
Abstract
The loss and degradation of wetlands worldwide has adversely affected waterbirds, which depend on wetland habitats. Many studies have indicated that effectively managed wetlands can provide alternative or complementary habitats for waterbirds and mitigate the adverse effects of wetland loss and degradation. We review the studies on the habitat variables affecting use of wetlands by waterbirds, and we suggest how wetlands can be managed to provide waterbird habitat. These habitat variables include water depth, water level fluctuation, vegetation, salinity, topography, food type, food accessibility, wetland size, and wetland connectivity. Overall, the practice of wetland management requires integrated knowledge related to the entire wetland ecosystem, and ecosystem-based approach is needed to improve the habitat quality of managed wetlands with considering multiple spatial scales, temporal variability, and trade-off among diverse habitat requirements of different waterbirds. Several priorities for future research and management are also suggested in this paper.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Over 50% of wetlands in the world have been lost in the past century, and the remaining wetlands have been degraded to different degrees because of the adverse influences of human activities (Fraser and Keddy 2005). The loss and degradation of wetlands has negatively affected waterbirds, which depend on wetland habitats. How to provide high quality habitats for waterbirds through effective management is a critical issue in waterbird conservation (Weber and Haig 1996; Erwin 2002; Taft et al. 2002).
Currently, the management of wetlands focuses on artificial and restored wetlands, which by definition are greatly affected by human activities. Artificial wetlands are those that are created or extensively modified by humans, including paddy fields, salt ponds, aquacultural ponds, impoundments, and reservoirs. Although artificial wetlands cannot completely replace the functions of natural ones as waterbird habitats (e.g., Ma et al. 2004; Desrochers et al. 2008), researchers and managers widely recognize that artificial wetlands can provide alternative or complementary habitats for waterbirds in all life stages (e.g., Weber and Haig 1996; Elphick and Oring 1998; Connor and Gabor 2006) and consequently, can partially mitigate the adverse influences of loss and degradation of natural wetlands. For example, paddy fields are among the most important artificial habitats for waterbirds worldwide (Czech and Parsons 2002). Rice prairie in Texas provides wintering habitat for over two million waterfowl (Hobaugh et al. 1989), and herons and egrets largely depend on paddy fields as their major foraging habitats in Europe and Asia, especially where the natural wetlands have been lost or degraded (Kushlan and Hafner 2000). With the continuous loss of natural wetlands globally, we can expect artificial wetlands to become increasingly important as habitats for waterbirds (Czech and Parsons 2002).
In contrast to artificial wetlands, restored wetlands are areas that historically supported a natural wetland ecosystem but were modified and used for other purposes before being altered so as to regain the characteristics of natural wetlands (Hammer 1997). Because wetlands represent important natural capital and provide essential ecological services, restoration of wetlands is a worldwide activity, especially in America (e.g., Zedler 2001; Erwin and Beck 2007), Europe (e.g., Wolters et al. 2005), and Asia (e.g., Nakamura et al. 2006). Some countries and regions have developed policies or laws to ensure the implementation of wetland restoration (e.g., the ‘no net loss’ policy of USA enacted in 1988 and the Habitat Directive of European Union adopted in 1992). By providing resting, roosting, foraging, and nesting habitats, restored wetlands benefit diverse waterbird species (Erwin and Beck 2007; O’Neal et al. 2008).
Although artificial and restored wetlands mitigate loss of wetlands in terms of area, maintenance of wetland area is completely different from maintenance of wetland function or quality. Many studies have indicated that effective management plays a critical role in enhancing habitat quality of wetlands for waterbirds (e.g., Erwin 2002; Balcombe et al. 2005). Over the past half century, especially the past two decades, the variables affecting habitat use by waterbirds have been intensively studied in both artificial and natural wetlands. In this paper, we review recent studies on the habitat variables affecting habitat use by waterbirds, and we suggest how these variables could or should be managed to benefit waterbirds.
Habitat Variables
Water Depth
Many studies have indicated that water depth is an important variable affecting the use of wetland habitats by waterbirds (Velasquez 1992; Elphick and Oring 1998; Colwell and Taft 2000; Isola et al. 2002), and this relationship has served as the basis for the guidelines of wetland management (Bolduc and Afton 2004). Water depth directly determines the accessibility of foraging habitats for waterbirds because of the restrictions of bird morphology, such as the lengths of tarsometatarsi (for wading birds, Powell 1987; Baker 1979; Ntiamoa-Baidu et al. 1998; Collazo et al. 2002; Darnell and Smith 2004) or necks (for dabbling ducks, Poysa 1983). Larger species with longer necks, bills, and legs can feed in deeper habitats than smaller taxa.
Non-diving waterbirds, such as wading and dabbling birds, generally require shallow water to forage, and their access to foraging habitat is limited by water depth. In contrast, diving waterbirds require deep water, and their access to foraging habitat is limited by the minimum water depth that allows them to dive (Fig. 1). Because the wading and dabbling birds are the dominant waterbird groups in most regions worldwide, the greatest waterbird diversity and density generally occur at a relatively shallow water depth, where the depth requirements of different waterbird groups overlap (e.g., 10–20 cm, Elphick and Oring 1998, 2003; Colwell and Taft 2000; Isola et al. 2002; Taft et al. 2002). Habitats with deeper water, however, support the greatest density of waterbirds in areas where diving birds are dominant (Stapanian 2003), and where the wetlands provide roosting sites for waterfowl (Hattori and Mae 2001). From a management perspective, the overlapping of water depth requirements among waterbird groups suggests that wetlands can be managed to meet the water depth needs for different waterbird groups. At the same time, it may be necessary to manage wetlands exclusively for species associated with the extreme ends of the depth spectrum (e.g., diving waterbirds and small shorebirds), especially where such species constitute a large component of the waterbird community (Taft et al. 2002).

Variation of water depths at foraging sites among waterbird groups. Small shorebirds (such as sandpipers) forage in water less than 5 cm deep; large shorebirds (such as godwits) forage in water up to 15 cm deep; dabbling ducks (such as teals and mallards) and large waders (such as herons, egrets and ibis) forage in water up to 30 cm deep. Diving waterbirds (such as cormorants and grebes) require a minimum water depth of >25 cm and can forage in water up to several meters deep (Refer to data in Pöysä 1983; Baldassarre and Fischer 1984; Fredrickson and Reid 1986; Accurso 1992; Davis and Smith 1998; Elphick and Oring 1998; Ntiamoa-Baidu et al. 1998; Colwell and Taft 2000; Isola et al. 2002; Bolduc and Afton 2004)
In addition to limiting access to foraging habitats, water depth affects the net energy intake of waterbirds because foraging efficiency decreases with increasing water depth. Gawlik (2002) indicated that for wading birds that forage on prey in the water column, the locomotion of the birds might be slowed in deep water because of increased water resistance with depth. Moreover, deeper water can also reduce foraging efficiency because prey can escape not only horizontally, as is the case in shallow water, but also vertically. In addition, prey in deep water may be more difficult to detect, especially if the water is turbid. Consequently, drawdown can increase the foraging efficiency of wading birds by concentrating fish and other prey in limited, low-lying areas (Kushlan 1978; Bancroft et al. 2002; Sustainable Ecosystems Institute 2007). Foraging efficiency for the non-diving herbivores may also be reduced in deeper water. When foraging in shallow water, these birds feed by dipping their heads and necks only (neck-dipping), but when feeding in deeper water, they must tilt their entire body forward (up-ending). The latter results in a lower food intake rate and higher energy expenditure (Guillemain and Fritz 2002; Nolet et al. 2002). This suggests that waterbirds obtain a higher net energy intake in shallow than deep water, even with the same food abundance. Holm and Clausen (2006) also indicated that non-diving herbivores prefer foraging submerged vegetation in shallow water until depletion, even when food is more abundant in deeper water.
Because larger species of waterbirds generally have longer necks, bills, and legs, they usually can access greater range of water depths than smaller species (Baker 1979; Isola et al. 2002). The range of accessible water depths also depend on foraging behavior. Compared to shorebirds, waterfowl can use diverse foraging behaviors (such as surface dabbling, tipping-up, and head-submerging), each of which is suitable for foraging at a different water depth (Isola et al. 2002). Therefore, waterfowl and large wading birds can use a wider range of water depths than small shorebirds.
