
Abstract
Exercise training in combination with optimal nutritional support is an effective strategy to maintain or increase skeletal muscle mass. A single bout of resistance exercise undertaken with adequate protein availability increases rates of muscle protein synthesis and, when repeated over weeks and months, leads to increased muscle fiber size. While resistance-based training is considered the ‘gold standard’ for promoting muscle hypertrophy, other modes of exercise may be able to promote gains in muscle mass. High-intensity interval training (HIIT) comprises short bouts of exercise at or above the power output/speed that elicits individual maximal aerobic capacity, placing high tensile stress on skeletal muscle, and somewhat resembling the demands of resistance exercise. While HIIT induces rapid increases in skeletal muscle oxidative capacity, the anabolic potential of HIIT for promoting concurrent gains in muscle mass and cardiorespiratory fitness has received less scientific inquiry. In this review, we discuss studies that have determined muscle growth responses after HIIT, with a focus on molecular responses, that provide a rationale for HIIT to be implemented among populations who are susceptible to muscle loss (e.g. middle-aged or older adults) and/or in clinical settings (e.g. pre- or post-surgery).
Similar content being viewed by others
Avoid common mistakes on your manuscript.
High-intensity interval training (HIIT) possesses inherent similarities to resistance training and may be a viable exercise modality to maintain muscle mass. However, no scientific literature has evaluated the anabolic effects of HIIT alone or combined with optimal nutritional support (i.e. increased protein availability) on skeletal muscle adaptations. |
Emerging evidence from human studies shows that HIIT can modulate expression of genes and proteins implicated in muscle mass regulation, increase muscle protein synthesis and activate muscle satellite cells. |
Despite stimulation of acute molecular events that precede gains in muscle mass with HIIT alone or with increased protein availability, further research assessing chronic muscle mass/size responses is needed to elucidate the role of HIIT in counteracting muscle loss with age. |
1 Introduction
Human skeletal muscle comprises ~ 40% of body mass and plays fundamental roles in locomotion, thermoregulation and metabolic health [1]. Sarcopenia, the gradual loss of skeletal muscle mass with age, is closely linked with adverse health outcomes including reduced physical function, independence, quality of life and increased risk of premature death [2,3,4]. The global prevalence of sarcopenia is contentious due to different definitions of diagnosis [5], although this condition is now recognised as a reportable medical condition by the World Health Organisation [6]. Sarcopenia is associated with substantial personal suffering along with loss of independence, and places a significant economic burden on healthcare systems worldwide [7, 8]. Skeletal muscle mass decreases by ~ 1–2% per year after the age of 60 years [9,10,11] whereas muscle strength declines by up to 3% per year after the sixth decade [11, 12]. As peak muscle mass in mid-adulthood is related to muscle mass and strength in older age [13], maintenance of muscle mass throughout early and mid-life is crucial for mobility and prevention of physical inactivity-related chronic diseases.
Resistance-based exercise and optimal nutritional support (i.e. increased protein availability) are potent modulators of skeletal muscle protein turnover that work synergistically to promote gains in skeletal muscle mass [14, 15]. Divergent exercise modalities initiate unique molecular programs resulting in the accumulation of skeletal muscle proteins that, over time, modulate muscle function and phenotype [1]. Resistance training, characterised by repeated contractions against near-maximal external loads, increases the synthesis of myofibrillar proteins and promotes satellite cell or nuclear addition to pre-existing myofibers [15]. Resistance training increases the cross-sectional area (CSA) of muscle fibers (hypertrophy) and muscle mass, which is related to the capacity to move an external load (strength) [1]. Crucially, fat-free mass (as an estimate of muscle mass) and strength are both independent predictors of all-cause mortality in older adults [16, 17], highlighting the importance of regular exercise to counteract the negative effects of sarcopenia. Although resistance training is regarded as the ‘gold standard’ for promoting gains in muscle mass and maximal strength, participation rates remain low among older adults [18,19,20,21], in part due to a need for specialised equipment and correct technique to prevent injury. In contrast, moderate intensity continuous training (MICT) performed at submaximal intensities (45–75% maximal oxygen uptake [VO2max]) for 30 min or longer can be performed safely with little/no equipment or supervision. Aerobic training primarily stimulates cell signalling cascades that modulate the expression of nuclear and mitochondrial genes encoding mitochondrial proteins [1]. Increases in mitochondrial density enhance skeletal muscle oxidative capacity and cardiorespiratory fitness, the latter of which declines with age [22], and is inversely associated with cardiovascular disease risk and all-cause mortality [23]. Several factors preclude the performance of any form of exercise training, with a ‘lack of time’ remaining the most commonly cited barrier to regular participation [24]. As such, time-efficient, practical and stimulating exercise prescription that maintains muscle mass and function is important countermeasure to delay the onset of sarcopenia.
Interval training, such as high-intensity interval training (HIIT) and sprint interval training (SIT), has recently risen in popularity [25], and may be a viable, yet overlooked alternative to resistance training for promoting muscle mass accrual. HIIT is infinitely variable, but typically characterised by brief periods (≤ 4 min) of intense continuous exercise [80–100% peak heart rate (HRpeak)] interspersed with short periods of rest or recovery. In contrast, SIT involves shorter (≤ 30 s) ‘all-out’ work periods performed at ≥ 100% of the power output/speed that elicits an individual’s VO2max [26]. The enhanced metabolic and cardiorespiratory effects of HIIT and SIT in skeletal muscle have been well documented [27, 28]. However, much less is known about the impact of aerobic-based HIIT and SIT modalities (e.g. cycling, running, swimming) on muscle growth responses and whether these interventions can promote gains in muscle hypertrophy, lean mass and strength, particularly when undertaken with optimal nutritional support. Furthermore, it is of interest to exercise physiologists, clinicians and the general population alike to determine if HIIT can maintain and/or improve cardiorespiratory fitness and muscle mass concurrently, given the reduced time commitment compared to traditional resistance or aerobic exercise modalities. Accordingly, the principal aim of this review is to critically evaluate emerging evidence that HIIT, either in isolation or combined with increased protein availability, can promote skeletal muscle anabolism. We conducted a thorough PubMed search of literature up until October 2020 that examined skeletal muscle and whole body responses to HIIT in young, middle-aged and older adults. First, studies investigating the cellular responses related to muscle hypertrophy following HIIT are discussed. Subsequently, investigations that have determined whole body functional outcomes in response to HIIT are considered. Collectively, the findings from these studies provide a molecular basis for the efficacy of HIIT to complement resistance-based exercise training or, under certain conditions, provide an alternative stimulus for skeletal muscle anabolism.
2 Defining the Diversity in Skeletal Muscle Adaptations with HIIT
Various HIIT and SIT protocols in both healthy and clinical populations have been shown to improve cardiometabolic health outcomes [26, 29, 30]. Some of the more popular and well researched protocols are the ‘Norwegian’ [31,32,33,34,35], ‘Gibala’ [36,37,38,39] and ‘Tabata’ models [40,41,42], as well as Wingate-based training [43,44,45,46] and reduced exertion HIIT (REHIT) [47,48,49] (Table 1). Importantly, various interval training regimes [50,51,52], particularly the ‘Norwegian’ model [31, 34, 53,54,55] have been shown to increase aerobic fitness in older adults. HIIT improves skeletal muscle oxidative capacity, primarily via activation of signalling cascades that stimulate mitochondrial biogenesis and angiogenesis [27]. SIT and MICT can induce similar improvements in oxidative metabolism with vastly different training volumes [43, 49]. As such, interval training is often considered a time-efficient alternative to traditional aerobic training.
Despite similarities in skeletal muscle oxidative capacity and subjective ratings of enjoyment between HIIT and MICT [56], two weeks of HIIT induce a different neuromuscular profile to MICT in young men [57, 58]. Using high-density electromyography (EMG), HIIT increases muscle fiber conduction velocity [58], maximal knee extensor torque and discharge rate of high-threshold motor units [57], all factors related to maximal force production. In contrast, only minor changes in these functional measures have been observed after MICT. There is a greater activation of type II muscle fibers with increasing exercise intensity/contractile force [59, 60], with these fibres having the greatest potential for hypertrophy following resistance training [61]. Considering that changes to neural factors and muscle fiber CSA following resistance training are directly linked to maximal strength gains [62], HIIT may also increase muscle strength, albeit to a lesser magnitude, particularly in populations sensitive to relatively intense exercise (e.g. untrained/ageing populations). While the results from these studies [57, 58] provide information regarding neuromuscular adaptations with HIIT, the focus of this review is on the capacity for HIIT to increase skeletal muscle mass and size as a practical strategy to offset the inevitable loss in muscle mass with advancing age.
3 Molecular Responses to HIIT/SIT
3.1 Effects of HIIT/SIT on Skeletal Muscle Transcriptome and Proteome
In response to skeletal muscle contraction, a multitude of cellular events are initiated that modulate the expression of specific gene sets that encode proteins that ultimately form the basis of adaptation responses [63]. The majority of studies investigating molecular responses to HIIT have focused on pathways regulating mitochondrial biogenesis and insulin sensitivity in skeletal muscle [27]. Recent studies incorporating high-throughput ‘-omics’ techniques have explored the ‘global’ effects of HIIT at the transcriptional and translational levels. Key findings from these studies reveal unique molecular ‘signatures’ supporting the notion that genes and proteins implicated in muscle mass regulation are up-regulated through interval-style exercise (Table 2).
