
Abstract
The purpose of this study was to investigate the effect of two cycling velocities on power output and concomitant metabolic and cardiorespiratory responses to repeated all-out exercises. Mean power output (P m), total work (W tot), total oxygen consumption (VO2tot) and blood lactate accumulation (Δ[La]b) were evaluated in 13 male subjects who performed two series of twelve 5-s bouts of sprint cycling. Recovery periods of 45-s were allowed between trials. One series was executed at optimal velocity (V opt: velocity for greatest power) and the other one at 50% V opt (0.5V opt). Velocities obtained in these conditions were Vopt=116.6 (4.7) rpm; 0.5Vopt=60.6 (4.9) rpm. After a phase of adaptation in oxygen uptake in the first part of the series, the data from the 6th to the 12th sprint were as follows: P m, 924.6 (73.9) versus 689.2 (61.8) W; W tot, 29.95 (4.14) versus 22.04 (3.17) kJ; VO2tot, 12.80 (1.36) versus 10.58 (1.37) l; Δ[La]b, 2.72 (1.22) versus 0.64 (0.79) mmol.l−1, respectively (P<0.001). Both W tot and VO2tot were consistently higher at optimal velocity (+21 and +35.8%, respectively). The present findings demonstrate that during intermittent short-term all-out exercise requiring maximal activation, the energy turnover is not necessarily maximal. It depends on muscle contraction velocity. The increase, lower than expected, in metabolic response from 0.5V opt to V opt suggests also that mechanical efficiency is higher at V opt.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In a single short-term (<10-s) all-out dynamic cycle exercise, the maximum capacity of human muscle to generate power is described by a polynomial power–velocity relationship (Sargeant et al. 1981; Seck et al. 1995; Arsac et al. 1996).The power output reaches a maximum value (P max) at optimal velocity (V opt). Studies performed on an isokinetic (McCartney et al. 1983) or friction-loaded cycle ergometer (Linossier et al. 1996) confirmed pedalling rate to be the major factor governing the amount of work generated during a single sprint (5-s). The mechanical output is different for maximal exercises performed at different velocities; i.e. the closer the velocity is to V opt, the higher the total amount of work performed. On the other hand it is commonly accepted that the rate of ATP resynthesis is maximal during brief "all-out" effort (Gaitanos et al. 1993). The key question is therefore to understand how during all-out exercise this variation in work and power output, which is determined by the mechanical conditions of muscle contraction (in terms of velocity), might be related to a change of metabolic requirements and hence energy turnover. In studies on isolated muscles, Hill (1938) established a relationship between energy consumption and work generated during maximal contractions. Ferguson et al. (2000, 2001) recently described the influence of contraction frequency on total power output and energy cost during volitional knee extensor exercise of submaximal intensities. Little is known about the effect of velocity on both mechanical output and general metabolic and cardiorespiratory responses to all-out exercise in vivo.
During high-intensity intermittent exercise, short-term bouts lead to primarily anaerobic energy production, whereas recovery processes depend upon aerobic energy metabolism in order to resynthesize phosphocreatine, restore the acid–base balance and recharge oxygen bound to myoglobin. From a physiological perspective, the actual oxygen uptake cannot attain a steady state and is likely to be complex within each work–recovery cycle. However, during a prolonged series of maximal sprints interspersed with adequate recovery periods, the average oxygen uptake for each work–recovery cycle (V̇O2) can stabilize at a constant submaximal level. For instance, Balsom et al. (1992) reported an oxygen uptake of 57% of VO2max during intermittent 15×40-m sprint running (5.6 to 5.8 s) with a 60-s rest period between sprints. It is well established that the recovery oxygen uptake (i.e. oxygen debt) markedly overestimates anaerobic energy release (Bangsbo et al. 1990). Nevertheless, Knuttgen and Saltin (1972) verified the proportional relationship between high-energy phosphate depletion during periods of exercise and oxygen debt. Taking these observations into consideration, it appears that the magnitude of average oxygen uptake, associated with the measure of the rate of lactate accumulation in blood (di Prampero and Ferretti 1999), should provide an estimate of physiological stress during intermittent exercise, at least with the aim of comparing two experimental conditions.
