
Abstract
The ability of the human body to generate maximal power is linked to a host of performance outcomes and sporting success. Power-force-velocity relationships characterize limits of the neuromuscular system to produce power, and their measurement has been a common topic in research for the past century. Unfortunately, the narrative of the available literature is complex, with development occurring across a variety of methods and technology. This review focuses on the different equipment and methods used to determine mechanical characteristics of maximal exertion human sprinting. Stationary cycle ergometers have been the most common mode of assessment to date, followed by specialized treadmills used to profile the mechanical outputs of the limbs during sprint running. The most recent methods use complex multiple-force plate lengths in-ground to create a composite profile of over-ground sprint running kinetics across repeated sprints, and macroscopic inverse dynamic approaches to model mechanical variables during over-ground sprinting from simple time-distance measures during a single sprint. This review outlines these approaches chronologically, with particular emphasis on the computational theory developed and how this has shaped subsequent methodological approaches. Furthermore, training applications are presented, with emphasis on the theory underlying the assessment of optimal loading conditions for power production during resisted sprinting. Future implications for research, based on past and present methodological limitations, are also presented. It is our aim that this review will assist in the understanding of the convoluted literature surrounding mechanical sprint profiling, and consequently improve the implementation of such methods in future research and practice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Power-force-velocity relationships can be assessed during maximal sprinting using a variety of methods and technologies — from multiple trials performed on friction-braked cycle ergometers and specialised treadmills, to ‘simplified' techniques employing a single over ground trial measured via timing gates, radar, or even cellular devices. |
Although the direct development of mechanical profiling spans almost a century, the rapid expansion of these and other methods in recent years has led to limited data on modern equipment. |
While there is growing evidence to support the value of these techniques, future studies should look to collect normative data on highly trained cohorts and examine their usefulness in orienting and assessing training outcomes. |
1 Background
The ability of skeletal muscle to generate force and the maximal rate of movement is described in the force-velocity (Fv) relationship. The relationship postulates that for a given constant level of muscular activation, increasing shortening velocity progressively decreases the force produced by the neuromuscular system [1]. Mechanical power output (i.e. the rate of performing mechanical work), in this instance, is defined as the product of force and velocity. As the maximal abilities of skeletal muscle to generate both force and velocity are intertwined, the Fv relationship characterizes the ability to produce and maximize power. The term maximal power (\(P_{ \text{max} }\)) describes the peak combination of velocity and force achieved in a given muscular contraction, or movement task [2–4]. Fv and power-velocity (Pv) relationships (i.e. PFv) have been examined, in vitro and in vivo, to give insight into the mechanical determinants of performance and further our understanding of movement.
1.1 History of Force-Velocity Profiling
The first studies to report concepts of force, velocity and maximal work were based on theoretical methods derived from hydraulicians, where fluid within the muscle has a certain velocity, and any work performed (effort) is proportional to the square of velocity (\(v\)) [5]. The first experimental studies in this area [6, 7] showed that skeletal muscle performed similarly to other mechanical systems where increasing velocity resulted in decreasing work, and this work-time relationship corresponded to the force-time relationship [8]. A decade after these studies, an exponential function was developed from in vitro experimentation [9, 10], after which Hill [1] derived the well-known hyperbolic equation (Table 1; Eq. 1) which has been widely used in power-based sprint-cycling research [11, 12]. While Hill’s rectangular hyperbola accurately fits the data provided by many single-joint actions across differing testing procedures, this relationship does not describe external force production occurring during multi-joint actions [13, 14].
There are several modalities through which these characteristics are assessed in in vivo movement tasks: (1) control and manipulation of the force imposed on the movement, and measurement of velocity (isotonic) [15]; (2) control and manipulation of movement velocity and the subsequent measurement of force (isokinetic) [16]; and (3) control and manipulation of the external constraints (inertia and/or weight) and measurement of force and/or velocity (isoinertial) [17]. Regardless of the testing conditions, the effort presented is maximal given that the goal is to determine the mechanical limit of the neuromuscular system. Isoinertial experiments are most common as they best represent the natural movement patterns found in sporting contexts, and generally represent a less costly and complex alternative to isokinetic and isotonic modalities. Typically, in an isoinertial experiment external loading conditions are manipulated and the responses of the dependent variables of force and/or velocity are measured across single or multiple trials.
1.2 Characterisation of Mechanical Capacity in Multi-Joint Tasks
While non-linear relationships are typically observed in Fv profiles with individual joints or muscle fibres [10], multi-joint tasks appear to present quasi-linear relationships. Notably, the Fv relationships observed in multiple joint movements are a product of complex interactions between muscular coordination characteristics, activation patterns, the anatomy of joints, and the orientation of moments occurring (among others) (for an extensive recent review see Jaric [14]). While neural mechanisms were considered to primarily cause these observations [15, 18, 19], recent evidence suggests segmental dynamics may instead be the main determinant of linear Fv profiles in these tasks, with each joint progressively impeding muscular production of force with increasing velocity, thus decreasing external force [13, 20]. Inverse linear Fv and parabolic power-velocity (Pv) relationships have subsequently been used in recent practice to describe the mechanical capabilities of the neuromuscular system during a range of multi-joint lower-limb movements (for a detailed review of these methods see Soriano et al. [21]): primarily variations on jumping and similar acyclic extensions of the lower limbs [15, 22–29].
Changes in external force production across varying movement velocities are described by PFv relationships, and are most commonly displayed by the following three variables in literature: the theoretical maximum force the system can produce at zero velocity (\(F_{0}\)); the theoretical maximum velocity at which the system can contract/extend at zero force (\(v_{0}\)); and the \(P_{ \text{max} }\) the system can produce (either during cyclic or acyclic movements). \(F_{0}\) and \(v_{0}\) represent the y and x intercepts of the linear regression, respectively. \(P_{ \text{max} }\) corresponds to the apex of the parabolic Pv relationship and can be computed directly from \(F_{0}\) and \(v_{0}\) for linear Fv relationships: \(P_{ \text{max} } = \left( {F_{0} {\cdot} v_{0} } \right)/4\) (see Table 1; Eq. 5) [28, 30]. Principally, these variables characterize the maximal mechanical abilities of the total system (depending on the movement and definition used in each circumstance) pertaining to the generation of mechanical capacities. As the relationship between these macroscopic variables encompasses the entire capability of the neuromuscular system, it is inclusive of mechanical properties of individual muscles (e.g. rate of force development, and internal Fv and length-tension relationships), morphological features (e.g. muscle architecture and tendon characteristics), neural mechanisms underpinning motor-unit drive (e.g. motor unit recruitment, synchronization, firing frequency, and coordination between muscles), and segmental dynamics [13, 18, 31–33]. The Fv mechanical profile during explosive lower limb movements can be described by the ratio between \(F_{0}\) and \(v_{0}\), or the slope of the linear regression fit for the Fv relationship (\(S_{\text{Fv}}\)) when force is displayed on the x-axis [28]. Additional variables of interest are the combination of force and velocity that elicit \(P_{ \text{max} }\), often classified in literature as the ‘optimal’ level for maximal power (\(F_{\text{opt}}\) and \(v_{\text{opt}}\), respectively) [34]. These variables are of particular interest to practitioners as training implemented under a loading scheme representative of \(F_{\text{opt}}\) and \(v_{\text{opt}}\) (i.e. \(L_{\text{opt}}\)) may acutely and longitudinally improve the capacity of the system for maximal power production [35].
2 Mechanical Profiling in Sprint Cycling
2.1 Multiple-Trial Mechanical Profiling with Cyclic Cranks
To our knowledge, assessments of the Fv relationship using cycling ergometry began as early as 1928 with the experiments of Dickinson [36]. In this study, a mixed sex cohort of four subjects displayed a linear relationship between pedal rate and braking force, across increasing resistive loads on a friction-braked cycle ergometer. The researchers used a combination of spring balances to determine the application of braking to the rim of the wheel, calculated as a component of weight added to the device [37]. Despite the fact that the results obtained by Dickinson [36] are comparable to recent results [38], the focus of the article and subsequent release of the hyperbolic equation by Hill [1] limited their impact on the field of physical assessment.