Rundqvist et al. [64] were among the first to examine the global gene expression profiles in skeletal muscle (v. lateralis) from young (~ 26 years) adults in response to a single bout of sprint cycling exercise (3 × 30 s ‘all-out’ sprints). A biopsy obtained ~ 2 h following the final sprint revealed differential expression of 879 genes (471 genes upregulated, 408 genes downregulated). Notably, sprint exercise significantly increased expression of genes implicated in the regulation of muscle mass including frizzled class receptor 7 (FZD7) and myogenic differentiation 1 (MYOD1), while concomitantly downregulating myostatin (MSTN) expression, a key suppressor of skeletal muscle growth. However, the acute nature of the sprint exercise protocol makes it difficult to assess the contribution and involvement of the collective changes in these gene transcripts to promote the requisite molecular signals for muscle hypertrophy responses. Extending these findings, Miyamoto-Mikami et al. [65] examined global gene expression profiles (v. lateralis) following six weeks of Tabata-style SIT (cycling exercise, 6–7 × 20 s, 170% VO2max) in healthy young (~ 23 years) men [65]. The authors reported 79 genes were upregulated with 73 genes downregulated post-intervention [65]. Given the transient nature of many exercise-sensitive transcripts, and the time-course of sampling of the post-training muscle biopsy (48–72 h after the last training session), it is likely that changes in expression of many transcripts may have been missed. Nonetheless, the CSA of the quadriceps femoris and hamstring muscles (assessed by magnetic resonance imaging; MRI) were both increased following SIT, despite downregulation of several genes, such as myosin heavy chain 1 (MYH1), myosin light chain kinase 2 (MYLK2) and nebulin-related anchoring protein (NRAP) [65], genes that encode proteins with putative roles in contractile function. Furthermore, gene expression of myostatin was decreased following HIIT [65]. In addition to changes at the mRNA level, the authors also observed increased protein expression of carnosine synthase 1 (CARNS1), myosin light chain kinase family member 4 (MYLK4), protein phosphatase 1 regulatory subunit 3 C (PPP1R3C), serum/glucocorticoid regulated kinase 1 (SGK1) and peroxisome proliferator-activated receptor gamma (PPARGC1A) [65]. CARNS1 and MYLK4 are involved in events that may improve force-generating capacity. Carnosine synthase, an enzyme catalysing the β-alanyl-l-histidine dipeptide to carnosine encoded by the CARNS1 gene, improves pH buffering capacity and increases calcium (Ca2+) sensitivity to the contractile apparatus [66]. Skeletal muscle myosin light chain kinase, a Ca2+/calmodulin-dependent protein kinase encoded by the MYLK gene, phosphorylates the regulatory light chain of myosin in the sarcomere providing mechanical support during force generation [67]. Collectively, these data provide preliminary evidence for enhanced calcium handling that may support improved force-generating capacity and possibly muscle hypertrophy following SIT. Indeed, contraction-induced alterations in intracellular [Ca2+] may be linked to distinctive programs of gene expression that establish phenotypic diversity among skeletal muscle fibers and confer some of the whole body adaptations after SIT protocols [68].
Robinson et al. [34] compared transcriptome and proteome responses from skeletal muscle of young (~ 25 years) and older (~ 70 years) adults obtained 72 h after the final training session of a 12 week programme of either cycling HIIT [4 × 4 min at > 90% peak oxygen uptake (VO2peak)], resistance training (whole body, 2–4 sets × 8–12 reps) or combined training (HIIT and resistance training). They reported increased expression of 22 genes in older adults following HIIT, including the genes collagen type XIV alpha 1 chain (COL14A1) and lumican (LUM), with roles in extracellular matrix (ECM) organisation, and integrin subunit beta 2 (ITGB2), which is involved in integrin signalling. Increased basal expression of these genes may support enhanced ECM tensile strength, cell-to-ECM adhesions and mechanotransduction signalling [69]. Increased COL14A1 and LUM expression has been reported following 12 weeks of MICT, suggestive of enhanced mechanotransduction to ECM components [70], although the precise role of ECM reorganization in facilitating muscle growth responses following aerobic training remains unknown. HIIT also increased the expression of 11 genes in the older adults that were significantly downregulated prior to exercise training compared to younger adults including MYLK4 (actin cytoskeleton regulation) and KAZALD1 (insulin-like growth factor binding). Given both the MYLK4 gene and protein expression have previously been shown to increase following SIT in younger adults [65], increased mechanical support to the sarcomere to facilitate higher-intensity contractions may be a characteristic molecular response to HIIT/SIT independent of age. Another key finding from the study by Robinson et al. [34] was that the expression of genes upregulated with HIIT and resistance training showed considerable overlap in both older (81 genes) and younger adults (88 genes). Collectively, results from these investigations [34, 64, 65] provide evidence of transcriptional and translational responses implicated in muscle growth responses following exposure to SIT and HIIT.
3.2 Effects of HIIT/SIT on Rates of Muscle Protein Synthesis
Increases in rates of muscle protein synthesis and the concomitant increases in muscle mass occur in response to sustained periods of positive net protein balance (NPB), when the rates of muscle protein synthesis (MPS) exceed that of muscle protein breakdown [71]. Two recent studies have used deuterium oxide (D2O) tracer methodology to measure MPS following SIT and HIIT. Scalzo et al. [46] investigated the integrated v. lateralis MPS response over the course of a three-week SIT intervention (nine cycling sessions, 4–8 × 30 s, 100% VO2max) in young (~ 23 years) adults. Contrary to their hypothesis and previous literature demonstrating no sex differences in exercise-induced MPS [72, 73], males had greater rates of both mixed (~ 0.40 vs. ~ 0.25%.day−1) and cytoplasmic (~ 0.40 vs. ~ 0.29%.day−1) protein synthesis compared to females. Regardless of the sex-based differences, post-intervention increases in mixed and cytosolic protein synthesis demonstrated that three weeks of SIT can stimulate increases in MPS in human skeletal muscle.
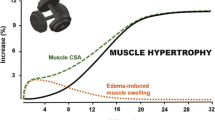
Bell et al. [74] compared protein fractional synthetic rates (FSR) following a single session of either HIIT (10 × 1 min cycling at ~ 95% HRpeak), resistance exercise (3 sets of leg press and leg extension at ~ 95% 10RM with last set to failure) or MICT (30 min cycling at ~ 70% HRpeak) in untrained older (~ 67 years) men. Participants consumed D2O for nine days and muscle biopsies were obtained from the v. lateralis on days 5–8 and used to estimate integrated myofibrillar and sarcoplasmic FSR during the 48-h period following each of the exercise bouts. Rates of myofibrillar FSR significantly increased in the 24 and 48 h period following the single bout of HIIT compared to rest [74]. The magnitude of the HIIT-induced increase in myofibrillar protein synthesis was less compared to resistance exercise (~ 50% versus ~ 80%) but greater than MICT (~ 10%) 24 h post exercise. Additionally, HIIT was the only exercise modality to increase sarcoplasmic protein synthesis 24 h post exercise (~ 25%), a response the authors suggested may be due to increased mitochondrial protein synthesis. The acute [74] and short-term training-induced [46] increases in MPS with HIIT and SIT demonstrate that HIIT can regulate the molecular machinery that underpins muscle hypertrophy. Some studies have reported that acute changes in integrated MPS following resistance exercise align with changes in the short-term (three weeks) adaptive hypertrophic response [75]. However, acute changes in MPS do not always reflect subsequent increases in muscle hypertrophy [76,77,78] particularly in the short-term where oedematous muscle swelling is likely an artefact contributing to measurements of ‘hypertrophy’ [79]. Whether acute rises in MPS align with HIIT-induced muscle hypertrophy are yet to be determined. In any case, the available literature suggests that skeletal muscle remodelling with HIIT may extend beyond established changes in oxidative capacity and substrate metabolism.
3.3 Effects HIIT/SIT on Satellite Cell Dynamics, Myonuclear Content and Muscle Fiber Cross-Sectional Area
Investigating the activity of satellite cells in response to HIIT can provide mechanistic insight into potential factors regulating the remodelling of skeletal muscle beyond that of gross measures of muscle hypertrophy [80]. However, compared to resistance exercise, few studies have investigated satellite cell responses following HIIT. Nederveen and co-workers [81] compared satellite cell responses in older (~ 67 years) men at 24 and 48 h post-exercise between HIIT (10 × 1 min cycling at ~ 95% HRpeak), resistance exercise (three sets of bilateral leg press and leg extension at 95% of 10RM with last set to failure) and MICT (30 min cycling at ~ 70% HRpeak). While satellite cell content (Pax7+ cells) in type I fibers was elevated at 24 and 48 h following resistance exercise, satellite cell activity (Pax7+/MyoD+ cells) increased at the same time points only following HIIT. As such, these findings do not exclude the possibility of HIIT can induce increases in satellite cell content later during post-exercise recovery (i.e. > 48 h post-exercise) in this cohort.
Joanisse et al. examined satellite cell dynamics following short-term HIIT using a 10 × 1 min cycling protocol performed three times per week over 6 weeks in untrained young (~ 27 years) women [36]. Despite no significant change in the myonuclear content or CSA of myosin heavy chain one (MHCI), myosin heavy chain two (MHCII) or hybrid fibers, a 96 h post-training muscle biopsy revealed that HIIT induced a significant increase in the number of quiescent and differentiating satellite cells associated with hybrid fibers (i.e. those displaying MHC I and II). There was also a significant 15% increase in the number of hybrid fibers containing centrally located nuclei, a commonly used marker of skeletal muscle remodelling and repair [36]. The absence of muscle fiber hypertrophy but increased satellite cell activity with hybrid fibers and fast-to-slow fiber type distribution (i.e. increase in hybrid fibers and decrease in MHCII fibers) led the authors to conclude that HIIT contributed to ‘non-hypertrophic’ remodelling of muscle fibers [36]. However, not all studies report such changes in fibre-type distribution with short-term (four weeks) HIIT in young (~ 21 years) adults [42].