Water Level Fluctuation
Water level fluctuation in wetlands can be caused by seasonal flooding, tides, and agricultural irrigation or drawdown. The effect of water level fluctuation on waterbirds varies among groups and seasons. Generally, water level fluctuation creates habitats with diverse water depths changing in time and space. This provides more foraging opportunities and consequently supports a high species richness and abundance of waterbirds (Dimalexis and Pyrovetsi 1997; Ntiamoa-Baidu et al. 1998). Water level fluctuation, however, may create “ecological traps” and be detrimental for the breeding, brood-rearing, and molting of waterbirds (Kaminski et al. 2006). For example, breeding attempts are abandoned when water rises submerge nests and water drops make birds more vulnerable to mammalian predators after nesting. Many studies have shown that the brood densities of waterbirds are greater on wetlands with stable water levels than on seasonally flooded wetlands because stable water level benefits the breeding of waterbirds by providing suitable nest sites (e.g., Ogden 1991; Connor and Gabor 2006). Impoundments with stable water level also can attract more dabbling ducks than intertidal flats with periodic tidewater (Gordon et al. 1998), although shorebirds may prefer the latter.
Vegetation
Beyond providing food such as seeds, leaves, tubers, and rhizomes for herbivorous waterbirds, vegetation is an important habitat element and greatly influences the habitat use of waterbirds. The effect and importance of vegetation depends on the season and on the waterbird group. In the breeding season, emergent and floating plants benefit the nest building of coots (Fulica atra), mallards (Anas platyrhynchos), and moorhens (Gallinula chloropus) and consequently enhance their breeding success (Froneman et al. 2001; Sánchez-Zapata et al. 2005). Emergent plants also provide shelter and decrease human disturbance, which often occurs in artificial wetlands, at both roosting and nesting sites (Hattori and Mae 2001). In addition, tree islands benefit colonial waterbirds by providing colony sites in open wetlands (Hoffman et al. 1994). Dense vegetation also provides habitat and food requirements for invertebrates, and improves the viability of eggs or diapausing invertebrates, which increases invertebrate density, biomass, and diversity (Wiggins et al. 1980; Rehfisch 1994). This increases food for waterbirds (Anderson and Smith 2000). Many studies have indicated that species richness and abundance of waterbirds increase with increasing emergent vegetation cover in wetlands, especially during breeding periods when waterbirds are less mobile and more sensitive to disturbance (Owen and Black 1990; Losito and Baldassarre 1995; VanRees-Siewert and Dinsmore 1996; Post 1998; Froneman et al. 2001).
High, dense vegetation, however, can limit the accessibility of wetlands and adversely affect foraging (Fujioka et al. 2001; Bancroft et al. 2002) and prey detection (White and Main 2004) by waterbirds, thus too much emergent vegetation can lead to decreased numbers of nesting waterbirds. Most waterbird groups, except the rails and bitterns, prefer unvegetated or short and sparsely vegetated foraging habitats (e.g., VanRees-Siewert and Dinsmore 1996; Maeda 2001; Darnell and Smith 2004; Sánchez-Zapata et al. 2005). Open habitats have been considered an important variable influencing shorebird abundance (Colwell 1993; Warnock and Takekawa 1995). Dimalexis and Pyrovetsi (1997) also indicated that fish-eating waterbirds, such as herons and egrets, avoid foraging in water bodies with submerged vegetation because of the reduced efficiency in searching for prey.
Salinity
Salinity is an important factor in the management of salt ponds for waterbird habitats. Generally, water of high salinity is harmful to waterbirds. Waterbirds that drink highly saline lose body weight by dehydration (e.g., Purdue and Haines 1977; Hannam et al. 2003), and waterbirds avoid water of high salinity even for roosting because salts reduce the waterproofing of feathers and thus increases the energy costs of thermoregulation (Rubega and Robinson 1997). Euliss et al. (1989) reported that high salinity water may cause carbonate to precipitate on tail feathers of Ruddy Ducks (Oxyura jamaicensis), which erodes the feathers and interferes with diving and flying.
Water salinity also affects the species composition of aquatic plant communities and consequently, indirectly affects herbivorous waterbirds. In the two coastal lagoons of Denmark, Chara and Potamogeton plants, which are high quality foods for herbivorous waterbirds (e.g., mute swans Cygnus olor), are vulnerable to high salinity and were more likely to be found in the lagoon with fresh or slightly brackish water ( salinity 2–7 ppt, mean 5 ppt), while Ruppia plants, which are relatively low quality foods for herbivorous waterbirds, are tolerant of high salinity and were abundant in the lagoon with high salinity (salinity 7–34 ppt, mean 17 ppt) (Holm 2002; Holm and Clausen 2006). The distribution of plants in areas with different salinities causes herbivorous waterbirds to aggregate at lower salinity ponds with higher quality food. However, the opposite condition was reported in Lake Wyara (salt water) and Lake Numalla (freshwater) in southwest Queensland, Australia, where there was more macrophyte vegetation providing food for herbivorous waterbirds in the saline lake than in the freshwater lake. This caused much more herbivorous waterbirds were recorded in the saline lake (Kingsford and Porter 1994).
Water salinity also determines the distribution of zoobenthos and aquatic animals and thus influences the use of foraging sites by waterbirds. The effects of water salinity on zoobenthos and aquatic animals are taxa dependent. For example, Velasquez (1992) and Takekawa et al. (2006) found that chironomid fly larvae, amphipods, and copepods predominate in relatively low-salinity (<50 ppt) water but are replaced by brine-adapted organisms such as Artemia and Ephydra in high-salinity water (>150 ppt). In the salt ponds in San Fransisco Bay estuary, zoobenthos are abundant in relatively low (<100 ppt) and high (>200 ppt) salinity water but are scarce at mid-salinities (100–200 ppt). However, the opposite relationship exists between water salinity and zooplankton abundance, i.e., zooplankton are most abundant in mid-salinity water (Takekawa et al. 2006). The different responses of various invertebrates to water salinity means that the preferred water salinity at foraging sites of waterbirds varies with waterbird group and prey composition. Velasquez (1992) found that the highest foraging densities of waterbirds occurred with salinities of 25–70 and 170–220 ppt at the Berg River estuary, South Africa. Warnock et al. (2002) reported that the highest numbers and species richness of waterbirds occur with salinities around 140 and 126 ppt, respectively, at San Francisco Bay, USA. Takekawa et al. (2006) also indicated that most waterbirds forage in mid-salinity (81–150 ppt) at San Francisco Bay, USA. All these suggest that, in saline environments, regulating water salinity according to the prey of waterbirds is critical in supporting foraging habitats for diverse waterbirds.
Topography
Because the topography (relief) of restored wetlands cannot be easily altered after construction, topography needs to be carefully considered and planned before wetland restoration begins. With a given water depth in a wetland, topographic variation broadens the range of water depths and thus provides a range of foraging habitats, from exposed mud flats to deep water, to meet the needs of shorebirds, wading birds, dabbling ducks, and diving waterbirds (Isola et al. 2002; Takekawa et al. 2006). Moreover, emergent plants can be distributed according to suitable water depth, enhancing the structural diversity of wetland habitats for waterbirds (Hoffman et al. 1994). In addition, island uplands and levees (including dredge-material islands) in wetlands can be used as resting, roosting, preening, and even nesting and molting sites by waterbirds (Parnell et al. 1986; Warnock and Takekawa 1995; Pyrovetsi 1997; Holm 2002; Warnock et al. 2002; Erwin and Beck 2007). Many studies have suggested that the combination of variable topography with suitable water depth provides habitats accessible for diverse waterbirds (Colwell and Taft 2000; Isola et al. 2002; Taft et al. 2002). Ponds with gentle sloping sides can also increase topographical variation and attract both short- and long-legged wading birds (Erwin et al. 1994).
Food and Its Accessibility to Waterbirds
Waterbirds use diverse foods, including seeds (dabbling ducks, geese, cranes), leaves (geese), tubers and rhizomes (geese, swans), invertebrates (shorebirds, waterfowl), and some vertebrates, such as fish and amphibians (wading birds). The amounts, composition, and spatiotemporal dynamics of these foods largely affect the use of foraging habitats by waterbirds and can be important indicators of habitat quality (Davis and Smith 1998; Taft and Haig 2005; Hartke et al. 2009). Many studies have clearly demonstrated that managed wetlands can be important foraging habitats for waterbirds, especially during pre-migration fattening and wintering periods (e.g., Davidson and Evans 1986; Masero et al. 2000). Understanding food resources in wetlands is important for determining the potential carrying capacity of the wetlands for waterbirds.