The purpose of this study, therefore, was to investigate the effect of two different pedal-crank velocities (V opt and 50% of V opt: 0.5V opt) on the average V̇O2 and potential blood lactate accumulation during two series of 12 repeated cycling sprints, both requiring maximal involvement of the subjects (i.e. all-out intensity). In order to control the influences of the relationship between the duration of work and recovery periods and of the form of recovery, subjects were submitted to the same work–recovery duration ratio of 5-s isokinetic sprints with 45-s of rest at both velocities. This protocol was designed to obtain a constant level of power and V̇O2 response and to avoid fatigue in both conditions. Moreover, a critical issue when studying the relationship between mechanical output and physiological response is to take into account the internal power (Widrick et al. 1992; Ferguson et al. 2000, 2001) or the oxygen cost of internal power (Gaesser and Brooks 1975; Francescato et al. 1995), in particular when different pedalling frequencies are to be compared. The oxygen cost of internal power (VO2int), that is defined by these authors as the oxygen consumed to keep the limbs in motion, was therefore evaluated at both velocities in order to determine to what extent it influences V̇O2. Muscle activation was measured in V opt and 0.5V opt conditions by EMG analysis in vastus lateralis (VL) and biceps femoris (BF) muscles. The main hypothesis was that different movement frequencies (V opt, 0.5V opt) would modify the amount of energy output (power and work) and consequently would determine the metabolic and cardiovascular response (oxygen consumption, ventilation, heart rate and blood lactate accumulation) during short-term all-out exercise. Finally, the possible effect of pedalling condition on mechanical efficiency was discussed.
Methods
Subjects
A total of 13 male cyclists volunteered to participate in this study. The test procedures were explained to the subjects before they gave their informed consent. The experimental design of the study was approved by the Ethical Committee of the Lyon University Hospital. The mean (SD) age, height and body mass were 25.4 (4.2) years, 176.3 (7.8) cm and 68 (7.1) kg, respectively. Percentage body fat (Durnin and Rahaman 1967) was 9.5 (1.5)%. Mean VO2max, measured on a cycle ergometer, was 4.57 (0.49) l.min−1. All subjects competed in endurance events and their yearly training distance averaged approximately 10 000 km (including competition).
Experimental procedures
Each subject performed four tests at the same time of the day and at least 4 h after a meal. Different test sessions were separated by 3–6 days.
Day 1. Force–velocity measurements and VO2max
After a 5-min warm-up, the subjects were asked to perform three maximal cycling sprints of 6-s duration, each one separated by at least 4-min rest. Three friction loads of 0.25, 0.50 and 0.75 N.kg−1 body mass (corresponding to torque of 0.238, 0.477 and 0.717 Nm.kg−1 body mass) were applied to the friction belt. The friction-loaded cycle ergometer (Monark 818E, Stockholm, Sweden) is described elsewhere (Arsac et al. 1996). The device was equipped with a strain gauge (200 N, bandwidth 500 Hz) to measure the frictional force and with an optical encoder (1969.2 points per metre of displacement or 11 815 points per pedal revolution) to record the flywheel displacement. A lever-arm (with weights) was added to apply the frictional load. Force and displacement signals were sampled (200 Hz) and stored on a PC via a 12-bit analog-to-digital interface card (DAS-8, 12 bits, Keithley Metrabyte, Taunton, Mass., USA). First- and second-order derivatives of the flywheel displacement were calculated to obtain flywheel velocity and acceleration. The external force produced was calculated as the sum of frictional force (given by the strain gauge) and force necessary to accelerate the flywheel (Lakomy 1986). Power, force recalculated as torque and velocity recalculated as pedalling rate were averaged for the period of each pedal down-stroke. After computation, the data from all the sprints were used to draw the torque– and power–velocity relationships respectively, using a linear and a second-order polynomial regression (Sargeant et al. 1981; Hintzy et al. 1999). Maximum power (P max) was identified as the apex of the power–velocity relationship. V opt was the pedalling rate at which P max occurred. The linear torque–velocity relationship was used to calculate optimal torque (T Vopt, torque at V opt), half the optimal pedalling rate (0.5V opt) and the torque at 0.5V opt (T 0.5Vopt). An example of typical data is presented in Fig. 1.

Torque (open circles) and power–velocity (filled circles) relationships of a subject, fitted by values obtained from three sprints of 6-s duration. Determination of maximal power (P max ), optimal cycling rate (V opt ), half optimal cycling rate (0.5V opt ) and corresponding torques (T Vopt and T 0.5Vopt )
After 10 min rest, VO2max was determined by a continuous incremental test on the cycle ergometer. Subjects warmed up again for 5-min at 60–80 W (70 rpm) and then the work rate was increased by 30–50 W (according to training status and endurance fitness level) every 3-min until exhaustion. Expired gases were collected during the last 30-s of each stage in Douglas bags through a low-resistance Hans Rudolph valve 2700 (Kansas City, Mo., USA). Subsequent volume determinations were carried out using a balanced Tissot spirometer. The O2 and CO2 fractions were determined by means of S3A/I Ametek (Pittsburgh, Pa., USA) and D-Fend Datex (Helsinki, Finland) analysers, respectively. The gas analysers were calibrated with gas mixtures of compositions determined by the Scholander method. All gas volumes were corrected for STPD conditions. Blood lactate concentrations ([La]b) were measured after completion of the exercise. Blood samples from fingertip were diluted in a haemolizing solution and stored at 5–10°C. The measurements were carried out with a Y.S.I. 2300 lactate analyser (Yellow Spring Instrument, Yellow Springs, Ohio, USA). The highest V̇O2 measured during exercise was considered as VO2max, provided the heart rate (HR, which was monitored throughout the test) was more than 90% of its maximum age-predicted value, the blood lactate concentration measured at the end of this period was >9 mmol.l−1 and respiratory exchange ratio (RER=V̇CO2/V̇O2) was >1.1. VO2max was attained during the last cycling period for all subjects.