Active satellite cells have been shown to reside in closer proximity to capillaries than quiescent satellite cells at rest and 24 h following a single bout of high-intensity resistance exercise in young men [82]. Given aerobic-based exercise training increases satellite cell activity [36, 41], an enhanced spatial relationship between active satellite cells and capillaries may increase uptake of circulating substrates known to modulate satellite cell function. Mitochondrial metabolism regulates satellite cell fate in vitro whereby oxidative phosphorylation promotes satellite cell differentiation and repression which is obligatory for satellite cell self-renewal [83]. Thus, aerobic-based exercise training-induced alterations in mitochondrial activity may modulate satellite cell function. Abreu et al. [84] reported no change in muscle fiber CSA in young mice following five weeks of MICT compared to non-exercised controls. However, MICT resulted in enhanced skeletal muscle repair, assessed by centrally located nuclei counts on individual fibers, seven days following muscle injury. Furthermore, isolated satellite cells from the exercised mice showed profiles indicative of improved activation and self-renewal while mRNA expression of mitochondrial markers remained unchanged and respiration at rest was repressed [84]. As such, aerobic-based exercise training-induced changes to skeletal muscle mitochondria and the microvasculature may regulate satellite cell function to facilitate enhance skeletal muscle remodelling [80].
In another investigation comprising two independent 6-week study protocols, Joanisse et al. implemented divergent aerobic training protocols to examine satellite cell activity [41]. The first six-week study compared Tabata-style interval training to MICT performed three times per week by recreationally active young (~ 21 years) adults. In the second six-week intervention, young (~ 29 years) overweight adults performed three all-out 20 s cycling sprints against a resistance set to 0.05 kg.kg body mass [BM]−1 (with 2 min of active rest between work bouts) three times per week [41]. Similar to their previous findings [36], no changes were observed in myonuclear content or muscle fiber CSA with either training intervention [41], despite an increase in the number of active and differentiating satellite cells following all three exercise protocols. Taken together, results from the aforementioned studies demonstrate that satellite cells are activated in response to different short-term aerobic training protocols in young adults, and such changes occur independent of muscle fiber hypertrophy. In contrast, Charifi et al. [85] observed increased v. lateralis satellite cell content in the absence of myonuclear accretion or mixed muscle fiber hypertrophy in response to 14 weeks of aerobic-based (i.e. cycling endurance and HIIT) training in older (~ 73 years) men. However, fiber-type analysis revealed larger type IIA fibers following the intervention. Verney et al. [86] reported v. lateralis fiber hypertrophy and increases in the satellite cell pool from both the v. lateralis and deltoid following 14 weeks’ combined lower body cycling HIIT and upper body resistance training (i.e. both modalities performed at each training session) in older (~ 73 years) men. Similarly, Snijders et al. [87] observed increased v. lateralis satellite cell content in the absence of muscle fiber hypertrophy or myonuclear content following 12 weeks of combined training (one session of cycling HIIT and two sessions of whole body resistance training per week) in older (~ 74 years) men. Taken together, results from these studies [85, 87] suggest that ~ 3–4 months of combined HIIT and resistance training can increase muscle fiber hypertrophy and satellite cell content in older men. Whether prolonged HIIT induces muscle fiber hypertrophy in young, middle-aged and older adults remain to be determined.
From the limited data available, HIIT performed for ~ six weeks in young untrained adults does not lead to muscle fiber hypertrophy or satellite cell-mediated myonuclei accretion. Whether HIIT can increase satellite cell and myonuclei content in middle-aged and older adults following short-term or prolonged interventions also remains to be determined. However, increases in satellite cell activation following acute and short-term HIIT indicate the potential for HIIT to regulate muscle satellite cell dynamics [36, 41, 81]. Muscle protein synthesis and satellite cell responses provide important mechanistic insight regarding HIIT-induced skeletal muscle remodelling. However, changes in body composition and skeletal muscle mass also require consideration given phenotypic changes to tissues (e.g. lean/fat-free mass) are often linked with health outcomes.
4 Body Composition and Skeletal Muscle Morphology Responses to HIIT/SIT
4.1 Effects of HIIT/SIT Lean or Fat-Free Mass
The majority of studies that have reported changes in body composition following HIIT or SIT have utilised dual-energy X-ray absorptiometry (DXA) where changes in total, appendicular lean tissue or fat-free mass are used as a proxy for skeletal muscle mass. A recent meta-analysis of 47 studies found no differences in body composition (i.e. increased lean body mass and/or decreased fat mass) between low volume HIIT and MICT or a non-exercise control [88]. However, several other investigations that were not included in that meta-analysis because they did not meet inclusion criteria (e.g. no MICT group or non-exercising control) have measured changes in lean/fat-free mass in response to various interval training protocols involving cycling [34, 37, 50, 89,90,91,92,93,94,95,96,97,98,99,100,101], running [102, 103], rowing [104], whole body [51, 105] and elliptical-based [106] HIIT/SIT. While some of these investigations failed to detect changes in lean/fat free mass [37, 50, 90,91,92, 96, 98, 100, 101, 103], others reported an increase in lean/fat-free mass in response to HIIT [34, 89, 93,94,95, 102, 104, 107,108,109]. Most studies that have observed increases in lean/fat-free mass with HIIT incorporated training durations of ≥ 12 weeks duration undertaken by young [34, 93, 95, 104], middle-aged [89, 109] and older adults [34]. Additionally, short-term HIIT interventions (6–8 weeks) have also induced increases in lean/fat-free mass in young [94, 107, 108] and middle-aged adults [102].
As previously noted, the aforementioned studies have estimated alterations in lean/fat-free mass using DXA methodology: however, several limitations need to be taken into account when considering exercise training-induced changes in body composition using this approach. For example, the precision (trueness) of whole body lean mass measurements, as estimated from the coefficient of variation (CV) ranges from ~ 0.5 to 1% depending on the densitometer used [110]. Additionally, DXA cannot distinguish muscle from intramuscular fluid and is affected by hydration status [111, 112]. These factors have raised questions regarding the validity of DXA-derived changes in muscle mass [113]. In contrast, techniques, such as magnetic resonance imaging (MRI) and computed tomography (CT), are considered as reference methods for measuring whole body and regional skeletal muscle mass [114]. However, notable limitations of MRI and CT include the cost and expertise to operate and maintain the scanners and in the case of CT, exposure to larger doses of ionising radiation.
4.2 Effects of HIIT/SIT on Skeletal Muscle Morphology
Using MRI, Osawa et al. [105] reported that 16 weeks of cycling HIIT (8–12 × 60 s, > 90% VO2peak) or whole-body HIIT (cycling followed by arm cranking ergometry [4–6 × 60 s, > 90% peak workload]) in healthy young (~ 35 years) men increased CSA of thigh and trunk muscles. Specifically, quadriceps femoris CSA increased with both types of HIIT (cycling: 7.7%, whole body: 5.2%) but only whole body HIIT increased trunk and abdominal muscle CSA. In contrast, DXA-derived total and regional lean mass remained unchanged following training. Similarly, HIIT has been shown to increase MRI-derived CSA and volume of skeletal muscle. In older (~ 68 years) men, eight weeks cycling HIIT (7 × 120 s, 80–90% VO2max) increased quadriceps femoris CSA (4.3%) and volume (5.8%) despite no change in total lean mass as assessed by DXA [90]. HIIT also increased maximal isometric torque at 60° of knee flexion but not at 90° of knee flexion or maximal isokinetic torque [90], while two-dimensional ultrasound detected increases in v. lateralis CSA in response to cycling [37] and running [115] HIIT in untrained young adults. In both studies that used both MRI and DXA to estimate training-induced changes in muscle size/mass following HIIT, MRI detected changes in CSA whereas DXA measures did not [90, 105]. In summary, findings from these studies, particularly those using MRI to assess changes in whole muscle CSA, provide data to suggest that HIIT has the potential to induce significant changes in whole muscle CSA.
5 HIIT and Increased Protein Availability
Nutrient availability is a key factor mediating exercise-induced skeletal muscle adaptations [116]. A key omission of the majority of studies discussed is the capacity for nutrient ingestion, particularly increased protein availability, to augment changes in skeletal muscle mass with HIIT. Dietary protein, principally the essential amino acid leucine, increases MPS [117, 118] and satellite cell activity [119]. Of the major macronutrients, post-exercise protein ingestion is crucial for skeletal muscle remodelling by stimulating rates of MPS through the transfer and incorporation of amino acids into skeletal muscle proteins [14]. Indeed, positive muscle protein turnover and net accretion of myofibrillar proteins are only achieved through post-exercise protein feeding [120]. However, the magnitude to which protein may promote anabolic training adaptations is dependent on several factors including the dose, source, timing and distribution of intake. A comprehensive discussion of these factors is beyond the scope of the present review and the reader is referred to other sources on these topics [14, 121, 122].
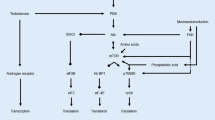
Protein supplementation in combination with resistance training amplifies changes in muscle fiber CSA, lean mass and strength [123, 124]. Protein intake in close proximity to aerobic exercise increases post-exercise rates of MPS [125,126,127] and attenuates muscle protein breakdown [128]. However, little is known about possible synergistic effects of HIIT and increased protein availability on anabolic skeletal muscle adaptations. One of the first studies to investigate the potential for protein ingestion to augment anabolic responses after HIIT/SIT was that of Coffey et al. [129] who reported that post-exercise rates of myofibrillar protein synthesis were ~ 48% higher (~ 0.083 vs ~ 0.056% h−1) in young (~ 21 years) men performing repeated cycling sprints (10 × 6 s, 0.75 N torque kg−1 interspersed by 60 s of recovery) following the ingestion of a pre-exercise meal containing 24 g whey protein (4.8 g leucine) and 50 g maltodextrin, or a placebo [129] (Fig. 1). While protein ingestion and repeated sprints resulted in higher rates of myofibrillar protein synthesis, there was little effect of feeding on the rates of mitochondrial protein synthesis during a ~ 4 h post-exercise recovery for nutrient or placebo treatments. The increase in myofibrillar protein synthesis with protein ingestion was associated with significant increases in the phosphorylation status of several key signalling proteins mediating translation initiation, such as protein kinase B (Akt), mTOR mammalian target of rapamycin (mTOR), ribosomal protein S6 kinase beta-1 (p70S6K) and ribosomal protein S6 (rpS6).

