
Abstract
Determination of a microbial strain for the joining into sustenance items requires both in vitro and in vivo assessment. A newly isolated bacteriocin-like inhibitory substance (BLIS) producing lactic acid bacterium, Lactococcus lactis Gh1, was isolated from a traditional flavour enhancer and evaluated in vitro for its potential applications in the food industry. Results from this study showed that L. lactis was tolerant to NaCl (≤ 4.0%, w/v), phenol (≤ 0.4%, w/v), 0.3% (w/v) bile salt, and pH 3. BLIS from L. lactis showed antimicrobial activity against Listeria monocytogenes ATCC 15313 and was susceptible to 10 types of antibiotics. The absence of haemolytic activity and the presence of acid phosphatase and naphthol-AS-BI-phosphohydrolase were observed in L. lactis. L. lactis could coagulate milk and showed a negative response to amylolytic and proteolytic activities and did not secrete β-galactosidase. The antimicrobial activity of BLIS was completely abolished at 121 °C. The BLIS was conserved at 4 °C in BHI and MRS medium up to 6–4 months, respectively. BLIS activity was more stable in BHI as compared to MRS after four freeze-thaw cycles and was not affected by a wide range of pH (pH 4–8). BLIS was sensitive to proteinase k and resistant to catalase and trypsin. The antimicrobial activity was slightly reduced by acetone, ethanol, methanol, and acetonitrile at 10% (v/v) and also towards Tween-80, urea, and NaCl 1% (v/v). Results from this study have demonstrated that L. lactis has a vast potential to be applied in the food industry, such as for the preparation of starter culture, functional foods, and probiotic products.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The idea of supplying functional foods that contain nutritious components is intended not only to relieve hunger and provide the essential nutrients but also to promote well-being for the host. These are one of the most important fields of food industry study and advancement, including foods with certain different nutrients or ingredients, including pro- and prebiotics [1]. In the human digestive system, probiotic properties and biological barrier resistance are different between species and even among individuals of the same species. Although a fair number of well-characterized probiotic strains are available commercially around the world, screening for new strains is still of great industrial interest [2]. In vitro evaluations are vital steps in searching for new potential lactic acid bacteria (LAB) strains to be applied in the food industry as the in vitro tests provide significant information concerning species and strain differences and are extremely helpful and strong instruments particularly for the rapid and efficient screening of bacterial probiotic activity [3].
LAB are one of the most significant groups of probiotic organisms which are commonly used in fermented dairy products [4]. LAB have been used extensively in food production for centuries without posing any health risks. As such, they are designated as generally regarded as safe (GRAS) microorganisms by the Food and Drug Administration (FDA) [5]. LAB are being used in the food industry due to their ability to inhibit or reduce contamination by spoilage and/or pathogen microorganisms through the production of various antimicrobial compounds such as organic acid, hydrogen peroxide, ethanol, diacetyl, carbon dioxide, bacteriocin, or antibiotic-like substances [6]. Bacteriocins are proteinaceous antimicrobial molecules with a diverse genetic origin, post-translationally modified or not, that can help the producer organism to outcompete other bacterial species [7]. Bacteriocins have been used as biopreservatives in various foods as an alternative to the use of chemical preservatives. Most bacteriocin producers belong to LAB, which occurs naturally in foods and has a long history of safe use in the dairy industry [8]. Besides possessing the GRAS status, LAB have also been granted the Qualified Presumption of Safety (QPS) status by the European Food Safety Authority (EFSA) [9]. Bacteriocins can be considered as safe since they are produced by LAB, and they can also be easily degraded by proteolytic enzymes of the mammalian gastrointestinal tract [8].
Bacteriocin-like inhibitory substance (BLIS) is often interchangeably used with bacteriocin since BLISs are uncharacterized substances that apparently share similar activity and can be identified by using the same quantification methods as bacteriocin [10]. Most of the food applications involving bacteriocins for the control of pathogenic and spoilage microorganisms in food can be applied on a purified/crude form or as a bacteriocin-producing LAB as a part of the fermentation process, or as adjuvant culture [8], and also in the form of cell-free supernatant (BLIS) [11].
Among the known bacteriocins reported, only nisin and pediocin have been approved as food preservatives. Their use in food, however, depends on many factors, such as pH, temperature, food composition, and target microbiota. The potential use of bacteriocin can thus be predicted through a detailed assessment of its sensitivity [8]. Like other proteins in the diet, many enzymes could digest bacteriocins and, hence, they are not detrimental to beneficial gut microflora. Bacteriocins are non-toxic and present in many foods and have been unwittingly consumed by mankind for thousands of years, being odourless, colourless, and tasteless [12]. Although bacteriocins are produced by many Gram-positive and Gram-negative bacterial species, only those produced by LAB are of particular importance in the food industry [13].
This study aims to provide a comprehensive assessment of potential probiotic strain Lactococcus lactis Gh1 and its BLIS to be used in the food industry. To achieve this purpose, the in vitro evaluation of the L. lactis Gh1 was designed to mimic the in vivo physiological environment of a gastrointestinal tract (GIT) to evaluate the desirable and undesirable characteristics of probiotic-related properties of this strain, the safety parameters that are usually considered for selection of probiotic, as well as their appropriateness to be applied in food handling. Apart from that, the antimicrobial characteristics of BLIS secreted by L. lactis Gh1 were also assessed to evaluate their potential in the food industry. Such observations are significant contributions for a better understanding of the functional characteristics of L. lactis Gh1 and the presence of BLIS in fermented foods.
Materials and Methods
Supplies
De Man Rogosa-Sharpe (MRS), Brain heart broth (BHI), Luria-Bertani (LB), and M17 broth and agar were purchased from Merck (Darmstadt, Germany). Antibiotic disc and bile salts were obtained from Oxoid Limited (Hampshire, England) and bromocresol purple from Friendemann Schmidt (Parkwood Western Australia). Taurodeoxycholic acid (TDC) and enzymes were purchased from Sigma (Sigma-Aldrich, St. Louis, MO, USA), and API ZYM from API Identification Systems (BioMérieux, France). All chemicals used in this study were analytical grade.
Bacterial Strains
The BLIS-producing strain, Lactococcus lactis Gh1, which was isolated from Ghara (Iranian traditional food enhancer), was used throughout this study [14]. The whole-genome shotgun sequence of this bacterium has been deposited in NCBI under GenBank accession number of SSBR01000004.1. The indicator microorganism was Listeria monocytogenes ATCC 15313, while Escherichia coli ATCC 25922 and Pediococcus acidilactici Kp10 were used as control microorganisms.
The stock cultures were MRS broth for L. lactis Gh1, BHI broth for L. monocytogenes ATCC 15313, LB broth for E. coli ATCC 25922 and M17 broth for P. acidilactici Kp10, each containing 20% (v/v) glycerol (BDH Laboratory Supplies, Poole, England), which were maintained at − 20 °C. Prior to use in experiments, the frozen stock was thawed and streaked onto respective agar plates and grown at their optimum temperatures. A single colony was picked and inoculated onto broth medium and incubated at 30 °C for L. lactis Gh1 and at 37 °C for L. monocytogenes ATCC 15313, E. coli ATCC 25922 and P. acidilactici Kp10 for 16 h. The culture was then sub-cultured in fresh broth and grown until the mid- or late exponential growth phase was reached (OD600 of 1.5–3.5).
Methodology for Characterization
The characterization methodology was divided into two sections, (i) physiological, biochemical, and characterization of L. lactis Gh1, and (ii) physio-chemical stability of BLIS activity produced by L. lactis Gh1. The parameter ranges tested were chosen based on the common practice applied in food-processing operations and imitate the conditions of the human GIT.
Physiological, Biochemical and Probiotic Characterization of L. lactis Gh1
Gram Stain, Cell Morphology and Catalase Test
Gram staining was performed using the standard Gram’s staining method. Meanwhile, cell morphology and colony appearance were examined by light microscopy and visual observation, respectively, based on basic microbiology guidelines. Catalase test was done by adding a drop of 3% (v/v) hydrogen peroxide solution to a heavily streaked colony. Instant formation of oxygen bubbles within 1 s indicated a positive reaction (presence of catalase in the cells).
Carbon Fermentation Test
To test the ability of the isolates to ferment carbon, nutrient agar was prepared with the addition of 1% (w/v) glucose or lactose and 0.004% (w/v) bromocresol purple as a pH indicator. About 10 μL of cultures was spotted on the prepared agar and incubated at 30 °C for 24 h. A yellow zone surrounding the culture after incubation indicated a positive result.
Effect of NaCl and Temperature on Growth of L. lactis Gh1
The culture of L. lactis Gh1 (1% v/v) was added into MRS broth containing different concentrations of NaCl (0.1–10%, w/v) and 0.004% (w/v) bromocresol purple followed by incubation at 30 °C for 48 h. To study the effect of temperature, the cultures (after the inoculation of MRS broth was added with 0.004% (w/v) bromocresol purple with L. lactis Gh1 (1%, v/v) inoculum) were incubated at various temperatures (− 20, 4, 10, 30, 35, 37, 40, 45, 50, 60, and 65 °C) for 48 h. Changes in colour from purple to yellow indicated positive results.
Effect of Low pH and Bile Salts on the Growth of L. lactis Gh1
MRS broth at pH 3 (adjusted using 0.5 N HCl) and MRS broth supplemented with 0.3% (w/v) bile salts were prepared. Then, 9 mL of the respective medium was inoculated with 1 mL of L. lactis Gh1 culture and incubated at 30 °C for 4 h. During incubation, growth was monitored as optical density at 650 nm using a spectrophotometer (Perkin Elmer, Lambda 25, Waltham, MA, USA) for every hour, and viable microorganisms were enumerated using the pour plate technique [14]. The number of colonies was determined using an electronic colony counter and compared with the control (0 h) to determine the acid and bile tolerance. The percentage of survival was calculated by dividing the log value of the colony-forming unit (CFU/mL) at any incubation time (1–4 h) with log CFU/mL at 0 h (control).