Days 2 and 3. Submaximal protocol and supramaximal intermittent exercises
Subjects performed, in random order, two sessions at pedalling rates of V opt and 0.5V opt. Each session consisted of a submaximal incremental exercise, followed by a 30-min recovery period and then a further supramaximal intermittent exercise bout (Fig. 2). After a period of at least 3 days the exercise protocol was repeated using the alternative pedalling rate.

The testing protocol is composed of 12 5-s sprints with 45-s of recovery between each trial. Subjects performed a submaximal cycling exercise at 15, 30, 40, 50 and 60% of their maximal aerobic power (MAP) before repeated sprints. Six Douglas bags (S1 to S6), each for one sprint and the corresponding rest period. LaS1, LaS2, La0, La6, La12: blood lactate samples
During submaximal cycling, each subject performed five continuous 4-min stages at power rates corresponding to 15, 30, 40, 50 and 60% of maximal aerobic power (MAP) at constant velocity (V opt or 0.5V opt). Expired gases were collected during the last 30-s of each stage and the V̇O2–power output relationships fit to a linear function (Gaesser and Brooks 1975; Francescato et al. 1995; Ferguson et al. 2000, 2001; Martin et al. 2002). The Y-intercept of these linear regressions was used to determine VO2int, which corresponded to the oxygen cost of moving the legs at both target velocities including resting V̇O2 (VO2int, at V opt and 0.5V opt). Blood lactate concentrations were measured at rest (LaS1) and after (LaS2) the submaximal exercise with the previously described method.
After 30-min of recovery, the subjects performed a series of 12 repeated 5-s sprints on the cycle ergometer interspersed by 45-s rest periods. The individual friction loads (T Vopt, T 0.5Vopt) were determined by preliminary F–V tests: based on the linear torque–velocity relationship (Fig. 1), these values were determined in order to allow subjects to attain both target cycling rates: V opt and 0.5V opt. Before the first sprint, subjects pedalled until the flywheel turned at the predetermined velocity corresponding to the pedalling rate V opt or 0.5V opt (no friction load: lever arm off). During the series, the friction load was applied (lever arm on) at the beginning of the sprint and was taken off at the end. During the 45-s rest periods the low residual internal friction force and the high inertia of the flywheel allowed an almost constant flywheel velocity. This protocol avoided the acceleration phase and made it possible to maintain an almost constant pedalling rate during the maximal effort. The mechanical variables were recorded for each bout. Considering the progressive adaptation of V̇O2 at the onset of the series, expired gases were collected only during the last six sprints with six Douglas bags (one bag for the 5-s sprint plus the 45-s rest). During this period, all ventilatory variables were measured and averaged over 50-s. Blood lactate measurements were performed before the first sprint and immediately after the 6th and 12th sprints (La0, La6, La12). Heart rate was monitored throughout the test in order to check the steady state of the cardiac response averaged for each work–recovery cycle. After the completion of 12 consecutive sprints of each session, subjects stopped pedalling and were asked to indicate the central rate of perceived exertion (RPE) for sensations involving the cardiorespiratory system and the local feeling of strain in the thighs (Hagen et al. 1993). The degree of subjective cardiorespiratory perception of the effort was assessed using the unmodified 15-grade (from 6 to 20) Borg RPE scale. Local RPE used the Borg's category-ratio scale from 0 to 10 (CR-10 scale), which appeared more appropriate to estimate the perception of exertion for the legs during each maximal repetitive cycling effort. Moreover, using two different scales prevented the subjects selecting the same number for both the local assessment and the central assessments.