Food preferences of waterbirds can vary greatly among species, even among those within the same group. For example, on saltmarsh in East China, little egret (Egretta garzetta) prefer to forage on fish and other aquatic animals, while cattle egret (Bubulcus ibis) prefer to forage on insects and other arthropods (Zhu and Zou 2001). Moreover, waterbirds can change their diet composition with season and site (e.g., red knots Calidris canutus prey on hard-shelled mollusks in winter and soft-bodied arthropods in summer, Dekinga et al. 2001). Prey size also affects the prey selection of waterbirds (Zwarts and Blomert 1992; Davis and Smith 2001). This might be related to the pursuit of maximum net energy intake by birds through tradeoffs among foraging efficiency, handling costs, and energy intake for prey of different sizes. However, the relationship between waterbirds and their food (especially invertebrate prey) is still largely unexplored, and the effects of management on food for waterbirds may vary among sites (Sanders 2000). Further studies are required on the life cycle of invertebrate food for waterbirds and their use and selection by waterbirds to ensure that managed wetlands provide high quality foraging habitats for waterbirds.
Although food abundance is generally used in assessing habitat quality of waterbirds, food accessibility, which often differs from food abundance, greatly influences habitat use by waterbirds (Bolduc and Afton 2004). Food accessibility to waterbirds depends on both intrinsic and extrinsic factors. Intrinsic factors include bird morphology (neck length, leg length, body size, and feeding structures), foraging methods, and diet selectivity. Extrinsic factors include water depth, vegetation density, and prey vulnerability. Because these intrinsic and extrinsic factors differ among waterbird species and groups, particular species of waterbirds generally feed in particular wetlands with features that maximize the abundance and accessibility of their foods (Taft and Haig 2003).
Wetland Size
Many studies have indicated that wetland size influences species richness and abundance of waterbirds (Froneman et al. 2001; Paracuellos and Telleria 2004; Sánchez-Zapata et al. 2005). Generally, the spatial configurations and habitat heterogeneity of wetlands are related to size (Paracuellos 2006). Because waterbirds have different preferences with respect to habitat configuration, the larger wetlands, which are more likely than smaller wetlands to have high habitat heterogeneity, can support a greater diversity of waterbirds than the smaller ones (Colwell and Taft 2000; Froneman et al. 2001; Warnock et al. 2002; Paracuellos and Telleria 2004; White and Main 2004). Generally, waterbird species foraging close to the shore persist in both large and small ponds and are considered area-independent species, while species foraging in open and deep-water habitats are considered area-dependent species and are restricted to relatively large ponds (Paracuellos 2006). A recent review paper indicated that area sensitivity was detected in about half the wetland species studied (Bayard and Elphick, unpublished data). Area-dependent species tend to be the first to leave when pond size is reduced and open and deep-water habitats disappear. This causes a hierarchical disappearance of waterbird species as a wetlands becomes smaller (Paracuellos 2006). Paracuellos and Telleria (2004) reported that the distribution of waterbird species had a “nested” pattern among wetlands of different sizes: the scarcer waterbirds generally occur exclusively in large wetlands, and large wetlands support high species diversity of both area-independent and area-dependent species, while small wetlands generally support lower species diversity and only area-independent species. Thus larger wetlands are of greater conservation value than smaller ones in supporting diverse waterbird species.
However, the debate about whether several small habitat patches might be better than a single large patch (single large or several small (SLOSS), Soulé and Simberloff 1986) also applies to wetland management. Some studies have suggested that a group of small wetlands can maintain the same or even more waterbird species as one large wetland with equivalent area (Brown and Dinsmore 1986; Craig and Beal 1992; Scheffer et al. 2006). This might be due to high habitat heterogeneity among small wetlands. Some small wetlands, although used by waterbirds only seasonally, remain important in supporting the entire local and regional populations (Kushlan 1986; Craig and Beal 1992; Skagen and Knopf 1993). Moreover, compared to large wetlands, a group of small wetlands can more easily be managed to meet the different requirements of different waterbirds. Also, constructing a series of small wetlands is obviously more feasible in regions lacking space for one large wetland.
Although the total size of a wetland determines waterbird diversity, the size of accessible habitat is more crucial in determining the suitability of the wetland for a particular waterbird group (Gawlik 2002). Being restricted by their morphology or ecological habits, waterbirds may avoid or be unable to access some specific areas, such as areas of deep water, dense vegetation, and high salinity, as mentioned earlier. This greatly decreases the habitats available to waterbirds. For example, of the total 161 ha of impoundments at Pea Island National Wildlife Refuge (Coastal North Carolina, USA), the area of accessible habitats for small shorebirds is only 21 ha to 22 ha (Collazo et al. 2002). Consequently, accessible area, although it is dynamic and can be affected by precipitation and water level fluctuation, could be a better predictor than the total area of use by specific waterbird groups.
Wetland Connectivity
Generally, an individual wetland seldom meets all the requirements (foraging, resting, roosting, nesting sites, etc.) of diverse waterbirds. Thus, the development and maintenance of multiple, complementary wetlands within a mosaic can provide the resources required by diverse waterbirds. From a landscape perspective, the number and reproductive performance of waterbird species in a wetland can be affected by other wetlands nearby (Craig and Beal 1992; Dunning et al. 1992; Froneman et al. 2001; Kelly et al. 2008). Although some waterbirds show regional and local fidelity to a special habitat (Warnock and Takekawa 1996), they move around at landscape scales when deciding where to settle (Skagen and Knopf 1993; Warnock et al. 1995; Warnock and Takekawa 1996; Melvin et al. 1999; Plissner et al. 2000). For species that consume superabundant but ephemeral prey, foraging by traveling among wetlands is more effective than remaining at a single one, even it is large (Craig and Beal 1992). The connectivity between and within wetlands also facilities the exchange and movement of aquatic animals and plants among wetlands and among different wetland patches, and thus increases potential food for waterbirds (Bancroft et al. 1994).
Other Habitat Variables
In addition to the habitat variables mentioned previously, other variables related to the characteristics of sediments (e.g., organic matter content and particle size) and water quality (e.g., clarity, temperature, dissolved oxygen, and pH) can also directly or indirectly affect the use of wetlands by waterbirds. For example, the organic matter content in water and sediments affects the growth of aquatic plants, and determines invertebrate abundance (Rehfisch 1994). Particle size of sediments determines how water and oxygen penetrate sediments and thus affects the presence of meiofauna and infaunal and epifaunal invertebrates (Little 2000). Water transparency and temperature affect the growth of algae (Nielsen et al. 2002). The temperature of water also affects the hatching of invertebrate eggs and the development of invertebrate fauna (Rehfisch 1994).
Kersten et al. (1991) have demonstrated that the dissolved oxygen in the water affects the foraging of waterbirds by changing the vertical distribution of prey. At their study site in Camargue, France, the nocturnal respiration of macrophytes depleted the dissolved oxygen in the surrounding water, which forced the mosquito fish (Gambusia spp.) to concentrate at the surface in open water during early morning. Soon after sunrise, the photosynthesis of macrophytes increases the dissolved oxygen level in the surrounding water, enabling the mosquito fish to return to greater water depths. Accordingly, in the early morning, egrets tend to feed in groups in the open water (where mosquito fish are concentrated) but tend to feed solitarily over a wider area later in the day (when mosquito fish have dispersed). This suggests that dissolved oxygen in the water affects prey vulnerability and thus affects the habitat use of waterbirds.
Although these other variables can also influence the habitat use of waterbirds, they might be difficult or costly to regulate and control artificially, and generally are seldom considered in the practice of wetland management.
Implications for Management of Wetlands
The variation in habitat requirements among waterbird species and groups suggests that wetland management must be based on the region-specific knowledge about waterbird communities, including the species and their abundances and habitat requirements. The managers also need to understand the seasonal dynamics of waterbirds, so that management can be timed to meet the special needs of breeding, stopover, and wintering periods (Isola et al. 2002; Parsons 2002). However, because different waterbird species have various, and even contrasting, habitat requirements, the same management measures could have completely different effects on different species and groups (Craig and Beal 1992; Mitchell et al. 2006). This suggests that management solutions benefiting all species may not exist (Stralberg et al. 2009). Consequently, optimal wetland management to multi-species comes from assessing priorities and trade-offs among different species and groups of conservation concern (Elphick 2004; Thébault et al. 2008; Stralberg et al. 2009).
