
Abstract
Bacteriophages are viruses that only infect bacterial cells. Phages are categorized based on the type of their life cycle, the lytic cycle cause lysis of the bacterium with the release of multiple phage particles where as in lysogenic phase the phage DNA is incorporated into the bacterial genome. Lysogeny does not result in lysis of the host. Lytic phages have several potential applications in the food industry as biocontrol agents, biopreservatives and as tools for detecting pathogens. They have also been proposed as alternatives to antibiotics in animal health. Two unique features of phage relevant for food safety are that they are harmless to mammalian cells and high host specificity, keeping the natural microbiota undisturbed. However, the recent approval of bacteriophages as food additives has opened the discussion about ‘edible viruses’. This article reviews in detail the application of phages for the control of foodborne pathogens in a process known as “biocontrol”.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Microorganisms may be beneficial or harmful to us, and we are constantly fighting with the harmful microorganisms to keep them away. One battle is in the food chain against spoilage and poisoning bacteria. Many technologies have been devised to combat bacteria, many a times at the cost of food quality. Heat treatments are associated with deterioration of organoleptic properties, extensive use of sanitizers have led to the development of resistant bacteria, chemical preservatives have negative effect not only on sensory parameters but also on health as many of them are carcinogenic. In addition, these strategies are not infallible which is proved by continuous rise in the number of food borne diseases and increasing loss in food production. Listeria and others have an enormous impact on public health (DuPont 2007). On the other hand most of these strategies cannot be applied to fresh fruits, vegetables and ready to eat products. Hence there is a need for new strategies that fulfill consumer demand for minimally processed foods with fewer chemical preservatives.
Novel technologies like radiation processing, plasma processing, high pressure processing, pulsed electric field and ultrasound are expensive but may be the answer. Another promising approach is the use of natural antagonist towards bacteria to control bacterial contamination in food in a process called “biocontrol” which may tackle the drawbacks of current processing and preservation technology and is likely to be acceptable to consumers.
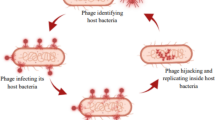
Bacteriophages are obligate parasites of bacteria, using the resources of the bacterial cell to replicate. They are typically highly specific, often being restricted to particular strains within a single bacterial species. However, some bacteriophages have a relatively broad host range, infecting multiple species within a genus and can even infect members of other genera closely related to their normal host. Bacteriophages will multiply when (and only when) their specific bacterial host is present, allowing the use of extremely low input doses (Monk et al. 2010).
This review attempts to describe bacteriophage as biocontrol agents and to review biocontrol strategies for major foodborne pathogens. It will also give an overview of other strategies based on bacteriophage to enhance food safety like phage therapy, biopreservation, biosanitation and use of phage lysin as an alternate to phage application.
Bacteriophage biology
Phages are the largest group of viruses, utilizing species in the Bacteria and Archaebacteria as hosts. Measuring between 20 and 200 nm (Ackermann and DuBow 1987), they are the most abundant form of life on the planet with estimated 1031phages in the biosphere (Kutter and Sulakvelidze 2005).
Phages may be roughly categorized by shape into tailed, polyhedral (icosahedral or quasiicosahedral bodies), filamentous, and pleomorphic phages (Ackermann and Prangishvili 2012). In tailed phages the tail fibers contain proteins that recognize molecules on the surface of bacterial cell walls, which provide the ability to attach only to host cells (Ackermann 2009; Guttman et al. 2005). Most tailed phages are stable in the pH range from 5 to 9 and are inactivated by heating at 60 °C for 30 min (Ackermann 2007).
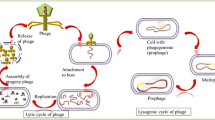
The phage life cycle can be one of two types, the productive or virulent cycle and the temperate or lysogenic cycle (Ackermann and Prangishvili 2012; Guttman et al. 2005). According to this, phages are classified as lytic (virulent) or lysogenic (temperate).
Lytic phages infect bacterial cells causing inhibition of host metabolism and subverting it to the production of phage progeny. The lytic cycle results in the lysis of the bacterium accompanied by the release of multiple phage particles. The new progeny phages produced by the host bacterium spread to infect other cells. The time for the whole cycle is usually within 1–2 h and the number of phage produced depends upon the phage type (Guttman et al. 2005).
