
Abstract
This study examined the distribution of wetland plants used in macrophyte-based index of biotic integrity (IBI) metrics to determine the effectiveness of zone sampling in assessing wetland condition. Using sampling data from a previous study of 74 emergent isolated wetlands, macrophyte taxonomic data and resulting IBI metrics were analyzed for various wetland zones and compared against the original site metrics. Zones were defined by parsing each sampling transect (from wetland edge to center) into thirds—creating an outer, intermediate, and inner zone—and constructing two additional zones representing two-thirds of the original sampling area—an outer plus intermediate zone and an intermediate plus inner zone. While a highly-significant decrease in mean species richness was observed from the wetland exterior to interior, there were no significant differences between metric values derived from site data and those derived from zone data. Linear correlations between metrics from the outer zone and site data were highly significant (R 2 > 0.94), but linear correlation strength decreased with the intermediate and inner zones, respectively. Thus, macrophyte-based indicators of biological integrity can be efficiently assessed by sampling the outer third of a wetland—a 66% reduction in the area and effort required to monitor isolated wetland condition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wetland condition assessment occurs worldwide due to the quantified and qualified relationships between condition and function in wetland systems, in addition to legal strictures associated with wetland mitigation in some countries (e.g., De Groot et al. 2006; Reiss et al. 2009; Ma et al. 2011). In the United States, the Clean Water Act (CWA) stipulates that states and tribes assess the condition of their aquatic resources, including wetlands. While few states currently report wetland condition in their CWA 305b reports to Congress (i.e., only 10 in the latest report [US EPA 2009]), most states have developed or are developing measures to quantify the condition of their wetland resources using macrophytes, macroinvertebrates, amphibians, or other organisms (e.g., Ohio—Mack 2004; Minnesota—Gernes and Helgen 1999; North Dakota—DeKeyser et al. 2003). In addition, the US Environmental Protection Agency (US EPA), federal partners, and states are undertaking the 2011 National Wetland Condition Assessment (NWCA), wherein a total of 900 wetland sites will be sampled across the country to provide the first nationwide characterization of wetland condition (Scozzafava et al. 2011). The NWCA methods include macrophyte sampling, as wetland plants have been identified as good indicators of relative wetland condition (e.g., Galatowitsch et al. 1999; US EPA 2002; Cohen et al. 2004; Mack 2004).
Florida has numerous wetlands as a result of low relief and positive water balance (Ewel 1990; Kushlan 1990; Fretwell et al. 1996; Houlahan and Findlay 2004). Small isolated wetlands—those completely surrounded by uplands (Tiner 2003)—in Florida occur where topographic depressions, generally caused by ground slumping in karst topography, receive overland and interstitial runoff from localized watersheds or groundwater (Kushlan 1990). Depending on phreatic water levels and seasonal variation in rainfall, these wetlands may function as ephemeral, temporary, or permanent wetlands (Sun et al. 1995). Isolated wetlands provide a variety of ecological functions, from seasonal breeding habitat for macroinvertebrates and amphibians to ecosystem services, such as maintenance of water quality and quantity (Ewel and Odum 1984; Leibowitz 2003; Lane and D’Amico 2010).
Several studies have developed indices of biotic integrity (IBIs) for wetlands using macrophytes to assess relative condition (US EPA 2002; Lane 2003; Mack 2004; Reiss et al. 2010). As a precursor to an extensive field survey, Doherty et al. (2000) conducted a detailed review of the literature and identified a series of possible plant metrics for use in biological assessment of wetlands. Subsequently, Lane et al. (2003) conducted wetland surveys and analyses and identified five macrophyte metrics for small isolated marshes in Florida that were strongly correlated to both land use-based measures of expected human disturbances (e.g., the Landscape Development Intensity Index [Brown and Vivas 2005]) and to rapid on-site assessments of disturbance (e.g., the Wetland Rapid Assessment Procedure [Miller and Gunsalus 1997]). In his study of 74 emergent marshes, Lane et al. (2003) found the following metrics, which are further described in the Methods section of this publication, to be strongly and significantly correlated with both the Brown and Vivas (2005) landscape disturbance measure and the Miller and Gunsalus (1997) measure of disturbance: 1) percentage of sensitive indicator taxa, 2) percentage of tolerant indicator taxa, 3) percentage of exotic species, 4) the ratio of annual to perennial species, and 5) the mean site coefficient of conservatism value (Wilhelm and Ladd 1988; Cohen et al. 2004; see Lane 2003; Lane et al. 2003; Reiss 2006 for additional information).