Many studies have indicated that hydrology is the most important variable determining the development and maintenance of wetland structure and functions (e.g., Winter and Woo 1990; Euliss et al. 2004), and that hydrology greatly affects the waterbird response to wetland dynamics (Bancroft et al. 1994; Ogden 1994; O’Neal et al. 2008; Hoover 2009). Unsuitable hydrological condition is usually the major reason for the failure of wetland restoration (Mitsch and Wilson 1996). Currently, construction of hydrological installations is an important practice in wetland management. For example, the practice of Structural Marsh Management (SMM) includes construction of water-control structures (e.g., floodgates, water pumps, canals, and levees) to regulate hydrological conditions according to the requirements of waterbirds; this includes drawdown or flooding to obtain a suitable water depth, opening and closing of flood gates to obtain desired fluctuations in water level, and excavating or banking to form various topographical features in wetlands (Collazo et al. 2002; Coops et al. 2004; Mitchell et al. 2006). The practice of Moist-soil Management (MSM) includes flooding or drawdown in spring or summer to create suitable water level in wetlands for promoting the germination, growth, and seed production of plants, followed by flooding in fall or winter to allow waterbirds access to habitats and food. SMM and MSM effectively increase waterbird richness, abundance in wetlands, and have been widely applied in North America (e.g., Anderson and Smith 1999, 2000; Kaminski et al. 2006) and Europe (e.g., Coops et al. 2004).
In coastal wetlands, periodic tidal flooding is the dominant force determining the structure and function of ecosystems (Pennings and Bertness 2001). Because most alteration or degradation of coastal wetlands is due to the isolation of tidal flooding through construction of dikes during the past century, diked areas are re-opened to reestablish tidal exchange and restore the hydrological conditions of coastal wetlands (Konisky et al. 2006; Mitchell et al. 2006). Combined with modification of topography (such as grading substrate to proper elevations, placing dredged spoils to raise low areas, or excavating uplands to create dense tidal creek networks), the re-opening of diked areas can generate a diverse spatial structure and provides various habitats for waterbirds on restored coastal wetlands (Morgan and Short 2002). However, the changes in plant communities need to be considered because the plant species composition generally changes dramatically due to salinity changes (see above).
Abundant food attracts large numbers of waterbirds and is important for the formation of waterbird colonies in breeding periods (Bancroft et al. 1994). However, increasing food for waterbirds through wetland management can be difficult. Although the enhancement of nitrogen and phosphorous levels in water bodies increases productivity (e.g., Frost et al. 2009), which improves the species richness and abundance of waterbirds (Acuna et al. 1994; Hoyer and Canfield 1994; Holm and Clausen 2006), a hypertrophic state causes eutrophication of water and reduced water quality, which is obviously undesirable. Artificial feeding (e.g. providing corn to attract cranes), despite attracting waterbirds to feeding stations, is usually rejected because it concentrates the birds and because subsequently dispersing the flocks can be difficult. The concentration of birds increases their vulnerability to disease and tends to domesticate the birds and reduce their wariness of people (Ma et al. 2009). In addition, the task of managing food for waterbirds is complex because of cascading food-web effects and because different waterbird groups have diverse foraging requirement (Potthoff et al. 2008).
In coastal regions where managed wetlands are close to the intertidal area, an effective method of enhancing food resources is to obtain complementary food from the tidewater by constructing floodgates and sluices and adjusting the periodic exchange of tidewater (e.g., Brusati et al. 2001; Collazo et al. 2002). Also, maintaining suitable water conditions to support the colonization and reproduction of aquatic plants and invertebrates can enhance food for waterbirds in managed wetlands (Rehfisch 1994; Anderson and Smith 2000; Coops et al. 2004). Moreover, regulation of water depth so as to increase food accessibility for waterbirds is a common practice in wetland management. At the Blacktoft Sands Lagoons in Humber Estuary, England, the integrated management of tidewater exchange and water depth maintains high chironomid biomass in the lagoons and provides suitable foraging conditions for waterbirds (Rehfisch 1994).
Reestablishment of vegetation is an important target and success criterion of wetland restoration because vegetation provides the food-web base for the entire ecosystem (Davis and Bidwell 2008; Matthews and Endress 2008). The vegetation in restored wetlands can reestablish gradually post-construction if suitable hydrological and sediment conditions are provided. The planting or seeding of native species can accelerate the recovery of vegetation and discourage the establishment of unwanted species (Morgan and Short 2002; Ruiz-Jaen and Aide 2005; Armitage et al. 2006; Erwin and Beck 2007). Many studies have indicated that there is a strong correlation between vegetation structure and bird diversity (Losito and Baldassarre 1995; Froneman et al. 2001; Erwin and Beck 2007). However, this relationship might only occur in the early stage of wetland restoration. Prescribed fire is generally used to improve the habitats for waterbirds in the management of well-developed marsh vegetation when dense vegetation obstructs the foraging and movements of waterbirds (Mitchell et al. 2006). Fire removes litter and dead vegetation, or removes vegetation of little or no value to waterbirds. This decreases vegetation cover and creates sparse or unvegetated habitats, which increases the accessibility of the habitat for waterbirds (Nyman and Chabreck 1995). This is important when the dominant bird groups are shorebirds, terns, and others that use bare sand-shell and bare intertidal flats. With elaborate control, prescribed fire can maintain a mixture of open water and vegetated cover for the resting and foraging of diverse waterbirds. Also, burns stimulate growth or seed production of food plants eaten by waterbirds (de Szalay and Resh 1997). However, organic carbon stocks within the marsh are reallocated in the form of CO2 upon burning, which has negative consequences for global warming.
The hemi-marsh with 50:50 mixture of open water and emergent vegetation (Weller and Spatcher 1965; Weller and Fredrickson 1974) is a common wetland management recommendation to maintain high species rich and diversity of waterbirds in North America. The high species and diversity in the hemi-marsh might be caused by an increase of food, visual isolation and spacing, or increased edge and habitat diversity (Murkin et al. 1982; Smith et al. 2004). Although the hemi-marsh principle has been tested in waterfowl, especially dabbling ducks, in both breeding and wintering periods (Kaminski and Prince 1981; Smith et al. 2004), other waterbird groups may not indicate a significant preference for hemi-marsh (e.g., Murkin et al. 1997; Smith et al. 2004). More studies are needed to understand the applicability of hemi-marsh principle to other regions and to waterbird communities with different species composition.
Multiple-species conservation requires that habitat-based conservation programs be applied to provide wetland complexes at the landscape scale, whereby different wetlands or sections of wetlands are managed to meet the needs of multiple waterbird groups. For example, the rotation of drawdowns increases habitat heterogeneity and consequently provides the greatest benefits to a wide range of waterbird species over large spatial scales. At the same time, some wetlands need to be managed to meet the special needs of waterbirds. For instance, in the California rice-growing region, the flooding of rice fields provides habitats for most waterbird species. However, unflooded fields are also needed to provide habitats for Sandhill Cranes (Grus canadensis), which prefer to forage on uncultivated dryland (Elphick and Oring 1998). Diverse habitats can also meet various requirements for different activities of waterbirds. For example, vegetated areas benefit nesting, island uplands benefit roosting, and open shallow waters benefit foraging.
For most artificial wetlands such as paddy fields, aquacultural ponds, and salt ponds, obtaining economic benefit (profit) is a higher priority than providing habitat for waterbirds. Consequently, management of artificial wetlands for waterbirds must be integrated with the management for production. Because most artificial wetlands are not managed for production throughout the whole year (such as the fallow of paddy fields, aquacultural ponds, and salt ponds in winter), it is possible to temporally stagger the two different kinds of management activities. For example, flooding fallow paddy fields in winter provides foraging habitats with abundant food (gleanings, aquatic plants, and invertebrates) for waterbirds (Elphick 2000). Also, maintenance of flooded conditions in the evaporation pans of saltworks in winter provides foraging and roosting habitats for shorebirds (Masero 2003). Although predation by waterbirds decreases the abundance and productivity of aquatic organisms (fish, crab, shrimp, etc.) in aquacultural ponds, Huner (2002) indicated that waterbirds might consume only small and less commercially valuable organisms thus enhancing the production of larger more valuable organisms. Moreover, foraging by waterbirds in paddy fields can suppress plant pests (Fasola and Ruíz 1997; van Groenigen et al. 2003) and accelerate straw decomposition (Bird et al. 2000), which benefit agricultural production. Producers must be compensated, however, when waterbirds reduce the economic benefits of artificial wetlands (Jensen et al. 2008).