Effect of Phenol on the Growth of L. lactis Gh1
The tolerance of L. lactis Gh1 to phenol was performed by inoculating the cultures in MRS broth with the addition of phenol (0.1–0.4%, w/v). The samples (100 μL) were spread-plated onto MRS agar and incubated at 30 °C for 24 h. Bacterial survivability was enumerated using the formula as proposed by Xanthopoulos et al. [15], and the inhibition effect of phenol was calculated using the equation as follows:
Inhibition = Log10 (CFU/mL) at 0 h − Log10 (CFU/mL) at 24 h.
Antibiotic Sensitivity of L. lactis Gh1
The sensitivity of L. lactis Gh1 towards 18 types of antibiotics was determined by using the disc diffusion method [16]. A single colony was inoculated into MRS broth and incubated for 16–18 h at 30 °C. After incubation, the culture was diluted equivalent to 0.5 McFarland (1.5 × 108 CFU/mL) standard. Then, 100 μL of bacterial suspension was spread evenly on the surface of MRS agar plates by using sterile cotton wool before the plates were dried by air blowing in the laminar airflow.
Commercially available antibiotics discs containing penicillin G (10 units), sulfamethoxazole (100 μg), sulfamethoxazole (25 μg), sulphonamide (1000 μg), compound sulphonamides (300 μg), vancomycin (30 μg), erythromycin (15 μg), erythromycin (10 μg), trimethoprim (1.25 μg), trimethoprim (5 μg), streptomycin (10 μg), tetracycline (30 μg), chloramphenicol (30 μg), chloramphenicol (10 μg), sulphafurazole (300 μg), gentamicin (10 μg), rifampin (5 μg), ampicillin (10 μg), sulfamethoxazole-trimethoprim (23.75 μg/1.25 μg), oxytetracycline (30 μg) or amoxicillin (10 μg) were placed on the surface of the agar plates and incubated at 30 °C for 24 h.
Inhibition zones (including the disc diameter) were measured and categorized as sensitive (≥ 21 mm), intermediate (16–20 mm) or resistant (≤ 15 mm). The zone diameters of each drug were interpreted according to the Clinical & Laboratory Standards Institute (CLSI) [17], as previously described by Liasi et al. [18] and Vlková et al. [19].
β-Galactosidase Activity of L. lactis Gh1
The ability of L. lactis Gh1 to secrete β-galactosidase was tested based on the method described by Karasova et al. [20]. The colony of L. lactis Gh1 was incubated at 30 °C for 24 h on an MRS agar plate containing 0.01% (v/v) X-gal (5-bromo-4-chloro-3-indolyl β-D-galactopyranoside (Vivantis, Malaysia) and 0.1 mM IPTG (iso-propyl β-D-1-thiogalactopyranoside, Vivantis)) dissolved in dimethyl sulfoxide. The appearance of blue/green colonies was considered a positive result. P. acidilactici Kp10 was used as a positive control.
Qualitative Proteolytic Activity and Starch Hydrolysis of L. lactis Gh1
Heavy inoculum of the L. lactis Gh1 was streaked on 10% (w/v) skim milk agar and MRS-starch agar (0.2%, w/v soluble starch) and incubated at 30 °C for 24–48 h. Proteolytic activity was indicated by a clear zone surrounding colonies on skim milk agar while MRS-starch agar was topped with an iodine solution for the detection of starch hydrolysis [21]. L. monocytogenes ATCC 15313 and E. coli ATCC 25922 were used as negative controls.
Bile Salt Deconjugation of L. lactis Gh1
The ability of the strains to deconjugate bile salts was assessed according to the method described by Sharma et al. [22]. Bile salt plates were prepared by adding 0.5% (w/v) sodium salts of taurodeoxycholic acid (TDC) to MRS agar. Colonies of L. lactis Gh1 were streaked on the media, and the plates were anaerobically (GasPak System-Oxoid) and aerobically incubated at 30 °C for 72 h to evaluate the effect of atmospheric oxygen on bile salt hydrolase (BSH) activity. L. lactis Gh1 grown in MRS without supplementation of TDC was used as a negative control. The appearance of precipitated bile acid around colonies or the formation of opaque, granular white colonies with a silvery shine was considered as a positive result.
Acidification and Coagulation Activities of L. lactis Gh1
The effects of acidification and coagulation of L. lactis Gh1 were evaluated by inoculation of 10% (w/v) skim milk with 24-h-old culture at 1% (v/v) level and incubated in an incubator oven at 30 °C. Commencement of clotting, aroma and pH were made every 24 h up to 72 h of incubation [23].
Haemolytic Activity of L. lactis Gh1
The haemolytic activity was determined by growing the L. lactis Gh1 on MRS agar at 30 °C for 18 h that was then streaked onto Columbia agar plates containing 5% (v/v) of sheep blood (BioMe ́rieux, Hazelwood, MO, USA). The plates were incubated at 30 °C overnight. Haemolytic reactions were recorded by the presence of a clear zone of hydrolysis around the colonies (β-haemolysis), a green zone of a partial hydrolysis around the colonies (α-haemolysis) or no reaction indicated by the absence of a zone around the colonies (γ-haemolysis) [24].
Enzymatic Profile of L. lactis Gh1
Enzymatic characteristics of L. lactis Gh1 were determined using a commercial system, API ZYM strips, according to the manufacturer’s instructions. Colonies of pure culture of L. lactis Gh1 were suspended in sterile water before the initial suspension was adjusted to McFarland barium sulphate turbidity standard 6 (approximate 1.8 × 109 CFU/mL). The strips were incubated at 30 °C for 4 h, followed by the addition of reagents. Enzymatic activity was reported as positive when a score of 1 or higher was obtained after assessment of the colour intensity using the manufacturer’s colour chart.
Characterization of BLIS
Preparation of Cell-Free Supernatants
L. lactis Gh1 was grown in MRS broth for 16–18 h at 30 °C. Cell-free supernatant (CFS) was obtained by centrifugation at 12,000×g for 20 min at 4 °C (Rotor model 1189, Universal 22R centrifuge, Hettich AG, Bäch, Zurich, Switzerland). The CFS was separated from the pellet, and the clear CFS obtained was used in the subsequent experiments.
Antimicrobial Activity Test
The antimicrobial activity of the isolates was determined by the agar well diffusion method using CFS, as explained by Tagg et al. [25], with a slight modification. The supernatant (100 μL) was placed into 6-mm wells of agar plates that were previously seeded (1%, v/v) with the actively growing (106 CFU/mL) L. monocytogenes ATCC 15313. The plates were kept at 4 °C for 2 h to allow the diffusion of CFS on the medium followed with incubation at 37 °C. After 24 h, the growth inhibition zones were measured, and antimicrobial activity (AU/mL) was determined as described by Parente et al. [26]. The bacteriocin-producing strain P. acidilactici Kp10 [10, 14] was used as a positive control.
Effect of pH and Enzymes on BLIS Activity
The effect of pH on BLIS activity was evaluated by adjusting the pH of the CFS to 2–11 with 1 N HCl or 1 N NaOH [27]. The CFS was incubated at 30 °C for 1 h before measuring the BLIS activity. Sensitivity to enzymes was determined after 2 h of incubation with trypsin, pepsin, α-amylase and catalase at final concentrations of 1 and 0.1 mg/mL. The samples were then incubated at 30 °C, except for the samples containing trypsin and catalase, which were incubated at 25 °C and 37 °C, respectively.
Effect of Temperatures on BLIS Stability
The effect of temperature on BLIS stability was determined by treating the samples in a water bath (Cole Palmer Model 12104, 05, USA) at 60, 80, 90 and 100 °C for 5, 10 and 20 min. To study the effect of temperature at 121 °C, the CFS was autoclaved for 10, 15 and 20 min. The impact of long-term storage at the moderate and low refrigeration temperatures on supernatant stability was evaluated by placing supernatants at 30, 4 and − 20 °C for up to 6 months (180 days). The aliquots were taken from storage every month and were measured for antimicrobial activity. To test the stability of BLIS during three freeze-thaw cycles, the supernatants were frozen at − 20 °C for 24 h and thawed for 20 min at 5 °C. The inhibitory activity was tested and compared with the non-treated control. Residual inhibitory activity was measured using the agar well diffusion assay. In all cases, a positive control consisting of freshly prepared CFS was tested in parallel.
Effect of Chemicals on BLIS Stability
The effect of chemicals on antimicrobial activity was investigated by incorporating non-ionic (Tween 20, Tween 80, Triton X-100), anionic [sodium dodecyl sulphate (SDS)] surfactant, EDTA and urea to BLIS to yield the final concentration of 1% (w/v). Untreated BLIS and chemicals at this concentration in sterile distilled water were used as controls. The effect of chemicals, including organic solvents and surfactants, on the antimicrobial activity of BLIS was also investigated. Various organic solvents such as acetone, ethanol, butanol, methanol, toluene, acetonitrile and isopropyl alcohol were added to BLIS at 10 and 50% (v/v) of the total volume. Untreated BLIS solution and organic solvents with an equal amount of sterile distilled water were used as controls. All samples were incubated at 30 °C for 5 h before proceeding to the antimicrobial test [28].
Statistical Analysis
Data are expressed as a mean ± standard deviation calculated from three independent replications in triplicate. The findings were evaluated with the use of the one-way analysis of variance (one-way ANOVA) using the statistical package for social sciences (SPSS Inc., Chicago, USA) program version 21. Duncan’s multiple range test was employed to determine the significance of the difference within treatments for each analysis at probability level p < 0.05.
Results and Discussion
Antimicrobial Activity of L. lactis Gh1
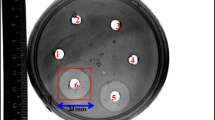
Pathogenic bacteria cause an important percentage of foodborne illnesses resulting from the consumption of contaminated food [29]. Among them, Escherichia coli and Listeria monocytogenes are responsible for a large number of outbreaks and deaths [30]. The CFS of L. lactis Gh1 showed an inhibitory effect (11.55 ± 0.14 mm diameter of clear zones) against L. monocytogenes ATCC 15313. The inhibition zones are shown in Online Resource 1. The antimicrobial activity of LAB may be attributed to the production of organic acids, with a consequent reduction in pH, hydrogen peroxide production or the production of bacteriocins or bacteriocin-like compounds [31]. In bacteria, the production of antimicrobial peptides provides a competitive advantage for the producer in certain ecological niches because of the peptide-mediated killing of other bacteria [32]. Antimicrobial peptides have been at the forefront of alternative antibiotic research for decades, but have not been extensively investigated for their utility. Bacteriocins, antimicrobial peptides of bacterial origin, are in particular positioned as potentially significant contributors to the paradigm shift due to the broad range of commercially available formats [33].