Some phages infect cells and incorporate their nucleic acid into the genome of the host cell or exist as an episomal element, leading to a permanent association as a prophage with the cell and all its progeny. During lysogeny, phages neither produce virions nor lyse bacteria. These phages are called temperate, and the cells that harbor a prophage are known as lysogenic. The lysogenic relationship between a temperate phage and its host bacterium provides a safe home to the temperate phage genome, blocks replication of non-virulent homologous phages, and has the potential to alter the phenotype of the host cell (Gill and Abedon 2003).
Characteristics of phage for food application
Phages intended for use in food should be strongly lytic. Their host range should cover all epidemiologically important strains of the target microorganism. They should also display minimum transduction, a process wherein host DNA is packaged into phage heads, rather than phage DNA (Ikeda and Tomizawa 1965). Phages selected should be stable within the intended use environment (Gill and Abedon 2003). In addition to all these phages should also have a broad host range, however this limitation can be mitigated using phage cocktails (McIntyre et al. 2007).
Influence of phage and host concentration on biocontrol
Historically, most research on phage biocontrol has been done in liquids and usually with a high concentration of pure target bacteria (Hagens and Loessner 2010). In liquid environments, thermal motion-driven particle diffusion and mixing due to either fluid flow or active swimming (bacterial motility) increase the likelihood of phages to encounter and infect susceptible host bacteria (Murray and Jackson 1992). When it comes to food applications, there are two major obstacles. First, a significant portion of targeted foods is solid rather than liquid in nature. Second, bacterial contamination would likely occur at very low numbers due to the expected high hygiene standards in place (Hagens and Loessner 2010). This problem can be overcome by inundating the food environment with overwhelming number of phages thereby increasing the chances of phage - target bacteria interaction (threshold of approximately 1 × 108 PFU/ml). In other words, low numbers of phages are unlikely to infect low numbers of bacteria simply because phages and bacteria are unlikely to come into contact with each other. The bacterial host concentration is not a limiting factor if the critical concentration of phage numbers is reached and is able to cover the entire available surface of the targeted food matrix (Hagens and Loessner 2010). Experimental verification of this claim has been achieved when a Salmonella phage (P7) was incubated with its respective host at 24 °C for up to 2 h in Luria-Bertani (LB) broth at varying ratios of phage and host cell concentrations, and the surviving host cells were counted (Bigwood et al. 2009). It was observed that inactivation of Salmonella by P7 seemed to be independent of the host concentration, with nearly complete inactivation occurring at a phage concentration of around 5 × 108 PFU/ml. This was again supported by studies on the control of spoilage bacteria on meat surfaces, which suggest that phages can be effective biocontrol agents when the population of host cells is as low as 46 CFU/cm2 (Greer 1988). Hence, the requirement of a minimum bacterial density as a prerequisite for successful phage biocontrol was not accepted (Kasman et al. 2002).
The results of phage-mediated inactivation of food borne pathogens in some reports using high phage concentrations may be due to lysis from without (Delbruck 1940). Lysis from without occurs when host cells to which numerous phage particles are adsorbed are inactivated rapidly in the absence of phage replication. In E. coli phage T4, this “lysis from without phenomenon” is mediated by a lysozyme on the baseplate (Abedon 1999). It occurs when more than 100 phages are adsorbed on a bacterial cell, which is followed by swelling and bulging of the membrane within 5–10 min after adsorption. Finally, this results in the formation of holes in the cell wall, through which cytoplasmic contents may escape (Tarahovsky et al. 1994).
Bacteriophage biocontrol of foodborne pathogens
There has been much success to control pathogens using bacteriophage. Table 1 summarizes a list of studies on bacteriophage biocontrol of foodborne pathogens. However, this list is not exhaustive.
All postharvest interventions to control E. coli O157:H7 have been successful. During a recent evaluation of the control of E. coli O157:H7 in broths, O’Flynn et al. (2004) reported that phages could eliminate the bacteria at temperatures of 30 or 37 °C where the organism was growing but could not lyse the cells in the absence of growth at 12 °C. Therefore it can be said that the replication of E. coli phage requires the metabolic processes associated with host cell growth.
Studies on reduction of Campylobacter contamination from chicken skin, raw and cooked beef have also been successful. The lysis from without was seen to be the mechanism of Campylobacter reduction on chicken skin as a 107 concentration of phage caused a 2-log CFU reduction where as 105 and 103 failed to reduce CFU.
All Salmonella phages reported have been able to decrease the number of viable cells present on raw meats, processed and ready-to-eat foods, and fresh products. Furthermore, the combined treatment of phage and Enterobacter asburiae, a strain exhibiting antagonistic activity against Salmonella, to control this pathogen on tomatoes, mung bean sprouts, and alfalfa seeds, represents a highly promising, chemical-free approach.