Social conditions also affect how wetlands are managed. Because wetland management projects, especially those mitigation and restoration projects, are often financially supported by the public via higher taxes, public preferences should be considered in developing and implementing wetland management so that the public continues to support that management (Bauer et al. 2004; Nakamura et al. 2006). Therefore, there is a need for scientists and managers to exchange information on wetland management practice with the public. Otherwise, the public support for management projects will decline gradually, especially considering the amount of money involved (Nakamura et al. 2006). Wetland management can also be affected by the stakeholders (Pyrovetsi 1997). Although different stakeholders have various, and generally conflicting, concerns, all the stakeholder concerns need to be identified and addressed early in the process to avoid conflict (Weishar et al. 2005). Failure to involve stakeholders can result in delays in permitting and possible litigation during or after the project has been completed (Weishar et al. 2005). Because 60% of the global human population lives in coastal and surrounding regions (Adam 2002), where most wetland management projects are implemented, the involvement of local social conditions are important for the success of wetland management projects.
Overall, ecosystem-based approach is needed in wetland management with various targets, including management with the goal of providing waterbird habitat. This requires integrated knowledge of the entire wetland ecosystem (including hydrology, geology, agrology, botany, aquatic biology, landscape ecology, engineering, and ornithology) with considering multiple spatial scales, temporal variability, and the diverse habitat requirements of waterbirds (Parsons 2002; Anteau and Afton 2008; Euliss et al. 2008; King et al. 2009). Moreover, the social and economic constrains need to be considered when making management targets and through the management practice to coordinate wetland management with local development. In addition, monitoring the dynamics of waterbirds in long-term is important for adjusting management measures whenever necessary according to the wetland dynamics and management targets. Figure 2 indicates the framework of wetland management for providing waterbird habitats. This provides a reference for the managers to design and implement wetland management projects.

Framework of wetland management practice for providing waterbird habitats
Priorities for Future Research and Management
The number of quantitative studies of how habitats affect the use of wetlands by waterbirds has been rapidly increasing over the past two decades. These studies provide a large reference database for the practice of wetland management. Priorities for future research and management include the following aspects:
-
1)
Most quantitative studies in managed wetlands have been conducted in artificial wetlands (especially in paddy fields and salt ponds), which are relatively simple systems, and there has been little in-depth research on how habitat variables affect waterbird communities in restored ones, which are generally more complex than the artificial ones. The restored wetlands, especially the newly restored ones, are very dynamic environments. Some aspects of restored wetlands may take several decades to stabilize after construction (e.g., soil organic matter content, Zedler 2001; Morgan and Short 2002). Consequently, the waterbird communities in recently restored wetlands also change over time. However, most knowledge about the temporal dynamics of waterbirds and habitat variables in response to management comes from short-term studies (e.g., prescribed fire, Mitchell et al. 2006). A long-term studies and monitoring is needed to understand the temporal and spatial dynamics of waterbird communities and the processes and functioning of restored wetlands. This will be of much help to the design and implementation of effective wetland management.
-
2)
In addition to directly affecting use of wetlands by waterbirds, habitat variables also interact to indirectly affect the waterbird use of wetlands (Fig. 3). This suggests that all the variables related to the habitat use of waterbirds should be considered together for wetland management. Currently, however, wetland management usually focuses on very few (one or a couple of) habitat variables. The interaction of the variables remains largely unexplored in both research and practice. This requires integrated experiments at the entire ecosystem scales with collaboration among experts in different disciplines. Simultaneously, rigorous experiments at small to medium scales are needed to separate variables in order to better understand their individual effects (see review of Elphick 1996).
Fig. 3 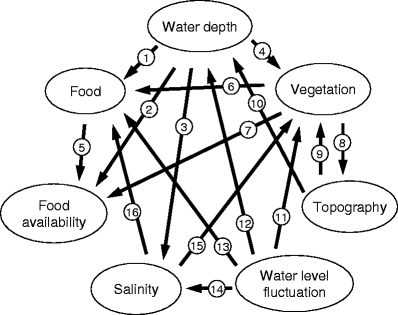
Interactions between the major habitat variables affecting habitat use of waterbirds at wetlands in situ. The directions of arrows indicate the directions of influences. Examples of these influences include 1: Water depth affecting the distribution of aquatic plants and animals serving as food for waterbirds; 2: Water depth limiting food availability of waterbirds; 3: Increase of salinity when water recedes; 4: Water depth affecting the distribution of vegetation; 5: Food size affecting its availability to waterbirds; 6: Vegetation supporting aquatic animals as food for waterbirds; 7: Dense vegetation obstructing the foraging of waterbirds; 8: vegetation functionally affecting topography because some waterbirds can walk on top of vegetation; 9: Topography affecting the distribution of vegetation in water body; 10: Topography affecting the variability of water depth; 11: Water level fluctuation affecting the growth of plants and the vegetation physiognomy and structure; 12: Water level fluctuation resulting in the changes of water depth; 13: Water level fluctuation affecting the growth of aquatic plants as food for waterbirds; 14: Water level fluctuation caused by precipitation and evaporation affecting water salinity; 15: Salinity affecting the distribution of plants; 16: Salinity affecting the aquatic plants and animals as food for waterbirds
-
3)
Most studies on waterbirds and their habitats on managed wetlands focus on species richness and abundance/density of waterbirds, which reflect habitat use by waterbirds. Species richness, population abundance and the derived indexes (e.g., saturation index of species pool, Wolters et al. 2005) are also generally used as succession criteria in evaluating wetland restoration (Neckles et al. 2002; Konisky et al. 2006). Habitat use, however, might not correctly reflect the habitat requirements of waterbirds or the quality of wetland habitats (van Horne 1983). Future research should assess habitat requirements and quality in terms of net energy acquired, time budgets of different behaviors, and population demographics (e.g., nesting and breeding success, survival rate) of waterbirds.
-
4)
Although there have been many studies providing reference for the practice of wetland management, the practice of management varies with, and is influenced by, region, wetland type, and function and degree of wetland degradation (Kentula 2000). Thus locally based research is needed to make a specific guidance for the practice of wetland management. However, our understanding of wetland management is still largely academic, and the practice of wetland management is generally out of touch with the academic research. It is now time to apply the knowledge gained by research to the decision-making processes and the practice of wetland management. Also, the effectiveness of the academic knowledge needs to be evaluated in the management practice.
-
5)
In order to guide the practice of wetland management for providing waterbird habitat, it is necessary to develop effective tools of predicting the effects of wetland management on the dynamics of waterbirds and their habitats. This requires simplified decision-supporting systems on the basis of complex multidisciplinary knowledge. The socioeconomic scenarios can also be involved in the systems to provide an integrated prediction of wetland management.