Day 4. EMG measurement
Another session was organized to evaluate the activation of the main knee-extensor and knee-flexor muscles (VL and BF), during supramaximal cycling at V opt and 0.5V opt. The main purpose of this additional session was to check for uncontrolled variation in EMG signals on recordings made on two different days. After a 10-min warm-up, subjects performed two series of three 5-s sprints interspersed with a 45-s rest at both V opt and 0.5V opt (in randomly assigned order). EMG signals from the VL and BF muscles were recorded via bipolar Ag-AgCl surface electrodes (interelectrode distance 1.2 cm), including an amplifier (gain 600) and a band-pass filter (6–600 Hz) (Biochip, Elmatek, Crolles, France) fixed longitudinally over the muscle belly. Raw EMG readings were electronically root mean squared (RMS) with a time averaging period of 25 ms (536AJ, Analog Devise, Norwood, Mass., USA), converted from analogical to digital, and stored on another desktop computer at the same sampling frequency as the mechanical data (200 Hz). According to Hautier et al. (2000), the activation levels of VL and BF were determined by measuring the mean values of the EMG RMS signals between the onset of activation and the end of each burst. The mean EMG RMS were calculated from all the bursts provided by the three sprints performed at the same velocity (15 to 30 EMG bursts).
Statistics
All data were analysed with Statview software and are expressed as means (SD). Linear (torque–velocity and V̇O2–power) and polynomial (power–velocity) regression models were fitted by the least-squares method. One-way analysis of variance (ANOVA) with repeated measures was employed to test the effect of sprint time (5-s) on the values of torque, velocity and power per down-stroke at both velocities. A two-factor (cycling rate × sprint repetition) ANOVA with repeated measures on the second factor was used to test the main effects of cycling rate and condition time throughout the repeated sprints on the physiological variables (ventilation: V̇E, V̇O2, HR, [La]b) as well as the mechanical variables averaged for each sprint (power: P m, velocity: V m). The same analysis was used for EMG RMS results at each velocity. When significant F ratios were found, the means were compared using a Tukey's post-hoc test. Since values of perceptual responses are discontinuous, the differences between V opt and 0.5V opt measurements were evaluated by use of the Wilcoxon signed rank test for paired samples. The P<0.05 level was used for all tests of statistical significance.
Results
F–V test
Mean torque, cycling rate and power values at Vopt and 0.5V opt obtained from individual torque- and power–velocity relationships are presented in Table 1.
Oxygen cost of internal work
Oxygen uptake increased linearly with mechanical power at V opt and 0.5V opt (Fig. 3). The regression coefficients averaged 0.991 (0.007) for both cycling rate conditions. Mean values of VO2int were significantly higher at V opt than at 0.5V opt [1.068 (0.205) l.min−1 versus 0.365 (0.061) l.min−1, respectively; P<0.001]. These values represented 23.4 and 8% of VO2max respectively. Post-exercise LaS2 were 3.51 (1.21) mmol.l−1 at V opt and 1.82 (0.47) mmol.l−1 at 0.5V opt.

Typical relationships between oxygen uptake (V̇O 2 ) and power output of a subject during the incremental submaximal exercise test at V opt (filled circles) and at 0.5V opt (open circles). The linear regression for V̇O2–power relationship is calculated from all data (15, 30, 40, 50 and 60% of MAP)
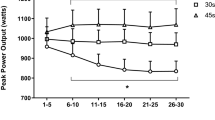
Mechanical data
Figure 4 shows a typical cycling rate, torque and power time course obtained for one subject during two 5-s sprints at V opt and at 0.5V opt. Mean cycling rate and power values for each 5-s sprint (P m, V m) at V opt and 0.5V opt are presented in Table 2. P m measured at 0.5V opt during sprints 1 and 2 were significantly lower than those measured in the subsequent sprints (P<0.05), but no differences were found from the third to the last sprint. At V opt, V m and P m data did not show a significant sprint repetition effect. The mean values of cycling velocity and power produced throughout the set of 12 sprints at V opt [116.6 (4.7) rpm; 924.6 (73.9) W] and at 0.5V opt [60.6 (4.9) rpm; 689.2 (61.8) W] were not significantly different from those expected from Table 1 (NS). The sum of work produced during the last six sprints (W tot) was significantly higher by 35.8% in the set performed at V opt compared to the set at 0.5V opt (P<0.001; Fig. 5).

Typical example of velocity–time (a), torque–time (b) and power–time (c) curves obtained for a subject during two sprints performed at V opt (filled squares) and 0.5V opt (open squares)

Total work output (W tot ), total oxygen consumption (V O2tot ) and blood lactate accumulation (Δ[La] b ) during the final six sprints for two different pedalling rates (V opt, 0.5V opt). ***Significantly different from V opt (P<0.001)
Metabolic variables
Figure 6 shows average V̇O2, V̇E and HR responses for each work–recovery period obtained in the last 5-min of both series (from sprint 7 to sprint 12). The repeated-measures ANOVA on HR did not show a significant sprint number effect at the two pedalling frequencies; mean values on the last six sprints were 156 (12) beats.min−1 for V opt and 145 (18) beats.min−1 for 0.5V opt (P<0.001). No differences were found in V̇O2 and V̇E from the 7th to the 11th sprint, but a slight unexplained decrease was observed in the 12th sprint at V opt and 0.5V opt (P<0.05). As the value of this decrease was low (~4%) and appeared at both velocities, and considering that there was no clear general trend for V̇O2 to increase or to decrease with sprint repetition, the V̇O2 of sprint 12 was taken into account to calculate the total cumulated O2 consumption during the last six sprints (VO2tot). Data showed that V opt condition resulted in a significantly higher VO2tot by 21% compared to 0.5V opt (P<0.001; Fig. 5). The mean values of V̇O2 at V opt [2.56 (0.27) l.min−1] and 0.5V opt [2.11 (0.28) l.min−1] represented approximately 56% and 46% of the subjects' VO2max respectively (P<0.001; Fig. 6).