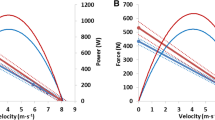
Modern attempts at profiling mechanical sprinting abilities originated based on a profiling method using a form of Monark cyclic cranks redesigned for use with the upper body [39] (for extensive information regarding cyclic ergometers, see the recent detailed reviews of Vandewalle and Driss [37] and Driss and Vandewalle [38]. The assessment protocol comprised of eight to ten sprints performed against progressively increasing braking forces (\(F\); +1 kg per trial) with consideration of a curvi-linear Pv relationship [40]. At each load, peak velocity (\(v_{ \text{max} }\)) was measured and plotted against the braking load applied (see Fig. 1). Given that at \(v_{ \text{max} }\) the force developed by the limbs is equal to the braking force (assuming zero acceleration), the linear Fv relationship can be plotted under a least-squares regression (see Table 1; Eq. 2) accounting for braking force and \(v_{ \text{max} }\) alone [40]. \(v_{0}\) and \(F_{0}\) were determined by the intercepts of the velocity and force axis respectively, with \(P_{ \text{max} }\) referring to the optimal combination between velocity \((0.5{\cdot} v_{0} )\) and a load (\(0.5{\cdot} F_{0}\); Eq. 3). These methods were later used to characterize lower body kinetics with the development of a new ergometer (Model: 864, Monark-Crescent AB, Varberg, Sweden) featuring higher load tolerances [30, 41], and a reduced volume of trials than these early studies (five to seven trials) due to the Fv relationship’s linearity. Similarly, \(P_{ \text{max} }\) of the lower limbs was determined as \(0.25{\cdot} v_{0} {\cdot} F_{0}\). The first methods to profile Fv characteristics on cycle ergometry only accounted for the force required to overcome resistive force against the flywheel inertia (i.e. \(F\)), and not that required to accelerate it. During the mid–late 1980s researchers [42–44] proposed a ‘corrected’ approach that considered the force required to accelerate the flywheel (\(F_{\text{acc}}\)) in addition to \(F\), to determine a corrected force (\(F_{\text{corr}}\)) (Eq. 4). Power-output per crank revolution (\(P_{\text{rev}}\)) was calculated as the combination of velocity per revolution (\(v_{\text{rev}}\)) and \(F_{\text{corr}}\), with its maximum value during acceleration described in corrected peak-power (\(PP_{\text{corr}}\)). However, caution should be exercised when interpreting the results of Lakomy [43] because of a small and mixed group of participants (five males and five females) and changes in sampling intervals [45].

Graphic representation of the relationship between force-velocity and power-velocity as profiled using a multiple-sprint method on a treadmill ergometer. Note that graphically the same relationship can be determined from cycle ergometry, but with torque (N·m) against velocity (rad·s−1). Each data point represents values derived from a single point during an individual trial at different loading or braking protocols. \(F_{0}\) and \(v_{0}\) represent the y and x intercepts of the linear regression, and the theoretical maximum of force and velocity able to be produced in the absence of their opposing unit. \(P_{\text{max} }\) represents the maximum power produced, determined as the peak of the polynomial fit between power and velocity
2.2 Single-Trial Profiling Method with Cycle Ergometers
Researchers realized that that once acceleration of the flywheel was accounted for, and with technology featuring a high enough sample rate, it was possible to plot instantaneous decreasing force production (inertial and friction force) with increasing velocity during a single maximal acceleration bout [46]. Following calculation of flywheel inertia (see the corrected method in Sect. 2.1), the relationship between instantaneous angular crank velocity and the torque exerted on the crank (\(T\)) were measured during a single maximal cycle sprint (Eqs. 6–8). Variables were assessed via a photoelectric cell measuring impulse bursts from the flywheel up to \(v_{ \text{max} }\), with \(T\) calculated as the combination of torque required for acceleration and torque to overcome the braking load (see Fig. 2) [43, 44]. Importantly, the torque-velocity relationship determined via this single trial method was similarly linear, as compared to the multiple-trial method. When comparing the multiple-trial method to the newly developed single trial method, there was no statistical difference between peak power metrics [i.e. \(P_{\text{corr}}\) vs. \(P_{ \text{max} } 2\) (determined as \(0.25\omega_{0} T_{0}\))]. Both variables were expectedly ~10% higher than the uncorrected \(P_{ \text{max} }\) determined from multiple trials, which is likely a product of fatigue, reminiscent of accelerating to a later \(v_{ \text{max} }\) [38, 47]. The usefulness of power correction was corroborated by Morin and Belli [48], who reported underestimation of ~20.4% for power without accounting for the effect of inertia; however, it should be noted that when computed from the Fv relationship, the errors are likely lower.

Graphic representation of force- and torque-velocity relationships determined from a single trial. Torque-velocity determined via a single-sprint method on a cycle ergometer. The method displayed describes linear regression determined from the peak of each cycle rotation, from the first down-stroke (First), to the last (Last). \(T_{0}\) and \(v_{0}\) represent the theoretical maximum of torque and velocity, determined via the y an x intercepts of the linear torque-velocity regression, respectively
As methods progressed, variables were averaged over each pedal down-stroke [19, 49, 50] rather than over non-descript periods of time. This was a biomechanically sound progression, given that the data now represented what the lower limbs could develop over one extension – similar to a squat, leg press, sprinting on dynamometric treadmill or force plate system in the ground. It should be noted, however, that power output during cycling does not only correspond to the pedal down stroke (when feet are strapped into the pedals), but instead is produced throughout each cycle with athletes pulling up against the pedal in combination with pushing down [51, 52]. Developments in data collection technology allowed for more exacting assessments of torque during cycle sprinting. For example, Buttelli et al. [53] were the first to measure the torque exerted over each pedal revolution during an all-out sprint instead of computing the torque from the acceleration of the flywheel by using a form of strain gauges bonded to the cranks of an electric ergometer. Buttelli et al. [53] further demonstrated that peak torque occurred later in the crank cycle when pedalling rate increased, which was confirmed later by Samozino et al. [19]. Furthermore, research has implemented similar analyses of power and effectiveness of force application, defined as the magnitude of effective force perpendicular to the crank expressed as a percentage of total force production [54]. This perpendicular oriented force vector is alone necessary to rotate the drive, and consequently has been related to mechanical efficiency [55] and positive pedalling technique [52]. Notably, Dorel et al. [52] clearly showed that Fv relationships are largely affected by the mechanical effectiveness of the force production onto the pedals, and the decrease in power output beyond the optimal pedalling rate can be partly explained by an important decrease in pedal effectiveness. These analyses are of note, and their impact will be discussed further with reference to its calculation and importance during sprint running.
3 Mechanical Profiling During Treadmill Sprint Running
3.1 The Beginnings of Mechanical Profiling of Sprint Running
In order to increase assessment specificity, researchers have endeavoured to assess mechanical capabilities during sprint running. To our knowledge, Furusawa et al. [56] performed the first published experiments to quantify acceleration of track and field sprinters using a system of wire coils, set at regular known intervals along a testing track, connected to a galvanometer [57]. The subject was equipped with a magnetized harness that recorded a deflection as each coil was passed during the sprint. As the distance (\(d\)) between each coil was known (1–10 yards), velocity was simply calculated as \(v = \Delta d/\Delta t\), and acceleration as \(a = \Delta v/\Delta t\), with time measured between each ping from the galvanometer (with a resolution of 5 ms). The first available experimental data concerning Fv relationships during bipedal load-bearing sprinting were derived from an experiment by Best and Partridge [8], based on the earlier work by Furusawa et al. [56]. This study used the same equipment with the addition of a spectrograph split to increase the accuracy of deflection measurement, and a customized tethered winch system to provide a constant external resistance to the athlete. The experiment effectively confirmed theories of the effects of internal resistance and viscosity of muscle impeding velocity production, and that these could be compared to the inhibiting effects of the external resistance provided in their study. Moreover, the study also noted the similarity of the results to work on air resistance [56], highlighting that the equations for estimating velocity decrement were accurate, with the exception of the application point of resistance (i.e. around the waist, in comparison to the whole body). The study has subsequently been replicated using updated technology in recent years [58].
Modern attempts at experimentally determining mechanical characteristics of the body during load-bearing sprinting commonly use specialized sprint treadmill ergometry. This method requires subjects to propel a treadmill belt while tethered around the waist to an immovable stationary point at the rear of the machine. These ergometers are either motorized [48, 59–62], with the motor set to apply a resistant torque to compensate for the friction of the treadmill track under the bodyweight of the subject, or non-motorized [63–68], with the track simply mounted on low-friction rollers.