References
Acuna R, Contreras F, Kerekes J (1994) Aquatic bird densities in two coastal lagoon systems in Chiapas State, Mexico, a preliminary assessment. Hydrobiologia 279(280):101–106
Adam P (2002) Salt marshes in a time of change. Environmental Conservation 29:39–61
Anderson JT, Smith LM (1999) Carrying capacity and diel use of managed playa wetlands by nonbreeding waterbirds. Wildlife Society Bulletin 27:281–291
Anderson JT, Smith LM (2000) Invertebrate response to moist-soil management of playa wetlands. Ecological Applications 10:550–558
Anteau MJ, Afton AD (2008) Amphipod densities and indices of wetland quality across the upper-midwest, USA. Wetlands 28:184–196
Armitage AR, Boyer KE, Vance RR, Ambrose RF (2006) Restoring assemblages of salt marsh halophytes in the presence of a rapidly colonizing dominant species. Wetlands 26:667–676
Baker MC (1979) Morphological correlates of habitat selection in a community of shorebirds (Charadriiformes). Oikos 33:121–126
Balcombe CK, Anderson JT, Fortney RH, Kordek WS (2005) Aquatic macroinvertebrate assemblages in mitigated and natural wetlands. Hydrobiologia 541:175–188
Bancroft CT, Strong AM, Sawicki RJ, Hoffman W, Jewell SD (1994) Relationships among wading bird foraging patterns, colony locations, and hydrology in the Everglades. In: Davis SM, Ogden JC (eds) Everglades: The ecosystem and its restoration. St. Lucie, Delray Beach, pp 615–657
Bancroft GT, Gawlik DE, Rutchey K (2002) Distribution of wading birds relative to vegetation and water depths in the Northern Everglades of Florida, USA. Waterbirds 25:265–277
Bauer DM, Cyr NE, Swallow SK (2004) Public preferences for compensatory mitigation of salt marsh losses: a contingent choice of alternatives. Conservation Biology 18:401–411
Bird JA, Pettygrove GS, Eadie JM (2000) The impact of waterfowl foraging on the decomposition of rice straw: mutual benefits for rice growers and waterfowl. Journal of Applied Ecology 37:728–741
Bolduc F, Afton AD (2004) Relationships between wintering waterbirds and invertebrates, sediments and hydrology of coastal marsh ponds. Waterbirds 27:333–341
Brown M, Dinsmore JJ (1986) Implications of marsh size and isolation for marsh bird management. Journal of Wildlife Management 50:392–397
Brusati ED, DuBowy PJ, Lacher TE Jr (2001) Comparing ecological functions of natural and created wetlands for shorebirds in Texas. Waterbirds 24:371–380
Collazo JA, O’Harra DA, Kelly CA (2002) Accessible habitat for shorebirds: factors influencing its availability and conservation implications. Waterbirds 25(Suppl. 2):13–24
Colwell MA (1993) Shorebird community patterns in a seasonally dynamic estuary. Condor 95:104–114
Colwell MA, Taft OW (2000) Waterbird communities in managed wetlands of varying water depth. Waterbirds 23:45–55
Connor KJ, Gabor S (2006) Breeding waterbird wetland habitat availability and response to water-level management in Saint John River floodplain wetlands, New Brunswick. Hydrobiologia 567:169–181
Coops H, Vulink JT, van Nes EH (2004) Managed water levels and the expansion of emergent vegetation along a lakeshore. Limnologia 34:57–64
Craig RJ, Beal KG (1992) The influence of habitat variables on marsh bird communities of the Connecticut River estuary. Wilson Bulletin 104:295–311
Czech HA, Parsons KC (2002) Agricultural wetlands and waterbirds: a review. Waterbirds 25(suppl. 2):56–65
Darnell T, Smith EH (2004) Avian use of natural and created salt marsh in Texas, USA. Waterbirds 27:355–361
Davidson NC, Evans PR (1986) The role and potential of man-made and man-modified wetlands in the enhancement of the survival of overwintering shorebirds. Colonial Waterbirds 9:176–188
Davis CA, Smith LM (1998) Ecology and management of migrant shorebirds in the Playa Lakes region of Texas. Wildlife Monographs 140:1–45
Davis CA, Smith LM (2001) Foraging strategies and niche dynamics of coexisting shorebirds at stopover sites in the southern Great Plains. The Auk 118:484–495
Davis CA, Bidwell JR (2008) Response of aquatic invertebrates to vegetation management and agriculture. Wetlands 28:793–805
de Szalay FA, Resh VH (1997) Responses of wetland invertebrates and plants important in waterfowl diets to burning and mowing of emergent vegetation. Wetlands 17:149–156
Dekinga A, Dietz MW, Koolhaas A, Piersam T (2001) Time course and reversibility of changes in the gizzards of red knots alternately eating hard and soft food. Journal of Experimental Biology 204:2167–2173
Desrochers DW, Keagy JC, Cristol DA (2008) Created versus natural wetlands: Avian communities in Virginia salt marshes. Ecoscience 15:36–42
Dimalexis A, Pyrovetsi M (1997) Effect of water level fluctuations on wading bird foraging habitat use at an irrigation reservoir, Lake Kerkini, Greece. Colonial Waterbirds 20:244–252
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65:169–175
Elphick CS (1996) Experimental approaches to shorebird habitat management. International Wader Studies 9:20–28
Elphick CS (2000) Functional equivalency between rice fields and seminatural wetland habitats. Conservation Biology 14:181–191
Elphick CS (2004) Assessing conservation trade-offs: identifying the effects of flooding rice fields for waterbirds on non-target bird species. Biological Conservation 117:105–110
Elphick CS, Oring LW (1998) Winter management of Californian rice fields for waterbirds. Journal of Applied Ecology 35:95–108
Elphick CS, Oring LW (2003) Conservation implications of flooding rice fields on winter waterbird communities. Agriculture, Ecosystems and Environment 94:17–29
Erwin RM (2002) Integrated management of waterbirds: beyond the conventional. Waterbirds 25(suppl. 2):5–12
Erwin RM, Beck RA (2007) Restoration of waterbird habitats in Chesapeake Bay: great expectations or Sisyphus revisited? Waterbirds 30 (Special Publication 1): 163–176
Erwin RM, Hatfield JS, Howe MQ, Klugman SS (1994) Waterbird use of saltmarsh ponds created for open water marsh management. Journal of Wildlife Management 58:516–524
Euliss NH Jr, Jarvis RL, Gilmer DS (1989) Carbonate deposition on tail feathers of Ruddy Ducks using evaporation ponds. Condor 99:803–806
Euliss NH Jr, LaBaugh JW, Fredrickson LH, Musher DM, Laubhan MK, Swanson GA, Winter TC, Rosenberry DO, Nelson RD (2004) The wetland continuum: a conceptual framework for interpreting biological studies. Wetlands 24:448–458
Euliss NH Jr, Smith LM, Wilcox DA, Browne BA (2008) Linking ecosystem processes with wetland management goals: charting a course for a sustainable future. Wetlands 28:553–562
Fasola M, Ruíz X (1997) Rice farming and waterbirds: Integrated management in an artificial landscape. In: Pain DJ, Pienkowski MW (eds) Farming and birds in Europe: the common agricultural policy and its implications for bird conservation. Academic, San Diego, pp 210–235
Fraser LH, Keddy PA (2005) The world’s largest wetlands: Ecology and conservation. Cambridge University Press, Cambridge
Fredrickson LH, Reid FA (1986) Wetland and riparian habitats: a nongame management overview. In: Hale JB, Best LB, Clawson RL (eds) Management of nongame wildlife in the Midwest: A developing art. North Central Section of the Wildlife Society, Chelsea, pp 59–96
Froneman A, Mangnall MJ, Little RM, Crowe TM (2001) Waterbird assemblages and associated habitat characteristics of farm ponds in the Western Cape, South Africa. Biodiversity and Conservation 10:251–270
Frost JW, Schleicher T, Craft C (2009) Effects of nitrogen and phosphorus additions on primary production and invertebrate densities in a Georgia (USA) tidal freshwater marsh. Wetlands 29:196–203
Fujioka M, Armacost JW, Yoshida H, Maeda T (2001) Value of fallow farmlands as summer habitats for waterbirds in a Japanese rural area. Ecological Research 16:555–567
Gawlik DE (2002) The effects of prey availability on the numerical response of wading birds. Ecological Monography 72:329–346
Gordon DH, Gray BT, Kaminski RM (1998) Dabbling duck-habitat associations during winter in coastal South Carolina. Journal of Wildlife Management 62:569–580