Phage and nisin combination used to control Listeria in ground beef revealed to be ineffective. This strategy had a synergistic effect once added to melon and apple resulting in an improved reduction of Listeria compared to phage or nisin alone. The efficacy of phage-nisin mixture was however significantly reduced in apples on the account of a decline of phage numbers possibly due to low pH. Other studies have used phage P100, which was highly effective in inhibiting Listeria growth.
Post harvest applications in pasteurized milk show that the use of combined phage treatments with nisin and high hydrostatic pressure could synergistically be used to reduce Staphylococcal contamination (Martınez et al. 2008; Tabla et al. 2012). Inactivation of S. aureus has also been accomplished in both fresh and hard-type cheeses using a phage cocktail during cheese manufacturing (Bueno et al. 2012).
Besides the above five pathogens, several other foodborne pathogens are responsible for illnesses, hospitalizations, and deaths, such as Clostridium spp., Shigella spp., Vibrio spp. and Cronobacter sakazakii. Only few studies have been done to control them using phage biocontrol.
Phage therapy
This is a strategy where phage treatment is applied preharvest during plant and animal growth to reduce the probability of plant or animal disease and to prevent the contamination of human pathogens in the food produced.
Many studies aimed at assessing the ability of phage to eliminate bacterial pathogens from food of plant origin and to control plant diseases have been carried out. Several promising trials have been carried out to control plant diseases such as bacterial blotch, bacterial spot, and fire blight in cultivated mushrooms, tomato, and apple. One of the earliest use of phage therapy to control bacterial spots in stone fruits was done by Civerolo and Kiel in 1969. He observed 42 % reduction of Xanthomonas arboricola pv. pruni on peach leaves when phage were applied prior to infection. Although phage application after infection had no effect. In another study by Ravensdale et al. (2007), Pectobacterium carotovorum load on Calla lily was reduced by phages. The phages however, were inactivated by fertilizer solutions. There have been many such successful interventions, but it should be remembered that the application of phages in an open field is associated with some difficulties such as uncontrolled environmental factors including temperature, sun exposure and humidity, uneven phage distribution and allocation, and rapid inactivation of the applied phages (Maura and Debarbieux 2011).
Phage treatment of food-producing animals reduces the probability of contamination of the resulting food products during processing. Risk assessment models indicate that a 1 and 2-log reduction in the number of pathogens shed in feces of the slaughtered animal could reduce the risks to the consumers by 45 and 75 %, respectively. For example, it is estimated that a reduction of 2 log on the Campylobacter loads in poultry intestines is sufficient to diminish 30 fold the incidence of campylobacteriosis associated with consumption of chicken meals (Rosenquist et al. 2003). Carvalho et al. (2010) administered campylobacter phage to poultry by oral gavage and reduced levels of C. coli and C. jejuni in feces by 2 log CFU/g. Studies have also been conducted on cattle to reduce fecal shedding of E. coli O157:H7 (Sheng et al. 2006). Many other trials have been conducted with success for reducing intestinal colonization and fecal shedding of E. coli, Salmonella and Campylobacter (Greer 2005).
Biosanitation
In food industry, biofilms are found on the surfaces of equipment used, for example, in the food handling, storage, or processing, especially on the surfaces that are not easy to clean or to sanitize.
Roy et al. (1993) studied the effectiveness of different phages to remove Listeria from stainless steel and polypropylene surfaces. They found that phage treatment alone was able to achieve approximately a 3-log cycle decrease in cell number. In another study Montanez-Izquierdo et al. (2012) evaluated Listeria phage P100 to control biofilm formation by L. monocytogenes on stainless steel surfaces and found a mean reduction of 5.29 log CFU/cm2. Apart from controlling Listeria biofilms, Campylobacter biofilms were successfully removed from the surface of glass (Siringan et al. 2011) and growth of E. coli O157:H7 was controlled using phage mixture BEC8 on stainless steel and ceramic tiles (Viazis et al. 2011). Use of phage for biosanitation is promising although very challenging due to the diversity of bacteria found in different environments (Sillankorva et al. 2012).
Biopreservation
Biopreservation is the use of bacteriophage as a preservative in perishable manufactured foods to extend its shelf life. Phages are excellent as food biopreservation agents since they are reported to lyse their hosts at temperatures as low as 1 °C (Greer 1982, 1988) limiting the growth of pathogenic and spoilage bacteria on even refrigerated foods (specially psychrotrophic bacteria).