Mean time course for oxygen uptake per minute (V̇O 2 ), pulmonary ventilation (V̇ E ) and peak heart rate (HR) during the final six sprints performed at V opt and at 0.5V opt. *Significantly different from the other sprints (P<0.05)
Blood lactate concentrations during the series of sprint are presented in Fig. 7. [La]b measured at V opt increased significantly (P<0.001) from rest (La0) to the end of the 6th sprint (La6) and from La6 to the end of the last sprint (La12). Similarly, blood lactate concentration measured at 0.5V opt increased significantly from La0 to La6 (P<0.01) and from La6 to La12 (P<0.05). La0 was not significantly different at V opt and 0.5V opt. La6 and La12 were significantly higher at V opt compared to 0.5V opt (P<0.001). Additionally, blood lactate accumulation during the last six sprints (Δ[La]b, Fig. 5) was significantly higher at V opt compared to 0.5V opt (P<0.001).

Blood lactate concentrations at rest (La0), and immediately after the 6th sprint (La6) and the 12th sprint (La12) in both pedalling conditions (V opt and 0.5V opt). ***Significantly different from V opt (P<0.001)
RPE and EMG results
Mean activation levels of the VL and BF muscles were not different from one down-stroke to another during the sprints at both cycling rates. No significant differences were found in EMG RMS values of VL and BF between V opt [514.3 (170.1) mV; 296 (79) mV] and 0.5V opt [501.1 (187.1) mV; 314,4 (84.2) mV]. No significant V opt to 0.5V opt differences were found in RPE response [13.92 (2.64) versus 12.54 (1.9), respectively]. Similarly, CR-10 did not differ significantly between cycling rate conditions [3.85 (1.86) versus 3.96 (2.11) at V opt and 0.5V opt, respectively].
Discussion
Isokinetic intermittent protocol
The present paper describes a method which allowed subjects to produce constant pedal-crank velocity and power output during short-term maximal efforts on a friction-loaded cycle ergometer. The variation in pedal-crank velocity over the 5-s period of each sprint was lower than 5 rpm, which is close to the value of 4% obtained on an isokinetic ergometer (Beelen and Sargeant 1993). Except for variations within the pedal down-strokes (range 2–8 rpm), this protocol nearly reproduced isokinetic conditions. This original utilization of a friction-loaded ergometer was the necessary condition for examining the influence of pedalling frequency on mechanical and concomitant physiological variables during all-out exercise. Similarly, power output remained constant during the 5-s of each sprint at 0.5V opt and declined slightly at the fifth second at V opt (~91% of the maximal value). This confirms the findings of previous studies from Sargeant et al. (1981), McCartney et al. (1983) and Beelen and Sargeant (1991), in which different time courses of power were observed at different cycling rates with a greater and more rapid decrease at the highest velocity. However, the mean cycling rate and power in the 5-s period of each repeated sprint at V opt and 0.5V opt remained similar to the predetermined maximal values obtained from the F–V test (Table 1). As a consequence, this result suggests that during the first 4-s of a single all-out cycling bout, only the variations in velocity and force can explain the observed increases or decreases in power output. Thus, metabolic stores (including CP stores) should no longer be considered as a limiting factor of power production during this period.
Performance was maintained over the series of sprints, at both target velocities, except for the first and second sprint at 0.5V opt (Table 2). This demonstrates that short-term all-out intermittent cycling (5-s/45-s) did not induce muscle fatigue (defined as the failure to maintain a required or expected power output; Edwards 1983), under the two pedalling conditions of our experiments. Gaitanos et al. (1993) and Hautier et al. (1998) observed that maximal intermittent exercises of 6-s and 5-s duration with 30-s and 25-s rest intervals led to 26.6% and 17.9% reductions in mean power respectively. It appears that a 45-s recovery interval would be sufficient to resynthesize metabolic stores, especially in endurance-trained subjects with predominantly fatigue-resistant muscle fibres. However, the final 9.18 mmol.l−1 [La]b observed at V opt might account for an uncompleted recovery resulting in impaired performance if the series of maximal exercises were prolonged beyond 12 sprints. Additionally, it is noteworthy that during the last six sprints oxygen uptake averaged for each work–recovery cycle (50-s) was constant and accompanied the constant power output at both cycling rates (Fig. 6). Associated with the absence of variation in mean heart rate throughout the same period, this stability of the mean physiological responses enabled us to calculate the total O2 consumption (VO2tot) and to relate it to the mechanical conditions of muscle contraction (velocity and power).
