
Abstract
Platynaspidius maculosus (Weise, 1910) (Coleoptera: Coccinellidae) is a ladybird beetle that preys on aphids and is distributed in China, Taiwan, and Vietnam. Recently, P. maculosus has invaded the southern parts of Japan; it was recorded on Okinawa Island in 1998, in Kagoshima Prefecture in 2006, and in Fukuoka Prefecture in 2010. In citrus groves in Shizuoka City, Central Japan in 2009–2012, the exotic ladybird was monitored by use of yellow sticky traps set inside citrus tree canopies. Adults of P. maculosus were captured in three out of the eight groves in 2010. This capture might be the first record of the ladybird from Honshu Island. Adults were trapped in three consecutive years, 2010–2012, in two groves; this suggests the possibility that the ladybird has become established in Shizuoka City. Adults were almost continuously caught on the traps from late April through October, being more abundant in early autumn (early September to early October), late spring (late April to mid-June), and mid-summer (early July to early August). During these periods, citrus shoots are growing and aphids are abundant. This finding suggests that the exotic ladybird might prey on citrus-infesting aphids, thereby reproducing in citrus groves. Based on the results obtained, potential impacts of the exotic ladybird on native aphidophagous ladybirds are also discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Numerous studies have documented that exotic species severely suppress or displace native species that occupy similar niches, thereby altering community structure (Elton 1958; Williamson 1996; Mack et al. 2000). Some alien aphidophagous ladybird species, such as Coccinella septempunctata L. (established in North America) and Harmonia axyridis Pallas (in North America and Europe), have been shown to cause such disruption, i.e., declines in the abundance of some indigenous ladybird species (Elliott et al. 1996; Colunga-Garcia and Gage 1998; Michaud 2002; Evans 2004; Brown et al. 2011).
Platynaspidius maculosus (Weise, 1910) (Coleoptera: Coccinellidae) is an aphidophagous ladybird (Tao and Chiu 1971) and is distributed in China, Taiwan, and Vietnam (Sasaji and Taniguchi 2003). In recent years, the ladybird has invaded the southern parts of Japan; it was recorded on Okinawa Island in 1989 (Matsubara et al. 1998) and 2002 (Sasaji and Taniguchi 2003), in Kagoshima Prefecture in 2006 (Imasaka 2006), and in Fukuoka Prefecture in 2010 (Kido 2010). In addition, the ladybird was detected in Shizuoka City, in the central part of Honshu Island (main island), Japan, in 2011 (Ishikawa 2011). The exotic ladybird might become established in Japan and adversely affect the abundance of native aphidophagous ladybirds. Ecological attributes of an aphidophagous ladybird species, such as prey species, feeding habitat, and seasonal occurrence, could play an important role in determining its interspecific relationship with other ladybirds. However, currently little information exists on these attributes of the exotic ladybird P. maculosus, even in the countries where it is originally distributed.
In this study, I report results from surveys of adults of the exotic ladybird P. maculosus that were conducted using yellow sticky traps in several citrus groves in the southern part of Shizuoka City in 2009–2012, thereby describing invasion of the city by the ladybird and its geographical distribution. I also present results from monitoring of seasonal changes in the number of the ladybird adults captured on the traps in each citrus grove, thereby estimating seasonal occurrence of the ladybird in citrus groves. Based on the results obtained, I discuss the possibility of establishment of the ladybird in Central Japan, its probable prey and reproduction in citrus groves, and its potential impacts on native aphidophagous ladybird species.
Materials and methods
Ladybird
The adult body is approximately 2–3 mm long. The pronotum is black, with its side parts being light yellow (Fig. 1a). The elytra are light red-orange (ground color), with two pairs of large black spots that surround a single smaller black spot. The pronotum and elytra are densely covered with short silver hairs. The larva has a peculiar feature (Fig. 1b): a flat, oval-shaped body, with the dorsum weakly convex and the entire outer margin very thin, i.e., coccid-like shape (Sasaji 1992a). This larval body is very similar to that of a native aphidophagous ladybird, Phymatosternus lewisii (Crotch), described by Sasaji and Tsubokawa (1983).