The role of phages in fish and red meats have been recognized for some time. However, their role as food preservatives has been explored only recently. Research on fishes (Delisle and Levin 1969) led to the discovery of phages in fish fillets that were active against psychrophilic spoilage pseudomonads and Shewanella putrefaciens of marine origin. However, those phages were evaluated only from the perspective of strain differentiation by means of a phage-typing scheme, and there has been no further work to examine them as biopreservation agents in fishes.
Preservative effects of Pseudomonas phages in raw chilled beef have also been examined. The retail shelf life of raw chilled beef was extended significantly after Pseudomonas specific lytic phage application (Greer 1988). However, when similar work was carried out using naturally contaminated beef samples and Pseudomonas phage mixture, the shelf life was not significantly affected (Greer 2005). This may be due to the narrow specificity of the used phages that were unable to infect all the spoilage bacteria present.
Another attempt has been made to investigate the ability of Brocothrix thermosphacta lytic phage to control the growth of its host and extend the shelf life of pork adipose tissue (Greer 2005). It was found that bacterial counts were reduced after 2 days of storage at both 2 and 6 °C but the growth of phage-sensitive and resistant strains were detected after this period. However, phage treatment extended the shelf life from 4 days in the control samples to at least 8 days.
Application of phage lysin in food
Lysins are enzymes produced by lytic phages, which play role in the degradation of the bacterial cell wall through targeting its various peptidoglycan bonds to allow the newly formed progeny phages to be released from the host cell (Borysowski et al. 2006). Because lysin enzymes attacks the cell wall peptidoglycan, they are highly effective against Gram-positive bacteria when added externally and may be used as biocontrol agents to enhance food safety (Fischetti 2008). Lysins generally have a narrow spectrum activity restricted to its host species. An exception is an enterococcal phage lysin that not only lyses enterococci but also Streptococcus pyogenes, group B streptococci, and S. aureus, making it one of the broadest acting lysins identified so far (Yoong et al. 2004).
Lysin can be added as a purified protein directly to food or feed. For example the growth of Staphylococcus aureus in pasteurized milk was controlled by addition of purified lysin at 37 °C (Obeso et al. 2008). Forty-eight strains of Clostridium perfringens were lysed by murein hydrolase (lysin) enzyme that is produced by C. perfringens phage j3626 (Zimmer et al. 2002). Another technique of using lyisn is via lysin- secreting recombinant bacteria (Borysowski et al. 2006). This was demonstrated in the case of recombinant Lactococcus lactis cells containing listerial lysin encoding genes to lyse L. monocytogenes in the surrounding medium (Gaeng et al. 2000). This study also showed that the expression of functional lysin by L. lactis was detected in the presence of lactose that is used in milk fermentation. These promising results suggested the possibility of using these recombinant starter lactococcal cultures to selectively protect dairy products against L. monocytogenes contamination.
Similar approaches for lysin application were investigated to control the growth of phytopathogenic bacteria. It was shown that when recombinant lysozyme of Erwinia amylovora phage Ea1h was applied on immature pears after inoculation with E. amylovora, disease symptoms such as ooze formation and necrosis were retarded or inhibited (Kim et al. 2004). Alternatively, transgenic plants able to produce lysin enzyme at the intercellular spaces of the plant to kill bacteria at a very early stage of infection could be developed (During et al. 1993).
The absence of bacterial resistance against lysin is considered as a major advantage of using phage lysins (Fischetti 2010), as the bacterial cell would have to modify the structure of its cell wall to avoid enzymatic action. It was found that exposing bacteria to a particular lysin for 40 reproductive cycles did not give any resistant strains (Fischetti 2010). However, the production of lysin is expensive and, moreover, they are relatively unstable large proteins that are prone to proteolysis and lose its activity in some foods (Coffey et al. 2010).
Advantages of bacteriophage biocontrol
There are many advantages of phages over traditional antimicrobials such as antibiotics and sanitizers.
They have a history of safe use, as they are bacterial viruses, infection of mammalian cells is unlikely. All available evidence indicates that their oral consumption is entirely harmless to humans as they represent a normal component of an everyday diet. Oral toxicity tests on rats that were given phages against Listeria monocytogenes at a dose of 2 × 1012 PFU/kg body weight/day showed no signs of abnormality with regards to histological changes, morbidity, or mortality (Carlton et al. 2005). Similar results were found in a human study with E. coli T4 phages that were added to drinking water (Bruttin and Brussow 2005). Individuals with HIV and other immunodeficiency diseases and healthy volunteers have also been intravenously injected with purified phages (e.g., FX174) without any apparent side effects (Atterbury 2009). Indeed, early phage therapy pioneers demonstrated safety by ingesting preparations themselves. The phages used were not only administered orally or superficially, but also were injected intramuscularly, intravenously, and even into the pericardium and carotid artery without any adverse effect being observed.