Acute rates of myofibrillar protein synthesis following cycling sprints with and without increased protein availability. Myofibrillar fractional synthesis rate figure (left) adapted from Coffey et al. [129], with permission. Combined exercise and diet interactions that stimulate myofibrillar protein synthesis induce muscle hypertrophy when repeated over time (i.e. weeks to months). Previous evidence of a single bout of sprint interval exercise with protein ingestion significantly increasing rates of myofibrillar protein synthesis compared to a placebo condition (*) raises the possibility that sustained increases may promote increases in muscle fiber hypertrophy. However, the extent to which acute increases in rates of muscle protein synthesis form the basis of chronic muscle hypertrophy responses is equivocal with accumulating evidence in resistance training models indicating such increases, in part, likely contribute to extensive muscle repair and remodelling of damaged proteins prior to facilitating muscle fibre hypertrophy [78, 130]
Recently, Rundqvist et al. [131] investigated the effect of an acute bout of SIT (3 × 30 s sprints separated by a 20 min recovery) in young (~ 26 years) healthy men with the co-ingestion of 300 mg kg BM−1 of essential amino acids and 1 g kg BM−1 of maltodextrin 5 min before the first sprint and 15 min after each of the remaining sprints. Compared to a placebo treatment, cycling sprints in the fed state resulted in greater gene and protein expression of sodium-coupled neutral amino acid transporter 2 (SNAT2), Akt and mamillian target of rapaymycin complex 1 (mTORC1) [131]. Moreover, post-exercise plasma (~ 42%) and muscle (~ 15%) FSR rates were higher with the nutrient compared to placebo condition [131]. Together, the results from these studies provide evidence of acute changes to molecular networks that support myofibrillar [129] and mixed muscle [131] protein synthesis with high-intensity interval-based exercise performed in the fed state, supporting the notion that HIIT undertaken with increased protein availability may be able to promote synergistic increases in muscle hypertrophy. Whether the cumulative effect of repeated HIIT sessions with increased protein availability can elicit similar or greater increases in rates of MPS over weeks/months remains an area for future investigation.
There is a paucity of information regarding the capacity for HIIT and increased protein availability to induce positive changes in muscle fiber hypertrophy or other measurements indicative of increases in skeletal muscle size/mass (e.g. MRI-derived muscle CSA/volume, DXA-derived lean body mass, ultrasound-derived muscle thickness). Leuchtmann et al. [132] observed no change in muscle fiber CSA following 12 weeks of cycling HIIT combined with post-exercise whey protein ingestion (30 g) in older (~ 66 years) men. However, total daily protein intake was not reported in that study. This is an important consideration as meeting a daily dietary protein intake appears to be critical for exercise-induced increases in muscle hypertrophy compared to protein feedings in close temporal proximity to an exercise bout [123].
In middle-aged (~ 55 years) adults with type 2 diabetes mellitus, 10 weeks of mixed model interval training (MMIT; consisting of HIIT and low-intensity high-volume resistance exercise performed on alternative days) combined with 20 g of whey protein before and after each exercise session provided no further increases in v. lateralis CSA compared to a non-protein isoenergetic control beverage [133]. In that study, participants were encouraged to maintain dietary habits during the experimental period, although macronutrient intake was not reported [133]. As such it is difficult to determine if both total daily protein intake and distribution were adequate to support skeletal muscle protein accretion. Furthermore, given that an increase in VO2max but not 1RM was reported in that study [117], it cannot be ruled out that the MMIT protocol may have contributed to blunted anabolic effects, particularly considering that combined strength and aerobic-based training typically negate some of the gains in muscle strength attained after single-mode training [134].
6 Conclusion and Future Directions
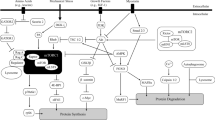
Evidence presented in this review supports the potential for HIIT to stimulate cellular responses that regulate metabolic pathways with putative roles in skeletal muscle hypertrophy. Acute and short-term HIIT increase rates of MPS and emerging evidence from ‘-omics’-based approaches suggests HIIT upregulates unique gene ‘signatures’ along with proteins implicated in anabolic signalling transduction. While HIIT does not increase muscle fiber CSA or satellite cell content, short-term and prolonged HIIT across a range of age groups have been associated with increases in whole muscle (i.e. MRI-derived muscle CSA/volume) and whole body (i.e. DXA-derived lean mass), surrogates of muscle hypertrophy. Notably, the majority of studies that have reported measures of muscle anabolism incorporating molecular and whole muscle/body measures have failed to provide optimal nutritional support (i.e. increased protein availability), which is considered essential to promote muscle hypertrophy in association with resistance exercise. Further work investigating the synergistic capacity for HIIT and long-term protein supplementation based on strategies that maximise muscle protein synthetic responses (i.e. timing, distribution, high-quality liquid and food sources) to promote lean mass accrual is urgently needed (Fig. 2). Moreover, we implore researchers to report relative daily intake values for protein intake (e.g. g kg BM−1) to better understand protein requirements for HIIT-induced muscle hypertrophy.

Schematic of putative factors that can be manipulated to induce muscle anabolism with combined HIIT and increased protein availability and cellular mechanisms that may underpin eventual gains in muscle mass. Exercise modalities that increase total musculature under load during work periods (e.g. running, whole body HIIT) and consuming a daily protein intake (≥ 1.6 g kg BM−1) known to augment resistance training-induced muscle hypertrophy [123] are pivotal to uncovering the anabolic potential of HIIT and increased protein availability. At the myocellular level, HIIT and increased protein availability increases phosphorylation of the Akt/mTOR pathway stimulating myofibrillar protein synthesis [129, 131]. HIIT increases sarcoplasmic [74] and mitochondrial [46] protein synthesis that over time may contribute to modest increases in muscle mass [34]. Increased expression of the myogenic regulatory factor MYOD1, amino acid transporter SNAT2 and Wnt signalling transmembrane receptor FZD7 as well as decreased expression of negative muscle growth regulator MTSN represents some of the gene-encoding proteins that may contribute to pathways regulating muscle fiber size with HIIT and increased protein availability [64, 131]. Furthermore, increased protein expression of CARNS1 and MYLK4 may increase calcium handling to support higher-intensity muscle contractions at the sarcomere [65]. Extracellular matrix remodelling may also play a supporting role in facilitating higher intensity muscle contractions by reorganization of collagen fibers to enhance transmission of tensile forces [34]. 4EBP1 eukaryotic translation initiation factor 4E-binding protein 1Akt, protein kinase B, BM body mass, Ca2+ calcium, CARNS1 carnosine synthase, FZD7 frizzled class receptor 7, MYLK4 myosin light chain kinase 4, MYOD1 myoblast determination protein 1, MSTN myostatin, mTOR mammalian target of rapamycin, p70S6K ribosomal protein S6 kinase beta-1, SNAT2 sodium-coupled neutral amino acid transporter 2. Green arrow denotes positive regulator of muscle growth, red arrow denotes negative regulator of muscle growth; blue dotted line denotes pathway that may be implicated in muscle growth with HIIT
There is currently a paucity of information that directly compares muscle growth response with HIIT and resistance training. Future studies including both single-mode HIIT and resistance training are warranted to understand where HIIT fits on an ‘exercise continuum’ with regard to stimulating muscle hypertrophy. Moreover, unravelling the potential of HIIT to induce muscle hypertrophy will require manipulation of variables known to promote resistance training-induced muscle hypertrophy. Thus, further investigation of HIIT following chronic training and in populations of different age groups/training status will be required. Specifically, prolonged interventions (≥ 12 weeks) assessing (a) rates of MPS in all muscle protein pools (i.e. myofibrillar, sarcoplasmic and mitochondrial), (b) global changes in gene and protein expression, and (c) muscle fiber-type specific changes in CSA/satellite cell dynamics, will provide novel insights into the hypertrophic potential of HIIT at the cellular level. Moreover, studies investigating potential dynamics between the microvasculature (e.g. flow, density), skeletal muscle mitochondria (e.g. biogenesis, function) and satellite cell activity following HIIT in various age groups will provide novel insight into mechanisms that contribute to muscle function with aging.
Another important consideration is that most studies assessing the anabolic effects of HIIT have used non-weight-bearing (cycling) protocols, presumably because ergometry cycling is a practical and safe exercise modality to implement in a laboratory-based setting. However, lower body aerobic exercise, such as cycling, does not engage upper body musculature to the same extent as whole body resistance training. In other words, it is difficult to observe whole body gains in muscle mass with cycling-only training protocols. As such, HIIT interventions incorporating dual arm and leg cycling ergometry would help to clarify whether robust changes in total lean mass reported with resistance training are also achievable with HIIT. Where cycling HIIT protocols are used, inclusion of segmental assessments of lean mass from the lower limb would be informative to ensure the regional anabolic effects of HIIT are detected. Even modest increases in leg or trunk lean mass may be of functional importance in clinical settings. Middle-aged or older adults may benefit from HIIT where either pre-operative physical conditioning (inclusive of simultaneous increases in muscle mass and cardiorespiratory fitness) is required for surgery clearance or exercise prehabilitation is recommended to improve post-operative outcomes. However, it must be noted that voluntary performance of HIIT may not always be suited to ageing populations despite its time-efficient nature. Moderate-intensity interval training may initially be more appropriate to help ‘condition’ previously sedentary/ageing populations when introducing interval-style training. Furthermore, modalities that aim to minimise potential adverse events arising from compromised balance (e.g. stationary cycling, seated upper body ergometry) should be considered when prescribing HIIT for older adults.