Zonation in Wetlands
Plant community zonation in wetlands is a temporal and spatial function of hydrology, which affects plant and microbial respiration, nutrient cycling, and soil redoximorphic potential (van der Valk 1981; Sharitz and Gresham 1998; Cronk and Fennessy 2001). In general, herbs, graminoids, and forbs are more tolerant of flooding throughout the growing season, while most woody taxa are less tolerant of seasonal flooding (Cronk and Fennessy 2001). In depressional wetlands, flooding gradients follow elevation gradients (Penfound 1952; Winchester et al. 1985). The topography of these systems can be simplistically viewed as a very gently-sloping, concavely-shaped conical depression (Hayashi and van der Kamp 2000; Haag et al. 2005; Lane and D’Amico 2010). Variation in ground and surface water input and retention clearly affects edaphic conditions, such as soil moisture and redoximorphic state, and these in turn are deterministic factors affecting the macrophyte ecology of these systems (Penfound 1952; Winchester et al. 1985). At the level of species assemblages, the topographical influence has frequently been expressed as a series of more or less discrete zones characterized by groups of species which have similar habitat preferences and hydrostatic tolerances (van der Valk 1981; Winchester et al. 1985; Cronk and Fennessy 2001). In a study of marshes and wet prairies in southwestern Florida, Winchester et al. (1985) identified six major vegetation zones in relation to basin shape (a surrogate for elevation differences), substrate type, and disturbance by herbivores. They distinguished these zones based on dominant and sub-dominant macrophyte species and found that the zones were related to wetland depths and peat thickness. LaClaire (1995) described vegetation zonation—in north and north-central Florida temporary ponds (<5 ha depressional wetlands) experiencing a recurrent dry phase of varying duration—as dependent primarily on the relative duration of inundation. Numerous researchers have also examined the response of individual taxa to varying inundation regimes in experimental settings (e.g., Megonigal and Day 1992; Ewing 1996; Lockhart et al. 1999; Kercher and Zedler 2004). Summary findings concluded that plant species are not uniformly distributed within wetland systems, and their distributions are related to ecological factors, particularly flooding depth, duration, and timing (van der Valk 1981; McKevlin et al. 1998; Cronk and Fennessy 2001).
Plant Metrics and Zonation
Since many emergent marsh wetlands exhibit distinct zonation, the effect of zonation on macrophyte-based IBIs may be significant depending on placement of sampling quadrants, sampling technique, sampling season, and data analysis (US EPA 2002). Some potential indicator species have evolved traits such that they are found only in areas of near-constant flooding (e.g., Nymphaea spp., Nuphar spp.), often near the center of the wetland. These are frequently obligate wetland species, but it should be noted that indicator status (sensu Reed 1997) implies fidelity to wetland conditions, not necessarily to hydrologic conditions (i.e., status is given based on the fidelity to wetlands versus non-wetlands). The outer wetland zone abutting the upland boundary may be characterized by higher variation in edaphic conditions or a shorter hydroperiod, while an intermediate set of conditions exists in between these zones. More facultative and facultative-upland species are likely to be found in the “outer” and shallower zones, but again these are guidelines with exceptions related to wetland geometry, topography, and climatic conditions (Brock and Casanova 1997).
The aim of this study was to examine the distribution of wetland plants used in macrophyte-based metrics to determine what level of sampling effort (as measured by area) might be needed to accurately assess wetland condition, while minimizing sampling effort. Using macrophyte data collected during a previous study of 74 small isolated emergent marshes (Lane et al. 2003), macrophyte taxonomic data and resulting IBI metrics were analyzed for various zones within each wetland and compared against the original site metrics developed by Lane et al. (2003). Differences in plant IBI metrics (and their components) were evaluated to determine the effectiveness of zone sampling as a cost-reduction measure, asking the question: How many or which zones must be sampled to provide sufficient information to qualify the condition of a wetland?