Physiological and Biochemical Characterization of L. lactis Gh1
In agreement with common characteristics of LAB, L. lactis Gh1 is a Gram-positive, catalase-negative and a diplococci (cocci in pair)-shaped bacterium. Catalase is a common enzyme presence in almost all oxygen-living organisms. It catalyses hydrogen peroxide decomposition into oxygen and water. Colonies were observed as smooth, cocci in shape, with convex elevation, entire margin, and cream in colour (shown in the Online Resources 2 and 3). The qualitative results for particular tests (carbon fermentation; antimicrobial susceptibility test; β-galactosidase activity; proteolytic activity and starch hydrolysis; determination of bile salts; hydrolases activity; acidifying activity) are shown in Online Resources 4.
Carbon Fermentation Test
It is found that L. lactis Gh1 was able to ferment both lactose and glucose as indicated by the formation of yellow zones surrounding the culture. Bromocresol purple added into the medium acted as a growth indicator. Positive results were indicated by the colour changes from purple to yellow caused by the production of lactic acid (pH below 5.2) as the production of this acid is growth-associated.
Effect of NaCl and Temperature on Growth of L. lactis Gh1
Results show that the bacterium was tolerant to 0.1 to 4.0% (w/v) NaCl. Additional data of the L. lactis Gh1 tolerance to different concentrations of NaCl and temperature are shown in Online Resource 5. Higher NaCl concentrations inhibit the growth of the bacterium. This finding is in accordance with Mulaw et al. [34], who reported that one of the LAB isolate namely E052 isolated from traditionally fermented Ethiopian food products was able to grow at 4% (w/v) NaCl and growth did not occur at 6.5% (w/v) NaCl. NaCl tolerance of the strains enables them to withstand the adverse effects of high osmotic pressure in the high-salt environment of the gastrointestinal tract and maintain a relative osmotic pressure balance under such conditions [35]. The NaCl tolerance test provides an indication of the osmotolerance degree of a LAB strain because bacterial cells grown in high salt concentrations will undergo a loss of turgor pressure that would affect the physiology, enzyme activity, water activity and cell metabolism [36]. Usually, bacteria protect themselves against the deleterious osmotic effects by the uptake or synthesis of a limited number of solutes under osmotic stress [37]. Furthermore, Lactococcus strains are homofermentative, and tolerance to pH, temperature and NaCl is species- and strain-dependent [38]. It is known that homofermentative LABs are more resistant to NaCl than heterofermentative LAB [39].
L. lactis Gh1 was able to grow at − 30 to 37 °C and growth was not detected at high temperatures (40–65 °C). In the food industry, the control of L. monocytogenes remains a challenge due to its ability to survive the most common food-processing conditions (extreme pH, high salt concentration, low water activity, and refrigeration temperatures) [40]. Therefore, the thermotolerance of L. lactis Gh1 at low temperatures (− 30 to 4 °C) might be advantageous to pawn this pathogen. The finding from this study is in good agreement with that of previous researchers, who stated that L. lactis usually grows at 30 °C [41] and 37 °C [42] as L. lactis is mesophilic in nature. At temperatures beyond 37 °C, L. lactis suffers from oxidative stress, which reduces NADH oxidase and pyruvate dehydrogenase activities [43].
Effect of Bile Salts and Low pH on Growth of L. lactis Gh1
The ability of L. lactis Gh1 to tolerate with bile salts and low pH up to 4 h of incubation is shown in Fig. 1. L. lactis Gh1 was resistant to 0.3% (w/v) of bile salt with comparable survival percentage (113–171%) to control culture (120–162%) throughout the incubation period. Furthermore, this strain was also able to tolerate the low pH condition (pH 3) and attained 96% of survivability after 4 h of incubation. Although there is a reduction in survivability as compared to control culture, this strain was still able to maintain its survivability consistently throughout the incubation period (96–101% survivability).

Tolerance of L. lactis Gh1 to bile salts and acidic condition. Results are expressed as mean and standard deviation; tests were performed in triplicate
Acid and bile salt tolerance is one of the essential requirements for probiotic strain selection as these are the few major factors that determine the likelihood of an exogenous culture surviving in the GIT [44]. The production of gastric acid by the stomach is an efficient mechanism for the destruction of pathogens ingested with food or water [45]. The resistance to 0.3–1.0% (w/v) of bile salt was also reported for various LAB species such as Lactococcus lactis, Lactobacillus acidophilus and Lactobacillus casei [46]. Bile salts interact with bacterial cell membranes, which are composed of lipids and fatty acids, inhibiting growth and killing many bacteria [47]. Bile salt tolerance is a prerequisite for bacterial colonization and metabolic activity in the host’s small intestine and contributes to the balance of intestinal microflora [48].
L. lactis Gh1 strain survived in the gastric juice with more than 95% of survivability after 4 h of incubation, suggesting that this bacterium has the potential for use as a probiotic. The secretion of gastric acid and transit through the stomach constitute a primary defence mechanism that all ingested microorganisms must overcome, including probiotics [49]. As probiotics are usually administrated orally, they must have the ability to survive passage through the stomach and small intestine. pH 3 and 4 h duration used in this study are to mimic the most common condition in the human stomach that assures the selection of the very acid-tolerant strain [50]. The transit time can be from < 1 to 3–4 h depending on the individual, the diet and other reigning conditions [51]. Acid tolerance of bacteria not only is necessary to withstand gastric stress but also helps the strain to live in high-acid foods such as yogurt for more extended periods without reducing the number of bacteria [52]. Corcoran et al. [53] reported that the resistance to low pH by Lactobacillus is due to the presence of F0F1-ATPase activity.
Effect of Phenol on the Growth of L. lactis Gh1
The ability of L. lactis Gh1 strain to tolerate different concentrations of phenol is shown in Table 1. After 24 h of incubation, L. lactis Gh1 showed a sufficient tolerance to phenol where there was no inhibition of viable cells as assured by the negative sign in the inhibition column at all phenol concentrations (0.1–0.4%, w/v). The viable cell count indicates the presence of growth since the initial viable cells were more than the final count. Even though the CFU per millilitre value was decreased with the increment of phenol concentration, it can be deduced that the growth inhibition has not occurred.
Phenol resistance of L. lactis Gh1 revealed its potential use in food applications. Xanthopoulos et al. [15] stated that the ability of strains to tolerate 0.4% (w/v) phenol indicates its capability to resist the possible bacteriostatic effect. This is because phenol has been reported to have a direct toxic effect on human colonic epithelial cells in vitro [54]. This metabolite affects the epithelial barrier function by disrupting the cellular membrane domains, thus indirectly altering the distribution of tight junction-associated proteins and can lead to gastrointestinal dysfunction or injury [55]. Furthermore, testing for resistance to phenol may further generate information for potential survivals of probiotic bacteria under digestive condition, because for a strain to be a probiotic, it has to survive the action of toxic metabolites, primarily phenols (a toxic metabolite produced by the deamination of some aromatic amino acids) [56]. In addition, phenol tolerance-probiotic bacteria living in the colon can bind or degrade phenol and consequently may reduce the risk of getting cancer [57].
Antimicrobial Susceptibility Test
The sensitivity of L. lactis Gh1 against 18 different types of antimicrobial agents by the disc diffusion method is presented in Table 2. Results showed that L. lactis Gh1 was resistant to 8 different types of antimicrobial agents namely sulphamethoxazole at 23–25 μg and compound sulphonamides from the sulphonamide group; trimethoprim from the diaminopyrimidine group; and all antibiotics from the aminoglycoside group (streptomycin, gentamycin) and rifampin. Furthermore, L. lactis Gh1 showed a susceptible effect to most of the antibiotics (10 types) specifically penicillin G, amoxicillin and ampicillin (β-lactam group); sulphafurazole and sulphonamide (sulphonamide group); vancomycin; erythromycin at 15 μg (macrolide group); tetracycline and oxytetracycline (tetracycline group); and chloramphenicol (amphenicol group). Besides, L. lactis Gh1 exhibited an intermediate result towards sulfamethoxazole at 100 μg from the sulphonamide group and erythromycin at 10 μg from the macrolide group.
Antimicrobial susceptibility screening of L. lactis Gh1 is an essential step to ensure its safe application to be incorporated in food products. In practice, antibiotics are used for opposing infectious diseases of humans and animals, but their efficacy has been resulting in an increase in antibiotic-resistant strains [58]. To date, the overwhelming increase in antibiotic resistance is presently recognized as a global crisis and, as such, requires the immediate attention of the pharmaceutical industry, academia and government institutions [59]. Therefore, by knowing the antimicrobial resistance properties of L. lactis Gh1 towards various antimicrobial agents, we can evaluate the security level of this strain before considering whether it is safe for human and animal consumption.
L. lactis Gh1 showed a susceptible effect towards β-lactams, vancomycin, macrolide, tetracyclines and amphenicol groups of antibiotics. The “susceptible” category implies that isolates are inhibited by the usually achievable concentrations of the antimicrobial agent when the dosage recommended treating the site of infection is used [17]. The susceptibility of L. lactis Gh1 to vancomycin is in line with the finding by Flórez et al. [60]. They also reported the resistance to vancomycin was not observed in 93 different L. lactis strains. Vancomycin has long been considered the antibiotic of last resort against serious and multi-drug-resistant infections caused by Gram-positive bacteria [61].
Also, the susceptibility of L. lactis Gh1 to tetracyclines is opposing the previous study by Flórez et al. [62] that reported six of their L. lactis strains were tetracycline resistant. Tetracycline is the corresponding resistance determinant and the most frequently described antibiotic-resistant in foodborne LAB. In comprehensive compilation work by Devirgiliis et al. [63], they found out that most of the resistance genes have been found in foodborne Lactococcus species is tet (M) gene carried by a functional Tn916 transposon,. Moreover, the majority of the tetracycline resistance genes found in L. lactis are located on resident plasmids, some of which have been proved to be transmissible by conjugation to lactococci and enterococci species [64].