Adult (a) and larva (b) of the exotic ladybird Platynaspidius maculosus captured in a citrus grove in Shizuoka City in September 2011
Monitoring sites and method
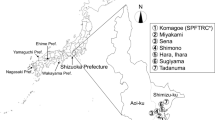
Monitoring was conducted in a total of seven, eight, and eleven groves (each more than 0.1 ha in area) of Satsuma mandarin, Citrus unshiu Marc, in 2009, 2010 (both from May through November), and 2011 and 2012 (both from April through November), respectively (Table 1). The groves are located in the southern part of Shizuoka City, mostly in Shimizu-ku, one of the major citrus-producing regions in Shizuoka Prefecture (Fig. 2). They include groves that are organically managed, a grove in which pesticide application was reduced to approximately 50 % that in conventionally managed groves (hereafter referred to as reduced-pesticide grove), and groves that are conventionally managed. These groves were selected because different levels of pesticide application might affect the occurrence of the ladybird P. maculosus directly and/or indirectly by reducing the abundance of aphids, its probable prey.

Map showing citrus groves where Platynaspidius maculosus adults were surveyed using sticky traps in Shizuoka City in 2009–2012
In 2009, P. maculosus adults were monitored in one organic, one reduced-pesticide, and five conventional groves. In 2010, monitoring was performed in two organic, one reduced-pesticide, and five conventional groves. In 2011 and 2012, this was done in four organic, one reduced-pesticide, and six conventional groves. The organically managed groves were in Sena-1, Shimono, Sugiyama, and Komagoe-1 (hereafter referred to as SE-1, SH, SU, and KO-1) and the reduced-pesticide grove was in Tadanuma-1 (hereafter referred to as TA-1) (Fig. 2). The conventionally managed groves were in Sena-2, Hara, Ihara, Tadanuma-2, Miyakami, and Komagoe-2 (hereafter referred to as SE-2, HA, IH, TA-2, MI, and KO-2).
Three citrus trees, approximately 5 m apart, were chosen randomly at the center of each grove. Inside each tree canopy, a single piece of double-sided, yellow sticky card traps (a yellow adhesive sheet of 100 × 200 mm attached to each side of a yellow acrylic plate of 200 × 200 mm) was set in a vertical position 1.3–1.5 m above the ground using a wooden pole and metal fittings. These traps were replaced at an interval of 12–16 days (generally 14 days). The number of P. maculosus adults captured on the collected traps was counted under a binocular microscope (Olympus SZX12); identification of the ladybird was based on external morphology. Voucher specimens are deposited at Shizuoka Prefectural Research Institute of Agriculture and Forestry, Fruit Tree Research Center, Shizuoka City.
Results
Geographical distribution
Adults of the exotic ladybird P. maculosus were captured on the sticky traps in three out of the eight citrus groves in 2010 and in seven out of the eleven groves in both 2011 and 2012 (Table 1). The adults were not detected in 2009 when the monitored groves were fewer in number (seven groves) and did not involve the grove SE-1 in which many adults occurred in 2010.
Adults of P. maculosus were caught in three consecutive years, 2010–2012, in SE-1 and HA (Table 1). On the other hand, in TA-1 and TA-2, the adults were collected in both 2011 and 2012, but not in the previous 2 years, 2009 and 2010. The adults were not recorded in MI, KO-1, and KO-2, in which monitoring was conducted for 4 years, 2009–2012. The adults were trapped in both organic or reduced-pesticide, and conventional groves, with a tendency of greater total numbers of adults in the former.
Seasonal prevalence
Adults of P. maculosus were captured more or less throughout the monitoring period, from April through November, in the citrus groves (Fig. 3). In particular, in the grove SU, where total P. maculosus adult numbers were largest among the groves, the adults were continuously trapped from late April through October in 2011 (Fig. 3a).