Methods
Site Sampling
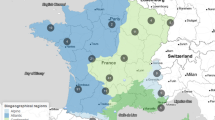
As part of a larger study developing wetland indicators of integrity for isolated marsh and forested wetland systems (Lane 2003; Reiss 2006; Lane and Brown 2007; Reiss et al. 2010), 74 small depressional emergent marsh wetlands were sampled a single time throughout peninsular Florida in either the summer of 1999 or 2000 (Fig. 1). The marshes ranged from 0.2 to 3.9 ha in area, with the majority (68%) between 0.3 and 1.0 ha (mean 0.87 ± 0.63 standard deviation [SD]). Sites were typically flooded when sampled, The wetland systems selected for sampling represented various levels of disturbance (based on visual assessment of surrounding land use), with least-disturbed sites often located in protected areas (e.g., state and national parks) and more disturbed sites found in landscapes with varying degrees of agriculture and urban development—from fallow farm and pasture lands to high-density cattle operations to residential and urban settings (see Lane 2003; Lane et al. 2003; Reiss et al. 2010).

Wetland sampling sites throughout peninsular Florida (n = 74)
At each site, the presence of rooted vascular macrophytes was determined through an extensive (2–3 h) cross-wetland sampling effort. Wetland boundaries were identified through vegetation characterization and soil typing and belted transects established from the approximate upland boundary (i.e., wetland edge) to the wetland center, at each of the four cardinal directions (Fig. 2). Each transect was divided into multiple 5-m2 quadrants measuring 1-m wide by 5-m long, and rooted species presence within each quadrant was recorded. Unknown taxa were field collected, preserved, and later identified by experts from the University of Florida.

Sampling schematic for hypothetical depressional geographically isolated wetland systems showing the three zones tested, as well as the combined outer and intermediate, and intermediate and inner zones
Data Analysis
Using total macrophyte presence found among all four transects within each wetland, Lane et al. (2003; see also Reiss et al. 2010) identified five macrophyte indicators (US EPA 2002) that strongly correlated with expected wetland condition, as determined by the Landscape Development Intensity Index (Brown and Vivas 2005). The five indicator metrics of biotic integrity were: 1) percentage of sensitive indicator taxa (% sensitive; i.e., those with fidelity and specificity to high-quality wetlands); 2) percentage of tolerant indicator taxa (% tolerant; i.e., those with fidelity and specificity to poor-quality wetlands); 3) percentage of exotic species (% exotics; FLEPPC 2001); 4) the ratio of annual to perennial species (A:P ratio); and 5) the mean site coefficient of conservatism value (mean CC). Sensitive and tolerant taxa were identified using indicator species analyses (Dufrêne and Legendre 1997; McCune and Grace 2002), which identified taxa at the species level with specificity and fidelity to the least and most impaired conditions. The first and second metrics were based on the percentage of sensitive and tolerant macrophytes, respectively, found at each site. The third metric was based on the percentage of species at each site that were identified by the Florida Exotic Pest Plant Council as exotic (FLEPPC 2001). The fourth metric was based on the ratio of the richness of annual to perennial species. To support development of the fifth metric, a CC value was assigned to each wetland macrophyte by a group of expert botanists in Florida. The CC provides a number from 1 to 10, representing the fidelity and specificity of a macrophyte to particular conditions, with high-scoring plants being found only in exceptional wetland habitats (Wilhelm and Ladd 1988). The fifth and last metric was the mean CC value for each site.
To identify possible zones for comparative analyses, the four transects from each site were divided into three zones, each representing approximately 33% of the sampling area: the outer zone, followed by the intermediate zone, and finally the most interior or inner zone (see Fig. 2). Two additional zones were developed by combining 1) the outer and intermediate zones (outer plus intermediate) and 2) the intermediate and inner zones (intermediate plus inner), for a total of five zone permutations—three that reduced the area sampled per wetland by 66% and two combined zones that reduced sampling area by 33%.