3.2 Multiple-Trial Methods Using Treadmill Ergometry
To our knowledge, the first author to publish direct measurements of sprint running kinetics was Lakomy [67], who used a combination of two tests previously proposed by Dal Monte and Leonardi [69] and Cheetham et al. [65]. Of note, the experiments presented by Dal Monte and Leonardi [69] featured the assessment of kinetics during load-bearing running, albeit without restricted arm movement due to the need to push against a bar to drive the treadmill belt. Consequently, Lakomy [67] used an early non-motorized treadmill (Woodway model AB, Germany) to show that power, horizontal force (\(F_{\text{h}}\)) and velocity (\(v_{\text{h}}\)) could be accurately measured during a 7-s maximal sprint. While the authors did not attempt to profile Fv relationships from the dataset, they did confirm collection of these variables was possible, and therefore paved the way for future investigation. Jaskólska et al. [61, 70] showed that a multiple-trial method could be used to accurately profile PFv relationships on a sprint-specialized motorized treadmill ergometer (Model: Gymroll 1800, Gymroll, Roche la Molière, France) (see Fig. 1 for an example of the multiple sprint method). To provide resistance for each loading condition the track motors were set to apply braking, as a percentage of a predetermined maximum value (i.e. ~1351 N) [61, 70, 71], to backward movement of the treadmill belt. Resistance was increased across six sprints (68, 108, 135, 176, 203 and 270 N), during which \(F_{\text{h}}\) was estimated via a tether-mounted force-transducer and goniometer (to correct for attachment angle, and separate vertically oriented forces), and \(v_{\text{h}}\) via a sensor system attached to the rear drum of the treadmill belt. Instantaneous power was calculated as the combination of horizontal force and belt velocity. These studies showed that not only were these measurement methods sensitive enough to determine differences between athletes of similar abilities, but the linear profile developed for each subject was independent of \(v_{ \text{max} }\) itself. Moreover, using various methods of variable sampling (instantaneous peak, greatest peak value assessed from 1-s averages, total mean and mean across 5 s), maximal power measures were shown to be reliable [intraclass correlation coefficient (ICC) = 0.80–0.89]. When comparing PFv relationships calculated from multiple sprints on cycle and treadmill ergometry, Jaskólska et al. [70] showed power indices were similar (r = 0.71–0.86; P < 0.01), albeit with lower readings on the treadmill attributed to the load-bearing nature of treadmill sprinting reducing maximal power output of the lower limbs. Furthermore, the study showed that athletes with a range of maximal speed abilities presented a linear Fv relationship when using the multiple-trial method, with high scores for individual correlation coefficients (R 2 > 0.989) and no significant difference with repeated measurement.
Since these original studies, modernized ‘two-dimensional’ sprint treadmills have been shown to provide reliable and accurate assessment of sprinting kinetics in various population groups [64, 72–76], although none have repeated the multiple trial PFv experiments of Jaskólska et al. [61]. There are, however, limitations inherent to treadmill-based sprinting assessment [67]. Earlier studies [48, 61, 63, 67, 77, 78] sampled instantaneous power values over non-descript brackets of time, often exceeding 1 s in duration, resulting in the inaccurate measurement of power and underestimation of velocity (among other errors) [44, 67]. While this limitation was often imposed by technology, force values should be averaged over distinct time periods relating to muscular events in the interest of gaining a holistic view of power production specific to step-cycles during sprint acceleration [79]. Although this error has typically been avoided in recent studies, with high sampling frequencies allowing averaging across definite time-windows, this needs to be considered when interpreting findings from earlier studies featuring this limitation. Arguably the most prominent limitation of treadmill sprints using force transducers is that the collection of horizontal force is an approximation, and is vulnerable to being affected by vertical ground reaction force (\(F_{\text{V}}\) or GRF-z) signals as the tether moves up and down with each step (movement of <4°; ~7% contribution of vertical force to horizontal readings) [62, 67]. While some studies have used goniometers in attempt to account for this occurrence [59, 61, 70], this is not common practice [74–76]. Moreover, although the output of power from this method is a propulsive measure of horizontally directed force and velocity, collection of these variables occurs in disparate locations (along the tether and from the track under the subject’s feet, respectively). Consequently, kinetic output does not register a null reading between foot-strikes (flight phase), but drops to approximately ~20% of the peak value [67], depending on technique. This is thought to be due either to body inertia acting on the tether during flight, some elasticity in the system, or a combination of these factors. Furthermore, significantly lower \(v_{ \text{max} }\) (e.g. 2.87 m·s−1; [67]) and acceleration on these ergometers are observed when they are compared to over-ground data from the same subjects [76]. These lower values are explained by the friction characteristics of the belt and inertia of rolling components, and a constant manufacturer pre-set track torque in the case of non-motorized variants, limiting velocity-ability. This is an issue that persists even with modernized instrumented treadmills, with the exception of feedback controlled models [80], and will be discussed in the following section.
3.3 Modern Instrumented Treadmills and the Single Treadmill-Sprint Method
Instrumented treadmills featuring the ability to obtain three-dimensional GRF data have been validated during walking, running and maximal sprinting [62, 78, 81]. These rare and costly machines allow collection of antero-posterior, medio-lateral and vertical GRF data (in association with velocity at foot-strike) from piezo-electric force sensors positioned under the treadmill (Model: KI 9007b; Kistler, Winterthur, Switzerland). Given the rarity and recent development of such ergometers, until very recently few studies have been published using such devices [62, 82–88]. Because GRF is averaged for each single foot contact (approximately 0.15–0.25 s) corresponding to a single ballistic event of one push [79, 89] at a high sampling rate (1000 Hz) [67, 79], and horizontal net power output is calculated instantaneously as the product of horizontal force and velocity (\(P_{\text{h}} = F_{\text{h}} {\cdot} v_{\text{h}} )\) collected at the same location (i.e. treadmill belt) (see Fig. 3), these machines meet many of the concerns raised by previous authors.

Graphic representation of force-velocity relationship determined from a single sprint on treadmill ergometry. The data points represent values averaged across each foot-strike, from the first (First) to the last (Last) at peak velocity. Similar to Fig. 3, \(F_{0}\) and \(v_{0}\) represent the y and x intercepts, and the theoretical maximum of force and velocity able to be produced in absence of their opposing unit
It was with this new instrumented sprint-treadmill technology (Model: ADAL3D-WR; Medical Development, HEF Tecmachine, Andrézieux-Bouthéon, France) that Morin et al. [62] showed the ability to determine sprint mechanics during a single sprint. In this method, \(F_{\text{h}}\) and \(v_{\text{h}}\) are averaged and plotted throughout the course of a sprint for each stance phase similarly to each downward stroke on pedals in early sprint-cycling studies. The steps from maximal \(F_{\text{h}}\) (\(F_{{{\text{h}}_{ \text{max} } }}\)) through to that producing \(v_{{{\text{h}}_{ \text{max} } }}\) are subsequently used to plot the linear Fv relationship [90]. The entire Fv relationship is described by the maximal theoretical horizontal force that the lower limbs could produce over one contact at a null velocity (\(F_{{{\text{h}}_{0} }}\) displayed in N·kg−1), and the theoretical maximum running velocity that could be reached in the absence of mechanical constraints (\(v_{{{\text{h}}_{0} }}\) displayed in m·s−1). A higher \(v_{{{\text{h}}_{0} }}\) value represents a greater ability to develop horizontal force at high velocities. Values of horizontal maximum power (\(P_{{{\text{h}}_{ \text{max} } }}\)) obtained via this method and mechanical variables (\(F_{{{\text{h}}_{0} }}\) and \(v_{{{\text{h}}_{0} }}\)) are congruent with results from comparable subject pools and loading parameters in earlier studies (when converted to similar time-periods) [48, 65, 66, 70], and are highly reliable for test-retest measurement (r = 0.94; P < 0.01; ICC > 0.90).
Similar to cycling literature [52, 54], where indexes of force application can be computed for each pedal downstroke, GRF output from modern treadmills can be expressed as a ratio of ‘effective’ horizontal portion of GRF data to the total resultant force averaged across each contact phase (i.e. ‘ratio of forces’; \({\text{RF}} = F_{\text{h}} /F_{\text{tot}}\)) (see Morin et al. [83]). Where it is possible (and encouraged) to perform with a technique utilizing maximal RF (i.e. 100%) in cycling, the requirement of a vertical component in sprint running means that it is impossible to present a maximal RF value without falling. Instead, when measured on a sprint treadmill (from a crouched start) sub-maximal values of RF are observed at the beginning of the sprint (28.9–42.4%), which decrease linearly with increasing velocity (R 2 < 0.707–0.975; P < 0.05) [83]. The linear decrease in RF with velocity is described as an index of force application technique (‘decrement in ratio of forces’ = \({\text{D}}_{\text{RF}}\)), and has been shown to be highly correlated with maximal speed, mean speed, and distance at 4 s in 100-m over-ground sprinting performance (r = 0.735–0.779; P < 0.01) [83]. Practically, these variables demonstrate the ability to maintain effective orientation of global force production throughout a sprint (independently from the magnitude of the resultant GRF output), and provide additional detailed analysis during sprint running acceleration.