Seasonal changes in the number of Platynaspidius maculosus adults captured on sticky traps in each citrus grove in Shizuoka City in 2010–2012. SU Sugiyama, SE-1 Sena-1, SH Shimono, TA-1 Tadanuma-1, SE-2 Sena-2, HA Hara, IH Ihara, TA-2 Tadanuma-2
In SU, adult numbers increased with time, being most abundant in early autumn (early to mid-September 2011 and late August to mid-September 2012; Fig. 3a). In addition, many adults were caught in late spring (late April to late May 2011) and mid-summer (early to late July 2011 and 2012). In SE-2, on the other hand, the adults were most abundant in late spring (early May to early June 2011 and late April to mid-June 2012; Fig. 3e).
Some adults were also recorded in early autumn (mid-September 2010 and early October 2011 in SE-1; Fig. 3b), late spring (late April to mid-June 2011 in TA-1; Fig. 3d), and mid-summer (early August 2010 and late July 2012 in HA; Fig. 3f). A few adults were also detected during these periods in other groves (Fig. 3c, g, h).
Thus, P. maculosus adults occurred almost continuously from late April through October in the citrus groves in Shizuoka City, being more abundant in early autumn (early September to early October), late spring (late April to mid-June), and mid-summer (early July to early August).
Discussion
In this study, adults of the exotic ladybird P. maculosus were captured on sticky traps in July 2010 in the two citrus groves, SE-1 and IH, in Shizuoka City, which is located in the central part of Honshu Island, Japan (Fig. 3). This is probably the first collection record of P. maculosus from Honshu Island. Subsequently, Ishikawa (2011) collected the ladybird adults in the same city in May 2011. Moreover, the present study confirmed the occurrence of this species several times in 2010–2012, suggesting that the ladybird had already invaded Honshu Island by 2010. Imasaka (2006) recorded adults of this ladybird in 2006 in Kagoshima Prefecture, which is located in the southern part of Kyushu Island and more than 800 km southwest of Shizuoka City. It would be difficult for the ladybird to reach the greatly distant location (Shizuoka City) from Kagoshima Prefecture during a relatively short period (4 years) by naturally expanding its geographical distribution. Artificial transport might have contributed to this long-distance dispersion. Alternatively, the ladybird might be transported to Shizuoka City with plants or other materials (e.g., lumber) directly from the countries where it is originally distributed.
Adults of the exotic ladybird P. maculosus were captured in three consecutive years, 2010–2012, in the two citrus groves, SE-1 and HA, in Shizuoka City (Table 1). This result suggests the possibility of establishment of P. maculosus at the groves and/or their surroundings. Kido (2010) found overwintering adults of the ladybird in Fukuoka Prefecture (Northern Kyushu Island), indicating its establishment on the island. The present study firstly provides the possibility that the ladybird has become established on Honshu Island. Ten species of ladybirds were newly recorded in Japan in 1985–1992; most were considered as exotic species (Sasaji 1992b). The exotic ladybird Adalia bipunctata (L.) was added to these records (Sakuratani 1994). Adalia bipunctata has become established in Osaka City (Sakuratani et al. 2000) and has gradually expanded its geographical distribution in the city and its surrounding areas (Toda and Sakuratani 2006). Results of the present study suggest that the ladybird P. maculosus had just arrived at a location (Tadanuma) in Shizuoka City in 2011, because it had not been found there in the previous two years (Table 1). This implies that P. maculosus is now expanding its geographical distribution. Therefore, the spread of P. maculosus in Shizuoka City and the surrounding areas needs to be monitored. In addition, further studies are urgently required to investigate the current distribution of P. maculosus on Honshu Island, Japan, including the eastern and northern parts. It would be interesting whether P. maculosus could invade and become established in the cooler, northern parts of Japan, because it is originally distributed in warmer, southern regions, such as southern China, Taiwan, and Vietnam (Sasaji and Taniguchi 2003).