Finally, future studies should aim to include simultaneous measurements at various ‘levels’ of interrogation when studying HIIT-induced muscle anabolism to ensure different attributes contributing to muscle growth responses and hypertrophy are assessed [111]. Such measures include biochemical assays to measure muscle protein concentrations at the molecular level, cell size (i.e. CSA) at the muscle fiber level and MRI/CT at the whole muscle level. In addition to such measures of muscle growth, it is also important to measure the ‘functionality’ of these potential increases in muscle hypertrophy with HIIT. In particular, determining whether HIIT-induced increases in lean mass are concomitant with improvements in maximal muscle strength or results from clinical assessments used to determine functional capacity (e.g. hand grip strength, timed up and go test, 6-min walk test, etc.) will ensure exercise physiologists and clinicians obtain the most meaningful information possible to assist in the rehabilitation of their patients. Overall, while further work is required, HIIT is a promising and viable strategy to promote and maintain muscle mass, and may serve as a useful adjunct to traditional resistance exercise.
References
Hawley JA, Hargreaves M, Joyner MJ, Zierath JR. Integrative biology of exercise. Cell. 2014;159(4):738–49. https://doi.org/10.1016/j.cell.2014.10.029.
Beaudart C, Zaaria M, Pasleau F, Reginster JY, Bruyere O. Health outcomes of sarcopenia: a systematic review and meta-analysis. PLoS ONE. 2017;12(1):e0169548. https://doi.org/10.1371/journal.pone.0169548.
Woo T, Yu S, Visvanathan R. Systematic literature review on the relationship between biomarkers of sarcopenia and quality of life in older people. J Frailty Aging. 2016;5(2):88–99. https://doi.org/10.14283/jfa.2016.93.
Fisher G, Brown AW, Bohan Brown MM, Alcorn A, Noles C, Winwood L, et al. High intensity interval- vs moderate intensity-training for improving cardiometabolic health in overweight or obese males: a randomized controlled trial. PLoS ONE. 2015;10(10):e0138853. https://doi.org/10.1371/journal.pone.0138853.
Mayhew AJ, Amog K, Phillips S, Parise G, McNicholas PD, de Souza RJ, et al. The prevalence of sarcopenia in community-dwelling older adults, an exploration of differences between studies and within definitions: a systematic review and meta-analyses. Age Ageing. 2019;48(1):48–56. https://doi.org/10.1093/ageing/afy106.
Falcon LJ, Harris-Love MO. Sarcopenia and the new icd-10-cm code: Screening, staging, and diagnosis considerations. Fed Pract. 2017;34(7):24–32.
Goates S, Du K, Arensberg MB, Gaillard T, Guralnik J, Pereira SL. Economic impact of hospitalizations in us adults with sarcopenia. J Frailty Aging. 2019;8(2):93–9. https://doi.org/10.14283/jfa.2019.10.
Norman K, Otten L. Financial impact of sarcopenia or low muscle mass - a short review. Clin Nutr. 2019;38(4):1489–95. https://doi.org/10.1016/j.clnu.2018.09.026.
Frontera WR, Hughes VA, Lutz KJ, Evans WJ. A cross-sectional study of muscle strength and mass in 45- to 78-year-old men and women. J Appl Physiol (1985). 1991;71(2):644–50.
Roubenoff R, Hughes VA. Sarcopenia: current concepts. J Gerontol A Biol Sci Med Sci. 2000;55(12):M716–24.
Macaluso A, De Vito G. Muscle strength, power and adaptations to resistance training in older people. Eur J Appl Physiol. 2004;91(4):450–72. https://doi.org/10.1007/s00421-003-0991-3.
von Haehling S, Morley JE, Anker SD. An overview of sarcopenia: facts and numbers on prevalence and clinical impact. J Cachexia Sarcopenia Muscle. 2010;1(2):129–33. https://doi.org/10.1007/s13539-010-0014-2.
Sayer AA, Syddall H, Martin H, Patel H, Baylis D, Cooper C. The developmental origins of sarcopenia. J Nutr Health Aging. 2008;12(7):427–32.
Stokes T, Hector AJ, Morton RW, McGlory C, Phillips SM. Recent perspectives regarding the role of dietary protein for the promotion of muscle hypertrophy with resistance exercise training. Nutrients. 2018. https://doi.org/10.3390/nu10020180.
McGlory C, Devries MC, Phillips SM. Skeletal muscle and resistance exercise training; the role of protein synthesis in recovery and remodeling. J Appl Physiol (1985). 2017;122(3):541–8. https://doi.org/10.1152/japplphysiol.00613.2016.
Bigaard J, Frederiksen K, Tjonneland A, Thomsen BL, Overvad K, Heitmann BL, et al. Body fat and fat-free mass and all-cause mortality. Obes Res. 2004;12(7):1042–9. https://doi.org/10.1038/oby.2004.131.
Li R, Xia J, Zhang XI, Gathirua-Mwangi WG, Guo J, Li Y, et al. Associations of muscle mass and strength with all-cause mortality among us older adults. Med Sci Sports Exerc. 2018;50(3):458–67. https://doi.org/10.1249/mss.0000000000001448.
Merom D, Pye V, Macniven R, van der Ploeg H, Milat A, Sherrington C, et al. Prevalence and correlates of participation in fall prevention exercise/physical activity by older adults. Prev Med. 2012;55(6):613–7. https://doi.org/10.1016/j.ypmed.2012.10.001.
Humphries B, Duncan MJ, Mummery WK. Prevalence and correlates of resistance training in a regional australian population. Br J Sports Med. 2010;44(9):653–6. https://doi.org/10.1136/bjsm.2008.048975.
Lin CY, Park JH, Hsueh MC, Sun WJ, Liao Y. Prevalence of total physical activity, muscle-strengthening activities, and excessive tv viewing among older adults; and their association with sociodemographic factors. Int J Environ Res Public Health. 2018. https://doi.org/10.3390/ijerph15112499.
Bennie JA, Pedisic Z, van Uffelen JG, Charity MJ, Harvey JT, Banting LK, et al. Pumping iron in australia: prevalence, trends and sociodemographic correlates of muscle strengthening activity participation from a national sample of 195,926 adults. PLoS ONE. 2016;11(4):e0153225. https://doi.org/10.1371/journal.pone.0153225.
Schneider J. Age dependency of oxygen uptake and related parameters in exercise testing: an expert opinion on reference values suitable for adults. Lung. 2013;191(5):449–58. https://doi.org/10.1007/s00408-013-9483-3.
Ekblom-Bak E, Ekblom B, Soderling J, Borjesson M, Blom V, Kallings LV, et al. Sex- and age-specific associations between cardiorespiratory fitness, cvd morbidity and all-cause mortality in in 266.109 adults. Prev Med. 2019;127:105799. https://doi.org/10.1016/j.ypmed.2019.105799.
Trost SG, Owen N, Bauman AE, Sallis JF, Brown W. Correlates of adults’ participation in physical activity: review and update. Med Sci Sports Exerc. 2002;34(12):1996–2001. https://doi.org/10.1249/01.mss.0000038974.76900.92.
Thompson WR. Worldwide survey of fitness trends for 2019. ACSM’s Health Fit J. 2018;22(6):10–7. https://doi.org/10.1249/fit.0000000000000438.
Weston KS, Wisløff U, Coombes JS. High-intensity interval training in patients with lifestyle-induced cardiometabolic disease: a systematic review and meta-analysis. Br J Sports Med. 2014;48(16):1227–34. https://doi.org/10.1136/bjsports-2013-092576.
Gibala MJ, Little JP, Macdonald MJ, Hawley JA. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J Physiol. 2012;590(5):1077–84. https://doi.org/10.1113/jphysiol.2011.224725.
Gibala M. Molecular responses to high-intensity interval exercise. Appl Physiol Nutr Metab (Physiologie appliquee, nutrition et metabolisme). 2009;34(3):428–32. https://doi.org/10.1139/h09-046.
Batacan RB, Duncan MJ, Dalbo VJ, Tucker PS, Fenning AS. Effects of high-intensity interval training on cardiometabolic health: a systematic review and meta-analysis of intervention studies. Br J Sports Med. 2017;51(6):494–503. https://doi.org/10.1136/bjsports-2015-095841.
Cassidy S, Thoma C, Houghton D, Trenell MI. High-intensity interval training: a review of its impact on glucose control and cardiometabolic health. Diabetologia. 2017;60(1):7–23. https://doi.org/10.1007/s00125-016-4106-1.
Wisloff U, Stoylen A, Loennechen JP, Bruvold M, Rognmo O, Haram PM, et al. Superior cardiovascular effect of aerobic interval training versus moderate continuous training in heart failure patients: a randomized study. Circulation. 2007;115(24):3086–94. https://doi.org/10.1161/circulationaha.106.675041.
Rognmo O, Hetland E, Helgerud J, Hoff J, Slordahl SA. High intensity aerobic interval exercise is superior to moderate intensity exercise for increasing aerobic capacity in patients with coronary artery disease. Eur J Cardiovasc Prev Rehabil. 2004;11(3):216–22.
Moholdt TT, Amundsen BH, Rustad LA, Wahba A, Lovo KT, Gullikstad LR, et al. Aerobic interval training versus continuous moderate exercise after coronary artery bypass surgery: a randomized study of cardiovascular effects and quality of life. Am Heart J. 2009;158(6):1031–7. https://doi.org/10.1016/j.ahj.2009.10.003.
Robinson MM, Dasari S, Konopka AR, Johnson ML, Manjunatha S, Esponda RR, et al. Enhanced protein translation underlies improved metabolic and physical adaptations to different exercise training modes in young and old humans. Cell Metab. 2017;25(3):581–92. https://doi.org/10.1016/j.cmet.2017.02.009.
Wyckelsma VL, Levinger I, McKenna MJ. Preservation of skeletal muscle mitochondrial content in older adults: relationship between mitochondria, fibre type and high-intensity exercise training. J Physiol. 2017;595(11):3345–59. https://doi.org/10.1113/jp273950.
Joanisse S, Gillen JB, Bellamy LM, McKay BR, Tarnopolsky MA, Gibala MJ, et al. Evidence for the contribution of muscle stem cells to nonhypertrophic skeletal muscle remodeling in humans. FASEB J. 2013;27(11):4596–605. https://doi.org/10.1096/fj.13-229799.