Phages are highly active and specific against their host with no adverse effects on the intestinal microbiota. Bacteriophages are auto-replicative, hence when bacterial contamination is high, low concentrations of phage can get the desired pathogen reduction. Phage production is relatively simple and has high storage stability under different environmental conditions.
Drawbacks of using phage for food preservation include limited host range, risk for the development of resistant mutants, and the potential for the transduction of virulent characters from one bacterial strain to another. Yet these are not very significant as overcoming them is simple. Phage cocktail can be used when multiple strains of host are present. Spontaneously occurring phage-resistant mutants are not likely to significantly influence treatment efficacy and the complex phage resistance mechanisms common in bacteria can be overcome by screening for broad host range phages and/or use of phage cocktails (Hagens and Loessner 2010). From another perspective, it has been noted that a phage-resistant strain of E. coli O157:H7 had a smaller, more coccoid cellular morphology than the parental strain and it reverted to phage sensitivity within 50 generations (O’Flynn et al. 2004). Likewise, phage-resistant mutant strains of Salmonella Enteritidis lost the O-polysaccharide layer, which is required for phage adsorption, and as a result became avirulent (Santander and Robeson 2007). Finally transduction of virulence can be prevented by choosing phages which show low transduction frequencies. Another disadvantage is that most current research on their efficacy have involved experiments with artificially inoculated foods that do not necessarily reflect the real commercial environments where phages will be applied.
Conclusion
Bacteriophages were created by nature to combat bacteria around us. We have manipulated these bacterial viruses to control and detect bacterial pathogens in food as well as in medicine and veterinary. Research is still needed to thoroughly understand the mechanisms of phage resistance acquired by the hosts and methods to overcome phage resistance.
Although the results of the studies appear to be encouraging, they should be interpreted with caution. For instance, some phage studies have proven that phages are inefficient in reducing their host, such as biocontrol on apple slices. Also, several authors have reported the emergence of phage-resistant phenotypes, but this has not significantly affected the results of the phage trials.
Application and commercialization of phage based technologies have begun and still have a long way to go. To name a few products, Ecoshield™ and Listshield™ are phage preparations by Intralyix, Inc that target E. coli and L. monocytogenes respectively and Listex™ and salmonlex™ are products of Micreos food safety that are active against Listeria and Salmonella.
Although phages are and will be present forever in foods, the consumer’s perception of adding viruses to foods will, arguably, be the most critical hurdle to be overcome in order for phages to be used widely for biocontrol of bacterial pathogens in food (Strauch et al. 2007).
References
Abedon ST (1999) Bacteriophage T4 resistance to lysis-inhibition collapse. Genet Res 74:1–11
Ackermann HW (2007) 5500 phages examined in the electron microscope. Arch Virol 152:227–243
Ackermann HW (2009) Phage classification and characterization. In: Bacteriophages. Humana press, pp 127–140
Ackermann HW, DuBow MS (1987) Viruses of prokaryotes: general properties of bacteriophage. CRC Press, Boca Raton
Ackermann HW, Prangishvili D (2012) Prokaryote viruses studied by electron microscopy. Arch Virol 157(10):1843–1849
Atterbury RJ (2009) Bacteriophage biocontrol in animals and meat products. Microb Biotechnol 2:601–612
Bigot B, Lee WJ, McIntyre L et al (2011) Control of Listeria monocytogenes growth in a ready-to-eat poultry product using a bacteriophage. Food Microbiol 28(8):1448–1452