Monitoring in the citrus groves showed that P. maculosus adults were more abundant in late spring (late April to mid-June), mid-summer (early July to early August), and early autumn (early September to early October) (Fig. 3). Citrus shoot flush normally occurs three times a year, and aphids that feed on citrus shoots, such as the cotton aphid Aphis gossypii Glover, the spirea aphid Aphis spiraecola Patch, and the brown citrus aphid Toxoptera citricidus (Kirkaldy), are abundant during the flushes, i.e., from early May to mid-June, from early July through August, and from early September to early October (Korenaga et al. 1992). The observed increase in the numbers of trapped P. maculosus adults during these aphid-abundant periods suggests that P. maculosus exploits mostly citrus-feeding aphids as its prey, thereby reproducing in citrus groves. Indeed, larvae of P. maculosus were observed preying on A. spiraecola on young citrus shoots in Shizuoka City in 2011 and 2012, and the larvae successfully became adults when provided with A. spiraecola in the laboratory (S. Kaneko, personal observation). The ladybird P. maculosus might feed on other aphid species infesting citrus shoots, as well as A. spiraecola; prey aphid species of P. maculosus and its prey preference need to be determined.
Numerous studies have documented that exotic aphidophagous ladybird species often adversely affect the abundance of some native ladybird species (Elliott et al. 1996; Colunga-Garcia and Gage 1998; Michaud 2002; Evans 2004; Brown et al. 2011). The exotic ladybird P. maculosus might also cause declines in the abundance of some indigenous ladybird species that exploit aphids on citrus, because it was captured throughout the period during which the aphids occur, i.e., from late April through October (Fig. 3). In addition, larvae of P. maculosus were found in A. spiraecola colonies that were being attended by the ants, Lasius japonicus Santschi or Pristomyrmex punctatus (Smith), on citrus shoots in Shizuoka City, and the larvae were never attacked by the honeydew-collecting ant workers (S. Kaneko, personal observation). This suggests that P. maculosus mainly attacks aphids attended by ants on citrus, and that its colonization in citrus groves might significantly reduce the abundance of some ladybird species that are adapted for utilizing ant-attended aphids, such as Phymatosternus lewisii and Scymnus posticalis Sicard (Kaneko 2007).
It has been suggested that the impacts of an exotic aphidophagous ladybird species on native species depend on whether the exotic species has advantages over the natives in terms of prey exploitation competition (Evans 2004) or intraguild predation (Kajita et al. 2000; Michaud 2002; Snyder et al. 2004; Yasuda et al. 2004). In addition, reproductive interference between exotic and native aphidophagous ladybirds might affect their abundance and distribution, as demonstrated for two species of native, closely related aphidophagous ladybirds (Suzuki et al. 2012). Alternatively, an exotic aphidophagous ladybird might be able to coexist with natives due to temporal and spatial segregation, as suggested by Toda and Sakuratani (2006). Further studies are necessary on the ecological characteristics of the exotic ladybird P. maculosus, including prey and habitat preference, prey-finding ability and consumption rate, growth rate and fecundity, occurrence and direction of intraguild predation with native aphidophagous ladybirds, and reproductive interference with native, closely related species, such as P. lewisii. Impacts of P. maculosus in fields could be assessed by comparing the abundance of native aphidophagous ladybirds prior to and following its invasion. Therefore, investigation of native aphidophagous ladybird assemblages also needs to be started immediately in regions not yet invaded by P. maculosus.