Lane’s (2003) original metric values were derived from total macrophyte presence found in all four transects within each wetland and called “site” metrics. To account for the non-independent nature of the tests and binomial metric distributions, PROC GLIMMIX, a procedure that fits statistical models to non-normally distributed data, was used to compare each of the zone-based metrics (e.g., outer, intermediate, outer plus intermediate, etc.) to the corresponding site metric in SAS (version 9.2, SAS Inc., Cary, North Carolina), using the CONTRAST statement. Linear regressions were calculated to predict and estimate variance of site metric distributions based on zone-specific metric distributions. Differences in mean richness measures between the zones were compared using the non-parametric Friedman’s multiple comparison test (Zar 1999) in Systat (version 13, Systat Software, Inc., Chicago IL).
Results
Site Characterization and Species Richness
The average transect length of the 74 emergent marshes was 45 m, with transects ranging in size from 5 m to 100 m. Outer zone length averaged 66 m (±19 m SD), while the intermediate zone averaged 60 m (±19 m SD) and the inner zone averaged 52 m (±19 m SD). The combined outer plus intermediate zone averaged 126 m (±38 m SD) in length and the intermediate plus inner zone averaged 112 m (±37 m SD). The highest richness (57 taxa) was found at a large, recovering site in a fallow pasture; the lowest richness (14 taxa) was found in a small undisturbed oligotrophic site. Three hundred ninety (390) plants were identified overall, and on average, sites had 31 taxa (±10 SD). The most commonly found taxa were Panicum hemitomon Schult. (maidencane; 63 sites), Andropogon virginicus L. (broomsedge bluestem; 48 sites), Pontederia cordata L. (pickerelweed; 45 sites), Centella asiatica (L.) Urb. (spadeleaf; 42 sites), and Eupatorium capillifolium (Lam.) Small (dogfennel; 34 sites). One hundred forty (140) species were found only once and 45 species were found twice. The outer zone averaged 27 taxa (±9 SD) and had the highest maximum richness (50 taxa) and highest minimum richness (12 taxa); the intermediate zone averaged 16 taxa (±8 SD) and ranged from 3 to 37 taxa; the inner zone averaged 11 taxa (±7 SD) and ranged from 2 to 30 taxa. The outer plus intermediate zone averaged 22 taxa (±10 SD), while the intermediate plus inner zone averaged 14 taxa (±8 SD). As a proportion of the total richness at all sites, 87% of the species found in wetland sites occurred in the outer third, 52% were found in the intermediate third, and just 36% were in the inner third. Friedman’s test identified significant differences (p < 0.0001) for richness measures between all zones compared (Fig. 3).

Highly significant (p < 0.0001) differences were found in Friedman’s test on richness between a) two zones and b) three zones
Zonal Distribution Analyses—Plant Distribution and Metric Response
Average site data metrics are given in supplementary Table S1. No significant differences were found between site metrics derived from the entire wetland and those derived using portions of the data (i.e., from each zone; Table 1). In all cases, the strength of the regression models between site and zone data decreased from the species-rich outer zones towards the inner zones (Figs. 4, 5, 6, 7 and 8). The proportion of site variance estimated by sampling a single zone (33% of the wetland sample area) ranged from high (R 2 = 0.985 for outer zone CC scores) to moderate (R 2 = 0.403 for inner zone ratio of annual to perennial species). Linear correlations between metrics from the outer zone and site data were highly significant (R 2 > 0.943), and sampling data from the outer plus intermediate zone (66% of the wetland sample area) resulted in regressions on site data with R 2 > 0.977. Metrics derived from sampling the inner and intermediate zones only explained moderate amounts of site metric scores, with regression estimates ranging from R 2 = 0.643 (annual to perennial ratio) to R 2 = 0.891 (percent sensitive).