Bigwood T, Hudson JA, Billington C (2009) Influence of host and bacteriophage concentrations on the inactivation of food-borne pathogenic bacteria by two phages. FEMS Microbiol Lett 291:59–64
Borysowski J, Weber-Dabrowska B, Gorski A (2006) Bacteriophage endolysins as a novel class of antibacterial agents. Exp Biol Med 231:366–377
Bruttin A, Brussow H (2005) Human volunteers receiving escherichia coli phage T4 orally: a safety test of phage therapy. Am Soc Microbiol 49:2874–2878
Bueno E, Garcıa P, Martınez B, Rodrıguez A (2012) Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int J Food Microbiol 158(1):23–27
Carlton RM, Noordman WH, Biswas B, de Meester ED, Loessner MJ (2005) Bacteriophage P100 for control of Listeria monocytogenes in foods: genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul Toxicol Pharmacol 43(3):301–312
Carvalho CM, Gannon BW, Halfhide DE, Santos SB, Hayes CM, Roe JM, Azeredo J (2010) The in vivo efficacy of two administration routes of a phage cocktail to reduce numbers of campylobacter coli and campylobacter jejuni in chickens. BMC Microbiol 10(1):232
Civerolo EL, Kiel HL (1969) Inhibition of bacterial spot of peach foliage by Xanthomonas pruni bacteriophage. Phytopathology 59:1966–1967
Coffey B, Mills S, Coffey A, McAuliffe O, Ross RP (2010) Phage and their lysins as biocontrol agents for food safety applications. Ann Rev Food Sci Technol 1:449–468
Delbruck M (1940) The growth of bacteriophage and lysis of the host. J Gen Physiol 23:643–660
Delisle AL, Levin RE (1969) Bacteriophages of psychrophilic pseudomonads. I. Host range of phage pools active against fish spoilage and fish-pathogenic pseudomonads. Anton Leeuw 35:307–317
DuPont HL (2007) The growing threat of foodborne bacterial enteropathogens of animal origin. Clin Infect Dis 45:1353–1361
During K, Porsch P, Fladung M, Lorz H (1993) Transgenic potato plants resistant to the phytopathogenic bacterium Erwinia carotovora. Plant J 3:587–598
Dykes GA, Moorhead SM (2002) Combined antimicrobial effect of nisin and a listeriophage against Listeria monocytogenes in broth but not in buffer or on raw beef. Int J Food Microbiol 73(1):71–81
El-Shibiny A, Scott A, Timms A, Metawea Y, Connerton P, Connerton I (2009) Application of a group II campylobacter bacteriophage to reduce strains of Campylobacter jejuni and campylobacter coli colonizing broiler chickens. J Food Prot 72(4):733–740
Fischetti VA (2008) Bacteriophage lysins as effective antibacterials. Curr Opin Microbiol 11:393–400
Fischetti VA (2010) Bacteriophage endolysins: a novel anti-infective to control gram-positive pathogens. Int J Med Microbiol 300:357–362
Gaeng S, Scherer S, Neve H, Loessner MJ (2000) Gene cloning and expression and secretion of Listeria monocytogenes bacteriophage-lytic enzymes in Lactococcus lactis. Appl Environ Microbiol 66:2951–2958
Gill J, Abedon ST (2003) Bacteriophage ecology and plants. APSnet Feature
Gill J, Sabour PM, Leslie KE, Griffiths MW (2006) Bovine whey proteins inhibit the interaction of staphylococcus aureus and bacteriophage K. J Appl Microbiol 101(2):377–386
Greer GG (1982) Psychrotrophic bacteriophages for beef spoilage pseudomonads. J Food Prot 45:1318–1325
Greer GG (1988) Effects of phage concentration, bacterial density, and temperature on phage control of beef spoilage. J Food Sci 53:1226–1227
Greer GG (2005) Bacteriophage control of foodborne bacteria. J Food Prot 68(5):1102–1111
Guenther S, Loessner MJ (2011) Bacteriophage biocontrol of Listeria monocytogenes on soft ripened white mold and red- smear cheeses. Bacteriophage 1(2):94–100
Guenther S, Herzig O, Fieseler L, Klumpp J, Loessner MJ (2012) Biocontrol of salmonella typhimurium in RTE foods with the virulent bacteriophage FO1-E2. Int J Food Microbiol 154(1–2):66–72
Guttman B, Raya R, Kutter E (2005) Basic phage biology. In: Kutter E, Sulakvelidze A (eds) Bacteriophages: biology and applications. CRC Press, Boca Raton, p 28
Hagens S, Loessner MJ (2010) Bacteriophage for biocontrol of foodborne pathogens: calculations and considerations. Curr Pharm Biotechnol 11(1):58–68
