearce D, Bazin MJ, Lynch JM (1995) The rhizosphere as a biofilm. In: Lappin-Scott HM, Costerton JW (eds) Microbial biofilms. Cambridge University Press, Cambridge, UK
Pearse JS, Hines AH (1979) Expansion of a central California kelp forest following the mass mortality of sea urchins. Mar Biol 51:83–91
Perry DR (1978) Factors influencing arboreal epiphytic phytosociology in Central America. Biotropica 10:235–237
Peterson CG (1996) Response of benthic algal communities to natural physical disturbance. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, San Diego
Peterson CG, Grimm NB (1992) Temporal variation in enrichment effects during periphyton succession in a nitrogen–limited desert stream ecosystem. J N Am Benthol Soc 11:20–36
Pike LH (1978) The importance of epiphytic lichens in mineral cycling. Bryologist 81:247–257
Pogoreutz C, Kneer D, Litaay M, Asmus H, Ahnelt H (2012) The influence of canopy structure and tidal level on fish assemblages in tropical Southeast Asian seagrass meadows. Estuar Coast Shelf Sci 107:58–68
Poorter L, Hawthorne W, Bongers F, Sheil D (2008) Maximum size distributions in tropical forest communities: relationships with rainfall and disturbance. J Ecol 96:495–504
Poorter L, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2012) Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol 193:30–50
Porter JW (1974) Zooplankton feeding by the Caribbean reef-building coral Montastrea cavernosa. Proc Second Int Coral Reef Symp 1:111–125
Porter JW (1976) Autotrophy, heterotrophy, and resource partitioning in Caribbean reef-building corals. Am Nat 110:731–742
Postma JA, Lynch JP (2012) Complementarity in root architecture for nutrient uptake in ancient maize/bean and maize/bean/squash polycultures. Ann Bot 110:521–534
Povinelli DJ, Cant JGH (1995) Arboreal clambering and the evolution of self–conception. Quart Rev Biol 70:393–421
Pringle CM (1985) Effects of chironomid (Insecta: Diptera) tube-dwelling activities on stream diatom communities. J Phycol 21:185–194
Putz FE (1995) Vines in treetops: consequences of mechanical dependence. In: Lowman MD, Nadkarni NM (eds) Forest canopies. Academic, San Diego
Ramage G, Mowat E, Jones B, Williams C, Lopez-Ribot J (2009) Our current understanding of fungal biofilms. Crit Rev Microbiol 35:340–355
Ramsing NB, Kühl M, Jørgensen BB (1993) Distribution of sulfate–reducing bacteria, O2, and H2S in photosynthetic biofilms determined by oligonucleotide probes and microelectrodes. Appl Environ Microbiol 59:3840–3849
Ramsing NB, Ferris MJ, Ward DM (2000) Highly ordered vertical structure of Synechococcus populations within the one-millimeter-thick photic zone of a hot spring cyanobacterial mat. Appl Environ Microbiol 66:1038–1049
Randlkofer B, Obermaier E, Hilker M, Meiners T (2010) Vegetation complexity: the influence of plant species diversity and plant structures on plant chemical complexity and arthropods. Basic Appl Ecol 11:383–395
Rasmussen L (1975) The bryophytic epiphyte vegetation in the forest, Slotved Skov, Northern Jutland. Lindbergia 3:15–38
Reed DC, Foster MS (1984) The effects of canopy shadings on algal recruitment and growth in a giant kelp forest. Ecology 65:937–948
Reichenbach H (1951) The rise of scientific philosophy. University of California Press, Berkeley
Reiners WA, Olson RK (1984) Effects of canopy components on throughfall chemistry: an experimental analysis. Oecologia 63:320–330
Reynolds TB, Fink GR (2001) Baker’s yeast, a model for fungal biofilm formation. Science 291:878–881
Richards AJ (1986) Plant breeding systems. George Allen and Unwin, London
Richards PW (1996) The tropical rain forest, an ecological study, 2nd edn. Cambridge University Press, Cambridge, UK
Richardson BA (1999) The bromeliad microcosm and the assessment of faunal diversity in a neotropical forest. Biotropica 31:321–336