An issue that persists even with the most updated instrumented treadmills is the compensatory friction of the belt appears to restrict the subject’s ability to obtain \(v_{{{\text{h}}_{ \text{max} } }}\) levels near to over-ground sprint running [62, 91], as previously observed by Lakomy [67]. Furthermore, determining individualized torque parameters is time-consuming, and familiarization persists as a limitation with even the most modern machines (>10 trials) [62]. Although the reduction in sprinting velocity with torque-compensated treadmills varies in significance between studies when compared to over-ground sprinting (e.g. ~20% of \(v_{ \text{max} }\); P < 0.001) [91], one could argue the ability to measure direct kinetics over a virtually unlimited time-period (e.g. change in mechanics with fatigue, across 100- to 400-m distances) [92, 93] potentially outweighs these limitations.
4 Mechanical Profiling in Over-Ground Sprint Running
Until recently, the assessment of sprint running kinetics was only possible via the specialized treadmill ergometers discussed in Sect. 3. While these methods have been markedly improved since their conception [62], the technology remains rare, assessment is costly and it requires athletes to travel to a clinical setting. While a ‘specific’ mode of assessment, treadmill sprinting remains a dissimilar modality of assessment compared to over-ground sprint running performance [76, 91], prompting authors to investigate the possibility of profiling such measures during over-ground sprinting.
Over-ground mechanical sprint profiling is somewhat difficult due to its non-stationary nature, unlike ergometer-based assessment, with requirements for such an approach being the collection of high-frequency data over an acceleration phase until \(v_{{{\text{h}}_{ \text{max} } }}\) (~20–40 m in team-sport athletes, ~50–70 m in pure-speed athletes) [94]. Despite this, literature has seen kinetics directly quantified during over-ground sprinting, either as steps within an entire sprint bout [95–101] or instrumented load cell technology used to determine resistive force against a weighted chariot [102] or pulley systems [58]. Unlike the techniques developed on cycle and treadmill ergometry, researchers have typically ignored multiple trial methods and instead placed emphasis on the development of PFv relationships for an unloaded (i.e. free-resisted) sprint [90, 103]. To the best of our knowledge, there is only a single instance of researchers attempting to use a multiple trial method overground [58], with the resultant study methodologically unclear owing to somewhat convoluted design and being available only in Italian. While there are works currently being developed (article under review) using a full length (~50-m) force plate system to fully measure sprinting kinetics throughout a single sprint, the current research uses either multiple unresisted sprint attempts over force platforms transposed together for a single linear Fv relationship [90, 103, 104], or a simple method of determining sprinting kinetics from a single sprint [90].
4.1 Composite Trial Force Plate Method
Several ground-breaking studies [105] were recently published using a method of constructing a single composite mechanical profile from multiple sprints performed over a force platform system [90, 103, 104]. This approach, first proposed by Cavagna et al. [106], generates an entire mechanical Fv relationship from seven maximal sprints performed at different starting distances behind a 6.6-m force-plate system of six force platforms connected in series [103]. The athletes [elite (N = 4) and sub-elite (N = 5) sprinters] performed 10- to 40-m sprints, which enabled the collection of a total of 18 foot-contacts (including those from blocks), at 3–5 contacts per trial for greater or lesser distances, respectively. Forward acceleration of centre of mass (COM) was calculated from contact-averaged force data, and then expressed over time to determine instantaneous velocity. Data were compiled to determine Fv and Pv relationships, both of which were well described by linear (mean R 2 > 0.892) and second-order polynomial regressions (mean R 2 > 0.732), respectively (similarly to those shown on earlier cycle and treadmill studies). Furthermore, mechanical effectiveness variables were determined for each contact phase, and correlated with overall 40-m performances. Notably, \({\text{RF}}\) averaged across the sprint performance was shown to be the second largest differentiating factor between elite and sub-elite sprinters [9.7%; effect size (ES) = 2.31] and the greatest correlation with overall 40-m performance (r > 0.933; P < 0.01). Peak values were much higher than those reported by Morin et al. [83] on an instrumented treadmill (theoretical maximum RF = 70.6 ± 5.4%), likely a result of the athlete starting from sprint blocks as opposed to from a standing crouched start. Although basic mechanical variables were shown to be related to performance in varying degrees (e.g. \(v_{0}\); r = 0.803; P < 0.01, and \(P_{ \text{max} }\); r = 0.932; P < 0.001), these results further illustrate the value in further analysis of force orientation characteristics underpinning the horizontal Fv relationship in sprint profiling. Overall, while the model showed that there were no perceivable differences between sprints for the effort involved by the sprinters (ICC = 0.686–0.958; CV = 1.84–3.76%, for a range of variables), suggesting the effects of fatigue were likely negligible for similar highly trained sprint athletes, the repeated nature and complexity of reproducing such measures limit its applicability in an applied setting.
4.2 Macroscopic Approach to Mechanical Profiling during Over-Ground Sprinting
In conjunction with the methods developed by Rabita et al. [103], Samozino et al. [90] developed a method for profiling the mechanical capabilities of the neuromuscular system using a macroscopic inverse dynamics approach [107], applied to the movement of COM during a single sprinting acceleration. This approach was similar to those proposed by Furusawa et al. [56] and Vandewalle and Gajer [108]. Models including energetics and biomechanics have been proposed by van Ingen Schenau et al. [109], Arsac and Locatelli [110] and di Prampero et al. [111]. Based on the measurement of simple velocity-time data, gathered either by a set of photo-voltaic cells (as in the case with the primary analysis of Samozino et al. [90]), high sample rate sports-radar devices [90, 112–114] or sports lasers [115], the method represents a simple alternative to many of the techniques discussed in this review. Such an approach makes several assumptions: (1) the entire body is represented in displacement of COM; (2) when averaged across the acceleration phase, no vertical acceleration occurs throughout a sprint (see limitations of the treadmill sprint method in Sect. 3 [67]); and (3) the coefficient of air drag remains constant (e.g. changes in wind strength). While not an inherent limitation due to its ease of implementation, variables are modelled over time without consideration of changes between and within steps, inclusive of both support and flight phases, rendering assessment and comparison of individual limb kinetics impossible.
In this method, a mono-exponential function [56, 108, 110, 116, 117] is applied to the raw velocity-time data (Table 2; Eq. 9). After this, the fundamental principles of dynamics in the horizontal direction enable the net horizontal antero-posterior GRF to be modelled for the COM over time, considering the mass (\(m\)) of the athlete performing the sprint in association with the acceleration of COM, and the constant aerodynamic friction of the body in motion (\(F_{\text{aero}}\)) (Eqs. 10, 11). \(F_{\text{h}}\) and \(v_{\text{h}}\) values are then plotted to determine Fv relationships and mechanical variables (\(F_{{{\text{h}}_{0} }}\) and \(v_{{{\text{h}}_{0} }}\)). As with previous methods, \(P_{{{\text{h}}_{ \text{max} } }}\) can be calculated as the interaction between \(F_{{{\text{h}}_{0} }}\) and \(v_{{{\text{h}}_{0} }}\) (Eq. 5) [28, 90, 118], and by the peak of the second-order polynomial fit between \(P_{\text{h}}\) and \(v_{\text{h}}\). Furthermore, technical variables (\({\text{RF}}\) and \(D_{\text{RF}}\)) can be calculated similarly to previous methods [83, 90, 103], with the resultant force (\(F_{\text{res}}\)) in this case being computed from estimated net vertically (see Eq. 12) and horizontally oriented GRFs. Where previous studies calculated technical variables from the second step, in this case the variables are instead calculated from 0.3 s, given determining individual step characteristics is impossible.
Importantly, Samozino et al. [90] highlighted that the macroscopic inverse dynamic approach was very similar to the multiple force plate method for GRF modelled and computed over each step (\(F_{\text{h}}\), \(F_{\text{res}}\), vertical force (i.e. \(F_{\text{v}}\)); r = 0.826–0.978; P < 0.001). Furthermore, low absolute bias was observed between methods for physical (1.88–8.04%) and technical variables (6.04–7.93%). Data were extremely well fitted with linear and polynomial regressions (mean R 2 = 0.997–0.999), with all variables presented as reliable (CV and standardised error of measurement <5%) [90]. These results serve to illustrate the strength of such an approach—that estimation of over-ground sprinting kinetics via this simple field method is practically identical to direct measurement via a complex force-plate setup. Furthermore, the method has been shown to be sensitive enough to highlight differences in mechanical variables between athletes with similar abilities, determine between playing positions and track return from injury of rugby and soccer athletes in the field [112–114]. Given the only data required is velocity-time measured with sufficient sampling rate, any practitioner with a reasonable set of photovoltaic timing gates (i.e. >4 sections), sports radar or even simple cellular devices (MySprint application) [119] could potentially use such a profiling method during their training and assessment batteries [120]. While it is technically possible to apply the same method to the data gained from widely available global positioning systems [121], the specifications of current commercial units limit the accuracy of such technology for meaningful performance inferences.