L. lactis Gh1 strain is resistant to antimetabolite antibiotics, specifically sulphonamides, diaminopyrimidines, aminoglycoside, and rifampin groups. Antimetabolites antibiotics are antimicrobial agents that inhibit the synthesis of nucleotides needed for nucleic acid synthesis and folic acid metabolism [65]. The “resistant” category implies that isolates are not inhibited by the usually achievable concentrations of the agent with regular dosage schedules [17]. The resistance of the probiotic strains to some antibiotics might be an advantage considering a joint-use for probiotic therapy in the future for both preventive and therapeutic purposes in controlling intestinal infections [66]. The information provides a reasonable basis for the application of this probiotic along with certain antibiotics for improving the balance of the host GIT biota [67].
β-Galactosidase Activity
L. lactis Gh1 was not able to secrete β-galactosidase enzyme, as confirmed by the absence of a blue/green colony on MRS agar supplemented with X-gal and IPTG. The β-galactosidase is an enzyme involved in the hydrolysis of β-galactosides into monosaccharides through the breaking of a glycosidic bond [68]. The β-galactosidase-producing bacteria may also be considered as potential probiotics to reduce lactose intolerance as their ability to catalyse lactose digestion in the small intestine. Other than that, the strain can also be used in food industries to manufacture lactose-free products for lactose intolerant people and the formation of glycosylated products [69].
The deficiency of this enzyme in L. lactis Gh1 was concurred with the previous work by Vinderola and Reinheimer [46] in which the enzyme was also not found in L. lactis but detected in other strains such as Lb. delbrueckii subsp bulgaricus, Lb. acidophilus, S. thermophilus, B. bifidum and B. longum. According to Mustapha et al. [70], the absence of β-galactosidase would not be a disadvantage for probiotic strains since permeabilized cells might be necessary for efficient lactose hydrolysis to occur in the small intestine.
Qualitative Proteolytic Activity and Starch Hydrolysis
After 48 h of incubation, there was a slight clear zone observed surrounding the viable cells after smearing the iodine solution onto the surface of the media. This finding exposed that L. lactis Gh1 possessed visible but weak amylolytic activity and was not able to degrade significant amounts of starch. In general, amylolytic LAB played roles in food processing as they perform partial hydrolysis of the starchy material, making it easily digestible [71], and contribute to the flavour and taste of the fermented products [72].
Meanwhile, in proteolytic activity, there was no clear zone spotted adjacent to the cells, indicating the incapability of the bacterium to degrade protein. However, it has been reported that Lactobacillus strains with low proteolytic activity are successfully applied as adjunct cultures in different types of cheeses [73]. They contribute in the secondary proteolysis via peptidolytic potential, increasing the number of small peptides and free amino acids, major precursors of specific flavour compounds. The proteolytic enzymes degrade casein by releasing low molecular weight peptides and amino acids, which have been identified as particular growth stimulants to the enzyme producer [74].
Qualitative Determination of Bile Salt Hydrolase Activity
The opaque white with shine (no halos) colonies were observed on the MRS agar plate supplemented with 0.5% (w/v) sodium taurodeoxycholate hydrate (TDCA), both under aerobic and anaerobic conditions indicating the strain was able to deconjugate TDCA with and without the presence of atmospheric oxygen. BSH is an enzyme that catalyses the deconjugation of bile salt to liberate free primary bile acids [48]. The ability of probiotic strains to detoxify bile salt by producing BSH enzyme activity has often been included among the criteria for probiotic strain selection as the presence of BSH in probiotics renders them more tolerant to bile salts, which also helps to reduce the blood serum cholesterol level of the host [75]. In the meantime, the deconjugation of bile salts by BSH also can protect the gut microbiota and probiotic bacteria from the toxicity of conjugated bile salts and may have significant impacts on the physiology of the host [76]. Hence, the required culture that can grow well in the presence of bile, deconjugate bile salts, and assimilate cholesterol would be more useful and well-functioning in the GIT [77].
Acidifying Activity
L. lactis Gh1 was able to acidify skim milk as the pH of the skim milk reduced from pH 6.34 to 4.74 after 96 h of fermentation (Table 3). The coagulation of skim milk started at 24 h and was completed at 96 h. The increase in skim milk viscosity is a characteristic symptom of its thickening and takes place in the particulate protein available [78]. Milk acidification directly affects the stability of casein micelles, reduces the charges, dissolves some insoluble crosslinks of calcium phosphate and modifies the internal bonding between proteins [79].
Generally, the acidification capability of potential strains is one of the most significant technological features of possible starter cultures, particularly for the manufacture of milk-based probiotic products. The reduction of pH can control the growth of a large number of pathogenic or undesirable microorganisms that cause fermented products to spoil and can improve the hygienic properties and storage of the final products [80].
Haemolytic Activity of L. lactis Gh1
L. lactis Gh1 exhibited γ-haemolytic activity (no haemolysis), which proves the non-pathogenic status of the probiotic. There is no rupture or destruction of red blood cells in the agar plate. Haemolytic activity is commonly used to differentiate suspected pathogens because there may be a relationship between the haemolytic activity and pathogenicity of strains [81]. The general absence of haemolysis or poor haemolytic activities expressed by LAB is indicative of their safety applications in food. In vitro assessment of haemolytic activity on blood agar is one of the safety requirements often used to assess potential probiotic strains [82]. During the introduction of new microorganisms (genera and strains) for novel products, the safety and risk-to-benefit ratio should be evaluated based on CODEX Alimentarius guidelines to perform the premarketing determination of product safety [83]. Microorganisms can be classified into three groups which are as follows: non-pathogenic (Lactobacillus, Lactococcus, Bifidobacterium, Saccharomyces), pathogenic (B. cereus) and opportunistic pathogens (Enterococcus and also other general LAB) [84].
Enzymatic Profile of L. lactis Gh1
Enzymes produced by L. lactis Gh1 using API-ZYM system are listed in Table 4. It was found that digestive enzymes such as acid phosphatase and naphthol-AS-BI-phosphohydrolase were abundantly produced, whereas alkaline phosphatase and leucine arylamides at a moderate amount, and small amounts, were recorded for esterase (C4), esterase lipase (C8) and lipase (C14). Although the API ZYM system does only a semiquantitative analysis, these results are relevant and allow the selection of LAB with useful technological and probiotic characteristics. In the human digestive system, acid phosphatase and naphthol-AS-BI-phosphohydrolase are necessary enzymes used to free attached phosphoryl groups from other molecules during digestion [85]. Alkaline phosphatase is a superfamily of metalloenzymes known to catalyse the hydrolytic removal of phosphate from a variety of molecules [86]. The role of intestinal alkaline phosphatase (IAP) is key regulator of inflammation, infection and gut microbiota. Diminished activity of IAP could increase the risk of disease through changes in the microbiome, increased intestinal inflammation, and intestinal permeability [87]. Furthermore, leucine-arylamidase is an enzyme that breaks down the amino acid leucine into acetyl acetate, and acetyl coA.
In this study, the absence of a few essential enzymes like α-glucosidase and β-glucosidase was noticed. Both enzymes’ activity could contribute to polysaccharide digestion [88], and the presence of glycosidase activities in food cultures seems to affect sensory properties as it is often linked to sugar metabolism [89]. Other than that, the deficiency of β-galactosidase in this experiment was also confirmed by β-galactosidase activity result (absence of blue/green colonies). This enzyme is a beneficial enzyme considering both probiotic and technological aspects, supporting the reduction of lactose intolerance as well as milk [90]. However, the absence of β-glucuronidase activity could be beneficial as the enzyme may have adverse effects in the colon and has been considered a carcinogenic enzyme [90]. Also, the absence of β-glucuronidase is in good agreement with the data from Leite et al. [91] and Aziz et al. [92].
Physio-chemical Stability of BLIS Activity
One of the drawbacks of the direct application of bacteriocins to a food matrix is that the activity loss often occurs over time. This is because the enzymatic degradation and the interaction with food components such as proteins, lipids [93] and curing ingredients (sodium chloride, sodium nitrite, ascorbic acid, alginate and sodium lactate) also can be influenced by the environmental conditions of storage, food components, solubility and food additives [94] and the hurdle of food processing [95]. Besides bacteriocins, LAB also can produce a wide range of inhibitory compounds to reduce pathogen invasion, which includes reutericyclin, reuterin, organic acids (lactic acid, acetic acid), ethanol, diacetyl, carbon dioxide and hydrogen peroxide [96]. Therefore, an investigation of CFS containing BLIS is essential and needs to be carried out. Applying a crude bacterial fermentate rather than a purified bacteriocin may offer the advantage of incorporating different biologically active substances with possible synergistic effects in one product.
Effect of pH on BLIS Activity
The stability of BLIS at pH ranging from 2 to 11 is shown in Fig. 2. It is found that the supernatant from L. lactis Gh1 showed strong resistance in a broad range of pH (pH 4–8). In comparison to control (untreated), both acidic and alkaline conditions gave only a slight reduction in the activity (up to 9.73%). The optimal pH range for BLIS of L. lactis Gh1 also concurred with that reported by Moreno et al. [97], who claimed that BLIS was active at pH ranged from pH 2 to 10 but completely inactivated at pH 12. The stability of CFS from L. lactis Gh1 at neutralized pH indicated that the antimicrobial activity was not due to the acidity of lactic acid or any organic acid presence in the CFS. Likewise, Lim et al. [98] also found that the neutralized CFS showed growth inhibition against P. gingivalis, and concluded that it was due to the presence of the antimicrobial substance, such as BLIS.

Stability of bacteriocin-like inhibitory substance (BLIS) activity from L. lactis Gh1 at different pH levels. The indicated values are the reduction percentages of the activity after pH treatment
In the meantime, a reduction of BLIS activity up to 54.38–56.79% was observed at pH lower than pH 4 and higher than pH 8. Similar observations have also been reported for several other bacteriocins, such as paracaseicin A [99] and antimicrobial compound produced by L. lactis [97]. Reduced activity at high pH was possibly due to the denaturation of the protein [14]. In general, BLIS possesses stability at a wide range of pH and has advantages in food application in which it can be applied in both acidic and alkaline food manufacturing. Woraprayote et al. [28] stated that the suitable utilization of certain bacteriocins could be decided based on their pH stability profile. Moreover, pH stability of bacteriocins would be beneficial for its application as a food preservative [100] as it is capable of combating against moulds, yeast, acetobacter and other deteriorating food microorganisms in neutral and acidic foods.