Linear correlations for % exotics values between site and a) outer zone, b) outer plus intermediate zone, c) intermediate zone, d) intermediate plus inner zone, and e) inner zone

Linear correlations for % sensitive values between site and a) outer zone, b) outer plus intermediate zone, c) intermediate zone, d) intermediate plus inner zone, and e) inner zone

Linear correlations for % tolerant values between site and a) outer zone, b) outer plus intermediate zone, c) intermediate zone, d) intermediate plus inner zone, and e) inner zone

Linear correlations for mean coefficient of conservatism (CC) values between site and a) outer zone, b) outer plus intermediate zone, c) intermediate zone, d) intermediate plus inner zone, and e) inner zone

Linear correlations for annual to perennial (A:P) ratio values between site and a) outer zone, b) outer plus intermediate zone, c) intermediate zone, d) intermediate plus inner zone, and e) inner zone
Discussion
Deterministic Factors Affecting Macrophyte Distribution
van der Valk and Welling (1988) identified four major mechanisms leading to differential distributions of species along an elevation water-depth gradient that are applicable to the wetlands in this study: 1) seed distribution, 2) recruitment rates, 3) survival rates after flood recession, and 4) survival after re-flooding. Where these hydrologic-based fluctuations are greatest—at the ecotone between wetland and terrestrial habitat—the highest richness is expected (and was found in this study) as both terrestrial and wetland plants, including invasive species (Miller and Zedler 2003) vie for a footing and expand or contract their distribution based on hydrologic characteristics (Fraser and Karnezis 2005).
Other aspects, beyond simple hydroperiod and hydropattern, also affect plant success in wetlands (Kirkman et al. 2000). For instance, hydrologically-mediated processes at the wetland-upland boundary can affect plant success and richness, such as changes in salinity, nutrient cycling and the leaching of nutrients, oxidation and organic matter, and pore-water oxygen levels (Cronk and Fennessy 2001). Small depressional wetlands have a high perimeter to area ratio, so small changes in stage height can impact a large wetland area (Haag et al. 2005); they are also highly reliant on rainfall for ephemeral ponding and soil saturation to withstand invasion from upland sources (Dierberg and Brezonik 1984; Cronk and Fennessy 2001). Both too much and too little water can have drastic effects on wetland vegetation. Thus, the vegetated upland-wetland boundary in ephemeral, hydrologically-dynamic wetlands can be expected to be somewhat indistinct and the locus of constant community structure change in response to wide variations in edaphic and phreatic conditions based on relative elevation and hydrologic conditions.
Zonal Effects on Wetland Characterization
In sampling circular or ellipsoid wetlands using belted transects and equal area quadrants, the peripheral quadrants constitute a small sample of a large area and conversely, central quadrants constitute a large sample of a small area. As a result of the constant quadrant size used in this study, the sampling design employed thus sampled the peripheral or outer zones at a lower intensity than the center or inner zones. However, the sampling method did include four transects in an attempt to capture the richness of the ecotone between wetlands and uplands (i.e., a minimum of 20 m2 was sampled on the periphery of each site). Furthermore, the four transects sampled areas of localized topography. For instance, peripheral wetland edges often included pools with hydrophytic vegetation where hogs had created wallows, allowing for more hydrophilic plants to colonize (e.g., Brasenia schreberi J.F. Gmel., watershield). Similarly, the peripheral edges also included tussocks and soil mounds wherein slight differences in elevation permitted vastly different floral diversity (Fraser and Karnezis 2005). Indeed, the disproportionately greater species richness in the outer zone may explain a portion of the correlation strength between the outer zone metrics and those derived from the full wetland site, which ranged from R 2 = 0.943 to R 2 = 0.983 (p < 0.0001).
Based on our findings using belt transects, we recommend sampling only the outer third of each wetland to obtain an accurate assessment of wetland condition; all correlations with site data are R 2 > 0.943 and highly significant for this outer zone. While surveying an additional third (i.e., 66% of the wetland) improves the correlation (i.e., all R 2 > 0.977), the savings in terms of effort otherwise needed to assess the site does not appear warranted for a slight improvement in correlation strength. Either method (i.e., sampling the outer third or the outer plus intermediate thirds) are recommended over sampling the entire wetland for condition assessments.
Had a different sampling scheme been employed, results would likely have differed. For instance, had a releve plot been established, vegetation zones and estimates of percent cover would have been captured. Analyses of sampling effort, based on the time and effort involved, could have also been conducted. Houlahan and Findlay (2004) conducted a study of macrophyte sampling effort in 58 wetlands based on time spent, using a modified Braun-Blanquet abundance estimator, and divided total sampling into six 30-min sampling efforts. They found that in most surveys, more than 60% of species were found in the first of these segments and that more than 95% had been found by the fifth segment. With estimates of plot abundance, different metrics would likely have been developed, too. For instance, percent exotic species would have been calculated for the releve rather than the wetland. However, the exploratory nature of the initial metrics focused on simplicity and repeatability in multiple wetland types. Subsequent efforts in isolated forested wetlands (Reiss 2006) and forested sloughs (Reiss et al. 2010) have successfully developed metrics for the sites using a similar presence measure along quadrants.