While limited in its ability to quantify individual limb kinetics, the simplicity and ease of implementation of this method suggests value for practitioners who might otherwise be unable to access the technology required to accurately assess sprinting mechanics.
5 Optimal Loading, Training Considerations and Future Research
Mechanical profiling allows the computation of the exact conditions underlying maximal power to be determined. These parameters, regularly termed ‘optimal’, represent a combination of force and velocity values (i.e. \(F_{\text{opt}}\) and \(v_{\text{opt}}\)), at which a peak metric of power is maximized (see Fig. 3) [34]. Of note, training in these conditions has been suggested as an effective method of increasing the capacity for power production [21, 35], which may improve practical performance measures provided the subject displays a favourable profile of Fv capacities [24]. Practically, in order for these data to prove valuable, they require translation into an easy-to-set normal load (\(L_{\text{opt}}\)), either as bandwidth or individual value of external stimulus, that stimulates the mechanical conditions necessary to maximize power production during training.
5.1 Optimal Loading in the Literature
Typically, the literature has shown that PFv and optimal loading characteristics are specific to movement type [28, 35, 122, 123], with a recent meta-analysis [21] describing bandwidths of 0–30% of one-repetition-maximum (1RM) for jump squat movements, 30–70% 1RM for squat movements, and >70% of 1RM for the power clean movement. While increases in mechanical capacity are likely dependent on a number of factors, the literature supports the value of training at levels around optimal (\(L_{\text{opt}}\), \(F_{\text{opt}}\), and \(v_{\text{opt}}\)) [124] in a movement transferrable to performance. Furthermore, in limited examples the application of optimal force can directly influence performance in competition scenarios [52]. Specifically, optimal loading conditions as assessed in sprint cycling [125], in near competition-specific conditions, can be replicated by manipulating crank length and gear ratios to enable the athlete to perform a cycle race in practically optimal conditions for power production [126]. There is evidence to suggest performing and training closer to \(F_{\text{opt}}\) may be beneficial for a host of acute performance properties in cycling, including increased mechanical effectiveness [127, 128], decreased movement energy cost [127, 129, 130], reduced negative muscle actions [131], increased metabolic ratio [127, 132] and increased resistance to fatigue [133, 134]. These factors strengthen the rationale for profiling these characteristics where the mechanical constraints during competition can be altered to replicate optimal levels.
Unfortunately, reviews of optimal loading [21, 35] have largely focused on acyclic ‘single extension’ movements using free-weights or smith machines. Furthermore, research examining these themes in cyclic movements has almost exclusively focused on cycling, with differing methods, equipment, varying athlete training backgrounds and performance levels rendering comparison between studies difficult [135, 136]. Of the few studies that examine optimal loading for sprint running on treadmill ergometers, Jaskólski et al. [71] showed that athletes produced peak power at a variable resistance (i.e. torque applied to the belt) of 137–195 N (10.1–14.5% of maximal inbuilt braking resistance) for a range of power indices, and proposed that the results may be dependent on athlete strength and anthropometric characteristics. A few years later, Jaskólska et al. [70] reported similar results in a group of students [N = 32; optimal loading = 176–203 N (13–15% braking force)]. However, both studies simply reported the protocol that presented the greatest level of power, rather than fitting the data with regression equations (i.e. second- or third-order polynomial), to determine the exact point on the Fv/Lv relationship at which power was maximized [49, 137, 138]. In the latter example, the authors acknowledge that 34% of the athletes did not reach a measurable peak or decline in their power-capabilities with the heaviest loading protocol, and consequently the results likely understated the mechanical capacity of the cohort. A more recent study by Andre et al. [139], suffering from the same limitations, found that most athletes in their sample (~73%) produced their peak power between 25 and 35% of body mass (BM), based on the unsubstantiated manufacturer pre-set electromagnetic braking resistance for treadmill ergometer (Model: Woodway Force 3.0, Eugene, OR, USA). In contrast, using a modern instrumented treadmill, Morin et al. [62] showed three increasing levels of braking resistance did not significantly alter \(P_{ \text{max} }\) determined during a single sprint, reminiscent of increased force and decreased velocity output. This was mainly due to the fact that (1) power output was measured continuously during the sprint acceleration (in contrast to \(v_{ \text{max} }\) plateau in previous multiple trials method) and (2) maximal power was reached during the acceleration phase (but not at the same time) whatever the braking resistance. In any case, while the determination of optimal loading on treadmill ergometry offers an additional value by which to measure athlete ability, its relation to training implementation is limited to the assessment modality itself. That is, the conditions determined in these studies may only be replicated in training with access to specialized treadmill ergometry.
5.2 Optimal Conditions for Loading in Over-Ground Sprinting
At this stage, to the best of our knowledge no literature has clearly reported the methods necessary to profile practical optimal loading conditions during over-ground sprinting. That is, no research has used a multiple trial method with progressive resistance, such as sprinting sleds, braking devices or cable winches, to profile PFv relationships that can be understood and replicated with scientific rigor. While optimal loading conditions for sprint running training modalities over ground have been discussed [120, 124, 135, 140], authors have typically limited loading to that which maximizes external stimulus without significantly altering kinematics of the unloaded sprint movement (e.g. <10% decrease in \(v_{ \text{max} }\), or <12.6% of BM) [140–142]. While training in this manner no doubt achieves the goal of maintaining absolute kinematic similarity to unresisted performance, there is evidence to suggest that these loading protocols are far from the conditions necessary for development of maximal power. Of note, the fact that \(F_{\text{opt}}\) and \(v_{\text{opt}}\) occur at approximately half of the maximum velocity attained in an unloaded sprint (\(0.5{\cdot} F_{0}\) and \(0.5{\cdot} v_{0}\), [34]) would appear to challenge these guidelines, provided increased \(P_{ \text{max} }\) is the goal. Recent evidence suggests training at heavier loads versus more traditional, lighter loading protocols benefits sprint running performance to a greater degree (i.e. 10 vs. 43% of BM loading onto a resistive sled device) [35, 95]. To date, while \(v_{\text{opt}}\) has been reported in elite rugby athletes at between 4.31 and 4.61 m·s−1 [113], no specific optimal loading/training strategy has been determined for over-ground sprinting regarding power development. The effects of sprint training using loading protocols of such magnitude (e.g. 50% velocity decrement), both acute and longitudinal performance outcomes, are yet to be quantified.
5.3 Implications for Training, and Future Research
The relationship between Fv properties, as illustrated by the slope of the linear regression fit (\(S_{\text{Fv}}\)), denotes that \(P_{ \text{max} }\) and \(S_{\text{Fv}}\) are independent from one another. Evidence suggests performance in both jumping and sprint running is reliant not only on the expression of \(P_{ \text{max} }\), but also on the absolute level and balance between Fv components [28, 83]. Practically, two athletes exhibiting identical \(P_{ \text{max} }\) values could present markedly different Fv relationships (as a function of either a higher or a lower \(F_{0}\) or \(v_{0}\)) [28, 83], which may be evident in practical performance measures [120]. Considering Fv characteristics in single extension movements have recently been determined as individualized [24, 28], it would seem exercise and load prescription should occur based on both \(S_{\text{Fv}}\) and \(P_{ \text{max} }\) qualities [120]. While training at \(L_{\text{opt}}\) may be a simple approach to increase \(P_{ \text{max} }\), targeted programming may see prescription of greater or lesser load (force or velocity dominant stimuli, respectively) depending on the orientation of \(S_{\text{Fv}}\) and the targeted task (e.g. sprint distance to optimize, or level of resistive force to overcome). Importantly, this is an integrative multi-factorial approach, and targeted training based on \(S_{\text{Fv}}\)orientation may not be as important to novice athletes, who will likely see increases in performance with basic prescription, as opposed to highly-trained athletes. Furthermore, there is currently little research investigating these theories in practice, none of which exists in the realm of sprint running; hence investigation of this nature is required.