Effect of Enzymes on BLIS Activity
The effect of enzymes on the activity of BLIS from L. lactis Gh1 at different culture media is presented in Table 5. The BLIS activity of L. lactis Gh1 was reduced by proteolytic enzymes, proteinase K in both culture media, MRS and BHI up to 40.43% and 58.25%, respectively. This test was done to determine the inhibition showed by L. lactis Gh1 that was exclusively due to the action of BLIS instead of lactic acid since bacteriocins by definition are proteinaceous substances and hence they must be sensitive to at least one of the proteolytic enzymes. Thus, bacteriocins can be identified in part by exposure to proteolytic enzymes [101, 102]. Besides, the inactivation of bacteriocin activity by proteolytic enzymes also ensured the safety for the consumer since these bacteriocins could be entirely deactivated by enzymes in human GIT and would then be inactive against beneficial bacteria in human intestine [28].
It is also found that other protease treatment with trypsin did not alter much the antimicrobial activity of BLIS in this study; however, proteolytic enzymes do not always reduce the antimicrobial activity of a bacteriocin [103]. Stability in the presence of a proteolytic enzyme could be due to unusual amino acids in the active site bacteriocin structure [104]. Other than that, BLIS activity was not reduced by catalase, indicating that hydrogen peroxide was not responsible for microbial inhibition. A parallel finding was observed by Mariam et al. [105] where treatment with catalase did not result in the abolition of activity, signifying that hydrogen peroxide was not responsible for the observed actions.
Heating, Storage Temperature, Freezing and Thawing Sensitivity of BLIS
The stability of antimicrobial activity of crude supernatant on heat treatment is shown in Table 6. Results show that metabolites produced by L. lactis Gh1 demonstrated a low thermal resistance. The activity decreased upon the exposure to elevated temperature at 60–90 °C up to 78% of decrement and was completely abolished at 100 °C and 121 °C for all heating periods tested so far. On the contrary, Ponce et al. [106] found that L. lactis isolated from organic leafy vegetable showed a durable resistance to heat treatment, retaining about 60% of antibacterial activity after 10 and 20 min at 60 °C and 80 °C. The heat stability may be due to the formation of small globular structures and the occurrence of strongly hydrophobic regions, stable cross-linkages and high glycine content [27]. In general, heat stability would be an advantageous characteristic for bacteriocin to be used as a food preservative, as many food-processing procedures involve a heating step [28].
The type of culture medium often has significant influences on the stability of CFS during the storage period. As tabulated in Table 7, the use of BHI medium in the cultivation of L. lactis Gh1 showed high stability indicated by lower and steady reduction of percentages in all storage temperatures throughout the study (30 °C: up to 120 days—68.41%; 4 °C: up to 180 days—68.19%) as compared to MRS medium (30 °C: up to 30 days—74.68%; 4 °C: up to 120 days—61.72%) (except at − 20 °C). Meanwhile, 4 °C was found to be the most suitable storage temperature for both media. The remarkable effect of 4 °C in maintaining BLIS activity in this study was in agreement with the finding reported by Pawar et al. [107]. They also noticed that there was a less pronounced inhibition effect of nisin when the storage temperature was increased to 37 °C as compared to 4 °C. Besides, Prudêncio et al. [108] stated that a reduction in temperature promotes a change in the structure of the outer membrane, and this alteration may permeabilize the cell, allowing the nisin to act on S. typhimurium and E. coli at refrigeration temperatures. Moreover, this study was also meant to access the stability of BLIS from L. lactis Gh1 at 4 °C and − 20 °C since L. monocytogenes has shown to be able to survive at low temperature even after several cycles of freezing and thawing [109].
The sensitivity of antimicrobial activity from CFS of L. lactis Gh1 towards a repetitive freezing and thawing process on two culture media, namely BHI and MRS, is presented in Table 8. The result shows that CFS obtained from the BHI broth-grown strain displayed a steady reduction on each cycle and recorded 59.36 ± 1.34% at cycle 4. Whereas in the MRS medium, the activity was drastically affected at cycle 2 with 51.65 ± 3.97% activity reduced and further diminished up to 66.10 ± 3.20% at cycle 4. The stability pattern of antimicrobial activity for the first 3 cycles (equivalent to 3 days) before being significantly reduced at cycle 4 in BHI medium was similar to the previous observation in storage study at − 20 °C. In contrast, in this experiment, the CFS from MRS medium exhibits a sensitivity to the freezing-thawing process throughout the cycles. MRS medium might have heat-sensitive ingredients/end-fermentation by-product that may cause BLIS to experience cold denaturation during storage. The morphology of bacteriocins can be changed by freezing and thawing, which may render these particles to be inactive [110]. The incorporation of nisin with stress caused by the freezing/thawing regime showed a much higher reduction in viability of tested bacteria due to transient injury to the outer membrane permeability barrier, allowing nisin temporary access to the cytoplasmic membrane [111]. The fact that CFS demonstrated freezing and thawing stability has technological importance as the demand for frozen foods is increasing. It may also broaden the potential applications of such compounds as biopreservatives especially for minimally processed vegetables [106].
Stability of BLIS to Different Organic Solvents and Chemicals
The effects of organic solvents and chemicals on BLIS activity are shown in Table 9. The antimicrobial activity of BLIS was slightly affected by organic solvents such as acetone (24.58 ± 0.03%), ethanol (23.57 ± 0.06%), methanol (22.00 ± 0.08%) and acetonitrile (23.94 ± 0.02%) at 10% (v/v) concentration. However, a complete denaturation was recorded at 50% (v/v), excluding butanol and toluene, which was extremely affected by both concentrations. On the contrary, BLIS activity was moderately affected at both concentrations of isopropyl alcohol (10%, v/v: 43.33 ± 0.03%; 50%, v/v: 58.27 ± 0.05% reduction).
The stability of CFS from L. lactis Gh1 was in line with the finding reported by Barman et al. [112]. They stated that L. lactis strains retained the activities in a mixture with 10% (v/v) organic solvents viz., acetone, benzene, chloroform, dimethyl sulfoxide (DMSO), ethanol, ethyl acetate and methanol. The inactivation of CFS activity might be due to the degradation, which is unfolding of the three-dimensional structure of the protein, and commonly leads to a loss of functionality and formation of potentially immunogenic products. This can be caused by various stresses such as heat, shear stress, exposure to interfaces or chemical factors [113]. Moreover, in the food industry, the application of bacteriocins in the film matrix for antimicrobial packaging required the bacteriocins to be well-suited with film polymers and/or needed to be soluble in the same solvent of the film [11].
Furthermore, the BLIS antimicrobial activity from L. lactis Gh1 was found to be tolerant towards Tween 80, urea and NaCl with percentages of reduction of only 27.38 ± 0.04, 7.74 ± 0.09 and 14.08 ± 0.07%, respectively. On the other hand, SDS, EDTA and Tween 20 exhibited a damaging effect on BLIS activity. In contrast, the bacteriocin produced by Weissella hellenica BCC 7293 was greatly affected by urea, indicating that the sensitivity towards surfactants and urea seems to be bacteriocin-dependent [28]. Meanwhile, the stability of BLIS on NaCl is important as BLIS is often formulated with NaCl in food processing. Pawar et al. [107] concluded that the addition of 2% (w/v) NaCl in combination with nisin increased the efficacy of nisin in controlling L. monocytogenes in minced raw buffalo meat at storage temperatures of 4 and 37 °C. EDTA is one of the food preservatives that may act in destabilizing the outer membrane, by the release of components of the structure or by intercalating in the layer, permitting the bacteriocin action [114]. In this study, EDTA has caused a massive reduction (90.85 ± 0.29%) of the antimicrobial activity of CFS from L. lactis Gh1.
In contrast, an increase in the activity of BLIS was observed when it was in combination with Triton X-100. Several studies have indicated that the non-ionic detergent, Triton X-100, has a specific solubilizing effect on various entities with the hydrophobic surfaces including membrane proteins, carbon nanotube, influenza vaccine, liposomes and native membranes. Triton X-100 solubilizes a portion of proteins that have affinity for rafts, especially transmembrane proteins and proteins of the inner leaflet [115]. This might also be a reason for the elevation of BLIS antimicrobial activity. Even though the crude supernatant was used in this study, the possibility of existence of cell debris due to improper settlement of pellets is possible.
Conclusions
Results from this study suggest that L. lactis Gh1 and its crude supernatant have the potential to be used in commercial food applications. This strain exhibited desirable probiotic characteristics such as tolerance to low pH, bile salt, phenol and a wide range of NaCl and susceptibility to a wide range of antibiotics. The strain also can produce essential enzymes and can coagulate milk. Whereas, the BLIS secreted by this bacterium has stability at 4 °C for an extended period of storage and can withstand the repetitive freezing-thawing cycles. Other than that, the BLIS activity was not affected by a wide range of pH values (pH 4–8), and was slightly reduced by organic solvents such as acetone, ethanol, methanol and acetonitrile at 10% (v/v), and chemicals such as Tween 80, urea and NaCl. Further research is required to thoroughly explore the possible application of this strain as a bioprotective starter or co-starter culture for its use in the development of safe and nutritious food for human consumption. These investigations include in vivo field tests to investigate the capacity to adhere to the intestinal mucosa, the antimicrobial activities with synergic effects against pathogenic bacteria, identification of the metabolites, exploring their mechanisms of action, suitable modes and potential applications as well as safety issues.