Conclusion
As a result of these analyses across isolated emergent wetlands of varying sizes, which revealed no significant differences in macrophyte metrics between any of the wetland zones (e.g., outer, intermediate, inner, outer plus intermediate, intermediate plus inner) and the complete site assessment, we concluded that resources for macrophyte-based isolated wetland assessment using belt transects should focus solely on the peripheral or outer third of each isolated emergent marsh wetland. Such a sampling approach would drastically reduce the time and sampling area required for wetland community characterization and assessment by removing the need for full transects through wetland vegetation and typically deeper water habitats. Indeed, sampling just the outer third of each isolated wetland—a 66% decrease in area sampled (with a smaller decrease in effort, considering the majority of the wetland plants are found in the outer zone)—will produce results of equal accuracy to the full transect approach.
References
Brock MA, Casanova MT (1997) Plant life at the edges of wetlands; ecological responses to wetting and drying patterns. In: Klomp N, Lunt I (eds) Frontiers in ecology: Building the links. Elsevier Science, Oxford, pp 181–192
Brown MT, Vivas MB (2005) Landscape development intensity index. Environmental Monitoring and Assessment 101:289–309. doi:10.1007/s10661-005-0296-6
Cohen MJ, Carstenn S, Lane CR (2004) Floristic quality indices for biotic assessment of depressional marsh condition in Florida. Ecological Applications 14:784–794. doi:10.1890/02-5378
Cronk JK, Fennessy MS (2001) Wetland plants: Biology and ecology. CRC Press, L.L.C., Boca Raton
De Groot RS, Stuip MAM, Finlayson CM, Davidson N (2006) Valuing wetlands: Guidance for valuing the benefits derived from wetland ecosystem services, Ramsar Technical Report No. 3/CBD Technical Series No. 27. Ramsar Convention Secretariat, Gland, Switzerland and Secretariat of the Convention on Biological Diversity, Montreal, Canada. ISBN 2-940073-31-7
DeKeyser ES, Kirby DR, Ell MJ (2003) An index of plant community integrity: development of the methodology for assessing prairie wetland plant communities. Ecological Indicators 3:119–133. doi:10.1016/S1470-160X(03)00015-3
Dierberg FE, Brezonik PL (1984) Water chemistry of a Florida cypress dome. In: Ewel KC, Odum HT (eds) Cypress swamps. University Presses of Florida, Gainesville, pp 34–50
Doherty SJ, Cohen MJ, Lane CR, Line L, Surdick JA (2000) Biological criteria for inland freshwater wetlands in Florida: a review of technical and scientific literature (19901999). Center for Wetlands, University of Florida, Gainesville
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67:345–366. doi:10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Ewel KC (1990) Swamps. In: Myers RL, Ewel JJ (eds) Ecosystems of Florida. University of Central Florida Press, Orlando, pp 281–323
Ewel K, Odum HT (1984) Cypress swamps. University of Florida Press, Gainesville
Ewing K (1996) Tolerance of four wetland plant species to flooding and sediment deposition. Environmental and Experimental Biology 36:131–146. doi:10.1016/0098-8472(96)01000-3
Florida Exotic Pest Plant Council (FLEPPC) (2001) 2001 List of Florida’s invasive species. Florida EPPC Newsletter 11:3–4
Fraser LH, Karnezis JP (2005) A comparative assessment of seedling survival and biomass accumulation for fourteen wetland plant species grown under minor water-depth differences. Wetlands 25:520–530. doi:10.1672/0277-5212(2005)025[0520:ACAOSS]2.0.CO;2
Fretwell JD, Williams JS, Redman PJ (compilers) (1996) National Water Summary on Wetland Resources. U.S. Geological Survey Water Supply Paper 2425. United States Government Printing Office, Washington, DC
Galatowitsch SM, Anderson NO, Ascher PD (1999) Invasiveness in wetland plants in temperate North America. Wetlands 19:733–755. doi:10.1007/BF03161781