Blue MNM, Smith-Ryan AE, Trexler ET, Hirsch KR. The effects of high intensity interval training on muscle size and quality in overweight and obese adults. J Sci Med Sport. 2017. https://doi.org/10.1016/j.jsams.2017.06.001.
Hood MS, Little JP, Tarnopolsky MA, Myslik F, Gibala MJ. Low-volume interval training improves muscle oxidative capacity in sedentary adults. Med Sci Sports Exerc. 2011;43(10):1849–56. https://doi.org/10.1249/MSS.0b013e3182199834.
Little JP, Gillen JB, Percival ME, Safdar A, Tarnopolsky MA, Punthakee Z, et al. Low-volume high-intensity interval training reduces hyperglycemia and increases muscle mitochondrial capacity in patients with type 2 diabetes. J Appl Physiol (1985). 2011;111(6):1554–60. https://doi.org/10.1152/japplphysiol.00921.2011.
Tabata I, Nishimura K, Kouzaki M, Hirai Y, Ogita F, Miyachi M, et al. Effects of moderate-intensity endurance and high-intensity intermittent training on anaerobic capacity and VO2max. Med Sci Sports Exerc. 1996;28(10):1327–30.
Joanisse S, McKay BR, Nederveen JP, Scribbans TD, Gurd BJ, Gillen JB, et al. Satellite cell activity, without expansion, after nonhypertrophic stimuli. Am J Physiol Regul Integr Comp Physiol. 2015;309(9):R1101–11. https://doi.org/10.1152/ajpregu.00249.2015.
Scribbans TD, Edgett BA, Vorobej K, Mitchell AS, Joanisse SD, Matusiak JBL, et al. Fibre-specific responses to endurance and low volume high intensity interval training: striking similarities in acute and chronic adaptation. PLoS ONE. 2014;9(6):e98119. https://doi.org/10.1371/journal.pone.0098119.
Burgomaster KA, Howarth KR, Phillips SM, Rakobowchuk M, Macdonald MJ, McGee SL, et al. Similar metabolic adaptations during exercise after low volume sprint interval and traditional endurance training in humans. J Physiol. 2008;586(1):151–60. https://doi.org/10.1113/jphysiol.2007.142109.
Granata C, Oliveira RS, Little JP, Renner K, Bishop DJ. Training intensity modulates changes in pgc-1alpha and p53 protein content and mitochondrial respiration, but not markers of mitochondrial content in human skeletal muscle. FASEB J. 2016;30(2):959–70. https://doi.org/10.1096/fj.15-276907.
Gibala MJ, Little JP, van Essen M, Wilkin GP, Burgomaster KA, Safdar A, et al. Short-term sprint interval versus traditional endurance training: similar initial adaptations in human skeletal muscle and exercise performance. J Physiol. 2006;575(Pt 3):901–11. https://doi.org/10.1113/jphysiol.2006.112094.
Scalzo RL, Peltonen GL, Binns SE, Shankaran M, Giordano GR, Hartley DA, et al. Greater muscle protein synthesis and mitochondrial biogenesis in males compared with females during sprint interval training. FASEB J. 2014;28(6):2705–14. https://doi.org/10.1096/fj.13-246595.
Metcalfe RS, Babraj JA, Fawkner SG, Vollaard NB. Towards the minimal amount of exercise for improving metabolic health: beneficial effects of reduced-exertion high-intensity interval training. Eur J Appl Physiol. 2012;112(7):2767–75. https://doi.org/10.1007/s00421-011-2254-z.
Metcalfe RS, Tardif N, Thompson D, Vollaard NB. Changes in aerobic capacity and glycaemic control in response to reduced-exertion high-intensity interval training (rehit) are not different between sedentary men and women. Appl Physiol Nutr Metab. 2016;41(11):1117–23. https://doi.org/10.1139/apnm-2016-0253.
Gillen JB, Martin BJ, MacInnis MJ, Skelly LE, Tarnopolsky MA, Gibala MJ. Twelve weeks of sprint interval training improves indices of cardiometabolic health similar to traditional endurance training despite a five-fold lower exercise volume and time commitment. PLoS ONE. 2016;11(4):e0154075. https://doi.org/10.1371/journal.pone.0154075.
Sogaard D, Lund MT, Scheuer CM, Dehlbaek MS, Dideriksen SG, Abildskov CV, et al. High-intensity interval training improves insulin sensitivity in older individuals. Acta Physiol (Oxf). 2017. https://doi.org/10.1111/apha.13009.
Hwang CL, Yoo JK, Kim HK, Hwang MH, Handberg EM, Petersen JW, et al. Novel all-extremity high-intensity interval training improves aerobic fitness, cardiac function and insulin resistance in healthy older adults. Exp Gerontol. 2016;82:112–9. https://doi.org/10.1016/j.exger.2016.06.009.
Mejías-Peña Y, Rodriguez-Miguelez P, Fernandez-Gonzalo R, Martínez-Flórez S, Almar M, de Paz JA, et al. Effects of aerobic training on markers of autophagy in the elderly. Age (Dordr). 2016;38(2):33. https://doi.org/10.1007/s11357-016-9897-y.
Stensvold D, Viken H, Steinshamn SL, Dalen H, Støylen A, Loennechen JP, et al. Effect of exercise training for five years on all cause mortality in older adults—the generation 100 study: randomised controlled trial. BMJ. 2020;371:m3485. https://doi.org/10.1136/bmj.m3485.
Kovacevic A, Fenesi B, Paolucci E, Heisz JJ. The effects of aerobic exercise intensity on memory in older adults. Appl Physiol Nutr Metab. 2020;45(6):591–600. https://doi.org/10.1139/apnm-2019-0495.
Kim HK, Hwang CL, Yoo JK, Hwang MH, Handberg EM, Petersen JW, et al. All-extremity exercise training improves arterial stiffness in older adults. Med Sci Sports Exerc. 2017;49(7):1404–11. https://doi.org/10.1249/mss.0000000000001229.
Oliveira BRR, Santos TM, Kilpatrick M, Pires FO, Deslandes AC. Affective and enjoyment responses in high intensity interval training and continuous training: a systematic review and meta-analysis. PLoS ONE. 2018;13(6):e0197124. https://doi.org/10.1371/journal.pone.0197124.
Martinez-Valdes E, Falla D, Negro F, Mayer F, Farina D. Differential motor unit changes after endurance or high-intensity interval training. Med Sci Sports Exerc. 2017;49(6):1126–36. https://doi.org/10.1249/mss.0000000000001209.
Martinez-Valdes E, Farina D, Negro F, Del Vecchio A, Falla D. Early motor unit conduction velocity changes to high-intensity interval training versus continuous training. Med Sci Sports Exerc. 2018;50(11):2339–50. https://doi.org/10.1249/mss.0000000000001705.
Krustrup P, Soderlund K, Mohr M, Gonzalez-Alonso J, Bangsbo J. Recruitment of fibre types and quadriceps muscle portions during repeated, intense knee-extensor exercise in humans. Pflugers Arch. 2004;449(1):56–65. https://doi.org/10.1007/s00424-004-1304-3.
Edgett BA, Foster WS, Hankinson PB, Simpson CA, Little JP, Graham RB, et al. Dissociation of increases in pgc-1alpha and its regulators from exercise intensity and muscle activation following acute exercise. PLoS ONE. 2013;8(8):e71623. https://doi.org/10.1371/journal.pone.0071623.
Folland JP, Williams AG. The adaptations to strength training : morphological and neurological contributions to increased strength. Sports Med. 2007;37(2):145–68.
Duchateau J, Semmler JG, Enoka RM. Training adaptations in the behavior of human motor units. J Appl Physiol (1985). 2006;101(6):1766–75. https://doi.org/10.1152/japplphysiol.00543.2006.
Camera DM, Smiles WJ, Hawley JA. Exercise-induced skeletal muscle signaling pathways and human athletic performance. Free Radic Biol Med. 2016;98:131–43. https://doi.org/10.1016/j.freeradbiomed.2016.02.007.
Rundqvist HC, Montelius A, Osterlund T, Norman B, Esbjornsson M, Jansson E. Acute sprint exercise transcriptome in human skeletal muscle. PLoS ONE. 2019;14(10):e0223024. https://doi.org/10.1371/journal.pone.0223024.
Miyamoto-Mikami E, Tsuji K, Horii N, Hasegawa N, Fujie S, Homma T, et al. Gene expression profile of muscle adaptation to high-intensity intermittent exercise training in young men. Sci Rep. 2018;8(1):16811. https://doi.org/10.1038/s41598-018-35115-x.
Dutka TL, Lamboley CR, McKenna MJ, Murphy RM, Lamb GD. Effects of carnosine on contractile apparatus Ca(2)(+) sensitivity and sarcoplasmic reticulum Ca(2)(+) release in human skeletal muscle fibers. J Appl Physiol. 2012;112(5):728–36. https://doi.org/10.1152/japplphysiol.01331.2011.
Kamm KE, Stull JT. Dedicated myosin light chain kinases with diverse cellular functions. J Biol Chem. 2001;276(7):4527–30. https://doi.org/10.1074/jbc.R000028200.
Gibala MJ, Hawley JA. Sprinting toward fitness. Cell Metab. 2017;25(5):988–90. https://doi.org/10.1016/j.cmet.2017.04.030.
Goody MF, Sher RB, Henry CA. Hanging on for the ride: adhesion to the extracellular matrix mediates cellular responses in skeletal muscle morphogenesis and disease. Dev Biol. 2015;401(1):75–91. https://doi.org/10.1016/j.ydbio.2015.01.002.
Hjorth M, Norheim F, Meen AJ, Pourteymour S, Lee S, Holen T, et al. The effect of acute and long-term physical activity on extracellular matrix and serglycin in human skeletal muscle. Physiol Rep. 2015. https://doi.org/10.14814/phy2.12473.