References
Fazilah NF, Ariff AB, Khayat ME, Rios-Solis L, Halim M (2018) Influence of probiotics, prebiotics, synbiotics and bioactive phytochemicals on the formulation of functional yogurt. J Funct Foods 48:387–399. https://doi.org/10.1016/j.jff.2018.07.039
Ayeni FA, Sánchez B, Adeniyi BA, de Los Reyes-Gavilán CG, Margolles A, Ruas-Madiedo P (2011) Evaluation of the functional potential of Weissella and Lactobacillus isolates obtained from Nigerian traditional fermented foods and cow’s intestine. Int J Food Microbiol 147:97–104. https://doi.org/10.1016/j.ijfoodmicro.2011.03.014
Zielińska D, Rzepkowska A, Radawska A, Zieliński K (2015) In vitro screening of selected probiotic properties of Lactobacillus strains isolated from traditional fermented cabbage and cucumber. Curr Microbiol 70(2):183–194. https://doi.org/10.1007/s00284-014-0699-0
Colombo M, Castilho NPA, Todorov SD, Nero LA (2018) Beneficial properties of lactic acid bacteria naturally present in dairy production. BMC Microbiol 18:219. https://doi.org/10.1186/s12866-018-1356-8
FDA (2010) Generally recognized as safe (GRAS) notifications. https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras. Accessed 12 July 2020
Reis JA, Paula AT, Casarotti SN, Penna ALB (2012) Lactic acid bacteria antimicrobial compounds: characteristics and application. Food Eng Rev 4:124–140. https://doi.org/10.1007/s12393-012-9051-2
Alvarez-Sieiro P, Montalbán-López M, Mu D, Kuipers OP (2016) Bacteriocins of lactic acid bacteria: extending the family. Appl Microbiol Biotechnol 100:2939–2951. https://doi.org/10.1007/s00253-016-7343-9
Silva C, Silva S, Ribeiro SC (2018) Application of bacteriocins and protective cultures in dairy food preservation. Front Microbiol 9(594):1–15. https://doi.org/10.3389/fmicb.2018.00594
EFSA (2007) Scientific committee. Introduction of a qualified presumption of safety (QPS) approach for assessment of selected microorganisms referred to EFSA1. EFSA J 587:1–16
Sidek NLM, Halim M, Tan JS, Abbasiliasi S, Mustafa S, Ariff AB (2018) Stability of bacteriocin-like inhibitory substance (BLIS) produced by Pediococcus acidilactici Kp10 at different extreme conditions. Biomed Res Int 2018:1–11. https://doi.org/10.1155/2018/5973484
Woraprayote W, Malila Y, Sorapukdee S, Swetwiwathana A, Benjakul S, Visessanguan W (2016) Bacteriocins from lactic acid bacteria and their applications in meat and meat products. Meat Sci 120:118–132. https://doi.org/10.1016/j.meatsci.2016.04.004
Cleveland J, Montville TJ, Nes IF, Chikindas ML (2001) Bacteriocins: safe, natural antimicrobials for food preservation. Int J Food Microbiol 71:1–20. https://doi.org/10.1016/S0168-1605(01)00560-8
Abbasiliasi S, Ramanan RN, Tengku Azmi TI, Shuhaimi M, Mohammad R, Ariff AB (2010) Partial characterization of antimicrobial compound produced by Lactobacillus paracasei LA07, a strain isolated from Budu. Minerva Biotecnologica 22
Abbasiliasi S, Tan JS, Ibrahim TAT, Ramanan RN, Vakhshiteh F, Mustafa S, Ling TC, Rahim RA, Ariff A (2012) Isolation of Pediococcus acidilactici Kp10 with ability to secrete bacteriocin-like inhibitory substance from milk products for applications in food industry. BMC Microbiol 12:1–12. https://doi.org/10.1186/1471-2180-12-260
Xanthopoulos V, Litopoulou-Tzanetaki E, Tzanetakis N (2000) Characterization of Lactobacillus isolates from infant faeces as dietary adjuncts. Food Microbiol 17(2):205–215. https://doi.org/10.1006/fmic.1999.0300
Bauer AW, Kirby WM, Sherris JC, Turck M (1966) Antibiotic susceptibility testing by standardized single disc method. Am J Clinl Pathol 45:493–496. https://doi.org/10.1093/ajcp/45.4ts.493
CLSI (2014) Performance standards for antimicrobial susceptibility testing. 24th informational supplement. Clinical and Laboratory Standards Institute, Wayne, PA
Liasi S, Azmi T, Hassan M, Shuhaimi M, Rosfarizan M, Ariff AB (2009) Antimicrobial activity and antibiotic sensitivity of three isolates of lactic acid bacteria from fermented fish product, Budu. Malaysian J Microbiol 5:33–37. https://doi.org/10.21161/mjm.15008
Vlková E, Rada V, Popelářová P, Trojanová I, Killer J (2006) Antimicrobial susceptibility of Bifidobacteria isolated from gastrointestinal tract of calves. Livestock Sci 105:253–259. https://doi.org/10.1016/j.livsci.2006.04.011
Karasova P, Spiwok V, Mala S, Kralova B, Russell NJ (2002) Beta-galactosidase activity in psychrotrophic microorganisms and their potential use in food industry. Czech J Food Sci 20:43–47. https://doi.org/10.17221/3508-CJFS
Pailin T, Kang DH, Schmidt K, Fung DYC (2001) Detection of extracellular bound proteinase in EPS-producing lactic acid bacteria cultures on skim milk agar. Lett Appl Microbiol 33:45–49. https://doi.org/10.1046/j.1472-765X.2001.00954.x
Sharma P, Sharma P, Kumar N, Suman DN (2017) Identification and characterization of bile salt hydrolyzing Lactobacillus isolates. Int J Curr Microbiol App Sci 6(3):1655–1675. https://doi.org/10.20546/ijcmas.2017.603.192
Chettri R, Tamang JP (2014) Functional properties of Tungrymbai and Bekang, naturally fermented soybean foods of North East India. Int J Ferment Foods 3:87–103. https://doi.org/10.5958/2321-712X.2014.01311.8
Abbasiliasi S, Tan J, Bashokouh F, Azmi T, Ibrahim T, Mustafa S, Vakhshiteh F, Sivasamboo S, Ariff A (2017) In vitro assessment of Pediococcus acidilactici Kp10 for its potential use in the food industry. BMC Microbiol 17:1–11. https://doi.org/10.1186/s12866-017-1000-z
Tagg JR, Dajani AS, Wannamaker LW (1976) Bacteriocins of Gram-positive bacteria. Bacteriol Rev 40(3):722–756. https://doi.org/10.1128/MMBR.40.3.722-756.1976
Parente E, Brienza C, Moles M, Ricciardi A (1995) A comparison of methods for the measurement of bacteriocin activity. J Microbiol Methods 22:95–108. https://doi.org/10.1016/0167-7012(94)00068-I
De Vuyst L, Vandamme EJ (1994) Bacteriocins of lactic acid bacteria: microbiology, genetics and applications. Blackie Academic & Professional, London
Woraprayote W, Pumpuang L, Tosukhowong A, Roytrakul S, Perez R, Zendo T, Sonomoto K, Benjakul S, Visessanguan W (2015) Two putatively novel bacteriocins active against Gram-negative food borne pathogens produced by Weissella hellenica BCC 7293. Food Control 55:176–184. https://doi.org/10.1016/j.foodcont.2015.02.036
Alegbeleye OO, Singleton I, Sant’Ana AS (2018) Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: a review. Food Microbiol 73(177–208):177–208. https://doi.org/10.1016/j.fm.2018.01.003
WHO (2015) Food safety. http://www.who.int/news-room/fact-sheets/detail/food-safety
Nishant T, Sathish Kumar D, Arun Kumar R, Hima Bindu K, Raviteja Y (2011) Bacteriocin producing probiotic lactic acid bacteria. J Microbial Biochem Technol 3:121–124. https://doi.org/10.4172/1948-5948.1000062
Hassan M, Kjos M, Nes IF, Diep DB, Lotfipour F (2012) Natural antimicrobial peptides from bacteria: characteristics and potential applications to fight against antibiotic resistance. J Appl Microbiol 113:723–736. https://doi.org/10.1111/j.1365-2672.2012.05338.x
Cavera VL, Arthur TD, Kashtanov D, Chikindas ML (2015) Bacteriocins and their position in the next wave of conventional antibiotics. Int J Antimicrob Agents 46(5):494–501. https://doi.org/10.1016/j.ijantimicag.2015.07.011
Mulaw G, Tessema TS, Muleta D, Tesfaye A (2019) In vitro evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented Ethiopian food products. Int J Microbiol 2019:1–12. https://doi.org/10.1155/2019/7179514
Xu Y, Tian Y, Cao Y, Li J, Guo H, Su Y, Tian Y, Wang C, Wang T, Zhang L (2019) Probiotic properties of Lactobacillus paracasei subsp. paracasei L1 and its growth performance-promotion in chicken by improving the intestinal microflora. Front Physiol 10:1–14. https://doi.org/10.3389/fphys.2019.00937
Ibourahema C, Dauphin RD, Jacqueline D, Thonart P (2008) Characterization of lactic acid bacteria isolated from poultry farms in Senegal. Afr J Biotechnol 7:2006–2012. https://doi.org/10.5897/AJB2008.000-5048
Bremer E, Kramer R (2000) Coping with osmotic challenges: osmoregulation through accumulation and release of compatible solutes. In: Storz G, Hengge-Aronis R (eds) Bacterial stress responses, vol 126. ASM Press, Washington, pp 79–97. https://doi.org/10.1016/S1095-6433(00)80031-8
Papagianni M, Anastasiadou S (2009) Pediocins: the bacteriocins of Pediococci. Sources, production, properties and applications. Microb Cell Factories 8:1–16. https://doi.org/10.1186/1475-2859-8-3
Simonson L, Hannu Salovaara H, Korhola M (2003) Response of wheat sourdough parameters to temperature, NaCl and sucrose variations. Food Microbiol 20:193–199. https://doi.org/10.1016/S0740-0020(02)00117-X
Law JWF, Ab Mutalib NS, Chan KG, Lee LH (2015) An insight into the isolation, enumeration, and molecular detection of Listeria monocytogenes in food. Front Microbiol 6(1227):1–15. https://doi.org/10.3389/fmicb.2015.01227
Chen J, Shen J, Hellgren LI, Jensen PR, Solem C (2015) Adaptation of Lactococcus lactis to high growth temperature leads to a dramatic increase in acidification rate. Sci Rep 5:1–15. https://doi.org/10.1002/jps.21825