Gernes MC, Helgen JC (1999) Indexes of biotic integrity (IBI) for wetlands: vegetation and invertebrate IBIs. Final Report to EPA, Assistance Number CD995525-01. Minnesota Pollution Control Agency, St. Paul
Haag KH, Lee TM, Herndon DC (2005) Bathymetry and vegetation in isolated marsh and cypress wetlands in the northern Tampa Bay Area, 2000–2004. U.S. Geological Survey Scientific Investigations Report 2005–5109. U.S. Geological Survey, Reston
Hayashi M, van der Kamp G (2000) Simple equations to represent the volume-area-depth relations of shallow wetlands in small topographic depressions. Journal of Hydrology 237:74–85. doi:10.1016/S0022-1694(00)00300-0
Houlahan JE, Findlay CS (2004) Effect of invasive plant species on temperate wetland plant diversity. Conservation Biology 18:1132–1138. doi:10.1111/j.1523-1739.2004.00391.x
Kercher SM, Zedler JB (2004) Flood tolerance in wetland angiosperms: a comparison of invasive and noninvasive species. Aquatic Botany 80:89–102. doi:10.1016/j.aquabot.2004.08.003
Kirkman LK, Goebel PC, West L, Drew MB, Palik BJ (2000) Depressional wetland vegetation types: a question of plant community development. Wetlands 20:373–385. doi:10.1672/0277-5212(2000)020[0373:DWVTAQ]2.0.CO;2
Kushlan JA (1990) Freshwater marshes. In: Myers RL, Ewel JJ (eds) Ecosystems of Florida. University of Central Florida Press, Orlando, pp 323–363
LaClaire LV (1995) Vegetation of selected upland temporary ponds in north and north-central Florida. Bulletin Florida Museum of Natural History 38:69–96
Lane CR (2003) Development of Biological Indicators of Freshwater Wetland Condition in Florida. Ph.D. Dissertation. Department of Environmental Engineering, University of Florida, Gainesville
Lane CR, Brown MT (2007) Diatoms as indicators of isolated herbaceous wetland condition in Florida, U.S.A. Ecological Indicators 7:521–540. doi:10.1016/j.ecolind.2006.06.001
Lane CR, D’Amico E (2010) Calculating the ecosystem service of water storage in isolated wetlands using LiDAR in north central Florida, USA. Wetlands 30:967–977. doi:10.1007/s13157-010-0085-z
Lane CR, Brown MT, Murray-Hudson M, Vivas MB (2003) The wetland condition index (WCI): Biological indicators of wetland condition for isolated depressional herbaceous wetlands in Florida. H.T. Odum Center for Wetlands, Gainesville
Leibowitz SG (2003) Isolated wetlands and their functions: an ecological perspective. Wetlands 23:517–531. doi:10.1672/0277-5212(2003)023[0517:IWATFA]2.0.CO;2
Lockhart C, Austin DF, Aumen NG (1999) Water level effects on growth of Melaleuca seedlings from Lake Okeechobee (Florida, USA) littoral zone. Environmental Management 23:507–518. doi:10.1007/s002679900205
Ma Z, Cai Y, Bo L, Chen J (2011) Managing wetland habitats for waterbirds: an international perspective. Wetlands 30(1):15–27. doi:10.1007/s13157-009-0001-6
Mack JJ (2004) Integrated Wetland Assessment Program. Part 4: a vegetation index of biotic integrity (VIBI) and tiered aquatic life uses (TALUs) for Ohio wetlands. Ohio EPA Technical Report WET/2004-4. Ohio Environmental Protection Agency, Wetland Ecology Group, Division of Surface Water, Columbus, OH
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
McKevlin MR, Hook DD, Rozelle AA (1998) Adaptations of plants to flooding and soil waterlogging. In: Messina MG, Conner WH (eds) Southern forested wetlands ecology and management. Lewis Publishers, Boca Raton, pp 173–204
Megonigal JP, Day FP (1992) Effects of flooding on root and shoot production of bald cypress in large experimental enclosures. Ecology 73:1182–1193. doi:10.2307/1940668
Miller RE, Gunsalus BE (1997) Wetland rapid assessment procedure (WRAP). Technical Publication REG-001. South Florida Water Management District, West Palm Beach
Miller RC, Zedler JB (2003) Responses of native and invasive wetland plants to hydroperiod and water depth. Plant Ecology 167:57–69. doi:10.1023/A:1023918619073