Robinson CT, Minshall GW (1986) Effects of disturbance frequency on stream benthic community structure in relation to canopy cover and season. J N Am Benthol Soc 5:237–248
Roemer SC, Hoagland KD, Rosowski JR (1984) Development of a freshwater periphyton community as influenced by diatom mucilages. Can J Bot 62:1799–1813
Roeselers G, van Loosdrecht MCM, Muyzer G (2007) Heterotrophic pioneers facilitate phototrophic biofilm development. Microbial Ecol 54:578–585
Roos PJ (1979) Architecture and development of periphyton on reed-stems in Lake Maarsseveen. Hydrobiol Bull 13:117
Roxburgh SH, Watkins AJ, Wilson JB (1993) Lawns have vertical stratification. J Veg Sci 4:699–704
Ruinen J (1961) The phyllosphere. I. An ecologically neglected milieu. Plant Soil 15:81–109
Ruinen J (1975) Nitrogen fixation in the phyllosphere. In: Stewart WDP (ed) Nitrogen fixation by free–living micro–organisms. Cambridge University Press, Cambridge, UK
Rupp CJ, Fux CA, Stoodley P (2005) Viscoelasticity of Staphylococcus aureus biofilms in response to fluid shear allows resistance to detachment and facilitates rolling migration. Appl Environ Microbiol 71:2175–2178
Russell BC (1977) Population and standing crop estimates for rocky reef fishes of North–Eastern New Zealand. N Z J Mar Freshw Res 11:23–36
Russell BD (2007) Effects of canopy-mediated abrasion and water flow on the early colonisation of turf-forming algae. Mar Freshw Res 58:657–665
Russell G, Marshall B, Jarvis PG (eds) (1989) Plant canopies: their growth, form and function. Cambridge University Press, Cambridge, UK
Sagan D, Margulis L (1988) Garden of microbial delights. Harcourt Brace Jovanovich, Boston
Santelices B, Ojeda FP (1984) Effects of canopy removal on the understory algal community structure of coastal forests of Macrocystis pyrifera from southern South America. Mar Ecol Prog Ser 14:165–173
Schaefer DA, Reiners WA (1989) Throughfall chemistry and canopy processing mechanisms. In: Lindberg SE, Page AL, Norton SA (eds) Acidic precipitation. Vol. 3: sources, deposition, and canopy interactions. Springer, New York
Schaudinn C, Stoodley P, Kainovic A, O’Keeffe T, Costerton B, Robinson D, Baum M, Ehrlich G, Webster P (2007) Bacterial biofilms, other structures seen as mainstream concepts. Microbe 2:231–237
Schenk HJ (2006) Root competition: beyond resource depletion. J Ecol 94:725–739
Schenk HJ, Jackson RB (2002) The global biogeography of roots. Ecol Monog 72:311–328
Schmitt RJ, Holbrook SJ (1990) Contrasting effects of giant kelp on dynamics of surfperch populations. Oecologia 84:419–429
Schowalter TD (2011) Insect ecology: an ecosystem approach, 3rd edn. Academic, New York
Scott JT, Back JA, Taylor JM, King RS (2008) Does nutrient enrichment decouple algal–bacterial production in periphyton? J N Am Benthol Soc 27:332–344
Seifert R (1975) Clumps of Heliconia inflorescences as ecological islands. Ecology 56:1416–1422
Sekar R, Venugopalan VP, Nandakumar K, Nair KVK, Rao VNR (2004) Early stages of biofilm succession in a lentic freshwater environment. Hydrobiologia 512:97–108
Shaw DC (1996) Northwest forest canopies: preface. NW Sci 70:i–ii
Shugart HH, Saatchi S, Hall FG (2010) Importance of structure and its measurement in quantifying function of forest ecosystems. J Geophys Res 115:G00E13
Sillett SC, Gradstein SR, Griffin D (1995) Bryophyte diversity of Ficus tree crowns from cloud forest and pasture in Costa Rica. Bryologist 98:251–260
Sinoquet H, Pincebourde S, Adam B, Donès N, Phattaralerphong J, Combes D, Ploquin S, Sangsing K, Kasemsap P, Thanisawanyangkura S, Groussier-Bout G, Casas J (2009) 3-D maps of tree canopy geometries at leaf scale. Ecology 90:283
Sládecková A (1962) Limnological investigation methods for the periphyton (“aufwuchs”) community. Bot Rev 28:286–350