Ahmed T, Kanwal R, Najma Ayub N (2006) Influence of temperature on growth pattern of Lactococcus lactis, Streptococcus cremoris and Lactobacillus acidophilus isolated from camel milk. Biotechnol Adv 5:481–488. https://doi.org/10.3923/biotech.2006.481.488
Chen J, Jing Shen J, Solem C, Jensen PR (2013) Oxidative stress at high temperatures in Lactococcus lactis due to an insufficient supply of riboflavin. Appl Environ Microbiol 79(19):6140–6147. https://doi.org/10.1128/AEM.01953-13
Halim M, Mohd Mustafa NA, Othman M, Wasoh H, Kapri MR, Ariff AB (2017) Effect of encapsulant and cryoprotectant on the viability of probiotic Pediococcus acidilactici ATCC 8042 during freeze-drying and exposure to high acidity, bile salts and heat. LWT - Food Sci Technol 81:210–216. https://doi.org/10.1016/j.lwt.2017.04.009
Beasley DE, Koltz AM, Lambert JE, Fierer N, Dunn RR (2015) The evolution of stomach acidity and its relevance to the human microbiome. PLoS One 10(7):1–12. https://doi.org/10.1371/journal.pone.0134116
Vinderola CG, Reinheimer JA (2003) Lactic acid starter and probiotic bacteria: a comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Res Int 36:895–904. https://doi.org/10.1016/S0963-9969(03)00098-X
Erkkilä S, Petäjä E (2000) Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci 55:297–300. https://doi.org/10.1016/S0309-1740(99)00156-4
Shehata MG, El Sohaimy SA, El-Sahn MA, Youssef MM (2016) Screening of isolated potential probiotic lactic acid bacteria for cholesterol lowering property and bile salt hydrolase activity. Ann Agric Sci 61(1):65–75. https://doi.org/10.1016/j.aoas.2016.03.001
Gueimonde M, Salminen S (2006) New methods for selecting and evaluating probiotics. Dig Liver Dis 38(2):S242–S247. https://doi.org/10.1016/S1590-8658(07)60003-6
Pennacchia C, Ercolini D, Blaiotta G, Pepe O, Mauriello G, Villani F (2004) Selection of Lactobacillus strains from fermented sausages for their potential use as probiotics. Meat Sci 67(2):309–317. https://doi.org/10.1016/j.meatsci.2003.11.003
Fazilah NF, Hamidon NH, Ariff AB, Khayat ME, Wasoh H, Halim M (2019) Microencapsulation of Lactococcus lactis Gh1 with gum Arabic and Synsepalum dulcificum via spray drying for potential inclusion in functional yogurt. Molecules 24:1–21. https://doi.org/10.3390/molecules24071422
Ammor MS, Mayo B (2007) Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: an update. Meat Sci 76(1):138–146. https://doi.org/10.1016/j.meatsci.2006.10.022
Corcoran BM, Stanton C, Fitzgerald GF, Ross RP (2005) Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl Environ Microbiol 71:3060–3067. https://doi.org/10.1128/AEM.71.6.3060-3067.2005
Pedersen GBJ, Saermark T (2002) Phenol toxicity and conjugation in human colonic epithelial cells. Scand J Gastroenterol 37:74–79. https://doi.org/10.1080/003655202753387392
McCall IC, Betanzos A, Weber DA, Nava P, Miller GW, Parkos CA (2009) Effects of phenol on barrier function of a human intestinal epithelial cell line correlate with altered tight junction protein localization. Toxicol Appl Pharmacol 241(1):61–70. https://doi.org/10.1016/j.taap.2009.08.002
Sathyabama S, Vijayabharathi R, Priyadarisini VB (2012) Screening for probiotic properties of strains isolated from feces of various human groups. J Microbiol 50:603–612. https://doi.org/10.1007/s12275-012-2045-1
Nowak A, Libudzisz Z (2007) Ability of intestinal lactic bacteria to bind or/and metabolise phenol and p-cresol. Ann Microbiol 57(3):329–335. https://doi.org/10.1007/BF03175068
Lawton EM, Ross RP, Hill C, Cotter PD (2007) Two-peptide lantibiotics: a medical perspective. Mini Rev Med Chem 7:1236–1247. https://doi.org/10.2174/138955707782795638
Rather IA, Kim BC, Bajpai V (2017) Self-medication and antibiotic resistance: crisis, current challenges, and prevention. Saudi J Biol Sci 24(4):808–812. https://doi.org/10.1016/j.sjbs.2017.01.004
Flórez AB, Danielsen M, Korhonen J, Zycka J, von Wright A, Bardowski J, Mayo B (2007) Antibiotic survey of Lactococcus lactis strains to six antibiotics by Etest, and establishment of new susceptibility-resistance cut-off values. J Dairy Res 74(3):262–268. https://doi.org/10.1017/S0022029907002543
Boneca IG, Chiosis G (2003) Vancomycin resistance: occurrence, mechanisms and strategies to combat it. Expert Opin Ther Targets 7(3):311–328. https://doi.org/10.1517/14728222.7.3.311
Flórez AB, Ammor MS, Mayo B (2008) Identification of tet(M) in two Lactococcus lactis strains isolated from a Spanish traditional starter-free cheese made of raw milk and conjugative transfer of tetracycline resistance to lactococci and enterococci. Int J Food Microbiol 121(2):189–194. https://doi.org/10.1016/j.ijfoodmicro.2007.11.029
Devirgiliis C, Zinno P, Perozzi G (2013) Update on antibiotic resistance in foodborne Lactobacillus and Lactococcus species. Front Microbiol 4:1–13. https://doi.org/10.3389/fmicb.2013.00301
Lampkowska J, Feld L, Monaghan A, Toomey N, Schjørring S, Jacobsen B, van der Voet H, Andersen SR, Bolton D, Aarts H, Krogfelt KA, Wilcks A, Bardowski J (2008) A standardized conjugation protocol to assess antibiotic resistance transfer between lactococcal species. Int J Food Microbiol 127:172–175. https://doi.org/10.1016/j.ijfoodmicro.2008.06.017
Bhattacharjee MK (2016) Antimetabolites: antibiotics that inhibit nucleotide synthesis. In: Chemistry of antibiotics and related drugs. Springer International Publishing, Switzerland, pp 95–108. https://doi.org/10.1007/978-3-319-40746-3_4
EI-Naggar MY (2004) Comparative study of probiotic cultures to control the growth of E. coli O157:H7 and Salmonella typhimurium. Biotechnol 3 (2):173–180. doi:https://doi.org/10.3923/biotech.2004.173.180
Huang R, Tao X, Wan C, Li S, Xu H, F X, Shah NP, Wei H (2015) In vitro probiotic characteristics of Lactobacillus plantarum ZDY 2013 and its modulatory effect on gut microbiota of mice. J Dairy Sci 98:5850–5861. doi:https://doi.org/10.3168/jds.2014-9153
Ghosh SK, Pandey A, Arora S, Dwivedi VD (2013) Comparative modelling and docking studies of β-galactosidase from Aspergillus niger. Netw Model Anal Health Inform Bioinforma 2:297–302. https://doi.org/10.1007/s13721-013-0046-6
Saqib S, Akram A, Halim SA, Tassaduq R (2017) Sources of β-galactosidase and its applications in food industry. 3 Biotech 7:79. doi:https://doi.org/10.1007/s13205-017-0645-5
Mustapha A, Jiang T, Savaiano DA (1997) Improvement of lactose digestion by humans following ingestion of unfermented acidophilus milk: influence of bile sensitivity, lactose transport, and acid tolerance of Lactobacillus acidophilus. J Dairy Sci 80(8):1537–1545. https://doi.org/10.3168/jds.S0022-0302(97)76083-1
Nguyen TTT, Loiseau G, Icard-Verniere C, Rochette I, Treche S, Guyot JP (2007) Effect of fermentation by amylolytic lactic acid bacteria, in process combinations, on characteristics of rice/soybean slurries: a new method for preparing high energy density complementary foods for young children. Food Chem 100:623–631. https://doi.org/10.1016/j.foodchem.2005.09.080
Blandino A, Al-Aseeri ME, Pandiella SS, Canter OD, Webb C (2003) Cereal-based fermented foods and beverages. Food Res Int 36:527–543. https://doi.org/10.1016/S0963-9969(03)00009-7
Briggiler-Marco M, Capra M, Quiberoni A, Vinderola G, Reinheimer J, Hynes E (2007) Nonstarter Lactobacillus strains as adjunct cultures for cheese making: in vitro characterization and performance in two model cheeses. J Dairy Sci 90:4532–4542. https://doi.org/10.3168/jds.2007-0180
Kivanç M, Yilmaz M, Cakir E (2011) Isolation and identification of lactic acid bacteria from boza, and their microbial activity against several reporter strains. Turk J Biol 35:313–324. https://doi.org/10.3906/biy-0906-67
Miremadi F, Ayyash M, Sherkat F, Stojanovska L (2014) Cholesterol reduction mechanisms and fatty acid composition of cellular membranes of probiotic lactobacilli and bifidobacteria. J Funct Foods 9:295–305. https://doi.org/10.1016/j.jff.2014.05.002
Margolles A, Yokota A (2011) Bile stress in lactic acid bacteria and bifidobacteria. In: Sonomoto K, Yokota A (eds) Lactic acid bacteria and bifidobacteria: current progress in advanced research. Caister Academic Press, Norfolk, pp 111–142
Thakkar P, Modi H, Dabhi B, Prajapati J (2014) Bile tolerance, bile deconjugation and cholesterol reducing properties of Lactobacillus strains isolated from traditional fermented foods. Int J Fermented Foods 3(2):157–165. https://doi.org/10.5958/2321-712X.2014.01316.7
Li Q, Ma Y, He S, Elfalleh W, Xu W, Wang J, Qiu L (2014) Effect of pH on heat stability of yak milk protein. Int Dairy J 35:102–105. https://doi.org/10.1016/j.idairyj.2013.10.015
Lucey JA (2016) Acid coagulation of milk. Advanced dairy chemistry. Springer, New York. doi:https://doi.org/10.1007/978-1-4614-4714-6
Todorov SD, Stojanovski S, Iliev I, Moncheva P, Nero LA, Ivanova IV (2017) Technology and safety assessment for lactic acid bacteria isolated from traditional Bulgarian fermented meat product “lukanka”. Braz J Microbiol 48:576–586. https://doi.org/10.1016/j.bjm.2017.02.005