Influence of velocity on the energy turnover
It was hypothesized that although both exercises corresponded to maximal efforts, the energy output and the oxygen uptake should increase with increased speed (up to V opt). W tot and VO2tot measured at V opt were respectively 35.8% and 21% higher than those measured at 0.5V opt. Moreover, as [La]b reflects the balance between lactate production and removal and recovery periods are similar in both conditions, the greater Δ[La]b observed at V opt (Fig. 5) suggests that different rates of glycolysis were operating during the two series of repeated sprints. The fact that more work was delivered at V opt is surely not surprising considering levels of power output and the absence of fatigue. However, the parallel influence of pedalling condition on energy demand remained to be interpreted in such maximal exercise. Previous studies done on isolated frog muscle (Fenn 1923; Wilkie 1968) indicated that the energy output (heat + work) of contracting muscle is a function of the imposed load ("Fenn effect"). Since free energy is proportional to the heat and the work produced during a maximal contraction, the greater work performed at V opt may be responsible for greater total O2 consumption and blood lactate accumulation. However, in the present study, other factors could contribute to the elevation of physiological responses.
We measured EMG activity, in order to verify that muscle activation was at a constant level in each velocity condition. EMG RMS of agonist and antagonist muscles (VL and BF) were similar in both conditions. This is line with the fact that the mean P m values developed at both velocities were not significantly different from the maximal values obtained during F–V tests. These results confirm that the subjects performed maximal effort. According to Perrin (1993) and Sargeant (1996), such all-out exercise requires maximal or near-maximal recruitment of both type I and type II muscle fibres, independently of velocity. In addition it was observed that the local thigh muscle perception (CR-10) and the central perceived exertion (RPE) were similar in both conditions, i.e. subjects were equally involved at V opt and 0.5V opt. It is therefore unlikely that the increase in VO2tot (extra V̇O2tot: 2.22 l) and blood lactate accumulation at V opt were due to differences in fibre type recruitment and/or subjects' involvement.
Moreover, when comparing the two pedalling conditions it seems important to study the proportion to which internal power is altered. At 60 rpm, Gaesser and Brooks (1975) and Francescato et al. (1995) measured VO2int values close to that obtained here at 0.5V opt (~60 rpm). Similarly, the mean value obtained at V opt (~117 rpm) was close to that measured at 120 rpm by Beelen and Sargeant (1993). As a consequence, VO2int measured at V opt was almost three times higher than that measured at 0.5V opt (P<0.001). During the last six bouts of 5-s, total VO2int represented 0.534 and 0.183 l at V opt and 0.5V opt respectively. This large difference of 0.351 l corresponded to almost 16% of the extra VO2tot observed at V opt. It must be noted that VO2int assessments take into account the fact that a twofold higher number of relaxation–contraction cycles at V opt might be responsible for an increase in energy consumption by Ca2+ pumping (Bergström and Hultman 1988; Ferguson et al. 2001).
Finally, the following question may be addressed: what fraction of extra VO2tot may be caused by the increase in V̇̇O2 consumed by the respiratory muscles and by the trunk and arm muscles? The energy demand of the respiratory musculature increases as a function of the rise in V̇E. In this study, using the equation of Coast et al. (1993), the difference observed in V̇E between V opt and 0.5V opt (~24 l.min−1) might have resulted in a 0.226 l higher O2 consumption in respiratory muscles at V opt during the period of the final six sprints (VO2resp). This parameter therefore represented slightly less than 10% of the extra VO2tot. The influence of movements of the trunk and upper limbs in both experimental conditions is more open to discussion. On the one hand Beelen and Sargeant (1993) and Francescato et al. (1995) explained that this energy consumption may be slightly larger at the higher pedalling frequencies. On the other hand it could be greater when higher levels of force are developed, i.e. at the lower cycling rate (Marzorati et al. 2000). The present data do not allow a quantitative analysis of this factor.
In conclusion, taking into account the effect of components discussed above (VO2int, VO2resp), extra VO2tot that could be attributed to the increase of W tot passes from 21% to 17%. However, subtracting resting VO2 (Altman and Ditmer 1968; Du Bois and Du Bois 1916) and including a lactate O2 equivalent of 3.30 ml O2.kg−1 per mmol.L−1 increase in [La]b (Margaria et al. 1971), extra VO2tot amounts to 24.1%.