Smit AL, Bengough AG, Engels C, van Noordwijk M, Pellerin S, van de Geijn SC (eds) (2000) Root methods: a handbook. Springer, Berlin
Sobczak WV (1996) Epilithic bacterial responses to variations in algal biomass and labile dissolved organic carbon during biofilm colonization. J N Am Benthol Soc 15:143–154
Sousa WP (1979) Experimental investigations of disturbance and ecological succession in a rocky intertidal algal community. Ecol Monogr 49:227–254
Spies TA (1998) Forest structure: a key to the ecosystem. N W Sci 72:34–39
Staal M, Borisov S, Rickelt LF, Klimant I, Kühl M (2011) Ultrabright planar optodes for luminescence life-time based microscopic imaging of O2 dynamics in biofilms. J Microbiol Methods 85:67–74
Stal LJ (2000) Cyanobacterial mats and stromatolites. In: Whitton BA, Potts M (eds) Ecology of cyanobacteria. Kluwer Academic, Dordrecht
Steele MA (1999) Effects of shelter and predators on reef fishes. J Exp Mar Biol Ecol 233:65–79
Steneck RS (1997) Crustose corallines, other algal functional groups, herbivores and sediments: complex interactions along reef productivity gradients. In: Proceedings of 8th international coral reef symposium, Panama City 1:695–700
Steneck RS, Dethier MN (1994) A functional group approach to the structure of algal–dominated communities. Oikos 69:476–498
Sterck FJ, Schieving F (2007) 3-D growth patterns of trees: effects of carbon economy, meristem activity, and selection. Ecol Monog 77:405–420
Sterck FJ, Bongers F, During HJ, Martínez-Ramos M, de Kroon H (2003) Module responses in a tropical forest tree analyzed with a matrix model. Ecology 84:2751–2761
Stevenson RJ (1983) Effects of current and conditions simulating autogenically changing microhabitats on benthic diatom immigration. Ecology 64:1514–1524
Stevenson RJ (1996) An introduction to algal ecology in freshwater benthic habitats. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, New York
Stewart PS, Franklin MJ (2008) Physiological heterogeneity in biofilms. Nat Rev Microbiol 6:199–2010
Stimson J (1985) The effect of shading by the table coral Acropora hyacinthus on understory corals. Ecology 66:40–53
Stock MS, Ward AK (1991) Blue–green algal mats in a small stream. J Phycol 27:692–698
Stolzenbach KD (1989) Particle transport and attachment. In: Characklis WG, Wilderer PA (eds) Structure and function of biofilms. Wiley, New York
Stoodley P, Boyle JD, DeBeer D, Lappin-Scott HM (1999a) Evolving perspectives of biofilm structure. Biofouling 14:75–90
Stoodley P, Lewandowski Z, Boyle JD, Lappin-Scott HM (1999b) The formation of migratory ripples in a mixed species bacterial biofilm growing in turbulent flow. Environ Microb 1:447–455
Stork NE, Adis J, Didham RK (1997) Canopy arthropods. Chapman and Hall, London
Svenning J-C (2000) Small canopy gaps influence plant distributions in the rain forest understory. Biotropica 32:252–261
Teske A, Stahl DA (2001) Microbial mats and biofilms: evolution, structure and function of fixed microbial communities. In: Staley JT, Reysenbach A-L (eds) Biodiversity of microbial life: foundation of earth’s biosphere. Wiley, New York
Tilman D (1982) Resource competition and community structure. Princeton University Press, Princeton
Tomlinson PB (1983) Tree architecture. Am Sci 71:141–149
Tsegaye T, Mullins CE, Diggle AJ (1995a) An experimental procedure for obtaining input parameters for the rootmap root simulation program for peas (Pisum sativum L). Plant Soil 172:1–16
Tsegaye T, Mullins CE, Diggle AJ (1995b) Modeling pea (Pisum sativum) root growth in drying soil: a comparison between observations and model predictions. New Phytol 131:179–189
Tuchman NC (1996) The role of heterotrophy in algae. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, New York
Tuchman NC, Stevenson RJ (1991) Effects of selective grazing by snails on benthic algal succession. J N Am Benthol Soc 10:430–443