6 Conclusions
The Fv relationship and maximal power capacity offer understanding of the limits of the human body for sprinting performance. These mechanical capabilities can be accurately measured by various methods during acceleration sprinting in cycling and sprint running (treadmill and overground). While it is well known that adaptations are specific to the velocity used in training, there is an overall paucity of research using PFv methods on updated equipment, and further investigation is required in longitudinal studies. Given the rapid development of easily accessible profiling methods, research providing normative data on athletes from varying performance levels using modern technologies would provide insight into the mechanical performance requirements of unique sporting cohorts. Elucidating these normative or optimal characteristics, including methods through which to implement meaningful changes, would prove invaluable in the guidance of individualised and targeted training programs to increase the capacities underlying maximal sprinting performance.
References
Hill AV. The heat of shortening and the dynamic constants of muscle. Proc R Soc Lond B Biol Sci. 1938;126(843):136–95.
Gollnick PD, Matoba H. The muscle-fiber composition of skeletal-muscle as a predictor of athletic success—an overview. Am J Sports Med. 1984;12(3):212–7.
Kraemer WJ, Newton RU. Training for muscular power. Phys Med Rehabil Clin N Am; 2000. p. 341–68, vii.
Newton RU, Kraemer WJ. Developing explosive muscular power: implications for a mixed methods training strategy. Strength Cond J. 1994;16(5):20–31.
Amar J, Le Chatelier H. Le moteur humain et les bases scientifiques du travail professionel. Pinat: H Dunod et E; 1914.
Hill AV. The maximum work and mechanical efficiency of human muscles, and their most economical speed. J Physiol. 1922;56(1–2):19–41.
Lupton H. The relation between the external work produced and the time occupied in a single muscular contraction in man. J Physiol. 1922;57(1–2):68–75.
Best CH, Partridge RC. The equation of motion of a runner, exerting a maximal effort. Proc R Soc Lond B Biol Sci. 1928;103(724):218–25.
Aubert X. Structure and physiology of the striated muscle. I. Contractile mechanism in vivo; mechanical and thermal aspects. J Physiol (Paris). 1956;48(2):105–53.
Fenn WO, Marsh BS. Muscular force at different speeds of shortening. J Physiol. 1935;85(3):277–97.
Yoshihuku Y, Herzog W. Optimal-design parameters of the bicycle-rider system for maximal muscle power output. J Biomech. 1990;23(10):1069–79.
Yoshihuku Y, Herzog W. Maximal muscle power output in cycling: a modelling approach. J Sport Sci. 1996;14(2):139–57.
Bobbert MF. Why is the force-velocity relationship in leg press tasks quasi-linear rather than hyperbolic? J Appl Physiol. 2012;112(12):1975–83.
Jaric S. Force-velocity relationship of muscles performing multi-joint maximum performance tasks. Int J Sports Med. 2015;36(9):699–704.
Yamauchi J, Ishii N. Relations between force-velocity characteristics of the knee-hip extension movement and vertical jump performance. J Strength Cond Res. 2007;21(3):703–9.
Perrine JJ, Edgerton VR. Muscle force-velocity and power-velocity relationships under isokinetic loading. Med Sci Sports. 1977;10(3):159–66.
Murphy AJ, Wilson GJ, Pryor JF. Use of the iso-inertial force mass relationship in the prediction of dynamic human-performance. Eur J Appl Physiol Occup Physiol. 1994;69(3):250–7.
Yamauchi J, Mishima C, Nakayama S, et al. Force-velocity, force-power relationships of bilateral and unilateral leg multi-joint movements in young and elderly women. J Biomech. 2009;42(13):2151–7.
Samozino P, Horvais N, Hintzy F. Why does power output decrease at high pedaling rates during sprint cycling? Med Sci Sports Exerc. 2007;39(4):680–7.
Bobbert MF, Casius LJ, van Soest AJ. On the relationship between pedal force and crank angular velocity in sprint cycling. Med Sci Sports Exerc. 2015 (Published ahead of print).
Soriano MA, Jimenez-Reyes P, Rhea MR, et al. The optimal load for maximal power production during lower-body resistance exercises: a meta-analysis. Sports Med. 2015;45(8):1191–205.
Cuk I, Markovic M, Nedeljkovic A, et al. Force-velocity relationship of leg extensors obtained from loaded and unloaded vertical jumps. Eur J Appl Physiol. 2014;114(8):1703–14.
Rahmani A, Viale F, Dalleau G, et al. Force/velocity and power/velocity relationships in squat exercise. Eur J Appl Physiol. 2001;84(3):227–32.
Samozino P, Edouard P, Sangnier S, et al. Force-velocity profile: imbalance determination and effect on lower limb ballistic performance. Int J Sports Med. 2014;35(6):505–10.
Sheppard JM, Cormack S, Taylor KL, et al. Assessing the force-velocity characteristics of the leg extensors in well-trained athletes: the incremental load power profile. J Strength Cond Res. 2008;22(4):1320–6.
Samozino P, Morin JB, Hintzy F, et al. Jumping ability: a theoretical integrative approach. J Theor Biol. 2010;264(1):11–8.
Samozino P, Morin JB, Hintzy F, et al. A simple method for measuring force, velocity and power output during squat jump. J Biomech. 2008;41(14):2940–5.
Samozino P, Rejc E, Di Prampero PE, et al. Optimal force-velocity profile in ballistic movements—altius: citius or fortius? Med Sci Sports Exerc. 2012;44(2):313–22.
Bosco C, Luhtanen P, Komi PV. A simple method for measurement of mechanical power in jumping. Eur J Appl Physiol Occup Physiol. 1983;50(2):273–82.
Vandewalle H, Peres G, Monod H. Standard anaerobic exercise tests. Sports Med. 1987;4(4):268–89.
Cormie P, McGuigan MR, Newton RU. Influence of strength on magnitude and mechanisms of adaptation to power training. Med Sci Sports Exerc. 2010;42(8):1566–81.
Cormie P, McGuigan MR, Newton RU. Adaptations in athletic performance after ballistic power versus strength training. Med Sci Sports Exerc. 2010;42(8):1582–98.
Cormie P, McGuigan MR, Newton RU. Developing maximal neuromuscular power: Part 1—biological basis of maximal power production. Sports Med. 2011;41(1):17–38.
Sargeant AJ, Dolan P, Young A. Optimal velocity for maximal short-term (anaerobic) power output in cycling. Int J Sports Med. 1984;5(supplement 1):124–5.
Kawamori N, Haff GG. The optimal training load for the development of muscular power. J Strength Cond Res. 2004;18(3):675–84.
Dickinson S. The dynamics of bicycle pedalling. Proc R Soc Lond B Biol Sci. 1928;103(724):225–33.
Vandewalle H, Driss T. Friction-loaded cycle ergometers: past, present and future. Cogent Eng. 2015;2(1):1–35.
Driss T, Vandewalle H. The measurement of maximal (anaerobic) power output on a cycle ergometer: a critical review. Biomed Res Int. 2013;2013:589361.
Pérès G, Vandewalle H, Monod H. Aspect particulier de la relation charge-vitesse lors du pédalage sur cycloergometre. J Physiol (Paris). 1981;77:10A.
Pirnay F, Crielaard J. Mesure de la puissance anaérobie alactique. Med Sport. 1979;53:13–6.
Vandewalle H, Peres G, Heller J, et al. All out anaerobic capacity tests on cycle ergometers—a comparative-study on men and women. Eur J Appl Physiol Occup Physiol. 1985;54(2):222–9.
Bassett DR. Correcting the Wingate test for changes in kinetic energy of the ergometer flywheel. Int J Sports Med. 1989;10(6):446–9.
Lakomy HKA. Effect of load on corrected peak power output generated on friction loaded cycle ergometers. J Sport Sci. 1985;3(3):240.
Lakomy HKA. Measurement of work and power output using friction-loaded cycle ergometers. Ergonomics. 1986;29(4):509–17.
Winter EM, Brown D, Roberts NK, et al. Optimized and corrected peak power output during friction-braked cycle ergometry. J Sport Sci. 1996;14(6):513–21.
Seck D, Vandewalle H, Decrops N, et al. Maximal power and torque-velocity relationship on a cycle ergometer during the acceleration phase of a single all-out exercise. Eur J Appl Physiol Occup Physiol. 1995;70(2):161–8.
Kyle CR, Caiozzo VJ. Experiments in human ergometry as applied to the design of human powered vehicles. Int J Sport Biomech. 1986;2:6–19.
Morin JB, Belli A. A simple method for measurement of maximal downstroke power on friction-loaded cycle ergometer. J Biomech. 2004;37(1):141–5.
Arsac LM, Belli A, Lacour JR. Muscle function during brief maximal exercise: accurate measurements on a friction-loaded cycle ergometer. Eur J Appl Physiol Occup Physiol. 1996;74(1–2):100–6.
Morin JB, Hintzy F, Belli A, et al. Relations force–vitesse et performances en sprint chez des athlètes entraı̂nés. Sci Sports. 2002;17(2):78–85.