Guillemain M, Fritz H (2002) Temporal variation in feeding tactics: exploring the role of competition and predators in wintering dabbling ducks. Wildlife Biology 8:81–90
Hammer DA (1997) Creating freshwater wetlands, 2nd edn. Lewis, Boca Raton
Hannam KM, Oring LW, Herzog MP (2003) Impacts of salinity on growth and behavior of American Avocet chicks. Waterbirds 26:119–125
Hartke KM, Kriegel KH, Nelson GM, Merendino MT (2009) Abundance of wigeongrass during winter and use by herbivorous waterbirds in a Texas coastal marsh. Wetlands 29:288–293
Hattori A, Mae S (2001) Habitat use and diversity of waterbirds in a coastal lagoon around Lake Biwa, Japan. Ecological Research 16:543–553
Hoffman W, Bancroft CT, Sawicki RJ (1994) Foraging habitat of wading birds in the Water Conservation Areas of the Everglades. In: Davis SM, Ogden JC (eds) Everglades: The ecosystem and its restoration. St. Lucie, Delray Beach, pp 585–614
Hobaugh WC, Stutzenbaker CD, Flickinger EL (1989) The rice prairies. In: Smith LM, Pederson RL, Kaminski RM (eds) Habitat management for migrating and wintering waterfowl in North America. Texas Tech Univ. Press, Lubbock, pp 367–383
Holm TE (2002) Habitat use and activity patterns of Mute Swans at a molting and a wintering site in Denmark. Waterbirds 25(Special Publication 1):183–191
Holm TE, Clausen P (2006) Effects of water level management on autumn staging waterbird and macrophyte diversity in three Danish coastal lagoons. Biodiversity and Conservation 15:4399–4423
Hoover JP (2009) Effects of hydrologic restoration on birds breeding in forested wetlands. Wetlands 29:563–573
Hoyer MV, Canfield DE Jr (1994) Bird abundance and species richness on Florida lakes: influence of tropic status, lake morphology and aquatic macrophytes. Hydrobiologia 279(280):107–119
Huner JV (2002) Managing agricultural wetlands for waterbirds in the coastal regions of Louisiana, USA. Waterbirds 25(suppl. 2):66–78
Isola CR, Colwell MA, Taft OW, Safran RJ (2002) Interspecific differences in habitat use of shorebirds and waterfowl foraging in managed wetlands of California’s San Joaquin Valley. Waterbirds 25(suppl. 2):196–203
Jensen RA, Wisz MS, Madsen J (2008) Prioritizing refuge sites for migratory geese to alleviate conflicts with agriculture. Biological Conservation 141:1806–1818
Kaminski RM, Prince HH (1981) Dabbling duck and aquatic macroinvertebrate responses to manipulated wetland habitat. Journal of Wildlife Management 45:1–15
Kaminski MR, Baldassarre GA, Pearse AT (2006) Waterbird responses to hydrological management of Wetlands Reserve Program habitats in New York. Wildlife Society Bulletin 34:921–926
Kelly JP, Stralberg D, Etienne K, McCaustland M (2008) Landscape influence on the quality of heron and egret colony sites. Wetlands 28:257–275
Kentula ME (2000) Perspectives on setting success criteria for wetland restoration. Ecological Engineering 15:199–209
Kersten M, Britton RH, Dugan PJ, Hafner H (1991) Flock feeding and food intake in little egrets: the effects of prey distribution and behavior. Journal of Animal Ecology 60:241–252
King SL, Sharitz RR, Groninger JW, Battaglia LL (2009) The ecology, restoration, and management of southeastern floodplain ecosystems: A synthesis. Wetlands 29:624–634
Kingsford RT, Porter JL (1994) Waterbirds on an adjacent freshwater lake and salt lake in arid Australia. Biological Conservation 69:219–228
Konisky RA, Burdick DM, Dionne MD, Neckles HA (2006) A regional assessment of salt marsh restoration and monitoring in the Gulf of Maine. Restoration Ecology 14:516–525
Kushlan JA (1978) Feeding ecology of wading birds. In: Sprunt A IV, Ogden JC, Winckler S (eds) Wading birds. National Audubon Society, New York, pp 249–298
Kushlan JA (1986) The management of wetlands for aquatic birds. Colonial Waterbirds 9:246–248
Kushlan JA, Hafner H (2000) Heron conservation. Academic, London
Little C (2000) The biology of soft shores and estuaries. Oxford University Press, Oxford
Losito MP, Baldassarre GA (1995) Wetland use by breeding and postbreeding female mallards in the St. Lawrence River Valley. Wilson Bulletin 107:55–63
Ma ZJ, Li B, Jing K, Tang SM, Chen JK (2004) Are artificial wetlands good alternatives to natural wetlands for waterbirds? A case study on Chongming Island, China. Biodiversity and Conservation 13:333–350
Ma ZJ, Li B, Li WJ, Han NY, Chen JK, Watkinson A (2009) Conflicts between biodiversity conservation and development in a biosphere reserve. Journal of Applied Ecology 46:527–535
Maeda T (2001) Patterns of bird abundance and habitat use in rice fields of the Kanto Plain, central Japan. Ecological Research 16:569–585
Masero JA (2003) Assessing alternative anthropogenic habitats for conserving waterbirds: salinas as buffer areas against the impact of natural habitat loss for shorebirds. Biodiversity and Conservation 12:1157–1173
Masero JA, Pérez-Hurtado A, Castro M, Arroyo GM (2000) Complementary use of intertidal mudflats and adjacent salinas by foraging waders. Ardea 88:177–191
Matthews JW, Endress AG (2008) Performance criteria, compliance success, and vegetation development in compensatory mitigation wetlands. Environmental Management 41:130–141
Melvin SL, Gawlik DE, Scharff T (1999) Long-term movement patterns for seven species of wading birds. Waterbirds 22:411–416
Mitchell LR, Gabrey S, Marra PP, Erwin RM (2006) Impacts of marsh management on coastal-marsh birds habitats. Studies in Avian Biology 32:155–175
Mitsch WJ, Wilson RF (1996) Improving the success of wetland creation and restoration with know-how, time, and self-design. Ecological Applications 6:77–83
Morgan PA, Short FT (2002) Using functional trajectories to track constructed salt marsh development in the Great Bay Estuary, Maine/New Hampshire, USA. Restoration Ecology 10:461–473
Murkin HR, Kaminski RM, Titman RD (1982) Responses by dabbling ducks and aquatic invertebrates to an experimentally manipulated cattail marsh. Canadian Journal of Zoology 60:2324–2332
Murkin HR, Murkin EJ, Ball JP (1997) Avian habitat selection and prairie wetland dynamics: a 10-year experiment. Ecological Applications 7:1144–1159
Nakamura K, Tockner K, Amano K (2006) River and wetland restoration: lessons from Japan. Bioscience 56:419–429
Neckles HA, Dionne M, Burdick DM, Roman CT, Buchsbaum R, Hutchins E (2002) A monitoring protocol to assess tidal restoration of salt marshes on local and regional scales. Restoration Ecology 10:556–563
Nielsen SL, Sand-Jensen K, Borum J, Geertz-Hansen O (2002) Depth colonization of eelgrass (Zostera marina) and macroalgae as determined by water transparency in Danish coastal waters. Estuaries Coasts 25:1025–1032
Nolet BA, Bevan RV, Klaassen M, Langevoord O, van der Heijden YGJT (2002) Habitat switching by Bewick’s swans: maximization of average long-term energy gain? Journal of Animal Ecology 71:979–993
Ntiamoa-Baidu Y, Piersma T, Wiersma P, Poot M, Battley P, Gordon C (1998) Water depth selection, daily feeding routines and diets of waterbirds in coastal lagoons in Ghana. Ibis 140:89–103
Nyman JA, Chabreck RH (1995) Fire in coastal marshes: history and recent concerns. Proceedings of the Annual Tall Timbers Fire Ecology Conference 19:134–141
Ogden JC (1991) Nesting by wood storks in natural, altered, and artificial wetlands in central and northern Florida. Colonial Waterbirds 14:39–45
Ogden JC (1994) A comparison of wading bird nesting colony dynamics (1931–1946 and 1974–1989) as in indication of ecosystem conditions in the Southern Everglades. In: Davis SM, Ogden JC (eds) Everglades: The ecosystem and its restoration. St. Lucie, Delray Beach, pp 533–570
O’Neal BJ, Heske EJ, Stafford JD (2008) Waterbird response to wetlands restored through the conservation reserve enhancement program. Journal of Wildlife Management 72:654–664
Owen M, Black JM (1990) Waterfowl ecology. Blackie and Son Ltd., Glasgow
Paracuellos M (2006) How can habitat selection affect the use of a wetland complex by waterbirds. Biodiversity and Conservation 15:4569–4582
Paracuellos M, Telleria JL (2004) Factors affecting the distribution of a waterbird community: the role of habitat configuration and bird abundance. Waterbirds 27:446–453