Penfound WT (1952) Southern swamps and marshes. Botanical Review 18:413–446. stable URL: http://www.jstor.org/stable/4353488
Reed PB (1997) Revision of the national list of plant species that occur in wetlands. U.S. Department of the Interior, Washington, DC
Reiss KC (2006) Florida wetland condition index for depressional forested wetlands. Ecological Indicators 62:337–352. doi:10.1016/j.ecolind.2005.03.013
Reiss KC, Hernandez E, Brown MT (2009) Evaluation of permit success in wetland mitigation banking: a Florida case study. Wetlands 29(3):907–918. doi:10.1672/08-148.1
Reiss KC, Brown MT, Lane CR (2010) Characteristic community structure of Florida’s subtropical wetlands: the Florida wetland condition index for depressional marshes, depressional forested, and flowing water forested wetlands. Wetlands Ecology and Management 18:543–556. doi:10.1007/s11273-009-9132-z
Scozzafava M, Kentula ME, Riley E, Magee TK, Serenbetz G, Sumner R, Faulkner C, Price M (2011) The national wetland condition assessment: national data on wetland quality to inform and improve wetlands protection. National Wetlands Newsletter 33:11–13
Sharitz RR, Gresham CA (1998) Pocosins and carolina bays. In: Messina MG, Conner WH (eds) Southern forested wetlands. Lewis Publishers, Boca Raton, pp 343–378
Sun G, Riekerk H, Korhnak LV (1995) Shallow groundwater table dynamics of cypress wetland/pine upland systems in Florida flatwoods. Soil and Crop Science Society of Florida Proceedings 54:66–71
Tiner RW (2003) Geographically isolated wetlands of the United States. Wetlands 23(3):494–516. doi:10.1672/0277-5212(2003)023[0494:GIWOTU]2.0.CO;2
U.S. Environmental Protection Agency (US EPA) (2002) Methods for evaluating wetland condition: using vegetation to assess environmental condition. EPA-822-R-02-020. Office of Water, U.S. Environmental Protection Agency, Washington, DC
U.S. Environmental Protection Agency (US EPA) (2009) National water quality inventory: report to Congress, 2004 reporting cycle. EPA 841-R-08-001. Office of Water, U.S. Environmental Protection Agency, Washington, DC
van der Valk AG (1981) Succession in wetlands: a Gleasonian approach. Ecology 62:688–696. stable URL:http://www.jstor.org/stable/1937737
van der Valk A, Welling CH (1988) The development of zonation in freshwater wetlands: an experimental approach. In: During HJ, Werger MJA, Willems HJ (eds) Diversity and pattern in plant communities. SPB Academic Publishing, The Hague, pp 145–158
Wilhelm G, Ladd D (1988) Natural area assessment in the Chicago region. Transactions of the 53rd North American Wildlife and Natural Resource Conference. pp 361–375
Winchester BH, Bays JS, Higman JC, Knight RC (1985) Physiography and vegetation zonation of shallow emergent marshes in southwestern Florida. Wetlands 5:99–118. doi:10.1007/BF03160790
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, Inc, Englewood Cliffs
Acknowledgements
The United States Environmental Protection Agency through its Office of Research and Development partially funded and collaborated in the research described here under contract number EP-D-06-096 to Dynamac Corporation. Although reviewed by the US EPA and approved for publication, this manuscript does not necessarily reflect official US EPA policy. Justicia Rhodus, Dynamac Corporation, provided technical review and editing services, although any inadequacies in this manuscript remain our own.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table S1
Site metric values used in contrasts between site data and zone data (DOC 127 kb)
Rights and permissions
About this article
Cite this article
Murray-Hudson, M., Lane, C.R., North, S. et al. Macrophyte Species Distribution, Indices of Biotic Integrity, and Sampling Intensity in Isolated Florida Marshes. Wetlands 32, 449–460 (2012). https://doi.org/10.1007/s13157-012-0278-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-012-0278-8