Hypothesis about mechanical efficiency
It is worth noting that, between 0.5V opt and V opt, extra VO2tot remains consistently lower than the increase of W tot. This mismatch between the increase in mechanical work and the increase in metabolic requirement from 0.5V opt to V opt suggests that there is an increase in apparent mechanical efficiency. This is line with the work of Lodder et al. (1991), who states that during repeated contractions of the rat extensor digitorum longus muscle mechanical efficiency exhibits a velocity-dependent pattern that may have the same shape as the power–velocity curve. Concerning cycling exercise, Sargeant and Rademaker (1996) and Sargeant (1999) proposed also a general form of the efficiency/velocity relationships for type I and type II muscle fibres. By reference to this model, all muscle fibres are maximally activated during sprint cycling, and the contribution of type II fibres to the total power output and their mechanical efficiency are both higher at V opt. Thus efficiency would be improved at "optimum" pedal speed (V opt) as it was recently speculated by Bogdanis et al. (1996, 1998).
In conclusion, a method was employed to obtain two constant levels of power production in relation to two different pedalling velocities during brief all-out intermittent exercises performed on a cycle ergometer. Both work production and oxygen uptake remained significantly higher at V opt (38.5% and 21%, respectively) compared with 0.5V opt. The present study showed that during this type of exercise, the metabolic strain, based on V̇O2 and Δ[La]b, is higher when exercise is performed at optimal velocity. This confirms the hypothesis that despite similar maximal activation of agonist and antagonist muscles, the rate of energy turnover during repeated all-out exercises is not a fixed value. In fact the latter is mainly dependent on mechanical conditions of muscle contraction. However, the non-proportional increase in mechanical and physiological variables points out that efficiency is higher at V opt. In the light of the present findings, further studies are needed to investigate the influence of a higher pedalling velocity (i.e. lower power output), which enables dissociation of the effect of cycling velocity from the effect of power output on physiological responses.
References
Altman PL, Ditmer DS (1968) Metabolism, section VI. FASEB, Bethesda, Md., pp 327–406
Arsac LM, Belli A, Lacour J-R (1996) Muscle function during brief maximal exercise: accurate measurements on a friction-loaded cycle ergometer EurJ Appl Physiol 74:100–106
Balsom PD, Seger JY, Sjödin B, Ekblom B (1992) Maximal-intensity intermittent exercise: effect of recovery duration. Int J Sports Med 13:528–533
Bangsbo J, Gollnick PD, Graham TE, Juel C, Kiens B, Mizuno M, Saltin B (1990) Anaerobic energy production and O2 deficit-debt relationship during exhaustive exercise in humans. J Physiol (Lond) 422:539–559
Beelen A, Sargeant AJ (1991) Effect of fatigue on maximal power output at different contraction velocities in humans. J Appl Physiol 71:2332–2337
Beelen A, Sargeant AJ (1993) Effect of prior exercise at different pedalling frequencies on maximal power in humans. Eur J Appl Physiol 66:102–107
Bergström M, Hultman E (1988) Energy cost and fatigue during intermittent electrical stimulation of human skeletal muscle. J Appl Physiol 65:1500–1505
Bogdanis GC, Nevill ME, Boobis LH, Lakomy HKA (1996) Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J Appl Physiol 80:876–884
Bogdanis GC, Nevill ME, Lakomy HKA, Boobis LH (1998) Power output and muscle metabolism during and following recovery from 10 and 20-s of maximal sprint exercise in humans. Acta Physiol Scand 163:261–272
Coast JR, Rasmussen SA, Krause KM, O'Kroy JA, Loy RA, Rhodes J (1993) Ventilatory work and oxygen consumption during exercise and hyperventilation. J Appl Physiol 74:793–798
di Prampero PE, Ferretti G (1999) The energetics of anaerobic muscle metabolism: a reappraisal of older and recent concepts. Respir Physiol 118:103–115
Du Bois D, Du Bois E (1916) Clinical calorimeter: a formula to estimate the approximate surface area if height and weight be known. Arch Intern Med 17:863–871
Durnin JVGA, Rahaman MM (1967) The assessment of the amount of fat in the human body from measurements of skinfold thickness. Br J Nutr 21:681–689
Edwards RHT (1983) Biochemical bases of fatigue in exercise performance: catastrophe theory of muscular fatigue. In: Biochemistry of exercise. Human Kinetics, Champaign, Ill., pp 3–28
Fenn WO (1923) A quantitative comparison between the energy liberated and the work performed by the isolated sartorius muscle of the frog. J Physiol (Lond) 58:173–203