Martin JC, Brown NA. Joint-specific power production and fatigue during maximal cycling. J Biomech. 2009;42(4):474–9.
Dorel S, Couturier A, Lacour JR, et al. Force-velocity relationship in cycling revisited: benefit of two-dimensional pedal forces analysis. Med Sci Sports Exerc. 2010;42(6):1174–83.
Buttelli O, Vandewalle H, Peres G. The relationship between maximal power and maximal torque-velocity using an electronic ergometer. Eur J Appl Physiol Occup Physiol. 1996;73(5):479–83.
Davis RR, Hull ML. Measurement of pedal loading in bicycling: II. Analysis and results. J Biomech. 1981;14(12):857–72.
Zameziati K, Mornieux G, Rouffet D, et al. Relationship between the increase of effectiveness indexes and the increase of muscular efficiency with cycling power. Eur J Appl Physiol. 2006;96(3):274–81.
Furusawa K, Hill AV, Parkinson JL. The dynamics of “sprint” running. Proc R Soc Lond B Biol Sci. 1927;102(713):29–42.
Bassett DR. Scientific contributions of A. V. Hill: exercise physiology pioneer. J Appl Physiol. 2002;93(5):1567–82.
Bisciotti GN, Necchi P, Iodice GP, et al. The biomechanical parameters of the sprint. Med Sport (Roma). 2005;58(2):89–96.
Chelly SM, Denis C. Leg power and hopping stiffness: relationship with sprint running performance. Med Sci Sports Exerc. 2001;33(2):326–33.
Falk B, Weinstein Y, Dotan R, et al. A treadmill test of sprint running. Scand J Med Sci Sports. 1996;6(5):259–64.
Jaskólska A, Goossens P, Veenstra B, et al. Treadmill measurement of the force-velocity relationship and power output in subjects with different maximal running velocities. Sports Med Train Rehab. 1998;8(4):347–58.
Morin JB, Samozino P, Bonnefoy R, et al. Direct measurement of power during one single sprint on treadmill. J Biomech. 2010;43(10):1970–5.
Belli A, Lacour J-R. Treadmill ergometer for power measurement during sprint running. J Biomech. 1989;22(10):985.
Brughelli M, Cronin J, Chaouachi A. Effects of running velocity on running kinetics and kinematics. J Strength Cond Res. 2011;25(4):933–9.
Cheetham ME, Williams C, Lakomy HK. A laboratory running test: metabolic responses of sprint and endurance trained athletes. Br J Sports Med. 1985;19(2):81–4.
Funato K, Yanagiya T, Fukunaga T. Ergometry for estimation of mechanical power output in sprinting in humans using a newly developed self-driven treadmill. Eur J Appl Physiol. 2001;84(3):169–73.
Lakomy HKA. The use of a non-motorized treadmill for analysing sprint performance. Ergonomics. 1987;30(4):627–37.
Nevill ME, Boobis LH, Brooks S, et al. Effect of training on muscle metabolism during treadmill sprinting. J Appl Physiol; 1989. p. 2376–82.
Dal Monte A, Leonardi L. Nouvelle méthode d evaluation de la puissance anaérobic maximale alactacide. In: Congrès Groupement Latin Médicine Du Sport, S. Nice; 1977.
Jaskólska A, Goossens P, Veenstra B, et al. Comparison of treadmill and cycle ergometer measurements of force-velocity relationships and power output. Int J Sports Med. 1999;20(3):192–7.
Jaskólski A, Veenstra B, Goossens P, et al. Optimal resistance for maximal power during treadmill running. Sports Med Train Rehab. 1996;7(1):17–30.
McKenna M, Riches PE. A comparison of sprinting kinematics on two types of treadmill and over-ground. Scand J Med Sci Sports. 2007;17(6):649–55.
Sirotic AC, Coutts AJ. The reliability of physiological and performance measures during simulated team-sport running on a non-motorised treadmill. J Sci Med Sport. 2008;11(5):500–9.
Cross MR, Brughelli ME, Cronin JB. Effects of vest loading on sprint kinetics and kinematics. J Strength Cond Res. 2014;28(7):1867–74.
Ross RE, Ratamess NA, Hoffman JR, et al. The effects of treadmill sprint training and resistance training on maximal running velocity and power. J Strength Cond Res. 2009;23(2):385–94.
Highton JM, Lamb KL, Twist C, et al. The reliability and validity of short-distance sprint performance assessed on a nonmotorized treadmill. J Strength Cond Res. 2012;26(2):458–65.
Frishberg BA. An analysis of overground and treadmill sprinting. Med Sci Sports Exerc. 1983;15(6):478–85.
Kram R, Griffin TM, Donelan JM, et al. Force treadmill for measuring vertical and horizontal ground reaction forces. J Appl Physiol; 1998. p. 764–9.
Martin JC, Wagner BM, Coyle EF. Inertial-load method determines maximal cycling power in a single exercise bout. Med Sci Sports Exerc. 1997;29(11):1505–12.
Bowtell MV, Tan H, Wilson AM. The consistency of maximum running speed measurements in humans using a feedback-controlled treadmill, and a comparison with maximum attainable speed during overground locomotion. J Biomech. 2009;42(15):2569–74.
Belli A, Bui P, Berger A, et al. A treadmill ergometer for three-dimensional ground reaction forces measurement during walking. J Biomech. 2001;34(1):105–12.
Morin JB, Samozino P, Edouard P, et al. Effect of fatigue on force production and force application technique during repeated sprints. J Biomech. 2011;44(15):2719–23.
Morin JB, Edouard P, Samozino P. Technical ability of force application as a determinant factor of sprint performance. Med Sci Sports Exerc. 2011;43(9):1680–8.
Girard O, Brocherie F, Morin JB, et al. Comparison of four sections for analysing running mechanics alterations during repeated treadmill sprints. J Appl Biomech. 2015;31(5):389–95.
Girard O, Brocherie F, Morin JB, et al. Intra- and inter-session reliability of running mechanics during treadmill sprints. Int J Sports Physiol Perform. 2015 (Published ahead of print).
Girard O, Brocherie F, Morin JB, et al. Neuro-mechanical determinants of repeated treadmill sprints–usefulness of an “hypoxic to normoxic recovery” approach. Front Physiol. 2015;6:1–14.
Brocherie F, Millet GP, Morin JB, et al. Mechanical alterations to repeated treadmill sprints in normobaric hypoxia. Med Sci Sports Exerc. 2016 (Published ahead of print).
Morin JB, Bourdin M, Edouard P, et al. Mechanical determinants of 100-m sprint running performance. Eur J Appl Physiol. 2012;112(11):3921–30.
Williams JH, Barnes WS, Signorile JF. A constant-load ergometer for measuring peak power output and fatigue. J Appl Physiol; 1988. p. 2343–8.
Samozino P, Rabita G, Dorel S, et al. A simple method for measuring power, force, velocity properties, and mechanical effectiveness in sprint running. Scand J Med Sci Sports. 2016;26(6):648–58.
Morin JB, Seve P. Sprint running performance: comparison between treadmill and field conditions. Eur J Appl Physiol. 2011;111(8):1695–703.
Girard O, Brocherie F, Tomazin K, et al. Changes in running mechanics over 100-m, 200-m and 400-m treadmill sprints. J Biomech. 2016 (Published ahead of print).
Tomazin K, Morin J, Strojnik V, et al. Fatigue after short (100-m), medium (200-m) and long (400-m) treadmill sprints. Eur J Appl Physiol. 2012;112(3):1027–36.
Brüggemann G-P, Koszewski D, Müller H. Biomechanical Research Project, Athens 1997: Final Report: Meyer and Meyer Sport; 1999.
Kawamori N, Nosaka K, Newton RU. Relationships between ground reaction impulse and sprint acceleration performance in team sport athletes. J Strength Cond Res. 2014;27(3):568–73.
Komi PV. Biomechanical features of running with special emphasis on load characteristics and mechanical efficiency. In: Nigg BN, Kerr BA, editors. Biomechanical aspects of sport shoes and playing surfaces. Calgary: University of Calgary; 1983. p. 123–34.
Mero A. Force-time characteristics and running velocity of male sprinters during the acceleration phase of sprinting. Res Q Exerc Sport. 1988;59(2):94–8.
Mero A, Komi PV. Force-velocity, Emg-velocity, and elasticity-velocity relationships at submaximal, maximal and supramaximal running speeds in sprinters. Eur J Appl Physiol Occup Physiol. 1986;55(5):553–61.
Bezodis IN, Kerwin DG, Salo AI. Lower-limb mechanics during the support phase of maximum-velocity sprint running. Med Sci Sports Exerc. 2008;40(4):707–15.