Wilkinson SB, Phillips SM, Atherton PJ, Patel R, Yarasheski KE, Tarnopolsky MA, et al. Differential effects of resistance and endurance exercise in the fed state on signalling molecule phosphorylation and protein synthesis in human muscle. J Physiol. 2008;586(15):3701–17. https://doi.org/10.1113/jphysiol.2008.153916.
Dreyer HC, Fujita S, Glynn EL, Drummond MJ, Volpi E, Rasmussen BB. Resistance exercise increases leg muscle protein synthesis and mtor signalling independent of sex. Acta Physiol (Oxf). 2010;199(1):71–81. https://doi.org/10.1111/j.1748-1716.2010.02074.x.
Short KR, Vittone JL, Bigelow ML, Proctor DN, Nair KS. Age and aerobic exercise training effects on whole body and muscle protein metabolism. Am J Physiol Endocrinol Metab. 2004;286(1):E92-101. https://doi.org/10.1152/ajpendo.00366.2003.
Bell KE, Seguin C, Parise G, Baker SK, Phillips SM. Day-to-day changes in muscle protein synthesis in recovery from resistance, aerobic, and high-intensity interval exercise in older men. J Gerontol A Biol Sci Med Sci. 2015;70(8):1024–9. https://doi.org/10.1093/gerona/glu313.
Brook MS, Wilkinson DJ, Mitchell WK, Lund JN, Szewczyk NJ, Greenhaff PL, et al. Skeletal muscle hypertrophy adaptations predominate in the early stages of resistance exercise training, matching deuterium oxide-derived measures of muscle protein synthesis and mechanistic target of rapamycin complex 1 signaling. FASEB J. 2015;29(11):4485–96. https://doi.org/10.1096/fj.15-273755.
Mitchell CJ, Churchward-Venne TA, Parise G. Acute post-exercise myofibrillar protein synthesis is not correlated with resistance training-induced muscle hypertrophy in young men. PLoS ONE. 2014. https://doi.org/10.1371/journal.pone.0089431.
Mayhew DL, Kim JS, Cross JM, Ferrando AA, Bamman MM. Translational signaling responses preceding resistance training-mediated myofiber hypertrophy in young and old humans. J Appl Physiol (1985). 2009;107(5):1655–62. https://doi.org/10.1152/japplphysiol.91234.2008.
Damas F, Phillips SM, Libardi CA, Vechin FC, Lixandrao ME, Jannig PR, et al. Resistance training-induced changes in integrated myofibrillar protein synthesis are related to hypertrophy only after attenuation of muscle damage. J Physiol. 2016;594(18):5209–22. https://doi.org/10.1113/jp272472.
Damas F, Phillips SM, Lixandrao ME, Vechin FC, Libardi CA, Roschel H, et al. Early resistance training-induced increases in muscle cross-sectional area are concomitant with edema-induced muscle swelling. Eur J Appl Physiol. 2016;116(1):49–56. https://doi.org/10.1007/s00421-015-3243-4.
Joanisse S, Snijders T, Nederveen JP, Parise G. The impact of aerobic exercise on the muscle stem cell response. Exerc Sport Sci Rev. 2018;46(3):180–7. https://doi.org/10.1249/jes.0000000000000153.
Nederveen JP, Joanisse S, Seguin CM, Bell KE, Baker SK, Phillips SM, et al. The effect of exercise mode on the acute response of satellite cells in old men. Acta Physiol (Oxf). 2015;215(4):177–90. https://doi.org/10.1111/apha.12601.
Nederveen JP, Joanisse S, Snijders T, Ivankovic V, Baker SK, Phillips SM, et al. Skeletal muscle satellite cells are located at a closer proximity to capillaries in healthy young compared with older men. J Cachexia Sarcopenia Muscle. 2016;7(5):547–54. https://doi.org/10.1002/jcsm.12105.
Khacho M, Slack RS. Mitochondrial activity in the regulation of stem cell self-renewal and differentiation. Curr Opin Cell Biol. 2017;49:1–8. https://doi.org/10.1016/j.ceb.2017.11.003.
Abreu P, Kowaltowski AJ. Satellite cell self-renewal in endurance exercise is mediated by inhibition of mitochondrial oxygen consumption. J Cachexia Sarcopenia Muscle. 2020. https://doi.org/10.1002/jcsm.12601.
Charifi N, Kadi F, Feasson L, Denis C. Effects of endurance training on satellite cell frequency in skeletal muscle of old men. Muscle Nerve. 2003;28(1):87–92. https://doi.org/10.1002/mus.10394.
Verney J, Kadi F, Charifi N, Feasson L, Saafi MA, Castells J, et al. Effects of combined lower body endurance and upper body resistance training on the satellite cell pool in elderly subjects. Muscle Nerve. 2008;38(3):1147–54. https://doi.org/10.1002/mus.21054.
Snijders T, Nederveen JP, Bell KE, Lau SW, Mazara N, Kumbhare DA, et al. Prolonged exercise training improves the acute type ii muscle fibre satellite cell response in healthy older men. J Physiol. 2019;597(1):105–19. https://doi.org/10.1113/jp276260.
Sultana RN, Sabag A, Keating SE, Johnson NA. The effect of low-volume high-intensity interval training on body composition and cardiorespiratory fitness: a systematic review and meta-analysis. Sports Med. 2019. https://doi.org/10.1007/s40279-019-01167-w.
Bagley L, Slevin M, Bradburn S, Liu D, Murgatroyd C, Morrissey G, et al. Sex differences in the effects of 12 weeks sprint interval training on body fat mass and the rates of fatty acid oxidation and VO2max during exercise. BMJ Open Sport Exerc Med. 2016;2(1):e000056. https://doi.org/10.1136/bmjsem-2015-000056.
Bruseghini P, Calabria E, Tam E, Milanese C, Oliboni E, Pezzato A, et al. Effects of eight weeks of aerobic interval training and of isoinertial resistance training on risk factors of cardiometabolic diseases and exercise capacity in healthy elderly subjects. Oncotarget. 2015;6(19):16998–7015. https://doi.org/10.18632/oncotarget.4031.
Cassidy S, Thoma C, Hallsworth K, Parikh J, Hollingsworth KG, Taylor R, et al. High intensity intermittent exercise improves cardiac structure and function and reduces liver fat in patients with type 2 diabetes: a randomised controlled trial. Diabetologia. 2016;59(1):56–66. https://doi.org/10.1007/s00125-015-3741-2.
Dohlmann TL, Hindso M, Dela F, Helge JW, Larsen S. High-intensity interval training changes mitochondrial respiratory capacity differently in adipose tissue and skeletal muscle. Physiol Rep. 2018;6(18):e13857. https://doi.org/10.14814/phy2.13857.
Gahreman D, Heydari M, Boutcher Y, Freund J, Boutcher S. The effect of green tea ingestion and interval sprinting exercise on the body composition of overweight males: a randomized trial. Nutrients. 2016. https://doi.org/10.3390/nu8080510.
Gillen JB, Percival ME, Ludzki A, Tarnopolsky MA, Gibala MJ. Interval training in the fed or fasted state improves body composition and muscle oxidative capacity in overweight women. Obesity (Silver Spring). 2013;21(11):2249–55. https://doi.org/10.1002/oby.20379.
Heydari M, Freund J, Boutcher SH. The effect of high-intensity intermittent exercise on body composition of overweight young males. J Obes. 2012;2012:480467. https://doi.org/10.1155/2012/480467.
Kong Z, Sun S, Liu M, Shi Q. Short-term high-intensity interval training on body composition and blood glucose in overweight and obese young women. J Diabetes Res. 2016;2016:4073618. https://doi.org/10.1155/2016/4073618.
Maillard F, Rousset S, Pereira B, Traore A, de Pradel Del Amaze P, Boirie Y, et al. High-intensity interval training reduces abdominal fat mass in postmenopausal women with type 2 diabetes. Diabetes Metab. 2016;42(6):433–41. https://doi.org/10.1016/j.diabet.2016.07.031.
Sawyer BJ, Tucker WJ, Bhammar DM, Ryder JR, Sweazea KL, Gaesser GA. Effects of high-intensity interval training and moderate-intensity continuous training on endothelial function and cardiometabolic risk markers in obese adults. J Appl Physiol (1985). 2016;121(1):279–88. https://doi.org/10.1152/japplphysiol.00024.2016.
Sculthorpe NF, Herbert P, Grace F. One session of high-intensity interval training (hiit) every 5 days, improves muscle power but not static balance in lifelong sedentary ageing men: a randomized controlled trial. Medicine (Baltimore). 2017;96(6):e6040. https://doi.org/10.1097/md.0000000000006040.
Trapp EG, Chisholm DJ, Freund J, Boutcher SH. The effects of high-intensity intermittent exercise training on fat loss and fasting insulin levels of young women. Int J Obes (Lond). 2008;32(4):684–91. https://doi.org/10.1038/sj.ijo.0803781.
Ziemann E, Grzywacz T, Luszczyk M, Laskowski R, Olek RA, Gibson AL. Aerobic and anaerobic changes with high-intensity interval training in active college-aged men. J Strength Cond Res. 2011;25(4):1104–12. https://doi.org/10.1519/JSC.0b013e3181d09ec9.
Ravnholt T, Tybirk J, Jorgensen NR, Bangsbo J. High-intensity intermittent “5-10-15” running reduces body fat, and increases lean body mass, bone mineral density, and performance in untrained subjects. Eur J Appl Physiol. 2018;118(6):1221–30. https://doi.org/10.1007/s00421-018-3851-x.
Stensvold D, Tjonna AE, Skaug EA, Aspenes S, Stolen T, Wisloff U, et al. Strength training versus aerobic interval training to modify risk factors of metabolic syndrome. J Appl Physiol (1985). 2010;108(4):804–10. https://doi.org/10.1152/japplphysiol.00996.2009.