Turner JS (2000) The extended organism: the physiology of animal-built structures. Harvard University Press, Cambridge, MA
Vadas RL, Steneck RS (1988) Zonation of deep water benthic algae in the Gulf of Maine. J Phycol 24:338–346
Van der Putten WH, Vet LEM, Harvey JA, Wackers FL (2001) Linking above- and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol Evol 16:547–554
Van Gemerden H (1993) Microbial mats, a joint venture. Mar Geol 113:3–25
Venugopalan VP, Kuehn M, Hausner M, Springael D, Wilderer PA, Wuertz S (2005) Architecture of a nascent Sphingomonas sp. biofilm under varied hydrodynamic conditions. Appl Environ Microbiol 71:2677–2686
Vogel S (1996) Blowing in the wind: storm-resisting features of the design of trees. J Arboriculture 22:92–98
Vytopil E, Willis B (2001) Epifaunal community structure in Acropora spp. (Scleractinia) on the Great Barrier Reef: implications of coral morphology and habitat complexity. Coral Reefs 20:281–288
Wahl M (1997) Living attached: aufwuchs, fouling, epibiosis. In: Nagabhushanam R, Thompson M-F (eds) Fouling organisms of the Indian Ocean. A.A. Balkema, Rotterdam
Walla TR, Engen S, DeVries PJ, Lande R (2004) Modeling vertical beta–diversity in tropical butterfly communities. Oikos 107:610–618
Walter H (1973) Vegetation on earth. Springer, New York
Ward DM, Bateson MM, Ferris MJ, Kühl M, Wieland A, Koeppel A, Cohan FM (2006) Cyanobacterial ecotypes in the microbial mat community of Mushroom Spring (Yellowstone National Park, Wyoming) as species–like units linking microbial community composition, structure and function. Philos Trans R Soc B 361:1997–2008
Wardle DA, Bardgett RD, Klironomos JN, Setälä H, van der Putten WH, Wall DH (2004) Ecological linkages between aboveground and belowground biota. Science 304:1629–1633
Watnick P, Kolter R (2000) Biofilm, city of microbes. J Bacteriol 182:2675–2679
Weaver JE, Clements FE (1929) Plant ecology. McGraw-Hill, New York
Wernberg T, Kendrick GA, Toohey BD (2005) Modification of the physical environment by an Ecklonia radiata (Laminariales) canopy and implications for associated foliose algae. Aquat Ecol 39:419–430
Weitzel RG (1979) Periphyton measurements and applications. In: Weitzel RG (ed) Methods and measurements of periphyton communities: a review. American Society for Testing and Materials, Philadelphia
Wetzel RG (1983) Opening remarks. In: Wetzel RG (ed) Periphyton of freshwater ecosystems. Dr. W. Junk, The Hague
Wetzel RG (1993) Microcommunities and microgradients: linking nutrient regeneration, microbial mutualism, and high sustained aquatic primary production. Neth J Aquat Ecol 27:3–9
Whipps JM, Hand P, Pink D, Bendin GD (2008) Phyllosphere microbiology with special reference to diversity and plant genotype. J Appl Microbiol 105:1744–1755
Williams SL, Carpenter RC (1990) Photosynthesis/photon flux density relationships among components of coral reef algal turfs. J Phycol 26:36–40
Williams SL, Heck KL Jr (2001) Seagrass Community ecology. In: Bertness M, Gaines S, Hay M (eds) Marine community ecology. Sinauer Press, New York
Williams GA, Seed R (1992) Interactions between macrofaunal epiphytes and their host algae. In: John DM, Hawkins SJ, Price JH (eds) Plant–animal interactions in the marine benthos. Clarendon, Oxford, UK
Wilson JB, Steel JB, Steel SLK (2007a) Do plants ever compete for space? Folia Geobotanica 42:431–436
Wilson SK, Graham NAJ, Polunin NVC (2007b) Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Mar Biol 151:1069–1076
Wiman BLB, Agren GI, Lannefors HO (1985) Aerosol concentration profiles within a mature coniferous forest–model versus field results. Atmos Environ 19:363–367
Wimpenny JWT, Colasanti R (1997) A unifying hypothesis for the structure of microbial biofilms based on cellular automaton models. FEMS Microbiol Ecol 22:1–16