Holck A, Berg J (2009) Inhibition of Listeria monocytogenes in cooked ham by virulent bacteriophages and protective cultures. Appl Environ Microbiol 75(21):6944–6946
Hooton SPT, Atterbury RJ, Connerton IF (2011) Application of a bacteriophage cocktail to reduce salmonella typhimurium U288 contamination on pig skin. Int J Food Microbiol 151(2):157–163
Ikeda H, Tomizawa J (1965) Transducing fragments in generalized transduction by phage P1. I. Molecular origin of the fragments. J Mol Biol 14:85–109
Kasman LM, Kasman A, Westwater C, Dolan J, Schmidt MG, Norris JS (2002) Overcoming the phage replication threshold: a mathematical model with implications for phage therapy. J Virol 76:5557–5564
Kim W-S, Salm H, Geider K (2004) Expression of bacteriophage fEa1h lysozyme in escherichia coli and its activity in growth inhibition of Erwinia amylovora. Microbiology 150:2707–2714
Kim KP, Klumpp J, Loessner MJ (2007) Enterobacter sakazakii bacteriophages can prevent bacterial growth in reconstituted infant formula. Int J Food Microbiol 115:195–203
Kutter E, Sulakvelidze A (2005) Introduction. In: Kutter E, Sulakvelidze A (eds) Bacteriophages: biology and applications. CRC Press, Boca Raton, p 1
Leverentz B, Conway WS, Alavidze Z (2001) Examination of bacteriophage as a biocontrol method for salmonella on fresh-cut fruit: a model study. J Food Prot 64(8):1116–1121
Leverentz B, Conway WS, Camp MJ et al (2003) Biocontrol of Listeria monocytogenes on fresh-cut produce by treatment with lytic bacteriophages and a bacteriocin. Appl Environ Microbiol 69(8):4519–4526
Leverentz B, Conway WS, Janisiewicz W, Camp MJ (2004) Optimizing concentration and timing of a phage spray application to reduce Listeria monocytogenes on honeydew melon tissue. J Food Prot 67(8):1682–1686
Martınez B, Obeso JM, Rodrıguez A, Garcıa P (2008) Nisin- bacteriophage crossresistance in Staphylococcus aureus. Int J Food Microbiol 122(3):253–258
Maura D, Debarbieux L (2011) Bacteriophages as twenty-first century antibacterial tools for food and medicine. Appl Microbiol Biotechnol 90:851–885
McIntyre L, Hudson JA, Billington C, Withers H (2007) Biocontrol of bacteria: past, present and future strategies. Food N Z 7:25–32
Modi R, Hirvi Y, Hill A, Griffiths MW (2001) Effect of phage on survival of salmonella enteritidis during manufacture and storage of cheddar cheese made from raw and pasteurized milk. J Food Prot 64(7):927–933
Monk AB, Rees CD, Barrow P, Hagens S, Harper DR (2010) Bacteriophage applications: where are we now? Lett Appl Microbiol 51:363–369
Montanez-Izquierdo VY, Salas-Vazquez DI, Rodrıguez-Jerez JJ (2012) Use of epifluorescence microscopy to assess the effectiveness of phage P100 in controlling Listeria monocytogenes biofilms on stainless steel surfaces. Food Control 23:470–477
Murray AG, Jackson GA (1992) Viral dynamics: a model of the effects size, shape, motion and abundance of single-celled planktonic organisms and other particles. Mar Ecol Prog Ser 89:103–116
O’Flaherty S, Coffey A, Meaney WJ, Fitzgerald GF, Ross RP (2005) Inhibition of bacteriophage K proliferation on staphylococcus aureus in raw bovine milk. Lett Appl Microbiol 41(3):274–279
O’Flynn G, Ross RP, Fitzgerald GF, Coffey A (2004) Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157: H7.Applied and. Environ Microbiol 70(6):3417–3424
Obeso JM, Martınez B, Rodrıguez A, Garcıa P (2008) Lytic activity of the recom- binant staphylococcal bacteriophage FH5 endolysin active against staphylococcus aureus in milk. Int J Food Microbiol 128:212–218
Patel J, Sharma M, Millner P, Calaway T, Singh M (2011) Inactivation of Escherichia coli O157:H7 attached to spinach harvester blade using bacteriophage. Foodborne Pathog Dis 8(4):541–546
Ravensdale M, Blom TJ, Gracia-Garza A, Svircev AM, Smith RJ (2007) Bacteriophages and the control of Erwinia carotovora subsp. carotovora. Can J Plant Pathol 29(2):121–130