Parnell JF, Needham RN, Soots RF Jr, Fussell JOIII, Dumond DM, McCrimmon DA Jr, Bjork RD, Shields MA (1986) Use of dredged-material deposition sites by birds in coastal North Carolina, USA. Colonial Waterbirds 9:210–217
Parsons KC (2002) Integrated management of waterbird habitats at impounded wetlands in Delaware Bay, USA. Waterbirds 25(suppl. 2):25–41
Pennings SC, Bertness MD (2001) Salt marsh communities. In: Bertness MD, Gaines SD, Hay M (eds) Marine community ecology. Sinauer, Sunderland, pp 289–316
Plissner JH, Haig SM, Oring LW (2000) Postbreeding movements of American avocets and implications for wetland connectivity in the Western Great Basin. The Auk 117:290–298
Poysa H (1983) Resource utilization pattern and guild structure in a waterfowl community. Oikos 40:295–307
Post W (1998) Reproduction of least bitterns in a managed wetland. Colonial Waterbirds 21:268–273
Potthoff AJ, Herwig BR, Hanson MA, Zimmer KD, Butler MG, Reed JR, Parsons BG, Ward MC (2008) Cascading food-web effects of piscivore introductions in shallow lakes. Journal of Applied Ecology 45:1170–1179
Powell GVN (1987) Habitat us by wading birds in a subtropical estuary: implications of hydrography. The Auk 104:740–749
Purdue JR, Haines H (1977) Salt water tolerance and water turnover in the snowy plover. The Auk 94:248–255
Pyrovetsi M (1997) Integrated management to create new breeding habitat for Dalmatian pelicans (Pelecanus crispus) in Greece. Environmental Management 21:657–667
Rehfisch MM (1994) Man-made lagoons and how their attractiveness to waders might be increased by manipulating the biomass of an insect benthos. Journal of Applied Ecology 31:383–401
Rubega MA, Robinson JA (1997) Water salinization and shorebirds: emerging issues. International Wader Studies 9:45–54
Ruiz-Jaen MC, Aide TM (2005) Restoration success: how is it being measured? Restoration Ecology 13:569–577
Sánchez-Zapata JA, Anadón JD, Carrete M, Giménez A, Navarro J, Villacorta C, Botella F (2005) Breeding waterbirds in relation to artificial pond attributes: implications for the design of irrigation facilities. Biodiversity and Conservation 14:1627–1639
Sanders MD (2000) Enhancing food supplies for waders: inconsistent effects of substratum manipulations on aquatic invertebrate biomass. Journal of Applied Ecology 37:66–76
Scheffer M, van Geest GJ, Zimmer K, Butler MG, Hanson MA, Declerck S, de Meester L, Jeppesen E, Sondergaard M (2006) Small habitat size and isolation can promote species richness: second-order effects on biodiversity in shallow lakes and ponds. Oikos 112:227–231
Skagen SK, Knopf FL (1993) Toward conservation of midcontinental shorebird migrations. Conservation Biology 7:533–541
Smith LM, Haukos DA, Prather RM (2004) Avian response to vegetative pattern in playa wetlands during winter. Wildlife Society Bulletin 32:474–480
Soulé ME, Simberloff D (1986) What do genetics and ecology tell us about the design of nature reserves? Biological Conservation 35:19–40
Stapanian MA (2003) Species density of waterbirds in offshore habitats in western Lake Erie. Journal of Field Ornithology 74:381–393
Stralberg D, Applegate DL, Phillips SJ, Herzog MP, Nur N, Warnock N (2009) Optimizing wetland restoration and management for avian communities using a mixed integer programming approach. Biological Conservation 142:94–109
Sustainable Ecosystems Institute (2007) Everglades multi-species avian ecology and restoration review, Final Report, November 2007. Sustainable Ecosystems Institute, Portland, Oregon. http://www.sei.org/everglades/reports.htm
Taft OW, Haig SM (2003) Historical wetlands in Oregon’s Willamette valley: implications for restoration of winter waterbird habitat. Wetlands 23:51–64
Taft OW, Haig SM (2005) The value of agricultural wetlands as invertebrate resources for wintering shorebirds. Agriculture, Ecosystems and Environment 110:249–256
Taft OW, Colwell MA, Isola CR, Safran RJ (2002) Waterbird responses to experimental drawdown: implications for multispecies management of wetland mosaics. Journal of Applied Ecology 39:987–1001
Takekawa JY, Miles AK, Schoellhamer DH, Athearn ND, Saiki MK, Duffy WD, Kleinschmidt S, Shellenbarger GG, Jannusch CA (2006) Trophic structure and avian communities across a salinity gradient in evaporation ponds of the San Francisco Bay estuary. Hydrobiologia 567:307–327
Thébault J, Schraga TS, Cloern JE, Dunlavey EG (2008) primary production and carrying capacity of former salt ponds after reconnection to San Francisco bay. Wetlands 28:841–851
van Groenigen JW, Burns EG, Eadie JM, Horwath WR (2003) Effects of foraging waterfowl in winter flooded rice fields on weed stress and residue decomposition. Agriculture, Ecosystems and Environment 95:289–296
van Horne B (1983) Density as a misleading indicator of habitat quality. Journal of Wildlife Management 47:893–901
VanRees-Siewert KL, Dinsmore JJ (1996) Influence of wetland age on bird use of restored wetlands in Iowa. Wetlands 16:577–582
Velasquez CR (1992) Managing artificial saltpans as a waterbird habitat: species responses to water level manipulation. Colonial Waterbirds 15:43–55
Warnock SE, Takekawa JY (1995) Habitat preferences of wintering shorebirds in a temporally changing environment: Western Sandpipers in the San Francisco Bay estuary. The Auk 112:920–930
Warnock SE, Takekawa JY (1996) Wintering site fidelity and movement patterns of Western Sandpipers Calidris mauri in the San Francisco Bay estuary. Ibis 138:160–167
Warnock N, Page GW, Stenzel LE (1995) Non-migratory movements of dunlins on their California wintering grounds. Wilson Bulletin 107:131–139
Warnock N, Page GW, Ruhlen TD, Nur N, Takekawa JY, Hanson JT (2002) Management and conservation of San Francisco Bay salt ponds: effects of pond salinity, area, tide, and season on Pacific flyway waterbirds. Waterbirds 25(Suppl. 2):79–92
Weber LM, Haig SM (1996) Shorebird use of south Carolina managed and natural coastal wetlands. Journal of Wildlife Management 60:73–82
Weishar LL, Teal JM, Hinkle R (2005) Designing large-scale wetland restoration for Delaware Bay. Ecological Engineering 25:231–239
Weller MW, Spatcher CE (1965) Role of habitat in the distribution and abundance of marsh birds. Iowa Agriculture and Home Economics Experiment Station, Special Report 43. Ames, USA
Weller MW, Fredrickson LH (1974) Avian ecology of a managed glacial marsh. Living Bird 12:269–291
White CL, Main MB (2004) Habitat value of golf course wetlands to waterbirds. USGA Turfgrass and Environmental Research Online 3(16):1–10
Wiggins GB, Mackay RJ, Smith IM (1980) Evolutionary and ecological strategies of animals in annual temporary pools. Archives of Hydrobiology Supplement 58:97–206
Winter TC, Woo MK (1990) Hydrology of lakes and wetlands. In: Wolman MG, Riggs HC (eds) The geology of North America, Vol. 1: Surface water hydrology. The Geological Society of America, Boulder, pp 159–187
Wolters M, Garbutt A, Bakker JP (2005) Salt-marsh restoration: evaluating the success of de-embankments in north-west Europe. Biological Conservation 13:249–268
Zedler J (ed) (2001) Handbook for restoring tidal marshes. CRC, Boca Raton
Zhu X, Zou XP (eds) (2001) The herons, egrets and bitterns in China. China Forestry Publishing House, Beijing
Zwarts L, Blomert AM (1992) Why knot Calidris canutus take medium-sized Macoma balthica when six prey species are available. Marine Ecology Progress Series 83:113–128
Acknowledgments
This study was financially supported by the National Basic Research Program of China (2006CB403305), National Natural Science Foundation of China (30670269), Shanghai Education Commission (08ZZ04) and the Science & Technology Department of Shanghai (07DZ12038). We thank R. Michael Erwin, Chris S. Elphick, and Bruce Jaffee for their constructive comments on earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ma, Z., Cai, Y., Li, B. et al. Managing Wetland Habitats for Waterbirds: An International Perspective. Wetlands 30, 15–27 (2010). https://doi.org/10.1007/s13157-009-0001-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-009-0001-6