Hunter JP, Marshall RN, McNair P. Relationships between ground reaction force impulse and kinematics of sprint-running acceleration. J Appl Biomech. 2005;21(1):31–43.
Kugler F, Janshen L. Body position determines propulsive forces in accelerated running. J Biomech. 2010;43(2):343–8.
Martinez-Valencia MA, Romero-Arenas S, Elvira JL, et al. Effects of sled towing on peak force, the rate of force development and sprint performance during the acceleration phase. J Hum Kinet. 2015;46(1):139–48.
Rabita G, Dorel S, Slawinski J, et al. Sprint mechanics in world-class athletes: a new insight into the limits of human locomotion. Scand J Med Sci Sports. 2015;25(5):583–94.
Morin JB, Slawinski J, Dorel S, et al. Acceleration capability in elite sprinters and ground impulse: push more, brake less? J Biomech. 2015;48(12):3149–54.
Clark K, Weyand P. Sprint running research speeds up: a first look at the mechanics of elite acceleration. Scand J Med Sci Sports. 2015;25(5):581–2.
Cavagna GA, Komarek L, Mazzoleni S. The mechanics of sprint running. J Physiol. 1971;217(3):709–21.
Helene O, Yamashita MT. The force, power, and energy of the 100 meter sprint. Am J Phys. 2010;78(3):307–9.
Vandewalle H, Gajer B. Relation force-vitesse et puissance maximale au cours d’un sprint maximal en course à pied. Arch Physiol Biochem. 1996;104:D74–5.
van Ingen Schenau GJ, Jacobs R, de Koning JJ. Can cycle power predict sprint running performance? Eur J Appl Physiol Occup Physiol. 1991;63(3–4):255–60.
Arsac LM, Locatelli E. Modeling the energetics of 100-m running by using speed curves of world champions. J Appl Physiol. 2002;92(5):1781–8.
di Prampero PE, Botter A, Osgnach C. The energy cost of sprint running and the role of metabolic power in setting top performances. Eur J Appl Physiol. 2015;115(3):451–69.
Mendiguchia J, Samozino P, Martinez-Ruiz E, et al. Progression of mechanical properties during on-field sprint running after returning to sports from a hamstring muscle injury in soccer players. Int J Sports Med. 2014;35(8):690–5.
Cross MR, Brughelli M, Brown SR, et al. Mechanical properties of sprinting in elite rugby union and rugby league. Int J Sports Physiol Perform. 2015;10(6):695–702.
Mendiguchia J, Edouard P, Samozino P, et al. Field monitoring of sprinting power-force-velocity profile before, during and after hamstring injury: two case reports. J Sport Sci. 2016;34(6):535–41.
Buchheit M, Samozino P, Glynn JA, et al. Mechanical determinants of acceleration and maximal sprinting speed in highly trained young soccer players. J Sport Sci. 2014;32(20):1906–13.
Volkov NI, Lapin VI. Analysis of the velocity curve in sprint running. Med Sci Sports Exerc. 1979;11(4):332–7.
di Prampero PE, Fusi S, Sepulcri L, et al. Sprint running: a new energetic approach. J Exp Biol. 2005;208(14):2809–16.
Vandewalle H, Peres G, Heller J, et al. Force-velocity relationship and maximal power on a cycle ergometer: correlation with the height of a vertical jump. Eur J Appl Physiol Occup Physiol. 1987;56(6):650–6.
Romero-Franco N, Jiménez-Reyes P, Castaño-Zambudio A, et al. Sprint performance and mechanical outputs computed with an iPhone app: comparison with existing reference methods. Eur J Sport Sci. 2016 (Published ahead of print).
Morin JB, Samozino P. Interpreting power-force-velocity profiles for individualized and specific training. Int J Sports Physiol Perform. 2016;11(2):267–72.
Nagahara R, Botter A, Rejc E, et al. Concurrent validity of GPS for deriving mechanical properties of sprint acceleration. Int J Sports Physiol Perform. 2016 (Published ahead of print).
Suzovic D, Markovic G, Pasic M, et al. Optimum load in various vertical jumps support the maximum dynamic output hypothesis. Int J Sports Med. 2013;34(11):1007–14.
Cormie P, McCaulley GO, Triplett NT, et al. Optimal loading for maximal power output during lower-body resistance exercises. Med Sci Sports Exerc. 2007;39(2):340–9.
Wilson GJ, Newton RU, Murphy AJ, et al. The optimal training load for the development of dynamic athletic performance. Med Sci Sports Exerc. 1993;25(11):1279–86.
Linossier MT, Dormois D, Fouquet R, et al. Use of the force-velocity test to determine the optimal braking force for a sprint exercise on a friction-loaded cycle ergometer. Eur J Appl Physiol Occup Physiol. 1996;74(5):420–7.
Dorel S, Hautier CA, Rambaud O, et al. Torque and power-velocity relationships in cycling: relevance to track sprint performance in world-class cyclists. Int J Sports Med. 2005;26(9):739–46.
Dorel S, Bourdin M, Van Praagh E, et al. Influence of two pedalling rate conditions on mechanical output and physiological responses during all-out intermittent exercise. Eur J Appl Physiol. 2003;89(2):157–65.
Sargeant AJ, Hoinville E, Young A. Maximum leg force and power output during short-term dynamic exercise. J Appl Physiol Respir Environ Exerc Physiol; 1981. p. 1175–82.
Francescato MP, Girardis M, di Prampero PE. Oxygen cost of internal work during cycling. Eur J Appl Physiol Occup Physiol. 1995;72(1–2):51–7.
McDaniel J, Durstine JL, Hand GA, et al. Determinants of metabolic cost during submaximal cycling. J Appl Physiol. 2002;93(3):823–8.
Neptune RR, Herzog W. The association between negative muscle work and pedaling rate. J Biomech. 1999;32(10):1021–6.
Morel B, Clemencon M, Rota S, et al. Contraction velocity influence the magnitude and etiology of neuromuscular fatigue during repeated maximal contractions. Scand J Med Sci Sports. 2015;25(5):432–41.
Beelen A, Sargeant AJ. Effect of fatigue on maximal power output at different contraction velocities in humans. J Appl Physiol. 1991;71(6):2332–7.
McCartney N, Heigenhauser GJ, Jones NL. Power output and fatigue of human muscle in maximal cycling exercise. J Appl Physiol Respir Environ Exerc Physiol. 1983;55(1):218–24.
Jaafar H, Rouis M, Coudrat L, et al. Effects of load on wingate test performances and reliability. J Strength Cond Res. 2014;28(12):3462–8.
Vargas NT, Robergs RA, Klopp DM. Optimal loads for a 30-s maximal power cycle ergometer test using a stationary start. Eur J Appl Physiol. 2015;115(5):1087–94.
Hautier C, Linossier M, Belli A, et al. Optimal velocity for maximal power production in non-isokinetic cycling is related to muscle fibre type composition. Eur J Appl Physiol Occup Physiol. 1996;74(1–2):114–8.
Hintzy F, Belli A, Grappe F, et al. Optimal pedalling velocity characteristics during maximal and submaximal cycling in humans. Eur J Appl Physiol Occup Physiol. 1999;79(5):426–32.
Andre MJ, Fry AC, Lane MT. Appropriate loads for peak-power during resisted sprinting on a non-motorized treadmill. J Hum Kinet. 2013;38:161–7.
Petrakos G, Morin JB, Egan B. Resisted sled sprint training to improve sprint performance: a systematic review. Sports Med. 2016;46(3):381–400.
Lockie RG, Murphy AJ, Spinks CD. Effects of resisted sled towing on sprint kinematics in field-sport athletes. J Strength Cond Res. 2003;17(4):760–7.
Alcaraz PE, Palao JM, Elvira JL. Determining the optimal load for resisted sprint training with sled towing. J Strength Cond Res. 2009;23(2):480–5.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
No funding was received for this review which may have affected study design, data collection, analysis or interpretation of data, writing of this manuscript, or the decision to submit for publication.
Conflict of interest
Matt R. Cross, Matt Brughelli, Pierre Samozino and Jean-Benoit Morin declare that they have no conflicts of interest that are directly relevant to the content of this review.
Author contributions
All authors were involved in the preparation of the entire content of this manuscript.
Rights and permissions
About this article

Cite this article
Cross, M.R., Brughelli, M., Samozino, P. et al. Methods of Power-Force-Velocity Profiling During Sprint Running: A Narrative Review. Sports Med 47, 1255–1269 (2017). https://doi.org/10.1007/s40279-016-0653-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40279-016-0653-3