Rosenquist H, Nielsen NL, Sommer HM, Nørrung B, Christensen BB (2003) Quantitative risk assessment of human campylobacteriosis associated with thermophilic campylobacter species in chickens. Int J Food Microbiol 83(1):87–103
Roy B, Ackermann HW, Pandian S, Picard G, Goulet J (1993) Biological inactivation of adhering Listeria monocytogenes by listeriaphages and a quaternary ammonium compound. Appl Environ Microbiol 59:2914–2917
Santander J, Robeson J (2007) Phage-resistance of salmonella enterica serovar enteritidis and pathogenesis in Caenorhabditis elegans is mediated by the lipopolysaccharide. Electron J Biotechnol 10:627–632
Sharma M, Patel JR, Conway WS, Ferguson S, Sulakvelidze A (2009) Effectiveness of bacteriophages in reducing Escherichia coli O157:H7 on fresh-cut cantaloupes and lettuce. J Food Prot 72(7):1481–1485
Sheng H, Knecht HJ, Kudva IT, Hovde CJ (2006) Application of bacteriophages to control intestinal Escherichia coli O157:H7 levels in ruminants. Appl Environ Microbiol 72(8):5359–5366
Sillankorva SM, Oliveira H, Azeredo J (2012) Bacteriophages and their role in food safety. Int J Microbiol 2012:1–13
Siringan P, Connerton PL, Payne RJH, Connerton IF (2011) Bacteriophage- mediated dispersal of campylobacter jejuni biofilms. Appl Environ Microbiol 77(10):3320–3326
Soni A, Nannapaneni R, Hagens S (2010) Reduction of Listeria monocytogenes on the surface of fresh channel catfish fillets by bacteriophage listex p100. Foodborne Pathog Dis 7(4):427–434
Strauch E, Hammerl J, Hertwig S (2007) Bacteriophages: new tools for safer food? J Verbr Lebensm 2:138–143
Tabla R, Martínez B, Rebollo JE, González J, Ramírez MR, Roa I, Rodríguez A, García P (2012) Bacteriophage performance against staphylococcus aureus in milk is improved by high hydrostatic pressure treatments. Int J Food Microbiol 156(3):209–213
Tarahovsky YS, Ivanitsky GR, Khusainov AA (1994) Lysis of escherichia coli cells induced by bacteriophage T4. FEMS Microbiol Lett 122:195–199
Tomat D, Mercanti Q, Balague C, Quiberoni A (2013) Phage biocontrol of enteropathogenic and Shiga toxin-producing escherichia coli during milk fermentation. Lett Appl Microbiol 57:3–10
Viazis S, Akhtar M, Feirtag J, Diez-Gonzalez F (2011) Reduction of Escherichia coli O157:H7 viability on hard surfaces by treatment with a bacteriophage mixture. Int J Food Microbiol 145(1):37–42
Wagenaar JA, Bergen MAPV, Mueller MA, Wassenaar TM, Carlton RM (2005) Phage therapy reduces Campylobacter jejuni colonization in broilers. Vet Microbiol 109(3–4):275–283
Ye J, Kostrzynska M, Dunfield K, Warriner K (2009) Evaluation of a biocontrol preparation consisting of enterobacter asburiae JX1 and a lytic bacteriophage cocktail to suppress the growth of salmonella javiana associated with tomatoes. J Food Prot 72(11):2284–2292
Ye J, Kostrzynska M, Dunfield K, Warriner K (2010) Control of salmonella on sprouting mung bean and alfalfa seeds by using a biocontrol preparation based on antagonistic bacteria and lytic bacteriophages. J Food Prot 73(1):9–17
Yoong P, Schuch R, Nelson D, Fischetti VA (2004) Identification of a broadly active phage lytic enzyme with lethal activity against antibiotic resistant Enterococcus faecalis and Enterococcus faecium. J Bacteriol 186:4808–4812
Zhang H, Wang R, Bao H (2013) Phage inactivation of foodborne Shigella on ready- to-eat spiced chicken. Poult Sci 92:211–217
Zimmer M, Vukov N, Scherer S, Loessner MJ (2002) The murein hydrolase of the bacteriophage j3626 dual lysis system is active against all tested Clostridium perfringens strains. Appl Environ Microbiol 68:5311–5317
Zuber S, Boissin-Delaporte C, Michot L, Iversen C, Diep B, Brussow H et al (2008) Decreasing Enterobacter sakazakii (Cronobacter spp.) food contamination level with bacteriophages: prospects and problems. Microb Biotechnol 1:532–543
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kazi, M., Annapure, U.S. Bacteriophage biocontrol of foodborne pathogens. J Food Sci Technol 53, 1355–1362 (2016). https://doi.org/10.1007/s13197-015-1996-8
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13197-015-1996-8