Brown EC, Hew-Butler T, Marks CRC, Butcher SJ, Choi MD. The impact of different high-intensity interval training protocols on body composition and physical fitness in healthy young adult females. Biores Open Access. 2018;7(1):177–85. https://doi.org/10.1089/biores.2018.0032.
Osawa Y, Azuma K, Tabata S, Katsukawa F, Ishida H, Oguma Y, et al. Effects of 16-week high-intensity interval training using upper and lower body ergometers on aerobic fitness and morphological changes in healthy men: a preliminary study. Open Access J Sports Med. 2014;5:257–65. https://doi.org/10.2147/oajsm.s68932.
Fex A, Leduc-Gaudet JP, Filion ME, Karelis AD, Aubertin-Leheudre M. Effect of elliptical high intensity interval training on metabolic risk factor in pre- and type 2 diabetes patients: a pilot study. J Phys Act Health. 2015;12(7):942–6. https://doi.org/10.1123/jpah.2014-0123.
Macpherson RE, Hazell TJ, Olver TD, Paterson DH, Lemon PW. Run sprint interval training improves aerobic performance but not maximal cardiac output. Med Sci Sports Exerc. 2011;43(1):115–22. https://doi.org/10.1249/MSS.0b013e3181e5eacd.
Matsuo T, Saotome K, Seino S, Eto M, Shimojo N, Matsushita A, et al. Low-volume, high-intensity, aerobic interval exercise for sedentary adults: VO(2)max, cardiac mass, and heart rate recovery. Eur J Appl Physiol. 2014;114(9):1963–72. https://doi.org/10.1007/s00421-014-2917-7.
Nybo L, Sundstrup E, Jakobsen MD, Mohr M, Hornstrup T, Simonsen L, et al. High-intensity training versus traditional exercise interventions for promoting health. Med Sci Sports Exerc. 2010;42(10):1951–8. https://doi.org/10.1249/MSS.0b013e3181d99203.
Toombs RJ, Ducher G, Shepherd JA, De Souza MJ. The impact of recent technological advances on the trueness and precision of dxa to assess body composition. Obesity (Silver Spring). 2012;20(1):30–9. https://doi.org/10.1038/oby.2011.211.
Haun CT, Vann CG, Roberts BM, Vigotsky AD, Schoenfeld BJ, Roberts MD. A critical evaluation of the biological construct skeletal muscle hypertrophy: Size matters but so does the measurement. Front Physiol. 2019. https://doi.org/10.3389/fphys.2019.00247.
Nana A, Slater GJ, Stewart AD, Burke LM. Methodology review: Using dual-energy X-ray absorptiometry (dxa) for the assessment of body composition in athletes and active people. Int J Sport Nutr Exerc Metab. 2015;25(2):198–215. https://doi.org/10.1123/ijsnem.2013-0228.
Tavoian D, Ampomah K, Amano S, Law TD, Clark BC. Changes in dxa-derived lean mass and mri-derived cross-sectional area of the thigh are modestly associated. Sci Rep. 2019;9(1):10028. https://doi.org/10.1038/s41598-019-46428-w.
Mitsiopoulos N, Baumgartner RN, Heymsfield SB, Lyons W, Gallagher D, Ross R. Cadaver validation of skeletal muscle measurement by magnetic resonance imaging and computerized tomography. J Appl Physiol (1985). 1998;85(1):115–22. https://doi.org/10.1152/jappl.1998.85.1.115.
Estes RR, Malinowski A, Piacentini M, Thrush D, Salley E, Losey C, et al. The effect of high intensity interval run training on cross-sectional area of the vastus lateralis in untrained college students. Int J Exerc Sci. 2017;10(1):137–45.
Hawley JA, Burke LM, Phillips SM, Spriet LL. Nutritional modulation of training-induced skeletal muscle adaptations. J Appl Physiol (1985). 2011;110(3):834–45. https://doi.org/10.1152/japplphysiol.00949.2010.
Moore DR, Churchward-Venne TA, Witard O. Protein ingestion to stimulate myofibrillar protein synthesis requires greater relative protein intakes in healthy older versus younger men. J Gerontol A Biol Sci Med Sci. 2015. https://doi.org/10.1093/gerona/glu103.
Wilkinson DJ, Hossain T, Hill DS, Phillips BE, Crossland H, Williams J, et al. Effects of leucine and its metabolite beta-hydroxy-beta-methylbutyrate on human skeletal muscle protein metabolism. J Physiol. 2013;591(11):2911–23. https://doi.org/10.1113/jphysiol.2013.253203.
Dai JM, Yu MX, Shen ZY, Guo CY, Zhuang SQ, Qiu XS. Leucine promotes proliferation and differentiation of primary preterm rat satellite cells in part through mtorc1 signaling pathway. Nutrients. 2015;7(5):3387–400. https://doi.org/10.3390/nu7053387.
Biolo G, Tipton KD, Klein S, Wolfe RR. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol. 1997;273(1 Pt 1):E122–9.
Vliet SV, Beals JW, Martinez IG, Skinner SK, Burd NA. Achieving optimal post-exercise muscle protein remodeling in physically active adults through whole food consumption. Nutrients. 2018. https://doi.org/10.3390/nu10020224.
Phillips SM. The impact of protein quality on the promotion of resistance exercise-induced changes in muscle mass. Nutr Metab (Lond). 2016;13:64. https://doi.org/10.1186/s12986-016-0124-8.
Morton RW, Murphy KT, McKellar SR, Schoenfeld BJ, Henselmans M, Helms E, et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. Br J Sports Med. 2017. https://doi.org/10.1136/bjsports-2017-097608.
Cermak NM, Res PT, de Groot LC, Saris WH, van Loon LJ. Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: a meta-analysis. Am J Clin Nutr. 2012;96(6):1454–64. https://doi.org/10.3945/ajcn.112.037556.
Timmerman KL, Dhanani S, Glynn EL, Fry CS, Drummond MJ, Jennings K, et al. A moderate acute increase in physical activity enhances nutritive flow and the muscle protein anabolic response to mixed nutrient intake in older adults. Am J Clin Nutr. 2012;95(6):1403–12. https://doi.org/10.3945/ajcn.111.020800.
Breen L, Philp A, Witard OC, Jackman SR, Selby A, Smith K, et al. The influence of carbohydrate-protein co-ingestion following endurance exercise on myofibrillar and mitochondrial protein synthesis. J Physiol. 2011;589(Pt 16):4011–25. https://doi.org/10.1113/jphysiol.2011.211888.
Rowlands DS, Nelson AR, Phillips SM, Faulkner JA, Clarke J, Burd NA, et al. Protein-leucine fed dose effects on muscle protein synthesis after endurance exercise. Med Sci Sports Exerc. 2015;47(3):547–55. https://doi.org/10.1249/mss.0000000000000447.
Harber MP, Konopka AR, Jemiolo B, Trappe SW, Trappe TA, Reidy PT. Muscle protein synthesis and gene expression during recovery from aerobic exercise in the fasted and fed states. Am J Physiol Regul Integr Comp Physiol. 2010;299(5):R1254–62. https://doi.org/10.1152/ajpregu.00348.2010.
Coffey VG, Moore DR, Burd NA, Rerecich T, Stellingwerff T, Garnham AP, et al. Nutrient provision increases signalling and protein synthesis in human skeletal muscle after repeated sprints. Eur J Appl Physiol. 2011;111(7):1473–83. https://doi.org/10.1007/s00421-010-1768-0.
Mitchell CJ, Churchward-Venne TA, Cameron-Smith D, Phillips SM. What is the relationship between the acute muscle protein synthesis response and changes in muscle mass? J Appl Physiol (1985). 2015;118(4):495–7. https://doi.org/10.1152/japplphysiol.00609.2014.
Rundqvist HC, Esbjornsson M, Rooyackers O, Osterlund T, Moberg M, Apro W, et al. Influence of nutrient ingestion on amino acid transporters and protein synthesis in human skeletal muscle after sprint exercise. J Appl Physiol (1985). 2017. https://doi.org/10.1152/japplphysiol.00244.2017.
Leuchtmann AB, Mueller SM, Aguayo D, Petersen JA, Ligon-Auer M, Flück M, et al. Resistance training preserves high-intensity interval training induced improvements in skeletal muscle capillarization of healthy old men: a randomized controlled trial. Sci Rep. 2020;10(1):6578. https://doi.org/10.1038/s41598-020-63490-x.
Gaffney KA, Lucero A, Stoner L, Faulkner J, Whitfield P, Krebs J, et al. Nil whey protein effect on glycemic control after intense mixed-mode training in type 2 diabetes. Med Sci Sports Exerc. 2018;50(1):11–7. https://doi.org/10.1249/mss.0000000000001404.
Hickson RC. Interference of strength development by simultaneously training for strength and endurance. Eur J Appl Physiol Occup Physiol. 1980;45(2–3):255–63.
Esbjörnsson M, Rundqvist HC, Mascher H, Österlund T, Rooyackers O, Blomstrand E, et al. Sprint exercise enhances skeletal muscle p70s6k phosphorylation and more so in women than in men. Acta Physiol (Oxf). 2012;205(3):411–22. https://doi.org/10.1111/j.1748-1716.2012.02404.x.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
No sources of funding were used to assist in the preparation of this article.
Conflict of interest
Marcus Callahan, Evelyn Parr, John Hawley and Donny Camera declare that they have no conflicts of interest relevant to the content of this review.
Authorship contributions
MC wrote the first draft of the manuscript. MC, EBP, JH and DMC revised the original manuscript. All authors read and approved the final manuscript.
Rights and permissions
About this article

Cite this article
Callahan, M.J., Parr, E.B., Hawley, J.A. et al. Can High-Intensity Interval Training Promote Skeletal Muscle Anabolism?. Sports Med 51, 405–421 (2021). https://doi.org/10.1007/s40279-020-01397-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40279-020-01397-3