Wimpenny JWT, Kinniment SL (1995) Biochemical reactions and the establishment of gradients within biofilms. In: Lappin-Scott HM, Costerton JW (eds) Microbial biofilms. Cambridge University Press, Cambridge, UK
Wing SR, Leichter JJ, Denny MW (1993) A dynamic model for wave–induced light fluctuations in a kelp forest. Limnol Oceanogr 38:396–407
Wohlgemuth S, Ronacher B, Wehner R (2001) Ant odometry in the third dimension. Nature 411:795–798
Yeh P-Z, Gibor A (1970) Growth patterns and motility of Spirogyra sp. and Closterium acerosum. J Phycol 6:44–48
Young JW (2009) Substrate determines asymmetrical gait dynamics in marmosets (Callithrix jacchus) and squirrel monkeys (Saimiri boliviensis). Am J Phys Anthropol 138:403–420
Young TP, Isbell LA (1991) Sex differences in giraffe feeding ecology: energetic and social constraints. Ethology 87:79–89
Zotz G (2006) Substrate use of three epiphytic bromeliads. Ecography 20:264–270
Acknowledgments
I published Moffett (2001) in an up-and-coming journal for canopy research that unfortunately folded. I thank Margaret (“Meg”) Lowman, who, after I talked about this subject for the 2009 International Canopy Conference in Bangalore, gave me the chance to update the article here, so that it can come before a new audience. Luckily, I was able to contact many of the people who advised me on the first version. Although doubtless I missed a few things, I found that little has been done in the last decade on the issues I raise, except for the burgeoning research on biofilms. For assistance with the philosophy of discipline formation, I thank Mary Catherine Bateson, David L. Hull, Peter Harries-Jones, George Lakoff, Paul Ryan, and Carol Wilder; for thoughts on trees, forests, and the dimensionality of canopies, David Ackerly, Timothy F.H. Allen, Robert G. Bailey, Dennis Baldocchi, Frans Bongers, James H. Brown, Jerome Chave, Joel Clement, Raphael Didham, Brian J. Enquist, Stephen P. Ellner, Thomas J. Givnish, Juan Gouda, Paul G. Jarvis, David King, Steve Lindberg, Richard Law, Orie Loucks, Margaret Lowman, Gary Lovett, Jonathan Majer, Lauri Oksanen, Karl J. Niklas, John M. Norman, Geoffrey Parker, Serguei Ponomarenko, Hank H. Shugart, Jonathan Silvertown, Anthony R.E. Sinclair, Frank Sterck, Steven Sutton, Bastow Wilson, Neville Winchester, and Truman Young; for information on roots, Peter W. Barlow, Jan Čermák, Hans de Kroon, A. Roland Ennos, Lewis Feldman, Roman Gebauer, Robert B. Jackson, Donald R. Kaplan, Takashi Kohyama, Krista Lõhmus, Jonathan Lynch, Liesje Mommer, John Moore, James H. Richards, Wim van der Putten, Fernando Tuya, and Yoav Waisel; and for views on algal, bacterial, coral reef, and other canopies, Mohammad A Al-Najjar, Yves Basset, David Benzing, James D. Bryers, Robert C. Carpenter, the late and great J. William Costerton, Paul Dayton, Dirk de Beer, Phil DeVries, Peg Dirckx, Michael Dolan, David Duggins, Michael Franklin, Elizabeth Gladfelter, Ken Heck, Brian Helmuth, Helmut Hillebrand, Walter R. Hill, Mark Hixon, Bo Barker Jørgensen, Ronald H. Karlson, Mimi Koehl, Michael Kühl, Scott Larned, Steven E. Lindow, Rex L. Lowe, my recently deceased friend Lynn Margulis, Susan Merkel, Peter J. Morin, Cindy Morris, James W. Porter, Jennifer H. Richards, Kenneth P. Sebens, Vaclav Smil, Robert S. Steneck, Valerie Behan-Pelletier, Alice Tangerini, Chantal Vis, Martin Wahl, and Susan L. Williams. Lapses in coverage and logic can be attributed to me.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Moffett, M.W. (2013). Comparative Canopy Biology and the Structure of Ecosystems. In: Lowman, M., Devy, S., Ganesh, T. (eds) Treetops at Risk. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-7161-5_3
Download citation
DOI: https://doi.org/10.1007/978-1-4614-7161-5_3
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-7160-8
Online ISBN: 978-1-4614-7161-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)