References
Brown PMJ, Frost R, Doberski J, Sparks T, Harrington R, Roy HE (2011) Decline in native ladybirds in response to the arrival of Harmonia axyridis: early evidence from England. Ecol Entomol 36:231–240
Colunga-Garcia M, Gage SH (1998) Arrival, establishment, and habitat use of the multicolored Asian lady beetle (Coleoptera: Coccinellidae) in a Michigan landscape. Environ Entomol 27:1574–1580
Elliott N, Kieckhefer R, Kauffman W (1996) Effects of an invading coccinellid on native coccinellids in an agricultural landscape. Oecologia 105:537–544
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Evans EW (2004) Habitat displacement of North American ladybirds by an introduced species. Ecology 85:637–647
Imasaka S (2006) New record of Platynaspidius maculosus from Kyushu Island in Japan. Gekkan-Mushi 425:45 (in Japanese)
Ishikawa H (2011) New record of Platynaspidius maculosus from Shizuoka Prefecture in Japan. Gekkan-Mushi 487:46 (in Japanese)
Kajita Y, Takano F, Yasuda H, Agarwala BK (2000) Effects of indigenous ladybird species (Coleoptera: Coccinellidae) on the survival of an exotic species in relation to prey abundance. Appl Entomol Zool 35:473–479
Kaneko S (2007) Larvae of two ladybirds, Phymatosternus lewisii and Scymnus posticalis (Coleoptera: Coccinellidae), exploiting colonies of the brown citrus aphid Toxoptera citricidus (Homoptera: Aphididae) attended by the ant Pristomyrmex pungens (Hymenoptera: Formicidae). Appl Entomol Zool 42:181–187
Kido K (2010) Overwintering by Platynaspidius maculosus in Fukuoka Prefecture in Japan. Gekkan-Mushi 475:47 (in Japanese)
Korenaga R, Koizumi M, Ushiyama K, Furuhashi K (1992) A handbook of diseases and insect pests of fruit trees, vol 1. Citrus, loquat, and kiwifruit. Japan Plant Protection Association, Tokyo (in Japanese)
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Matsubara Y, Ueno T, Sasaji H (1998) Platynaspidius maculosus found in Okinawa Island: new record from Japan. Gekkan-Mushi 332:38 (in Japanese)
Michaud JP (2002) Invasion of the Florida citrus ecosystem by Harmonia axyridis (Coleoptera: Coccinellidae) and asymmetric competition with a native species, Cycloneda sanguinea. Environ Entomol 31:827–835
Sakuratani Y (1994) New record of Adalia bipunctata (Linnaeus) (Coleoptera, Coccinellidae) from Japan. Jpn J Entomol 62:627–628
Sakuratani Y, Matsumoto Y, Oka M, Kubo T, Fujii A, Uotani M, Teraguchi T (2000) Life history of Adalia bipunctata (Coleoptera: Coccinellidae) in Japan. Eur J Entomol 97:555–558
Sasaji H (1992a) Descriptions of four coccinellid larvae of Formosa, with the phylogenetic importance (Coleoptera: Coccinellidae). Mem Fac Educ Fukui Univ Ser II (Nat Sci) 42:1–11
Sasaji H (1992b) Recently new added coccinellids. Col News 100:10–13 (in Japanese)
Sasaji H, Taniguchi M (2003) New record of Platynaspidius maculosus (Weise, 1910) from Japan. Gekkan-Mushi 391:13 (in Japanese)
Sasaji H, Tsubokawa K (1983) Supplementary descriptions of the coccinellid larvae of Japan (Coleoptera). Mem Fac Educ Fukui Univ Ser II (Nat Sci) 32:33–66
Snyder WE, Clevenger GM, Eigenbrode SD (2004) Intraguild predation and successful invasion by introduced ladybird beetles. Oecologia 140:559–565
Suzuki N, Osawa N, Nishida T (2012) Asymmetric reproductive interference between specialist and generalist predatory ladybirds. J Anim Ecol 81:1077–1085
Tao C, Chiu SC (1971) Biological control of citrus, vegetable and tobacco aphids. Special Publication No. 10, Taiwan Agricultural Research Institute
Toda Y, Sakuratani Y (2006) Expansion of the geographical distribution of an exotic ladybird beetle, Adalia bipunctata (Coleoptera: Coccinellidae), and its interspecific relationships with native ladybird beetles in Japan. Ecol Res 21:292–300
Williamson M (1996) Biological invasions. Chapman and Hall, London
Yasuda H, Evans EW, Kajita Y, Urakawa K, Takizawa T (2004) Asymmetric larval interactions between introduced and indigenous ladybirds in North America. Oecologia 141:722–731
Acknowledgments
I thank Y. Matsubara for identification of the ladybird Platynaspidius maculosus. This work was partially supported by a grant from the Ministry of Agriculture, Forestry and Fisheries of Japan (Selection of functional biodiversity indicators and development of assessment methods 1111).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kaneko, S. Occurrence of the exotic predatory ladybird Platynaspidius maculosus (Coleoptera: Coccinellidae) in citrus groves in Shizuoka City, Central Japan: seasonal prevalence of adults captured on sticky traps. Appl Entomol Zool 48, 189–194 (2013). https://doi.org/10.1007/s13355-013-0170-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13355-013-0170-x