Sha Y, Wang L, Liu M, Jiang K, Xin F, Wang B (2016) Effects of lactic acid bacteria and the corresponding supernatant on the survival, growth performance, immune response and disease resistance of Litopenaeus vannamei. Aquaculture 452:28–36. https://doi.org/10.1016/j.aquaculture.2015.10.014
FAO/WHO (2002) Report of a joint FAO/WHO working group on drafting guidelines for the evaluation of probiotics in food. Food and agriculture organization of the United Nations. https://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf. Accessed 12 July 2020
Panghal A, Janghu S, Virkara K, Gat Y, Kumar V, Chhikara N (2018) Potential non-dairy probiotic products – a healthy approach. Food Biosci 21:80–89. https://doi.org/10.1016/j.fbio.2017.12.003
Anadón A, Martínez-Larrañaga MR, Ares I, Martínez MA (2016) Probiotics: safety and toxicity considerations. In: Gupta RC (ed) Nutraceuticals efficacy, safety and toxicity. Academic Press, London, pp 777–798
Bakde C, Poddar A (2011) Effect of steel plant effluent on acid and alkaline phosphatases of gills, liver and gonads of Cyprinus carpio Linn. Int J Environ Sci 1(6):1305–1316. https://doi.org/10.6088/ijes.00106020025
Millán JL (2006) Mammalian alkaline phosphatases: from biology to applications in medicine and biotechnology. WILEY-VCH Verlag GmbH & Co, Weinheim doi:https://doi.org/10.1002/3527608060
Bilski JM-B, Wojcik DA, Zahradnik-Bilska J, Brzozowski B, Magierowski M, Mach T, Magierowska K, Brzozowski T (2017) The role of intestinal alkaline phosphatase in inflammatory disorders of gastrointestinal tract. Mediat Inflamm 2017:1–9. https://doi.org/10.1155/2017/9074601
Papamanoli E, Tzanetakis N, Litopoulou-Tzanetaki E, Kotzekidou P (2003) Characterisation of lactic acid bacteria isolated from a Greek dry-fermented sausage in respect of their technological and probiotic properties. Meat Sci 65:859–867. https://doi.org/10.1016/S0309-1740(02)00292-9
Mesas JM, Rodríguez MC, Alegre MT (2011) Characterization of lactic acid bacteria from musts and wines of three consecutive vintages of Ribeira sacra. Lett Appl Microbiol 52:258–268. https://doi.org/10.1111/j.1472-765X.2010.02991.x
Monteagudo-Mera A, Caro I, Rodriguez-Aparicio LB, Rua J, Ferrero MA, Garcia-Armesto MR (2011) Characterization of certain bacterial strains for potential use as starter or probiotic cultures in dairy products. J Food Prot 74:1379–1386. https://doi.org/10.4315/0362-028X.JFP-10-392
Leite AMO, Miguel MAL, Peixoto RS, Ruas-Madiedo P, Paschoalin VMF, Mayo B, Delgado S (2015) Probiotic potential of selected lactic acid bacteria strains isolated from Brazilian kefir grains. J Dairy Sci 98(6):3622–3632. https://doi.org/10.3168/jds.2014-9265
Aziz G, Fakhar H, ur Rahman S, Tariq M, Zaidi A (2019) An assessment of the aggregation and probiotic characteristics of Lactobacillus species isolated from native (desi) chicken gut. J Appl Poul Res 28(4):846–857. https://doi.org/10.3382/japr/pfz042
Stergiou VA, Thomas LV, Adams MR (2006) Interactions of nisin with glutathione in a model protein system and meat. J Food Prot 69:951–956. https://doi.org/10.4315/0362-028X-69.4.951
Settanni L, Corsetti A (2008) Application of bacteriocins in vegetable food biopreservation. Int J Food Microbiol 121:123–138. https://doi.org/10.1016/j.ijfoodmicro.2007.09.001
Deegan LH, Cotter PD, Hill C, Ross P (2006) Bacteriocins: biological tools for bio-preservation and shelf-life extension. Int Dairy J 16:1058–1071. https://doi.org/10.1016/j.idairyj.2005.10.026
Vieco-Saiz N, Belguesmia Y, Raspoet R, Auclair E, Gancel F, Kempf I, Drider D (2019) Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front Microbiol 10:1–17. https://doi.org/10.3389/fmicb.2019.00057
Moreno I, Lerayer ALS, Baldini VLS, Leitão MFF (2000) Characterization of bacteriocins produced by Lactococcus lactis strains. Braz J Microbiol 31:183–191. https://doi.org/10.1590/S1517-83822000000300007
Lim HS, Yeu JE, Hong SP, Kang MS (2018) Characterization of antibacterial cell-free supernatant from oral care probiotic Weissella cibaria, CMU. Molecules 23(8):1–13. https://doi.org/10.3390/molecules23081984
Bendjeddou K, Fons M, Strocker P, Sadoun D (2012) Characterization and purification of a bacteriocin from Lactobacillus paracasei subsp paracasei BMK2005, an intestinal isolate active against multidrug-resistant pathogens. World J Microb Biot 8:1543–1552. https://doi.org/10.1007/s11274-011-0958-1
Lü X, Yi L, Dang J, Dang Y, Liu B (2014) Purification of novel bacteriocin produced by Lactobacillus coryniformis MXJ 32 for inhibiting bacterial foodborne pathogens including antibiotic-resistant microorganisms. Food Control 46:264–271. https://doi.org/10.1016/j.foodcont.2014.05.028
Abbasiliasi S, Nagasundara RR, Azmi TIT, Shuhaimi M, Rosfarizan M, Ariff AB (2010) Partial characterization of antimicrobial compound produced by Lactobacillus paracasei LA 07, a strain isolated from Budu. Minerva Biotecnologica 22
Riley MA, Chavan MA (2007) Bacteriocins: ecology and evolution. Springer-Verlag Berlin Heidelberg, Berlin. doi:https://doi.org/10.1007/978-3-540-36604-1
Korenblum E, von der Weid I, Santos ALS, Rosado AS, Sebastian GV, Coutinho CMLM, Magalhães FCM, de Paiva MM, Seldin L (2005) Production of antimicrobial substances by Bacillus subtilis LFE-1, B. firmus H2O-1 and B. licheniformis T6-5 isolated from an oil reservoir in Brazil. J Appl Microbiol 98:667–675. https://doi.org/10.1111/j.1365-2672.2004.02518.x
Ansari A, Zohra RR, Tarar OM, Ul Qader SA, Aman A (2018) Screening, purification and characterization of thermostable, protease resistant bacteriocin active against methicillin resistant Staphylococcus aureus (MRSA). BMC Microbiol 18:1–10. https://doi.org/10.1186/s12866-018-1337-y
Mariam SH, Zegeye N, Tariku T, Andargie E, Endalafer N, Aseffa A (2014) Potential of cell-free supernatants from cultures of selected lactic acid bacteria and yeast obtained from local fermented foods as inhibitors of Listeria monocytogenes, Salmonella spp. and Staphylococcus aureus. BMC Res Notes 7:1–9. https://doi.org/10.1186/1756-0500-7-606
Ponce AG, Moreira MR, del Valle CE, Roura SI (2008) Preliminary characterization of bacteriocin-like substances from lactic acid bacteria isolated from organic leafy vegetables. LWT - Food Sci Technol 41(3):432–441. https://doi.org/10.1016/j.lwt.2007.03.021
Pawar DD, Malik SVS, Bhilegaonkar KN, Barbuddhe SB (2000) Effect of nisin and its combination with sodium chloride on the survival of Listeria monocytogenes added to raw buffalo meat mince. Meat Sci 56:215–219. https://doi.org/10.1016/s0309-1740(00)00043-7
Prudêncio CV, dos Santos MT, Vanetti MCD (2015) Strategies for the use of bacteriocins in gram-negative bacteria: relevance in food microbiology. J Food Sci Technol 52(9):5408–5417. https://doi.org/10.1007/s13197-014-1666-2
Martinez RCR, de Martinis ECP (2006) Effect of Leuconostoc mesenteroides 11 bacteriocin in the multiplication control of Listeria monocytogenes 4b. Cienc Tecnol Aliment 26:52–55. https://doi.org/10.1590/S0101-20612006000100009
Daw MA, Falkiner FR (1996) Bacteriocins: nature, function and structure. Micron 27:467–479. https://doi.org/10.1016/S0968-4328(96)00028-5
Boziaris IS, Adams MR (2001) Temperature shock, injury and transient sensitivity to nisin in Gram negatives. J Appl Microbiol 91:715–724. https://doi.org/10.1046/j.1365-2672.2001.01433.x
Barman S, Ghosh R, Mandal DC (2018) Production optimization of broad spectrum bacteriocin of three strains of Lactococcus lactis isolated from homemade buttermilk. Annals of Agrarian Science 16:286–296. https://doi.org/10.1016/j.aasci.2018.05.004
Manning MC, Chou DK, Murphy BM, Payne RW, Katayama DS (2010) Stability of protein pharmaceuticals: an update. Pharm Res 27:544–575. https://doi.org/10.1007/s11095-009-0045-6
Alakomi H-L, Skyttä E, Saarela M, Mattila-Sandholm T, Latva-Kala K, Helander IM (2000) Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl Environ Microbiol 66:2001–2005. https://doi.org/10.1128/aem.66.5.2001-2005.2000
Delaunay JL, Breton M, Trugnan G, Maurice M (2008) Differential solubilization of inner plasma membrane leaflet components by Lubrol WX and Triton X-100. Biochim Biophys Acta Biomembr 1778(1):105–112. https://doi.org/10.1016/j.bbamem.2007.09.017
Funding
This research was funded by the Fundamental Research Grant Scheme (Grant No.: FRGS/1/2015/SG05/UPM/01/3) (Vot. No. 5524710) from the Ministry of Education Malaysia.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
ESM 1
(DOCX 4102 kb)
Rights and permissions
About this article

Cite this article
Jawan, R., Abbasiliasi, S., Mustafa, S. et al. In Vitro Evaluation of Potential Probiotic Strain Lactococcus lactis Gh1 and Its Bacteriocin-Like Inhibitory Substances for Potential Use in the Food Industry. Probiotics & Antimicro. Prot. 13, 422–440 (2021). https://doi.org/10.1007/s12602-020-09690-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12602-020-09690-3