Ferguson RA, Aagaard P, Ball D, Sargeant AJ, Bangsbo J (2000) Total power output generated during dynamic knee extensor exercise at different contraction frequencies. J Appl Physiol 89:1912–1918
Ferguson RA, Ball D, Krustrup P, Aagaard P, Kjaer M, Sargeant AJ, Hellsten Y, Bangsbo J (2001) Muscle oxygen uptake and energy turnover during dynamic exercise at different contraction frequencies in humans. J Physiol (Lond) 536:261–271
Francescato MP, Girardis M, di Prampero PE (1995) Oxygen cost of internal work during cycling. Eur J Appl Physiol 72:51–57
Gaesser GA, Brooks GA (1975) Muscular efficiency during steady-rate exercise: effects of speed and work rate. J Appl Physiol 38:1132–1139
Gaitanos GC, Williams C, Boobis LH, Brooks S (1993) Human muscle metabolism during intermittent maximal exercise. J Appl Physiol 75:712–719
Hagen KB, Hallen J, Harms-Ringdahl K (1993) Physiological and subjective responses to maximal repetitive lifting employing stoop and squat technique. Eur J Appl Physiol 67:291–297
Hautier CA, Belli A, Lacour JR (1998) A method for assessing muscle fatigue during sprint exercise in humans using a friction-loaded cycle ergometer. Eur J Appl Physiol 12:231–235
Hautier CA, Arsac LM, Deghdegh K, Souquet J, Belli A, Lacour JR (2000) Influence of fatigue on EMG/force ratio and cocontraction in cycling. Med Sci Sports Exerc 32:839–843
Hill AV (1938) The heat of shortening and the dynamic constants of muscle. Proc Roy Soc B 126:136–195
Hintzy F, Belli A, Grappe F, Rouillon JD (1999) Optimal pedalling velocity characteristics during maximal and submaximal cycling in humans. Eur J Appl Physiol 79:426–432
Knuttgen HG, Saltin BG (1972) Muscle metabolites and oxygen uptake in short-term submaximal exercise in man. J Appl Physiol 32:690–694
Lakomy HKA (1986) Measurement of work and power output using friction-loaded cycle ergometers. Ergonomics 29:509–517
Linossier MT, Dormois D, Fouquet R, Geyssant A, Denis C (1996) Use of the force-velocity test to determine the optimal braking force for a sprint exercise on a friction-loaded cycle ergometer. Eur J Appl Physiol 74:420–427
Lodder MAN, de Haan A, Sargeant AJ (1991) Effect of shortening velocity on work output and energy cost during repeated contractions of the rat EDL muscle. Eur J Appl Physiol 62:430–443
Margaria R, Aghemo P, Sassi G (1971) Lactic production in supramaximal exercise. Pflugers Arch 326:152–161
Martin R, Hautier C, Bedu M (2002) Effect of age and pedalling rate on cycling efficiency and internal power in humans. Eur J Appl Physiol 86:245–250
Marzorati M, Perini R, Milesi S, Veicsteinas (2000) Metabolic and cardiorespiratory responses to maximal intermittent knee isokinetic exercise in young healthy humans. Eur J Appl Physiol 81:275–280
McCartney N, Heigenhauser GJF, Jones NL (1983) Power output and fatigue of human muscle in maximal cycling exercise. J Appl Physiol 55:218–224
Perrin DH (1993) Isokinetic exercise and assessment. Human Kinetics, Champaign, Ill., pp 52–53
Sargeant AJ (1996) Human power output-determinants of maximum performance. In: Marconnet P, Saltin B, Komi P, Poortmans J (eds) Human muscular function during dynamic exercise. Med Sport Sci 41:269–273
Sargeant AJ (1999) Neuromuscular determinants of human performance. In: Whipp BJ, Sargeant AJ (eds) Physiological determinants of exercise tolerance in humans. Portland, London, pp 13–28
Sargeant AJ, Rademaker ACHJ (1996) Human muscle fibre types and mechanical efficiency during cycling. In: Steinacker, Ward (eds) The physiology and pathophysiology of exercise tolerance. Plenum, New York, pp 247–251
Sargeant AJ, Hoinville E, Young A (1981) Maximum leg force and power output during short-term dynamic exercise. J Appl Physiol 51:1175–1182
Seck D, Vandewalle H, Decrops N, Monod H (1995) Maximal power and torque-velocity relationship on a cycle ergometer during the acceleration phase of a single all-out exercise. Eur J Appl Physiol 70:161–168
Widrick JJ, Freedson PS, Hamill J (1992) Effect of internal work on calculation of optimal pedalling rate. Med Sci Sports Exerc 24:376–382
Wilkie DR (1968) Heat work and phosphorylcreatinine break-down in muscle. J Physiol (Lond) 195:157–183
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dorel, S., Bourdin, M., Van Praagh, E. et al. Influence of two pedalling rate conditions on mechanical output and physiological responses during all-out intermittent exercise. Eur J Appl Physiol 89, 157–165 (2003). https://doi.org/10.1007/s00421-002-0764-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-002-0764-4